
ABSTRACT
Purpose: Nitrogen (N) / potassium (K) nutrient balance has been studied for some ornamental plants, however, available information is limited. Here we investigate the optimum N and K balance and concentration for lisianthus production in soilless medium.
Materials and methods: The effect of three N / K balances: 1.43, 2.14 and 4.29, prepared by varying the concentration of N and K, were evaluated in lisianthus grown in soilless medium (volcanic rock).
Results: Plants fertigated with a N / K balance of 2.14 exhibited enhanced height and stem dry weight when compared to plants fertigated with a balance of 4.29, and a higher flower buds count and total dry weight than those fertigated with a balance of 1.43 or 4.29. Plants fertigated with a balance of 2.14 exhibited increased dry weight when N was reduced from 15 to 9 meq L−1 and K from 7 to 4.2 meq L−1, suggesting that lisianthus does not require high levels of these nutrients. Although shoot N concentration was not correlated with N concentration in the nutrient solution or balance, increasing N in the shoot was associated with higher P and Mg in the plant tissues. In general, P, Ca, and Mg in plant tissues were unaffected by the external N / K balance, however, the internal P and Mg concentrations were positively correlated with shoot dry weight. Shoot K concentration significantly increased when the N / K balance decreased, which is related to the higher concentrations of external K when the balance decreased.
Conclusions: The optimum N / K balance for lisianthus was 2.14. However, there was a concentration effect, as fertigation with solutions containing a N / K balance of 2.14 and an N and K concentration of 9 and 4.2 meq L−1 respectively, resulted in plants with the greatest dry weight.
Introduction
Lisianthus (Eustoma grandiflorum [Raf.] Shinn) is a relatively new ornamental crop that has gained popularity worldwide (Halevy and Kofranek Citation1984; Marchese et al. Citation2005; Harbaugh Citation2007). Native to the Southern United States and Northern México (Halevy and Kofranek Citation1984), lisianthus F1 hybrids have been introduced into several countries, including México. Flower growers in México started to cultivate lisianthus in the late 1990’s and its production has rapidly expanded as an attractive and profitable alternative to chrysanthemum (Chrysanthemum morifolium Ramat) and other cut flower ornamental species.
Compared to chrysanthemum, lisianthus demands low fertilisation rates (Sakata Citation2017) and fertilisation with calcium nitrate is recommended for sturdy stems (Sakata Citation2017), which are desirable during production and post-harvest. Electrical conductivity (EC) of fertigation solutions for lisianthus should be maintained at 1.5 dS m−1, as higher EC may delay flowering and promote rosseting in susceptible cultivars (PanAmerican Seed Citation2017). However, other reports indicate that lisianthus is tolerant to high EC (4–8 dS m−1), provided it is not caused by excess sodium or chloride (Valdez-Aguilar et al. Citation2013), and to high alkalinity in irrigation water provided calcium (Ca) is supplied at higher concentrations (Valdez-Aguilar et al. Citation2013; Gómez-Pérez et al. Citation2014).
Although high levels of nitrogen (N) (250–300 g m−3) were associated with high chlorophyll concentration and photosynthetic rate (Marchese et al. Citation2005), lisianthus seems to prefer low levels of this nutrient (45 mg N per pot per week) as well as low levels of Ca for maximum growth (75 mg L−1 Ca) (Frett et al. Citation1988). In contrast to recommendations, utilising nitrate (NO3−) as the preferred source of N (Armitage and Laushman Citation2003), other studies have demonstrated that increased ammonium (NH4+)-N nutrition enhanced plant growth when compared to NO3−-N (Mendoza-Villarreal et al. Citation2015), suggesting that lisianthus is a calcifuge species due to its low accumulation of Ca (Hernández-Pérez et al. Citation2016).
Nutrient efficiency in plants is higher when nutrients are provided in a balanced fashion, in that, nutrient uptake rates are determined by concentration in the root zone/rhizosphere and by the mutual relationship between nutrients, which is as crucial as the absolute concentration of the ions (Huber and Arny Citation1985; Sonneveld and Voogt Citation2009). Nitrogen and potassium (K) balance has been reported to affect yield, quality, cold hardiness, and postharvest physiology of ornamental plants (Joiner et al. Citation1983). The N / K balance is also of interest because K is the counterion for NO3− when transported from roots to shoots (White Citation2012). In addition, K is associated with the transport of photosynthates from shoot to root, which are used for the active uptake of NO3− through the carriers at the root plasma membrane (Fageria Citation2001). In ornamental species, a low N / K balance usually results in enhanced growth and improved quality; for example, a high N / K balance (2.00) resulted in reduced visual quality of areca palm (Dypsis lutescens) and canna lilies (Canna × generalis), whereas a low N / K balance (0.67) resulted in healthier plants (Broschat et al. Citation2008).
Although studies specifically designed to determine the optimum N / K balance in lisianthus have not been performed, some reports indicate that lisianthus requires higher rates of K (3.26 g m−2) than that of N (2.41 g m−2) (Castillo-González et al. Citation2017). This closely concurs with the optimum N / K balance suggested by Harbaugh (Citation2007) (0.67 on a meq L−1 basis) and Fiorenza (Citation2017) (0.70 on a meq L−1 basis), which resulted in high quality and long vase life flowers (Harbaugh Citation2007). However, the response of lisianthus to the combined effects of N and K concentration and balance has not been elucidated. The objective of the present study was to determine the effect of the N / K balance in the nutrient solution at varying N and K concentrations on plant growth and nutrient status of lisianthus plants cultivated in a soilless medium.
Materials and methods
The experiment was conducted in a glasshouse located at the Universidad Autónoma Chapingo, in central México (19° 20′ N Lat., 98° 53′ W Long., 2240 m above sea level). Average maximum / minimum temperatures were 29.8°C / 9.4°C, and maximum and minimum relative humidity for experiment duration averaged 70% and 40%, respectively. Lisianthus cv ABC 2 Blue plants were transplanted (6 September 2013) into 8 L black containers (four plants per container); each container was considered an experimental unit and contained volcanic rock as growing medium. Particle size of the growing medium was 2–3 mm in diameter.
Experimental units were fertigated with nutrient solutions with two concentrations of N, 9 or 12 meq L−1 (supplied as NO3−-N), while K concentration was adjusted to render a N / K balance, in meq L−1 terms, of 1.43, 2.14 and 4.29 (). The concentration of the reminder nutrients was according to Hoagland and Arnon’s formulation (meq L−1): 9 Ca, 4 Mg, 1 H2PO4−, and 2.9–9.3 SO42−. As a control, a group of plants were fed with the nutrient solution from Hoagland and Arnon, which contains a 2.14 N / K balance. Average EC and pH of the nutrient solution were maintained at ∼2.34 dS m−1 and 6.33, respectively. Micronutrients were supplied at the following concentration (mg L−1): 5.3 Fe, 0.4 Zn, 2.6 Mn, 0.5 Cu, 0.2 B, and 0.2 Mo. Irrigation with the nutrient solution was initiated 8 d after transplant and was provided every other day allowing a 30% leaching fraction.
Table 1. Nitrogen (N) and potassium (K) balance and concentration in the nutrient solution.
Plants were harvested on 15 December 2013 and separated into roots, stems, leaves, and flowers, washed twice with distilled water, and placed in an oven at 70°C for 72 h. Growth measurements were recorded in all the plants from each experimental unit, including leaf area (LI-3100, LI-COR, Lincoln, Nebraska) and dry weight of roots and shoots. Dry root and shoot tissues were ground to pass a 40 mesh sieve (Mini Willey Mill, Thomas Scientific. Swedesboro, NJ) and digested separately in 5 mL H2SO4 and 1 mL H2O2. The digested samples were completed to 25 mL and filtered previous determination of phosphorus (P), K, Ca and magnesium (Mg) with Inductively Coupled Plasma Emission Spectrometer (Agilent 725-ES ICP-OES, Mulgrave, Victoria, Australia) (Soltanpour et al. Citation1996). Nitrogen concentration was determined with micro-Kjeldalh procedure (Bremner Citation1996). Seven replicates of each experimental unit (container with four plants) were distributed in a completely randomised factorial block design. The effect of the N / K balance was analyzed with orthogonal contrast while the effect of the N and K concentration was analyzed using ANOVA and mean comparison test according to Duncan's procedure with SAS at p < 0.05 (SAS v. 9.2, SAS Institute).
Results and discussion
Effect of the N / K balance on plant growth
The balance between two nutrients is often considered as important, or even more important, as the concentration of each one (Joiner et al. Citation1983). In the present study, all of the N / K balances will be expressed on a meq L−1 basis. According to Joiner et al. (Citation1983), most ornamental species yield the best quality plants when the N / K balance ranges from 1.9 to 2.3. However, in chrysanthemum, the total plant fresh weight was reported not to be affected by N / K balances from 1.1 to 2.8 even though the concentration of both nutrients ranged from 14.39–18.94 to 6.8–12.92 meq L−1 of N and K, respectively (Barbosa et al. Citation2000). However, when the N / K balance was 4.2 or 5.6, there was a significant reduction in plant growth (Barbosa et al. Citation2000). The effect of the N / K balance is also demonstrated in the present study as it significantly affected the height, flower buds count, stem and total dry weight of lisianthus (). In general, plants fertigated with solutions containing a N / K balance of 2.14 exhibited higher plant height and stem dry weight than those fertigated with solutions with a N / K balance of 4.29, and higher flower buds count and total dry weight than that of those fertigated with a N / K balance of 1.43 or 4.29. Therefore, an increase in the N / K balance from 2.14 to 4.29, i.e. high N plus low K concentration, and a decrease in the N / K balance form 2.14 to 1.43, i.e. low N plus high K concentration, resulted in decreased dry weight of plants.
Table 2. Average plant growth and dry weight (DW) responses of lisianthus fertigated with nutrient solutions of varying nitrogen (N) and potassium (K) balances and orthogonal contrasts.
A similar N / K balance as the one that resulted in optimum growth and quality for lisianthus in the present study was used by Bernstein et al. (Citation2009) for growing Ranunculus asiaticus L. (Persian buttercup) in soilless culture (N / K balance = 2.3), however, it was demonstrated that maintaining the N / K balance while reducing the concentration of N from 7.1 to 3.6 meq L−1 and K from 3.1 to 1.5 meq L−1 did not affect growth and flower production. In our study the variations in the N and K concentration affected the growth of lisianthus despite the N / K balance being held constant. However, it would appear that R. asiaticus is also insensitive to the N / K balance as in another study, Bernstein et al. (Citation2011) demonstrated that N / K balances from 0.8 to 3.5 did not affect biomass production, although the yield and marketability of flowers decreased when the N / K balance was decreased from 2.1 to 0.8.
Effect of N and K concentrations on plant growth
The results of the present study demonstrate that the N / K balance was as important as the respective nutrient concentration for obtaining maximum growth. Furthermore, the response of lisianthus was associated not only to the N and K balance but also to the concentrations, as suggested by the root, shoot, and total plant dry weight, that were significantly affected by the levels of N and K (). The effect of N and K concentration is demonstrated in plants irrigated with solutions with a balance of 2.14 as they exhibited increased dry weight in all plant parts when the N and K concentration was reduced from 15 to 9 meq L−1 and from 7 to 4.2 meq L−1, respectively (). When compared to plants fertigated with the control solution, increasing the N / K balance to 4.29 resulted in increased dry weight only when the level of N and K was decreased to 12 and 2.8 meq L−1, respectively (). Further decreases in N and K were associated with growth similar to that of the control plants (). Plants irrigated with solutions with a N / K balance of 1.43 rendered similar dry weight as that of the control plants, regardless of the N and K concentration (). Other growth parameters such as plant height, number of flower buds and leaf area were unaffected by the N and K concentration in the nutrient solution ().
Figure 1. Effect of nitrogen (N) and potassium (K) balance and concentration in the nutrient solution on dry weight of lisianthus plants.
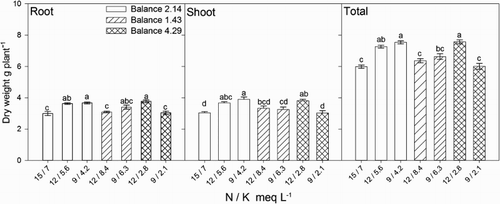
Table 3. Average plant growth and dry weight (DW) responses of lisianthus fertigated with nutrient solutions of varying nitrogen (N) and potassium (K) balances and concentrations.
The effect of the N and K concentration has also been demonstrated in chrysanthemum. Azeezahmed et al. (Citation2016) assessed the effect in five nutrient solutions which had a N / K balance of 3.5 during the vegetative phase and ∼2.0 during the reproductive phase. Their results indicated that there was a concentration effect as chrysanthemum fertigated with solutions containing N and K at 17.9 and 5.1 meq L−1, respectively, resulted in the greatest plant growth and quality during the vegetative phase, whereas in the reproductive phase the optimum concentration was 14.3 and 6.7 meq L−1 of N and K, respectively.
Nutrient allocation and concentration
Nitrogen and Ca concentration was on average 50% and 230% higher in the roots than in the shoots of lisianthus, respectively, while Mg was 60% higher in the shoots than in the roots (). Internal N concentration ranged from 866 to 1321 mmol kg−1 in the shoot and from 1089 to 2092 mmol kg−1 in the root (). These concentrations are low when compared to other ornamental species; for example, chrysanthemum cuttings contain from 2693 to 4029 mmol kg−1 of N (Druege et al. Citation2000) while in the leaves it ranged from 3000 to 3407 mmol kg−1 (Barbosa et al. Citation2000). Internal N of lisianthus was not associated with its concentration in the nutrient solution, as plants fertigated with N at 15 meq L−1 resulted in lower N in shoots and roots, when compared to plants fertigated with solutions of lower N concentrations (). This suggest that lisianthus does not require high levels of internal N and that this low demand may be met with low concentrations of N in the nutrient solution. Similar results were reported by Gómez-Pérez et al. (Citation2014), who demonstrated that lisianthus contained comparable concentrations of N and Mg in the shoots. However, our results and those by Gómez-Pérez et al. (Citation2014) are in contrast to other results, in that, high N and Ca concentration in the leaves (Mendoza-Villarreal et al. Citation2015) and total plants (∼2300 and 1948 mmol kg−1 respectively) (Hernández-Pérez et al. Citation2016), have been reported.
Figure 2. Effect of nitrogen (N) and potassium (K) balance and concentration in the nutrient solution on shoot and root N concentration of lisianthus plants.
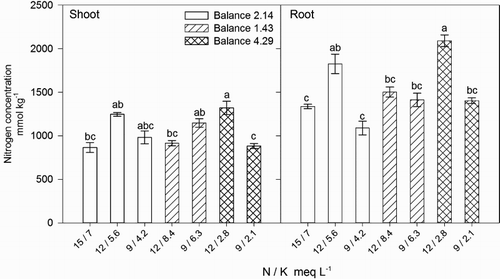
Table 4. Average nutrient concentrations (mmol kg−1) in plant parts of lisianthus fertigated with nutrient solutions of varying nitrogen (N) and potassium (K) balances and orthogonal contrasts.
The preference for higher N and Ca accumulation in the roots and, compared to other reports, the lower N concentrations that we report in the present study, may be due to the form in which it has been supplied, either as NO3− or NH4+. In the present study lisianthus was fertigated with solutions containing N exclusively in the NO3−-form, whereas in other lisianthus studies in which N was supplied as NH4+ there was a higher N and Ca concentrations. The higher concentration of N and Ca in the roots when fertigated with solutions containing N in NO3− form suggests that there is a restriction in transport of both nutrients to the shoots, which may be associated with the calcifuge trait of lisianthus (Hernández-Pérez et al. Citation2016). Calcifuge species are adapted to grow under low Ca concentrations (White and Broadley Citation2003), by increasing the Ca influx through the plasma membrane and decreasing Ca efflux from the cytosol (Lee Citation1999), and by preferring N in NH4+ form (Hawkesford et al. Citation2012).
Internal nutrient concentration and plant growth
Increasing shoot N concentration () was associated with higher shoot dry weight (), except when plants were fertirrigated with solutions containing N and K at 9 and 4.2 meq L−1, respectively, which resulted in plants with high shoot weight and low N concentration, probably due to a dilution effect. Low external N concentration, in the solution with N and K at 9 and 4.2 meq L−1, respectively, did not impede plant growth; we suggest that this was probably associated with the higher N allocation to the shoot as suggested by the lower N allocation to the roots (N was only 10% higher than that of the shoot), when compared to other plants receiving other N concentrations (N was up to 64% higher than that of the shoot) (). We did not observe this N allocation in plants irrigated with other low N solutions, i.e. 9 and 2.1 meq L−1 of N and K, respectively (), probably due to the extremely low K concentration in the nutrient solution, which, affected the transport of NO3− from the roots to the shoots (Hawkesford et al. Citation2012), as suggested by the 59% higher N concentration in the roots than in the shoot in plants fertigated with this nutrient solution ().
Shoot N concentration was not correlated with N concentration or the N / K balance in the nutrient solution, however, increasing N in the shoot () was associated with increasing shoot P concentration (Shoot P = 0.021[Shoot N] + 4.63, R2 = 0.929) () and Mg (Shoot Mg = 0.080[Shoot N] + 71.5, R2 = 0.936) (). In general, P, Ca, and Mg concentration in plant tissues were unaffected by the external N / K balance (), but the internal P () and Mg () concentrations were correlated with shoot dry weight (Shoot dry weight = 0.135[Shoot P] + 3.21, R2 = 0.621 and Shoot dry weight = 0.037[Shoot Mg] + 1.056, R2 = 0.693) (). We suggest that internal N concentration was regulated not by the external N concentration but by plant uptake in response to the effect of K on the allocation of NO3− to the shoots. However, N accumulation increased plant growth by enhancing the uptake of P and Mg.
Figure 3. Effect of nitrogen (N) and potassium (K) balance and concentration in the nutrient solution on shoot and root phosphorus concentration of lisianthus plants.
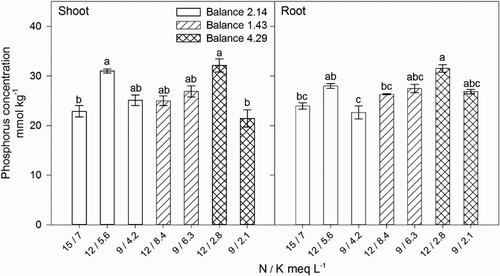
Figure 4. Effect of nitrogen (N) and potassium (K) balance and concentration in the nutrient solution on shoot and root magnesium concentration of lisianthus plants.
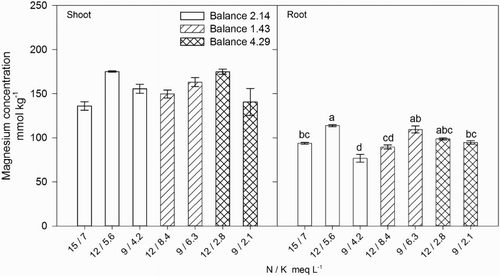
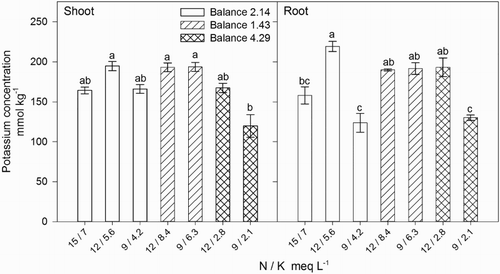
Shoot K concentration significantly increased when the N / K balance in the nutrient solution decreased (), which we suggest is related to the higher concentrations of external K in the nutrient solutions. Maximum growth was observed with a N / K balance of 2.14 and a N and K concentration of 12 and 5.6 meq L−1, respectively, which resulted in the highest root and shoot K concentrations (). Higher K concentration in the nutrient solution was directly correlated with increasing shoot K. Higher K uptake and accumulation when external K concentration increases has been attributed to the passive transport of K through the plasma membrane channels (Nieves-Cordones et al. Citation2014). Higher shoot dry weight was associated with internal K concentrations of ∼166–167 mmol kg−1, while concentrations of ∼120 or ∼193–195 mmol kg−1 were associated with reduced growth, except when the N / K balance was 2.14 (). This was probably due to a concentrations effect of K as plants resulted with lower dry weight.
Figure 5. Effect of nitrogen (N) and potassium (K) balance and concentration in the nutrient solution on shoot and root potassium concentration of lisianthus plants.
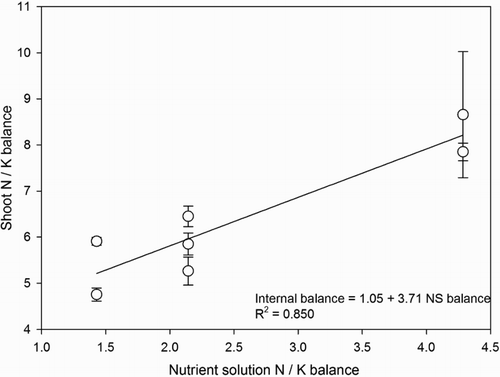
According to Joiner et al. (Citation1983), it is important to consider if plants are responding to the external nutrient balance imposed by the nutrient solution concentrations or to the internal balance. In our study, the internal N / K balance was directly influenced by the balance in the nutrient solution (), however, the internal balance was not associated with plant growth.
Figure 6. Correlation between nitrogen (N) and potassium (K) balance in the nutrient solution and N / K balance in the shoot of lisianthus plants.
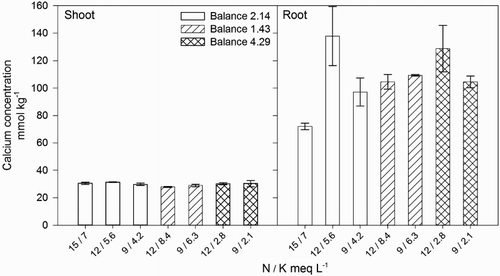
Shoot and root Ca concentration were unaffected by the external N / K balance or the N and K concentrations in the nutrient solutions (, ), however, Ca in the shoot exhibited a quadratic trend as it increased when the N / K balance in the nutrient solution was increased (Shoot Ca = −1.05[N / K]2 + 6.62[N / K ] + 21.2, R2 = 0.747). This trend indicated that maximum shoot Ca is achieved by plants when the external N / K balance was 2.14 or higher, which in turn suggests that low N / K balances, i.e. high K, may interfere with Ca uptake by roots and transport to the shoot due to the antagonism between both cations (Fageria Citation2001).
Figure 7. Effect of nitrogen (N) and potassium (K) balance and concentration in the nutrient solution on shoot and root calcium concentration of lisianthus plants.
In conclusion, the optimum N / K balance for lisianthus was 2.14, however, there was also a concentration effect as fertigation with solutions containing this balance but N and K concentration at 9 and 4.2 meq L−1, respectively, resulted in the plants with the highest dry weight. The external N / K balance had a direct effect on the internal N / K balance, however, plant growth was more affected by N concentration in plant tissues, which in turn was related with a linear increase in P and Mg concentration in plants. Nitrogen P and Mg tissue concentration had a direct linear impact on plant growth.
Disclosure statement
No potential conflict of interest was reported by the authors.
Notes on contributors
Dr Daniela Alvarado-Camarillo has a postdoctoral position at Universidad Autónoma Agraria Antonio Narro, Saltillo, Coah., México. She received her Doctor of Science in protected agriculture at the Universidad Autónoma Agraria Antonio Narro in 2017. She is currently working on hydroponic cultivation of ornamental plants, floriculture, and plant nutrition.
Dr Ana María Castillo-González is a senior full time researcher and professor in the Departamento de Fitotecnia in Universidad Autónoma Chapingo. Her research is focussed on the determination of optimum nutrition for horticultural plants, including ornamentals, top fruit, and soft fruit. She received her Doctor of Science in fruticulture at the Colegio de Postgraduados, México, in 1997 and has published 35 papers, one book, and four book chapters.
Dr Luis A. Valdez-Aguilar is a senior full time researcher and professor at Universidad Autónoma Agraria Antonio Narro. His research is mainly focussed on hydroponic cultivation of greenhouse ornamental and vegetable species, nutrient interactions, and design of fertilisation programmes for ornamentals and hydroponic systems. He received his PhD in horticulture from Texas A&M University in 2004 and has published 75 papers.
Dr Juana C. García-Santiago has a postdoctoral position at the Facultad de Ciencias Agropecuarias, Universidad Autónoma del Estado de Morelos, Cuernavaca, Morelos, México. She received her Doctor of Science in protected agriculture at Universidad Autónoma Agraria Antonio Narro in 2016. She is currently working on hydroponic cultivation of vegetables and plant nutrition.
ORCID
Luis A. Valdez-Aguilar http://orcid.org/0000-0002-2510-1962
References
- Armitage AM, Laushman JM. 2003. Specialty cut flowers. The production of annuals, perennials, bulbs, and woody plants for fresh and dried cut flowers. 2nd ed. Portland (OR): Timber Press, Inc.
- Azeezahmed SK, Dubey RK, Kukal SS, Sethi VP. 2016. Effect of different nitrogen-potassium concentrations on growth and flowering of chrysanthemum in a drip hydroponic system. J Plant Nutr. 39:1891–1898. doi: 10.1080/01904167.2016.1187749
- Barbosa JG, Kampf AN, Martinez HE, Koller OC, Bohnen H. 2000. Chrysanthemum cultivation in expanded clay. I. Effect of the nitrogen-phosphorus-potassium ratio in the nutrient solution. J Plant Nutr. 23:1327–1336. doi: 10.1080/01904160009382103
- Bernstein N, Ioffe M, Luria G, Bruner M, Nishri Y, Philosoph-Hadas S, Salim S, Irit Dori S, Matan E. 2009. Evaluation of a newly developed potassium and nitrogen fertilization regime for soil cultivation of Ranunculus asiaticus. Israel J Plant Sci. 57:411–420. doi: 10.1560/IJPS.57.4.411
- Bernstein N, Ioffe M, Luria G, Bruner M, Nishri Y, Philosoph-Hadas S, Salim S, Irit Dori S, Matan E. 2011. Effects of K and N nutrition on function and production of Ranunculus asiaticus. Pedosphere. 21:288–301. doi: 10.1016/S1002-0160(11)60129-X
- Bremner JM. 1996. Total nitrogen. In: Sparks DL, editor. Methods of soil analysis. Part 3. Chemical Methods. Madison (WI): Soil Science Society of America; p. 1085–1086.
- Broschat TK, Sandrock DR, Elliott ML, Gilman EF. 2008. Effects of fertilizer type on quality and nutrient content of established landscape plants in Florida. HortTechnology. 18:278–285.
- Castillo-González AM, Avitia-García E, Valdez-Aguilar LA, Velázquez-Maldonado J. 2017. Extracción nutrimental en lisianthus (Eustoma grandiflorum [Raf.] Shinn) cv. Mariachi Pink. Revista Mexicana de Ciencias Agrícolas. 8:345–354. doi: 10.29312/remexca.v8i2.55
- Druege U, Zerche S, Kadner R, Ernst M. 2000. Relation between nitrogen status, carbohydrate distribution and subsequent rooting of chrysanthemum cuttings as affected by pre-harvest nitrogen supply and cold-storage. Ann Bot. 85:687–701. doi: 10.1006/anbo.2000.1132
- Fageria VD. 2001. Nutrient interactions in crop plants. J Plant Nutr. 24:1269–1290. doi: 10.1081/PLN-100106981
- Fiorenza S. 2017. Lisianthus. http://www.sergiofiorenza.it/lisianthus.htm. [12 December 2017].
- Frett JJ, Kelly JW, Harbaugh BK, Roh M. 1988. Optimizing nitrogen and calcium nutrition of lisianthus. Commun Soil Sci Plant Anal. 19:13–24. doi: 10.1080/00103628809367916
- Gómez-Pérez L, Valdez-Aguilar LA, Sandoval-Rangel A, Benavides-Mendoza A, Mendoza-Villarreal R, Castillo-González AM. 2014. Calcium ameliorates the tolerance of lisianthus [Eustoma grandiflorum (Raf.) Shinn.] to alkalinity in irrigation water. HortScience. 49:807–811.
- Halevy AH, Kofranek AM. 1984. Evaluation of lisianthus as a new flower crop. HortScience. 19:845–847.
- Harbaugh BK. 2007. Lisiantus. Eustoma grandiflorum. In: Anderson NO, editor. Flower breeding and genetics. The Netherlands: Springer; p. 645–663.
- Hawkesford M, Horst W, Kichey TMR, Schjørring JK, Møller IS, White P. 2012. Functions of macronutrients. In: Marschner P, editor. Marschner’s mineral nutrition of higher plants. USA: Elsevier Science; p. 135–189.
- Hernández-Pérez A., Valdez-Aguilar LA, Villegas-Torres OG, Alía-Tejacal I, Trejo-Téllez LI, Sainz-Aispuro MDJ. 2016. Effects of ammonium and calcium on lisianthus growth. Hortic Environ Biotechnol. 57:123–131. doi: 10.1007/s13580-016-0004-1
- Huber DM, Arny DC. 1985. Interactions of potassium with plant disease. In: Munson RD, editor. Potassium in agriculture. Madison (WI): Soil Science Society of America; p. 467–488.
- Joiner JN, Poole RT, Conover CA. 1983. Nutrition and fertilization of ornamental greenhouse crops. In: Janick J (Ed.), Horticultural reviews, Volume 5. Westport (CT): The AVI Publishing Company Inc; p. 317–403.
- Lee JA. 1999. The calcicole–calcifuge problem revisited. Adv Bot Res. 29:1–30.
- Marchese JA, Katz I, Sousa AP, Rodrigues JD. 2005. Gas exchange in lisianthus plants (Eustoma grandiflorum) submitted to different doses of nitrogen. Photosynthetica. 43:303–305. doi: 10.1007/s11099-005-0050-x
- Mendoza-Villarreal R, Valdez-Aguilar LA, Sandoval-Rangel A, Robledo-Torres V, Benavides-Mendoza A. 2015. Tolerance of lisianthus to high ammonium levels in rockwool culture. J Plant Nutr. 38:73–82. doi: 10.1080/01904167.2014.920379
- Nieves-Cordones M, Alemán F, Martínez V, Rubio F. 2014. K+ uptake in plant roots. The systems involved, their regulation and parallels in other organisms. J Plant Physiol. 171:688–695. doi: 10.1016/j.jplph.2013.09.021
- PanAmerican Seed. 2017. Cut flower lisiatnhus. Grower Facts. https://www.panamseed.com/media/Culture/PAS/LisianthusCutflower.pdf [12 December 2017].
- Sakata. 2017. Cut flower lisianthus. http://www.sakataornamentals.com/_ccLib/attachments/plants/Lisianthus+Cut+Flower-0216-SAKATA.pdf [17 December 2017].
- Soltanpour PN, Johnson GW, Workman SM, Jones JB, Miller RO. 1996. Inductively coupled plasma emission spectrometry and inductively coupled plasma mass spectrometry. In: Sparks DL, editor. Methods of soil analysis. Part 3. Chemical Methods. Madison (WI): Soil Science Society of America; p. 91–139.
- Sonneveld C, Voogt W. 2009. Plant nutrition of greenhouse crops. New York: Springer.
- Valdez-Aguilar LA, Grieve CM, Poss JA. 2013. Response of lisianthus to irrigation with saline water: plant growth. J Plant Nutr. 36:1605–1614. doi: 10.1080/01904167.2013.799188
- White PJ. 2012. Ion uptake mechanisms of individual cells and roots: short-distance transport. In: Marscher P, editor. Marschner’s mineral nutrition of higher plants. USA: Elsevier Science; p. 7–47.
- White PJ, Broadley MR. 2003. Calcium in plants. England Ann Bot. 92:487–511. doi: 10.1093/aob/mcg164