
ABSTRACT
Determining genotypic responses to soil fertility may assist selection of cultivars that can be adapted to varied soil fertility regimes, and such selection under field conditions is still limited. A two-year field experiment was conducted in long-term field trials to investigate wheat genotype effects on early growth, yield and nutrient accumulation as affected by varied long-term soil fertility managements and nitrogen (N) fertilization. Results show that the early growth, grain yield and nutrient accumulation of spring wheat plants were strongly affected by soil fertility managements and genotypes. Early shoot and root biomass of singly grown plants and leaf canopy growth under standard growth density was associated with subsequent grain yield of plants under standard growth density across the gradient in soil fertility levels. Taifun and Thasos had stable higher yield and N and phosphorus (P) uptake across varied soil fertility regimes compared with other genotypes. Økilde, however, increased yield by 8–34% and N and P accumulation by 1–22% only when grown in the high organic manure treatment compared with other genotypes, indicating that it is more adapted to high organic fertility regimes. Therefore, the different responses and adaptations of genotypes to soil fertility regimes should be included during selection of cultivars.
Introduction
The global population is continually increasing, and to meet the food demand of this increasingly affluent population, the global crop production needs to double by 2050 (Ray et al. Citation2013). Global use of nitrogen (N) and phosphorus (P) fertilizer increased sevenfold and 3.5-fold between 1960 and 1995 to achieve high yield of crops; however, further increases in N and P application are unlikely to be as effective at increasing yields because of diminishing returns (Tilman et al. Citation2002). Moreover, the high level of fertilizer application in crop production leads to many unwanted environmental consequences. Therefore, it is crucial to optimize crop production practices and management to achieve higher crop yields with reduced resource use and lower environmental impacts.
In low-input and -organic agriculture, external inputs in the form of synthetic fertilizers are reduced or abolished. However, the currently available crop varieties are generally from conventional breeding programs with high input of synthetic fertilizers and pesticides, which have not been selected for traits favorable under less-intensive inputs. In organic management systems, grain yields are generally low compared to conventional production systems (Poutala et al. Citation1993; Mason et al. Citation2007; Reid et al. Citation2009). Varieties selected by plant breeders under conventional systems are widely used in organic farming, where they are grown under conditions they were not selected for. Moreover, many selections were performed in controlled environments rather than in field conditions. Genotypes may show different responses and adaptations to soil fertility managements and growth environment. Stagnari et al. (Citation2013) reported that the yield of wheat genotypes was affected significantly by N fertilization levels in organic farming. However, some of the genotypes showed good and stable yield levels in organic systems, whether grown under N-fertilization or unfertilized conditions, indicating higher ability to utilize inherent soil fertility conditions. Similarly, Ryan et al. (Citation2008) found significant yield responses of barley landraces to soil fertility levels in a greenhouse experiment, and some genotypes increased yield significantly when fertilized with N and P, whereas others remained at the same level or decreased yield. Therefore, it is important to assess the extent to which such genotypes respond to fertilizer inputs as this may affect the optimal breeding strategy, particularly for selection of germplasm adapted to a particular growing environment, for example, low-input and -organic agriculture (Ryan et al. Citation2008). Early shoot and root vigor is critical for resource acquisition, and plants with good early vigor improve light interception, biomass production, grain yield and nitrogen uptake efficiency (Rebetzke et al. Citation2004; Liao et al. Citation2008; Mullan & Reynolds Citation2010). When plants are grown singly, the effect of differences in early establishment would be easier to show in single plants than a crop, as the effect of early vigor in a crop is counteracted by subsequently increased competition within the crop, reducing its final effect and making it much more difficult to observe the impact among other differences. Moreover, singly grown plants, on the other hand, can benefit to a much larger extent from the advantages of early vigor throughout their growing period. Therefore, the early vigor growth of spring wheat genotypes grown both singly and under conventional standard density was examined. The objective of the present study was to investigate wheat genotype effect on early growth, yield and nutrient accumulation as affected by long-term soil fertility levels and fertilizer applications in the field. We also hypothesized that early above- and below-ground growth of singly grown plants and early leaf canopy growth under standard growth density are correlated to crop development and yield under standard growth density.
Materials and methods
Experimental site, treatment and growing conditions
Field experiments were conducted in two growing seasons, from April to August in 2013 and 2014, in two different long-term field trials at the experimental farm of University of Copenhagen, both located 20 km west of Copenhagen, Taastrup, Denmark, that is, the Nutrient Depletion Trial in 2013 and the CRUCIAL Trial in 2014.
The Nutrient Depletion Trial site used in 2013 did not receive P- or K-containing fertilizers from 1964 to 1995; only moderate N fertilizer rates were applied for the mainly cereal crops grown until 1995. Since 1996 different treatments with various application rates of mineral N, P, K and/or organic fertilizer have been applied. The soil type is sandy loam (clay 15%, silt 18%, sand 65%), and the soil contains total carbon 11.5 g kg−1, total N 1.3 g kg−1 and has a pH of 5.6 in the topsoil layer (0–30 cm). The Nutrient Depletion Trial comprises 14 long-term soil fertility management treatments, with four replicates in a block design; three of these treatments were used in this study, with five different spring wheat genotypes. Soil fertility management treatments comprise the ‘NoPK’ mineral fertilizer (60 N, 0 P, 0 K kg ha−1 yr−1), the ‘NPK’ mineral fertilizer (120 N, 20 P, 120 K kg ha−1 yr−1) and the ‘low organic manure’ fertilization (85 Ntot 17 P, 89 K kg ha−1 yr−1 in animal slurry). The P and K available for plants were 8, 12 and 10 mg Olsen-P kg−1 and 73, 113 and 94 mg exchangeable K kg−1 in the NoPK, NPK and low organic manure treatment, respectively.
The spring wheat genotypes included Taifun (origin: Lochow-Petkus GmbH, Germany; released 2002), Thasos (origin: Strube Research GmbH & Co. KG, Germany; released 1994), Økilde (origin: Nordic Seed A/S, Denmark; released 2012), Dacke (origin: Svalöf Weibull AB, Sweden; released 1990) and April Bearded (origin: landrace selected by Sir John Sinclair from the West Country, UK; released 1838). Four of the genotypes (April Bearded was omitted due to limited seed availability for full field plot trials) were grown in field plots with conventional standard density of 450 plant m−2. In addition, all five genotypes were grown as single plants in individual plots in each replicate in order to investigate the variations of early vigor development among genotypes without plants’ competition under standard growth density. Field plot size for each genotype at standard density was 3 m by 10 m. For single plants, seeds were sown by hand with a space of 30 cm both within and between rows and then seedlings were thinned after germination.
The CRUCIAL trial used in 2014 includes 11 long-term (since 2002) soil fertility management treatments, of which two were used in this study, namely the ‘unfertilized’ and the ‘high organic manure’ treatments. The unfertilized treatment received no external fertilizers, but a clover grass mixture was grown each autumn and incorporated as a green manure. The high organic manure treatment had received cattle manure at an accelerated application rate of 125 t ha−1 yr−1. The experiment was a randomized block design with three replicates. The soil type was sandy loam (16% clay, 17% silt and 66% sand), and the soil of the unfertilized treatment had a pH of 7.0, total C of 13.6 g kg−1, total N of 1.5 g kg−1, Olsen-P of 13 mg kg−1 and exchangeable K of 95 mg kg−1. The high organic manure treatment soil had a pH of 7.0, total C of 20 g kg−1, total N of 2.0 g kg−1, Olsen-P of 45 mg kg−1 and exchangeable K of 254 mg kg−1. In 2014, the ordinary practice of applying manure to the high manure treatment was omitted, and only N was given as fertilizer in different forms as described below. Four spring wheat genotypes (the same genotypes as in 2013 except April bearded) were grown at standard plant density (450 m−2) in each of the long-term soil fertility treatments. The N fertilization treatments included a mineral N and an organic N fertilizer. The mineral N was applied at the rate of 108 kg N ha−1, in the form of an NS fertilizer containing total N of 27% (as ammonium-nitrate) and total S of 4.5% . The organic N fertilizer was hoof and horn meal containing total N of 15.3% (Elixir Garden Supplies Ltd., Morecambe, UK) and was applied at the rate of 270 kg total N ha−1 (under the assumption of 40% of the N becoming available to the crop during the growing season, making it equivalent to 108 kg N mineral N ha−1). The hoof and horn meal was placed at 3 cm depth below soil surface using a sowing machine prior to the sowing of the seeds.
Daily weather data were recorded from a meteorological station located within 3 km from the experimental fields. The weather data during spring wheat growing season are listed in .
Table 1. Weather data during the two spring wheat growing seasons in 2013 and 2014 recorded at the climate station of the experimental farm of University of Copenhagen within 3 km of the two field trial sites.
Sampling, measurements and analyses
Singly grown plants were sampled at the early growth stages of GS11 (first leaf unfold), GS21 (main shoot and one tiller) and GS32 (second node detectable) (BBCH scale) in 2013. On each sampling, 25 single plants for each variety in each treatment were harvested to determine leaf area, dry biomass of shoot and root and root length. The leaf area of single plants was measured with a leaf area meter (model 3050A, Li-Cor Inc. Lincoln, NB, USA). The roots of sampled single plants were excavated and harvested together with soil adhering to the roots from the field. After transfer to the lab, roots were rinsed carefully by washing in tap water, and then scanned using a root scanner (Epson Perfection V700, CA, USA) and analyzed by the WinRHIZO image analysis system (Regents Instruments Inc., Quebec city, Canada) for root length. In 2014 the normalized difference vegetation index (NDVI) of plants grown in the plots of standard density was measured regularly each week as an index of leaf canopy area by Spectrosense 2+ (Skye Instruments, Powys, UK).
Plants in the field plots of standard density in 2013 and 2014 were harvested at maturity by an experimental combined harvester, and the weight of straw and grains were determined. For dry biomass determination, samples from all plant samples were oven dried at 70°C to constant weight. After grinding of dried plant samples, plant N concentration was analyzed using the Dumas dry combustion method in a system consisting of an ANCA-SL Elemental Analyzer coupled to a 20–20 Mass Spectrometer (Sercon Instruments, Crewe, UK). Plant P concentration was determined by the dry-ashing method at 500°C for 5 h (Jones Citation2001), followed by extraction in acid. Plant extracts were kept at 4°C, and the concentration of P in the extracts was determined using a flow-injection analyzer (FIAstar 5000, FOSS, Denmark). The accumulation of N and P was calculated by multiplying the element concentration by the dry weight.
Statistical analysis
Data were analyzed by analysis of variance (ANOVA) using SPSS software at a significance level of 5%. ANOVAs were done with genotype, soil fertility management or N fertilization as the main effects and including two-and three-way interactions. Duncan’s multiple range test was applied to assess the differences between treatments at a significance level of 5%. Pearson correlation coefficients were used to determine the relationships between the measured parameters.
Results
Early vigor growth
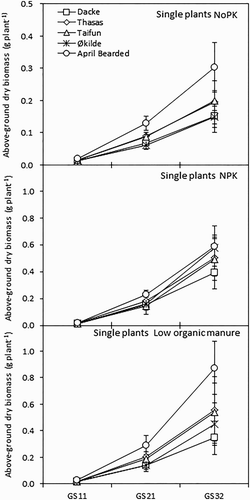
Above-ground dry biomass of single plants at early growth stages in the 2013 experiment was significantly affected by both genotypes and soil fertility management regimes (). April Bearded had the highest above-ground dry biomass, which was 32–98% higher than the other genotypes. Across the genotypes, the above-ground dry biomass under NoPK treatment was 12–64% lower than under the NPK and the low organic manure treatments. Similarly, the root dry biomass and length of single plants were significantly affected by both genotypes and soil fertility management regimes at most of the early growth stages. April bearded had the significantly highest root dry biomass and length compared with other genotypes. NPK treatment increased the growth of root biomass and length compared to NoPK and low organic manure treatments. During the 2014 experiment, leaf canopy area as measured by NDVI under standard crop density was affected significantly by genotypes only within the first 14 days after germination (DAG), whereafter it became similar among genotypes. The highest NDVI was found for Taifun, intermediate for Thasos and Dacke, and the lowest for Økilde. Soil fertility managements affected NDVI, with 3–51% higher values under organic manure than under unfertilized treatment.
Figure 1. Above-ground dry biomass of spring wheat genotypes in the plots of single plants during the early growth stages in 2013. Values are means ± standard error (SE).
Grain yield
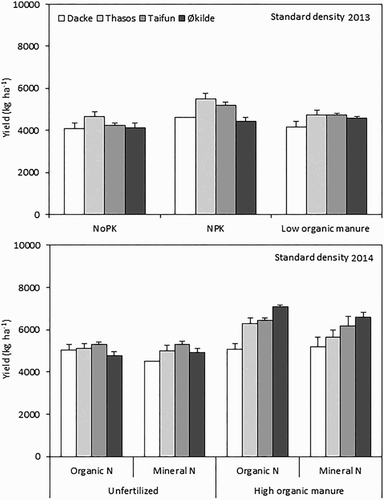
Grain yield was significantly affected by genotypes and long-term soil fertility management treatments in both years, whereas it was not affected significantly by actual N fertilization form (mineral vs. organic) in 2014 (). High soil fertility management treatments generally resulted in higher grain yields, that is, the NPK treatment gave 15% and 8% significantly higher yield compared with the NoPK and low organic manure treatment in 2013 and the high organic manure treatment gave 21% substantially higher yield than the unfertilized management treatment in 2014 across the genotypes. Analysis across the soil fertility treatments showed that Thasos and Taifun exhibited 7–18% higher grain yield compared with Dacke and Økilde, except under high organic manure treatment in 2014, where Økilde had 8–34% higher grain yield compared to other genotypes.
Figure 2. Grain yield of spring wheat genotypes grown under standard density in 2013 and 2014. Values are means ± SE.
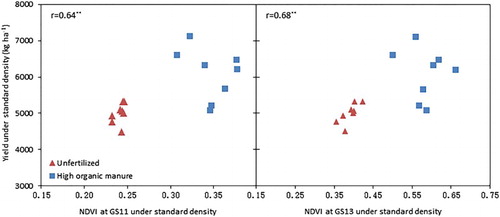
There were significant correlations between above-ground dry biomass or root dry biomass of single plants at the early growth stages and grain yield of the crop under standard growth density across soil fertility treatments in 2013 (), except root dry biomass at the very early GS11 (); however, most of the correlation is caused by the different soil fertility levels, and less so by the genotype differences. Similarly, the leaf canopy growth from the very early growth stages (GS11 and GS13) showed significant correlation to the final grain yield of the crop (). Likewise, most of the correlation is due to the soil fertility management history, but it appears that in the unfertilized treatment, there is a closer relation between early stage NDVI and final yield.
Figure 3. Relationship between above-ground (left) and root dry biomass (right) of single plants at early growth stages and the yield of spring wheat plants grown under standard density in the different soil fertility treatments in 2013. r denotes Pearson correlation coefficients, and * significance at P < .05.
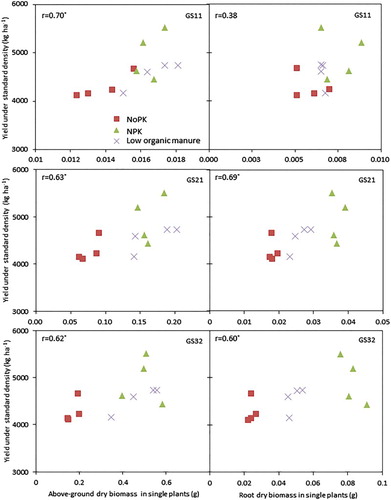
Figure 4. Relationship between NDVI at early growth stages and the harvested yield of spring wheat grown in 2014 in the different soil fertility management treatments. r denotes Pearson correlation coefficients, and ** significance at P < .01.
Grain N and P accumulation
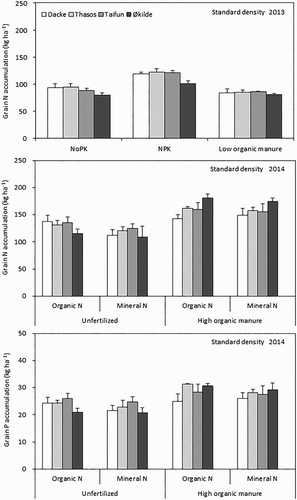
Grain N accumulation was significantly affected by both genotypes and soil fertility management treatments in 2013, whereas it was only affected by soil fertility management treatments in 2014 (). High soil fertility management increased grain N accumulation by 31% and 38% compared with the NoPK and low organic manure treatments across the genotypes. Dacke, Thasos and Taifun had 13–16% higher grain N accumulation than Økilde in 2013 across the soil fertility treatments. A similar tendency of grain N and P accumulation was found among genotypes within the unfertilized treatments in 2014; however, under the high organic manure management treatment, Økilde increased grain N and P accumulation by 1–22% compared with other genotypes.
Figure 5. Grain N and P accumulation of spring wheat genotypes grown under standard density in 2013 (N) and 2014 (N and P). Values are means ± SE.
Discussion
In the present study, early vigor growth as indicated by both the shoot and root growth of single plants in 2013 and NDVI measurements in 2014, harvest yield and nutrient accumulation were significantly affected by both genotypes and long-term soil fertility regimes. Consistently, Liao et al. (Citation2004) observed significant differences in vigorous early root and shoot growth of wheat plants. Likewise, in a sandy soil, Liao et al. (Citation2006) found that vigorous wheat genotypes had 33–83% higher total root length as well as earlier and more extensive root growth in the upper soil profile, contributing to the higher uptake of the N fertilizer. Comparing genotypes grown both singly and under standard density has the advantage that the genetic variations of early growth vigor can be examined both with and without competition between plants. There were significant genotypic differences in shoot and root growth at early growth stages for singly grown plants. Similarly, under standard growth density, the leaf canopy differed significantly before GS21. The early leaf canopy of Taifun, Thasos and Dacke was significantly higher than that of Økilde, and the same tendency was also found for final grain yield and N and P accumulation in both years except Økilde under the high organic manure treatment in 2014 (), indicating the importance of early vigor establishment in determining subsequent grain yield and nutrient acquisition. Moreover, the grain yields of genotypes grown under standard density were significantly correlated with shoot and root dry biomass of single plants in the 2013 trial () and with early growth NDVI of plants at standard density in the 2014 trial (). However, it is noteworthy that the main cause for these positive correlations was the soil fertility gradient. Across the cultivars within each fertility level, there was a tendency that under the low organic manure and NoPK management histories in 2013 and the unfertilized history in 2014, a positive correlation existed, whereas under the NPK (2013) or high organic manure management (2014) histories, there was no correlation whatsoever; however, conclusions on this are limited by the few numbers of genotypes in this study.
It has been proposed that organic farming and low-input agriculture rely on genotypes with high adaptability, particularly under low N and P fertilization levels (Rao et al. Citation1993; Lammerts van Bueren et al. Citation2011). Nass et al. (Citation2003) found significantly higher grain yield by the genotype AC Barrie under organic conditions compared to the genotype AC Walton, which generally outcompeted AC Barrie for grain yields under conventional conditions. Better performance of certain genotypes under organic field conditions has also been reported previously (Mason et al. Citation2007). Similarly, large genotypic variations in grain yield and responses to P fertilizer were found among wheat genotypes (McDonald et al. Citation2015). In the present study, the grain yield and nutrient accumulation in Thasos, Taifun and Dacke were similarly ranked under different long-term soil fertility regimes, with Taifun and Thasos having stable higher yield and nutrient uptake compared with other genotypes across low and high soil fertility conditions in both years. Most interestingly, Økilde, on the high organic manure management history in 2014, improved yield and nutrient accumulation significantly ( and ), indicating the varied adaptation of genotypes to soil fertility conditions. Økilde, which achieved high yield and nutrient accumulation under high organic manuring conditions, therefore appears to be more adaptive and suitable for high organic fertility management. When Økilde was grown singly, the above-ground dry biomass was lower compared with most of the other genotypes at early growth stages (). Moreover, similar to the single plants, under standard growth density, Økilde had the significantly lowest leaf canopy compared to other genotypes. Thus, the weak early vigor growth could lead to subsequent lower grain yield and nutrient uptake.
Conclusively, the early growth, yield and nutrient accumulation were significantly affected by genotypes and long-term soil fertility managements in the field. Early shoot and root dry biomass of singly grown plants and early leaf canopy growth under standard density were associated with subsequent grain yield under normal density across soil fertility gradients. Taifun and Thasos showed a higher yield and nutrient uptake across most soil fertility regimes. However, Økilde achieved a high yield and nutrient uptake only under the high organic fertility regime, and is apparently responsive to nutrient availability conditions than the others. Therefore, the varied responses and adaptation of genotypes to soil fertility regimes should be included during selection of cultivars.
Disclosure statement
No potential conflict of interest was reported by the authors.
Notes on contributors
Yaosheng Wang obtained Ph.D. and also worked as Postdoctoral fellow in the University of Copenhagen. Currently, he is a professor in the Institute of Environment and Sustainable Development in Agriculture, Chinese Academy of Agricultural Sciences. His researches are focusing on plant eco-physiology, biotic and abiotic stress, plant nutrition especially nitrogen and phosphorus, root and root hairs, soil biochemistry and agricultural water management.
Jakob Magid is an associate professor in the Department of Plant and Environmental Sciences, University of Copenhagen. His researches mainly focus on nutrient and waste management in land-based production systems, with an emphasis on carbon, nitrogen and phosphorus. In recent years emphasis has been placed on detailed studies of decomposition processes, and on a larger scale, the possibilities and barriers for increased nutrient and organic matter recycling from urban to peri-urban areas.
Kristian Thorup-Kristensen is a professor in the Department of Plant and Environmental Sciences, University of Copenhagen. The main aims of his researches are to study root growth and resource acquisition of crops, and to use this in the development of resource-efficient cropping systems. He is currently focusing on deep rooting and crop utilization of resources from deep soil layers as well as developing new methods for deep root observations.
Lars Stoumann Jensen is a professor in the Department of Plant and Environmental Sciences, University of Copenhagen. His major research focus has been on biological soil fertility, and the influence of organic matter decomposition processes on nutrient turnover in agro-ecosystems. Currently, he is also working on utilization of agricultural and urban waste, and their fertilizer value and effects on soil quality, gas emission and nutrient losses to the environment.
ORCiD
Yaosheng Wang http://orcid.org/0000-0002-2657-7057
Jakob Magid http://orcid.org/0000-0001-5867-0910
Kristian Thorup-Kristensen http://orcid.org/0000-0001-5476-985X
Lars Stoumann Jensen http://orcid.org/0000-0002-1446-2084
Additional information
Funding
References
- Jones JB. 2001. Laboratory guide for conducting soil tests and plant analysis. Boca Raton, FL: CRC Press LLC, 202–229.
- Lammerts van Bueren ET, Jones SS, Tamm L, Murphy KM, Myers JR, Leifert C, Messmer MM. 2011. The need to breed crop varieties suitable for organic farming, using wheat, tomato and broccoli as examples: a review. NJAS-Wagen J Life Sc. 58:193–205. doi: 10.1016/j.njas.2010.04.001
- Liao MT, Fillery IRP, Palta JA. 2004. Early vigorous growth is a major factor influencing nitrogen uptake in wheat. Funct Plant Biol. 31(2):121–129. doi: 10.1071/FP03060
- Liao MT, Hocking PJ, Dong B, Delhaize E, Richardson AE, Ryan PR. 2008. Variation in early phosphorus-uptake efficiency among wheat genotypes grown on two contrasting Australian soils. Aust J Agric Res. 59:157–166. doi: 10.1071/AR06311
- Liao MT, Palta JA, Fillery IRP. 2006. Root characteristics of vigorous wheat improve early nitrogen uptake. Aust J Agric Res. 57:1097–1107. doi: 10.1071/AR05439
- Mason HE, Navabi A, Frick BL, O’Donovan JT, Spaner DM. 2007. The weed competitive ability of Canada western red spring wheat cultivars grown under organic management. Crop Sci. 47:1167–1176. doi: 10.2135/cropsci2006.09.0566
- McDonald G, Bovill W, Taylor J, Wheeler R. 2015. Responses to phosphorus among wheat genotypes. Crop Pasture Sci. 66(5):430–444. doi: 10.1071/CP14191
- Mullan DJ, Reynolds MP. 2010. Quantifying genetic effects of ground cover on soil water evaporation using digital imaging. Funct Plant Biol. 37:703–712. doi: 10.1071/FP09277
- Nass HG, Ivany JA, MacLeod JA. 2003. Agronomic performance and quality of spring wheat and soybean cultivars under organic culture. Am J Alternative Agric. 18:164–170. doi: 10.1079/AJAA200348
- Poutala RT, Korva J, Varis E. 1993. Spring wheat cultivar performance in ecological and conventional cropping systems. J Sustain Agric. 3:63–84. doi: 10.1300/J064v03n03_05
- Rao ACS, Smith JL, Jandhyala VK, Papendick RI, Parr JF. 1993. Cultivar and climatic effects on the protein content of soft white winter wheat. Agron J. 85:1023–1028. doi: 10.2134/agronj1993.00021962008500050013x
- Ray DK, Mueller ND, West PC, Foley JA. 2013. Yield trends are insufficient to double global crop production by 2050. PLOS one. 8(6):e66428. doi: 10.1371/journal.pone.0066428
- Rebetzke GJ, Botwright TL, Moore CS, Richards RA, Condon AG. 2004. Genotypic variation in specific leaf area for genetic improvement of early vigour in wheat. Field Crops Res. 88:179–189. doi: 10.1016/j.fcr.2004.01.007
- Reid TA, Salmon DF, Yang R, Salmon DF, Spaner SD. 2009. Should spring wheat breeding for organically managed systems be conducted on organically managed land?. Euphytica. 169:239–252. doi: 10.1007/s10681-009-9949-9
- Ryan J, Masri S, Ceccarelli S, Grando S, Ibrikci H. 2008. Differential responses of barley landraces and improved barley cultivars to nitrogen-phosphorus fertilizer. J Plant Nutr. 31:381–393. doi: 10.1080/01904160801894939
- Stagnari F, Onofri A, Codianni P, Pisante M. 2013. Durum wheat varieties in N-deficient environments and organic farming: a comparison of yield, quality and stability performances. Plant Breeding. 132:266–275.
- Tilman D, Cassman KG, Matson PA, Naylor R, Polasky S. 2002. Agricultural sustainability and intensive production practices. Nature. 418:671–677. doi: 10.1038/nature01014