
ABSTRACT
The problem raised here is the intensification of technology in field crops of winter oilseed rape. Is there a justification for this by explaining what relationship between the yield and individual yielding traits are formed in hybrid and population cultivars. During years 2012–2015 field studies to evaluate five hybrid and five population cultivars response on the standard and high-input technology in winter oilseed rape production have been executed in Poland. High-input technology included: double foliar application (in autumn, and in spring) of micro-nutrients, additional top fertilisation of 40 kg N per ha in spring, double application of the amino-acid bio-stimulators, and additional growth regulator. According to the principle component analysis made for both technologies it was found that hybrids and population cultivars displayed varied response which is explained here on the basis of morphological, growth, and yield elements. The high-input technology improved the yield responded elements e.g. number of siliques and number of seeds per silique as well as winter hardiness. This impact was more evident for population cultivars than for hybrids. We state that high-input technology, as the proposal in this paper, that provided 9.6% increase in seed yield, is biologically justified for winter oilseed rape crop and can be recommended in North and central European countries.
Introduction
Over the past 50 years, there has been a two-fold increase in the production of oilseeds and almost five-fold increase in the area of oil seed rape (Brassica napus var. oleifera) crops in the world. Currently covers about 0.6% of the area of all crops in the world, which is 33 million ha, where the production of 67.7 million t provides 20 million t of edible oil and components needed to produce diesel oil (Jiang et al. Citation2014). The increase in the yield potential of rapeseed varieties and the possibility of a wider and fuller use of seeds contribute to the increase of crop area of this species in Europe (Jaradat Citation2016).
According to Balodis and Gaile (Citation2009) sowing technology winter oilseed rape is similar in European countries, where the date is from the beginning of August to the end of the first decade of September (Rathke et al. Citation2006; Rummukainen Citation2010; Crnobarac et al. Citation2015).
The environmental factors remain in interaction with the optimal sowing date such as soil temperature and moisture, which affects the preparation of the seed bed and germination process (BBCH 00-09) (Lancashire et al. Citation1991) of rapeseed. Aspects that should be taken into account when choosing the sowing date are: (1) ensuring that it will be possible to obtain a high field emergence capability, which is called the FEC index (field effective capacity), (2) optimisation of mass and amount of harvested plant parts, (3) control of the development cycle, in order to reduce the occurrence of periods unfavourable for crops, i.e. abiotic (frost, drought) and biotic (agrophages) stresses (Rathke et al. Citation2006). Rape responds more strongly to air humidity than to rainfall and in years with higher relative humidity, higher yields are harvested even with lower rainfall. The temperature is extremely important for the proper enzymatic activity and proper functioning of the plant (Bonhomme Citation2000; Ozolincius et al. Citation2005). Expected challenges in the face of climate change, including increased frequency of drought, higher temperatures, unpredictable and irregular cycles of precipitation and drought, soil salinity, flooding, hail and challenges resulting from existing and new diseases, in the coming years will set the directions of rape farming (Povilaitis and Lazauskas Citation2010; Salisbury and Barbetti Citation2011; Barbetti et al. Citation2012). Weymann et al. (Citation2015) state that the crucial phonological stages in yield formation are: the beginning of siliques (BBCH 50-65) and seed development (BBCH 71-79). The authors claim that in winter oilseed rape there are very large compensatory effects between the components of the seed yield, especially between the number of seeds per m2 and the TSW.
Winter rape crop has very high nutritional requirements and reacts with a significant increase in yield both under the influence of soil and foliar fertilisation. To produce 1 ton of rape seeds, the demand for macronutrients is: 45 kg N, 20 kg P2O5, 50 kg K2O, 12.5 kg MgO, 12.5 kg S (Barłóg and Grzebisz Citation2004a). The nitrogen dose must be determined precisely, both for economic reasons and due to the need to eliminate environmental hazards (Horst et al. Citation2002; Dresbøll et al. Citation2016). It is also very important to know the physiological way of nitrogen management in winter oilseed rape. The improvement of the efficiency of using N fertiliser in winter oilseed rape can be obtained in several ways: either by improving the intake of N from the soil (Garnett et al. Citation2009; Ulas et al. Citation2012), or by improving the ability to stop it in crop thanks to remobilisation (Masclaux-Daubresse et al. Citation2010), or by reducing aboveground biomass and N losses due to losses of old leaves (Sieling and Kage Citation2008). The first way to improve the uptake of N from the soil concerns the cultivation of varieties with more extensive and deeper root systems that reach the soil nitrogen resources. The highest uptake of winter oilseed rape occurs mainly in autumn and again in the spring from the elongation to flowering phase, while in the silique development phase and the maturation of seeds, N consumption is the smallest (Malagoli et al. Citation2005). Nitrogen for siliques and seeds is remobilized from previously accumulated in the stem and leaves (Avice and Etienne Citation2014). Nitrogen accumulated in the stem leaves is lost, because these leaves die during the growing season. Results of Wang et al. (Citation2016) suggest that efficient transport systems and developed root systems play important roles in B. napus N acquisition under N starvation. It is estimated that about 45 kg N per hectare per year is lost due to the loss of leaves from the stem forming phase to ripening (Dejoux et al. Citation2000; Malagoli et al. Citation2005). In addition, including the loss of rosette leaves during the winter, before forming the stem, these estimates reach 100 kg N per ha per year (Dejoux et al. Citation2000). It is certainly possible to develop new varieties of rapeseed with improved efficiency in the use of nitrogen in the near future. F1 hybrids tend to yield the highest compared to other varieties in different variants of nitrogen fertilisation. Research is also being carried out on the use of new hybrids in several varieties of fertilisation regimes. However, it should be remembered that not only the better use of nutrients by rapeseed is taken into account by breeders, but also the translocation in the plant and their metabolism (Berry et al. Citation2010; Ulas et al. Citation2012).
High-input technology is often practice in intensive crop production of winter rape. In the context of increasing nitrogen dose in the spring top fertilisation, foliar nutrition along with more intensive protection against diseases combine with bio stimulation we questioned whether it has the biological justification. To face this motivation for this research it was hypothesised that 10 cultivars selected for the study (5 population and 5 hybrids) will show a differentiated response to both technologies, which can be explained on the basis of morphological features, growth and yield structure elements. The hypothesis was to demonstrate of cooperation between selected rapeseed cultivars and technology which was used to determine for which cultivars the standard/high-input technology is profitable.
Material and methods
Field trial localisation
The field experiment was carried out from 2012 to 2015 as three-years. Plots were located in Grocholin (52°59′24″N 17°25′36″E). Topography is a very low-lying and the climate of the region is temperate and influenced both by Atlantic ocean and Asian continent. The average January temperature is −4 degrees and in July 18°C.
Experimental details
Plots were located on soil classified as Alfisols formed on sandy loams according to Polish Soil Classification, with regulated pH at 6.8. The soil’s availability in macronutrients was high (P2O5) and very high (K2O and MgO), according to the annual soil analysis made each year after harvest of pre-crop. The pre-crop for winter oilseed rape was: spring barley in 2012 and winter wheat in 2013 and 2014. After harvesting the pre-crop, the Grubber aggregate was used to a depth of 15 cm, then after about 2 weeks, ploughing was performed to a depth of 25 cm with adding Crosskill aggregate. Pre-sowing fertilisation in the form of multicomponent fertiliser was made 2–3 days before sowing, and immediately before sowing, the cultivator was applied to a depth of 5 cm. Sowing dates, in accordance with agro technical recommendations, as for the Cuyavia-Pomerania region, were between August 31 and September 4. Sowing was carried out with the Deppe seed drill, working width 1 m, in row spacing of 25 cm. The seed was characterised by very high parameters, i.e. 99% purity and sprouting ability also above 90%. The seed harvest was carried out in one stage, using the Zürn 150 combine, between 19 and 26 July. The size of the plots for sowing was 12 m2 and to harvest 10 m2.
Factors and treatments
Factor A: cultivation technology. The specification of all chemical practices is presented in . The treatments were set up as standard technology (A1) and high-input technology (A2) which is consisted of differentiation:
Double foliar application (in autumn, and in spring) of micro-nutrients.
Additional top fertilisation of 40 kg N per ha in spring.
Double application of the amino-acid bio-stimulators.
Additional growth regulator in spring.
Table 1. Treatments used in standard (A1) and high-input (A2) technology in winter oilseed rape.
Factor B: winter oilseed rape cultivar in population group: Adriana, Bellevue, Bogart, Kadore, Monolit, and in hybrid group F1: DK Exquisite, Müller 24, NK Technic, Poznaniak, Rohan.
The experiment was design as a split-plot with technology treatment in the sub-block and cultivars on the plots randomly arranged. Four blocks were set up of each 20 combinations.
Statistical analyses
The assumption that treatment variances are equal in three years of field trials was checked according to Bartlett’s test for homogeneity of variance. The working hypotheses concerning the effects of the cultivation technology (Factor A), cultivars (Factor B) and their interaction (AB) as the fixed factors were verified by the analysis of variance (ANOVA) with the test of significance at P ≤ 0.05. The GLM of ANOVA included three years and four replicates in split-plot model. For significant effects from ANOVA the dependent variables (presented in ) means were separated using HSD Tukey’s test at P < 0.05. The correlations were calculated using r-Pearson’s coefficient with significance at P < 0.05. The exploration technique of the main components PCA (principle component analysis) has been used to explain the multi-dimensional diversity of winter oilseed rape in standard and high-input technology and classification of winter rape cultivars in terms of the first two components.
Table 2. Vegetal characteristic of oil seed rape cultivars depending on standard (A1) and high-input technology (A2). Mean of 2013–2015 and deviation of mean [%].
Table 3. Morphological characteristics of oil seed rape cultivars depending on standard (A1) and high-input technology (A2). Mean of 2013–2015 and deviation of mean [%].
Table 4. Productivity of oil seed rape cultivars depending on standard (A1) and high-input technology (A2). Mean of 2013–2015 and deviation of mean [%].
Twelve variables (characteristics) of winter rape were used for PCA by giving them symbols from x1 to x12 according to the following assignment: x1 plant density in spring; x2 winter hardiness; x3 plant losses; x4 canopy height; x5 canopy loading; x6 branches per plant; x7 siliques per plant; x8 LAI (leaf area index); x9 HI (harvest index); x10 TSW (thousand seed weight); x11 seeds per silique; x12 seed yield; Y1 i Y2 1st and 2nd components.
The calculations were performed in the STATISTICA 12.5 programme, Stat Soft Poland.
Results and discussion
The FEC set for this research was 50 plants per m2 for population varieties and 40 plants per m2 for hybrid, as recommended by breeders. The emergencies were very even, however, the spring density was lower than in the autumn: from 38.3 in 2013 in A1 technology to 41.8 plants per m2 in A2 technology in 2012 (data not presented here). In the 3 year synthesis of variance the largest density in spring was obtained by population cultivars: Kadore and Bogart - 44.2, Bellevue and Monolit - 44.0 and 43.0 plants per m2 (). However, among the hybrids, the highest spring density displayed DK Exquisite and Müller 24, i.e. 38.2 and 39.1 plants per m2. In 3 year term, the intensity of rape technology had no impact on spring density, as well as no interaction between cultivar and technology was detected. The optimal density of plants varies depending on the rape growing region and it ranges from 80 to 150 before winter and 60–80 plants per m2 at the beginning of spring vegetation. For hybrid oilseed rape in Germany, smaller FEC are recommended (60–70 units per m2) (Diepenbrock Citation2000), while in the Nordic countries even at 30–70 plants per m2 (Velička et al. Citation2006). The required optimal density changes over time along with the availability of knowledge about the oil seed rape cultivars. Homogeneous distribution of plants on the field is a prerequisite for yield stability, because, as the results of earlier works indicate, plants are then less exposed to environmental stress (Sierts et al. Citation1987). Jankowski and Budzyński (Citation2007) demonstrated that during the spring vegetation proper and uniform water supply, where FEC was 60 plants per m2, guaranteed harvest density to the level of 46 plants per m2 and the highest plant productivity. This tendency to reduce the density of modern rapeseed cultivars was confirmed in our own research, where the overall plant density during harvest amounted to 35.9 per m2, and population cultivars had higher than hybrids about 3–5 plants per m2, while between standard (A1) and high-input (A2) technologies by 1.3 plants (). It is a proof that optimal weather conditions (and such prevailed here in entire study cycle) and standard technology successfully provide to the harvest density as to the first element that stabilise and lead to the high crop production. In practice, based on survey study conducted in 2004–2006 on nearly 250 winter rape plantations in Wielkopolska region, it was shown that the average density in autumn was 79.5 plants per m2, and most often it was in the range of 40–80 plants per m2 (Muśnicki and Gładysiak Citation2008). To reach an ideotype, i.e. a model of ideal rapeseed canopy, all yield elements should be consider (both primary and secondary) (Diepenbrock and Grosse Citation1995). According to Diepenbrock (Citation2000) FEC is the primary element of the yield structure that regulates the remaining elements of the crop, and thus it productivity. The density of plants, provided by FEC, has a huge impact on the growth, development, and yield of winter rape as well (Leach et al. Citation1999; Diepenbrock Citation2000).
The significant relationships were detected between the spring density of plant and further characteristics of plants and crop (–3). The increase of density resulted an increase in vegetative losses (r = 0.62) and a decrease in the canopy height (r = −0.62), and consequently an increase of the canopy lodging (r = 0.48), especially in high-input technology (r = 0.56). The increasing spring density positively affected LAI (r = 0.57), but at the same time reduced the number of siliques (r = −0.63) and the number of seeds in siliques (r = −0.78) (). LAI was measured in BBCH 53 phase i.e. in the budding phase. It is a trait that has an affinity for other morphological features, primarily to the number and size of leaves, but also to the number of branches and the height of the shoot. It was found that high-input caused that LAI reached higher values by 0.63 than in standard technology. Among the population varieties, Bogart and Monolit displayed LAI at the highest level, respectively 8.63 and 8.23 (). In exact research made by Kaczmarek et al. (Citation2003) an high-input and standard technology for winter rape was a premise for cultivars research in many localities. According to them, the productivity of oil seed cultivar is more dependent on weather conditions than on the potential productivity of habitat. However, the estimation of whether the cultivar will respond positively to high-input technology depends to a large extent on both weather and habitat conditions. Our research showed the positive effect of high-input technology on the overwintering of rapeseed cultivars at the level of 65.9%, i.e. by 2.3 percentage points higher than in standard technology was obtained (). This should be associated with additional microelement fertilisation in A2 technology, applied as foliar in 1 l per ha (Wenda-Piesik et al. Citation2017) plus boron in 1 kg per ha. In this study hybrid cultivars improved higher hardiness in the range of 63.3–71.2%, among which Müller 24 in 13.7% and DK Exquisite and Poznaniak in 8.8% even higher in A2 technology from overall cultivars. However, the range for population cultivars ranged from 57.0% for Adriana to 63.2% for Kadore and Bogart, which also responded positively to A2 technology in overwintering by 1.6 and 2.9% (). The increase of overwinter degree had a positive relationship with the yield of winter rape, through an increase in the number of siliques (r = 0.62) and an increase in the number of seeds in siliques (r = 0.63) (). Similar results, but without correlation analysis, were obtained by Jankowski and Budzyński (Citation2007), who describe that in the years with the mild winter, oilseed rape obtained the highest yield of seeds.
Table 5. Correlations (r – Pearson’s coefficient) between some characteristics of plant and crop of winter oilseed rape.
The siliques number on the plant is the next first-row element which is decisive for the rape productivity. This characteristic is ultimately determined by the number of branching shoots, flower buds and young pods. Here, in own study, confirmation was also found in high correlations between the number of branches and the number of siliques at the level of r = 0.62–0.85 (). Jankowski and Budzyński (Citation2007) reported that rape hybrids produced significantly more siliques on the plant and were better filled with seeds than the population cultivars, however, they produced lower TSW at 8%. Chay and Thurling (Citation1989), Leach et al. (Citation1999) reported that the number of siliques on the plant is negatively correlated with the plant density during harvest and this could be explain by the crop architecture elated to the number of siliques. Geisler and Henning (Citation1981) calculated that increasing density from 9 to 50 plants per m2 caused a drastic decrease in branching by 28%, and consequently the number of siliques on the plant. In European countries, including Poland, where rapeseed is sown at the end of summer, the plants produce very large biomass and about 5.000 floral buds on the cone during crypto genesis. From this huge amount, about 300–500 buds are formed, and further 200 siliques are produced (Sova et al. Citation1998). In this study, from the cultivars with the largest number of siliques on the plant, the population Bogart should be mentioned, which had the highest number of siliques (up to 391), and in high-input technology even 24.5% higher than in the standard one, also the Kadore, in which high-input technology caused the increase by 17.2% siliques number (). Among hybrids, the most abundant siliques were produced by Poznaniak (272), NK Technik and DK Exquisite (255–251), although only NK Technik proved a significant impact of A2 technology on the increase of siliques by 19.8% ().
The last yield elements are the number of seeds in silique and the weight of the seeds presented here in . Often, instead of the mass of a single seed, the concept of one thousand seeds weight (TSW) in grams is used. Here, the number of seeds in silique was very variable in the years of research, however it was modified by technology, cultivars and interaction between them. In A2 technology, the seeds in the silique were on average 3–4 more than in A1. Hybrids: Poznaniak, NK Technik and DK Exquisite had significantly more seeds (26.5–24.7). On the other hand, among the population cultivars, the more abundant seeds were produced by the Bogart type (21.9) and Kadore (18.3), and both responded positively to the high-input technology. The negative relationship shows this characteristic along with the plant density and the number of seeds in silique (Jenkins and Leitch Citation1986), while the number of seeds in the silique is positively correlated with the silique length Diepenbrock (Citation2000). Therefore, the author states that this is a trait that indirectly determines productivity in rapeseed plants. In TSW, the technology effect caused 0.10 g higher weight in A2 than in A1. In contrast, the genetic factor was clearly revealed in population cultivars: Bellevue (6.20 g), Adrian (6.14 g) and Monolit (5.99 g), and in hybrids DK Exquisite (5.91 g). The increase in the seed yield of hybrids is the result of combined a greater number of branches on the plant and a better setting of siliques (Wielebski Citation2009). This was partially confirmed in this study, because the highest yielding DK Exquisite had one of the largest number of siliques (an average of 251 per plant, from 154 to 334 in the years of research), but at the same time the TSW was the highest of all hybrid cultivars (5.91 g) ().
The biological yield of rapeseed is measured by total biomass, while the economic yield is the rape seed yield as an economically used part of plants (Diepenbrock Citation2000). The harvest index (HI) is the proportion of the dry mass of seeds to the dry matter of the aboveground part of the plant. In the present research, the seed yield and above-ground biomass were measured for the same dry matter content, i.e. 87%, in g per plant, and during study were obtained as follow: in 2013 (0.46), in 2015 (0.41) in 2014 (0.40). It was 100% interdependent with yielding of rape in the years 2013 - 63.4 dt/ha, 2015 - 55.5 dt/ha−1 and 2014 - 50.7 dt/ha (data not presented here). Of the 10 winter rapeseed cultivars studied here, the highest HI were obtained by DK Exquisite, Müller 24 and Bellevue (0.45–0.46), while the lowest by NK Technik and Poznaniak 0.36–0.37. There was also an interaction between the cultivars and the intensity of the technology to change the value of HI in the synthesis in the years of research (). Most preferably the hybrids reacted to the A2 technology, in which the HI was on average 0.50, i.e. Müller 24 - 19.7% and Rohan at 44% more in A2. However, among the population, two cultivars Monolit and Bellevue, significantly improved the HI in A2 technology (). Kessel et al. (Citation2012) also demonstrate the attribute of cultivars, especially hybrids, make better use of high-input nitrogen fertilisation. The author explains this with a longer period of vegetal development, during which the concentration of biomass occurs, up to phase BBCH 69-70. Comparing the breeding effect in winter oilseed rape with wheat, the HI in modern wheat varieties has been improved significantly, by 45–50%, while in winter oilseed rape this improvement is around 25–30% (Diepenbrock and Grosse Citation1995). The authors state that HI is the main parameter limiting the economic yield, while further increase in yield potential in rapeseed cultivars can be achieved by increasing this index, as, according to Grosse et al. (Citation1992) the genotypic correlation between the HI and the yield is stabilised. However, according to Jankowski et al. (Citation2016) more advantageous HI obtained a long-stemmed, hybrid Visby cultivar (38%) rather than two semi-dwarf hybrid cultivars Avenir - 35% and PR45D03 - 37%. In addition, these authors indicate that the highest HI was obtained by rape in the optimal sowing date (39%), while the index was significantly lower at the time of delayed or accelerated sowing.
The relationships between all vegetal traits, both plant and canopy, yielding elements and the seed yield, have been thoroughly analysed here using multidimensional principle component analysis PCA. This was done separately for standard (A1) and high-input (A2) technology. Twelve variables (characteristics) of winter rape were used for PCA by giving them symbols from x1 to x12 according to the following assignment: x1 plant density in spring; x2 winter hardiness; x3 plant losses; x4 canopy height; x5 canopy loading; x6 branches per plant; x7 siliques per plant; x8 LAI; x9 HI; x10 TSW; x11 seeds per silique; x12 seed yield; Y1 i Y2 1st and 2nd components.
For the standard technology (A1) the 1st component equation which explains 35.18% of variance (Y1) is: Y1 = −0.70 x1 + 0.44 x2 – 0.87 x3 + 0.79 x4 – 0.63 x5 + 0.17 x6 + 0.30 x7 – 0.74 x8 – 0.00 x9 + 0.24 x10 + 0.56 x11 + 0.74 x12. This component is positively associated with the yield of winter oilseed rape (0.74), and the canopy height (0.79), while significantly negatively with density in spring (−0.70), vegetative losses (−0.87) and LAI (−0.74). Thus, these characteristics stay in contrast to the seed yield. The 2nd component, responsible for 33.6% of total variance has the equation: Y2 = −0.50 x1 + 0.47 x2 – 0.13 x3 + 0.10 x4 + 0.57 x5 + 0.88 x6 + 0.91 x7 + 0.17 x8 – 0.68 x9 – 0.74 x10 + 0.76 x11 + 0.14 x12 that corresponds significantly and positively with the number of branches (0.88), the number of siliques (0.91) and the number of seeds (0.76), and significantly negatively for TSW (−0.74) and harvest index (−0.68). The impact of this component on the yield is neutral (0.14) ((A)). This is the confirmation of previous reports by Diepenbrock (Citation2000) that the strong associations concern the number of branches, the number of siliques and the number of seeds as first-row elements of yield. Factor coordinates for rape cultivars in the A1 technology show the greatest positive meaning of the 1st component for population cultivars, especially Kadore, Adriana and Monolit. This is component which keeps yield against density in spring, LAI, and plant losses. However, for four hybrids i.e. NK Technik, Poznaniak, DK Exquisite and Müller 24 these characteristics have a negative meaning, as they obtained negative coordinates to Y1. The meaning of the 2nd component (which corresponds positively to the number of branches, the number of siliques and the number of seeds and negatively for MTN and the harvest index) had the highest value for Bogart cv. and the smallest for the Bellevue cv. ((A)). He et al. (Citation2017) examined the N utilisation efficiency (NUtE) by agronomic and morphological traits of 50 oilseed rape genotypes and compared at contrasting N conditions across controlled (pot) and complex (field) environments. Seeds number per silique, harvest index and N harvest index were significantly correlated with NUtE under contrasting N conditions in all environments. Authors state that seeds number per silique may be a good indicator in rapidly selecting N efficient genotypes. Bouchet et al. (Citation2016) demonstrated that the effect of the trial was greater than the effect of nitrogen nutrition levels on seed yield-related traits under experimental conditions. Nevertheless, critical genomic regions associated with yield that were stable across environments were identified in rapeseed.
Figure 1. Projection of winter rape characteristics on two principle components at standard (A) and high-input (B) technology.
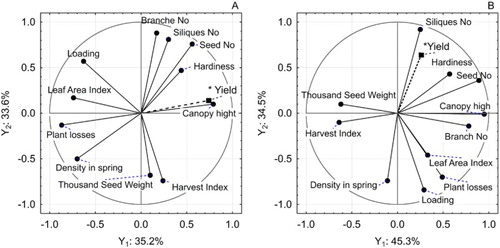
Figure 2. Classification of winter oil seed rape cultivars according to the principle components at standard (A) and high-input (B) technology.
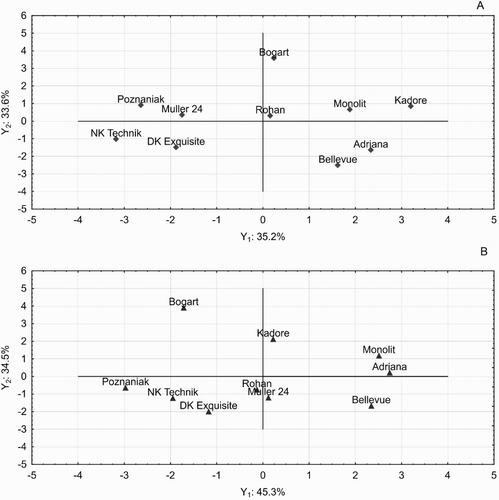
According to Weymann et al. (Citation2015), in patterning of the rapeseed yield are the compensation capacities of the structure elements that give opposite directions in relation to the yield. This is the relationship between the TSW and the number of siliques / seeds that signs are opposite. In addition, the different pattern of components and location of traits was discovered for high-input technology. In A2 the following PCA equations were calculated: Y1 = −0.11 x1 + 0.57 x2 + 0.49 x3 + 0.95 x4 + 0.29 x5 + 0.78 x6 + 0.25 x7 + 0.33 x8 − 0.62 x9 − 0.64 x10 + 0.89 x11 + 0.26 x12. For this component, the share of the explained variance is 45.33%. Y2 = −0.74 x1 + 0.43 x2 − 0.70 x3 + 0.01 x4 − 0.84 x5 − 0.14 x6 − 0.92 x7 − 0.46 x8 + 0.10 x9 − 0.10 x10 + 0.36 x11 + 0.64 x12. This component is responsible for 34.5% of the total variance. According to the criterion of maximising the explained variance for the conditions of technology A2, the model of the main components has the greatest credibility, because it explains a total of 79.83% of the total variance. In high-input technology, the 1st component is responsible for the morphological traits of plant and canopy, positively explaining the role of the branches per plant and canopy height as well as the seed filling per silique ((B)). The TSW and HI are on opposite side of this component. The classification of winter rape cultivars displayed that 1st component is the important for Adriana, Monolit and Bellevue cultivars. On the contrary, for hybrids Poznaniak, NK Technik and for the population Bogart the importance of this features turned out to be negative ((B)). The 2nd component according to the model generates the effect of traits such as: spring density, plant losses and loading in significantly negative way, while the number of siliques in positive and very important close to the yield. Therefore, one should look at the significance of these traits for yielding in high-input technology. Taking into account the direction of loads on the Y2 component, the yield is mostly connected to the hardiness, siliques per plant and seed number in silique. The most balanced effect of yield elements was displayed in case of Rohan cv. And in high-input technology also at Müller 24 cv. ((B)).
Our results confirmed the reports regarding the intensification of nitrogen fertilisation and the increase in seed yield. High-input technology (A2) assumed here based on the additional 40 kg N /ha, with total 211 N kg/ha (split on 18 kg pre-sown and 193 kg top fertilisation) – (). The average seed yield in 2014 was 13% higher in A2 technology than in A1, and in 2015 by 9.2% higher. Overall the 3 year cycle, the yield increase in technology A2 was 9.6% in relation to A1. Significantly higher yield was obtained for hybrids (56.3 dt/ha), especially DK Exquisite (58.0 dt/ha), except the population cultivar Bellevue, which obtained a record yield 58.1 dt/ha. On the background of the tested cultivars, the highest in plus of yield in favour of A2 technology showed hybrids (from + 3.1% for Rohan to 7.1% for DK Exquisite), whereas some population cultivars displayed the greatest deviations in minus in the yield with technology standard (−10.6% Kadore to 0.6% Bellevue) – (). The impact of mineral N fertilisation on oilseed rape yielding is described by many authors. Nitrogen fertilisation generates high costs in oilseed rape cropping, thus it optimisation is significant (Poreta et al. Citation2016). Fertilisation in a dose of 150–180 kg N/ha improved the yield of winter rape seeds (Schuster and Rathke Citation2001; Barłóg and Grzebisz Citation2004a, Citation2004b). In high input systems, higher doses of nitrogen fertilisation in the amount of 200 kg/ha (Zhao et al. Citation1993), 240–250 kg/ha (Wojanowska et al. Citation1993; Yusuf and Bullock Citation1993) and even 300 kg N /ha (Shepherd and Sylvester-Bradley Citation1996) also effectively increased yields. Moreover, according to Jankowski et al. (Citation2016) increasing the nitrogen dose to 240 kg N /ha might not improve the HI, thus the authors suggest that above 180 kg N /ha the vegetative mass improves more than seeds. The share of yield elements for shaping the winter rape seed yield in the context of nitrogen fertilisation was investigated by Li et al. (Citation2016). The results of these authors suggest that the pre-sowing fertilisation N was the most effective at a dose not exceeding 135 kg N /ha, along with 67.5–90 kg /per ha at the beginning of stem elongation. Although higher doses of nitrogen fertilisation increased the number of branches and the number of siliques, they did not change the number of seeds and the weight of 1000 seeds. According to Dresbøll et al. (Citation2016), a twice higher rape seed yield was achieved at a dose of 280 kg N /ha in the case of a hybrid type of low biomass, i.e. 69 dt/ha, than at a dose of 120 kg N /ha (i.e. 29 dt/ha). Nitrogen nutrition of winter rape plants at a dose of 280 kg resulted in a higher vitality of rape leaves, whereas at the dose of 120 N /ha there were no leaves on the stalk at the end of June. The drying of winter rape leaves occurs earlier in conditions of low nitrogen fertilisation (Svecnjak and Rengel Citation2006).
In standard technology population cultivars of winter oilseed rape formed the yield in relation to the height of canopy, while typical yield elements such as number of branches, the number of siliques and the number of seeds constituted as the second role. In our high-input technology, apart from of extra N, also amino acids, micronutrients and additional fungicidal treatment were provided to the practice. We can conclude that synergetic impact could have been included in the overall assessment of rapeseed production. The high-input technology improved the yield responded elements e.g. number of siliques and number of seeds per silique as well as winter hardiness. This impact was more evident for population cultivars than for hybrids. We state that high-input technology, that provided 9.6% increase in seed yield, scheduled as in this report is biologically justified for winter oilseed rape in North and central European countries.
Disclosure statement
No potential conflict of interest was reported by the authors.
Note on Contributors
Wenda-Piesik Anna has completed her DSc at the age of 41 years from UTP University of Science and Technology. Currently, her possition is Associate Professor, at The Department of Agronomy, UTP in Bydgoszcz, Poland. She is the Head of The Laboratory of Experimental Methods. She published 80 original articles, the total IF is at 32,751, total number of citations is at 229, h- index 9 and has been serving as an editorial board member of repute.
Hoppe Szymon has completed his PhD in Agronomy at the age of 35 years from UTP University of Science and Technology. He is Product Manager Seed at Polish Agro Company.
References
- Avice JC, Etienne P. 2014. Leaf senescence and nitrogen remobilization efficiency in oilseed rape (Brassica napus L.). J Exp Bot. 65:3813–3824. doi: 10.1093/jxb/eru177
- Balodis O, Gaile Z, et al. 2009. Influence of agroecological factors on winter oilseed rape (Brassica napus L.) autumn growth. In: Gaile Z., Ciproviča I., Kaķītis A., editor. Research for rural development – 2009. International scientific conference proceedings. Jelgava: LLU; p. 36–43.
- Barbetti MJ, Banga SS, Salisbury PA. 2012. Challenges for crop production and management from pathogen biodiversity and diseases under current and future climate scenarios – case study with oilseed Brassicas. Field Crop Res. 127:225–240. doi: 10.1016/j.fcr.2011.11.021
- Barłóg P, Grzebisz W. 2004a. Effect of timing and nitrogen fertilizer application on winter oilseed rape (Brassica napus L.). II. Nitrogen uptake dynamics and fertilizer efficiency. J Agron Crop Sci. 190:314–323. doi: 10.1111/j.1439-037X.2004.00109.x
- Barłóg P, Grzebisz W. 2004b. Effect of timing and nitrogen fertilizer application on winter oilseed rape (Brassica napus L.). I. Growth dynamics and seed yield. J Agron Crop Sci. 190:305–313. doi: 10.1111/j.1439-037X.2004.00108.x
- Bouchet AS, Laperche A, Bissuel-Belaygue C, Baron C, Morice J, Rousseau-Gueutin M, Dheu JE, George P, Pinochet X, Foubert T, et al. 2016. Genetic basis of nitrogen use efficiency and yield stability across environments in winter rapeseed. BMC Genet. 17:131. doi: 10.1186/s12863-016-0432-z
- Berry PM, Spink J, Foulkes MJ, White PJ. 2010. The physiological basis of genotypic differences in nitrogen use efficiency in oilseed rape (Brassica napus L.). Field Crop Res. 119:365–373. doi: 10.1016/j.fcr.2010.08.004
- Bonhomme R. 2000. Bases and limits to using ‘degree.day’ units. Eur J Agron. 13:1–10. doi: 10.1016/S1161-0301(00)00058-7
- Chay P, Thurling N. 1989. Variation in pod length in spring rape (Brassica napus) and its effect on seed yield and yield components. The Journal of Agricultural Science. 113:139–147. doi: 10.1017/S002185960008669X
- Crnobarac J, Marinković B, Jeromela-Marjanović A, Balalić I, Jaćimović G, Latković D. 2015. The effect of variety and sowing date on oilseed rape yield and quality. Agr Food Sci. 3:241–245.
- Dejoux JF, Recous S, Meynard JM, Trinsoutrot I, Leterme P. 2000. The fate of nitrogen from winter-frozen rapeseed leaves: mineralization, fluxes to the environment and uptake by rapeseed crop in spring. Plant Soil. 218/2:257–272. doi: 10.1023/A:1014934924819
- Diepenbrock W. 2000. Yield analysis of winter oilseed rape (Brassica napus L.): a review. Field Crop Res. 67:35–49. doi: 10.1016/S0378-4290(00)00082-4
- Diepenbrock W, Grosse F. 1995. Rapeseed (Brassica napus L.) physiology. [In:] Physiological potential for yield improvement of annual oil and protein crops. Adv Plant Breeding. 17:21–53.
- Dresbøll DB, Rasmussen IS, Thorup-Kristensen K. 2016. The significance of litter loss and root growth on nitrogen efficiency in normal and semi-dwarf winter oilseed rape genotypes. Field Crop Res. 186:166–178. doi: 10.1016/j.fcr.2015.12.003
- Garnett T, Conn V, Kaiser BN. 2009. Root based approaches to improving nitrogen use efficiency in plants. Plant Cell Environ. 32:1272–1283. doi: 10.1111/j.1365-3040.2009.02011.x
- Geisler G, Henning K. 1981. Untersuchungen zur Ertragsstruktur von Raps (Brassica napus L. var. napus). II. Die generative Entwicklung der Rapspfanze in Abhangigkeit von der Bestandesdichte. Bayer. Landwirthschaftliche Jahrbücher. 58:322–332.
- Grosse F, Leon J, Diepenbrock W. 1992. Ertragsbildung und ertragsstruktur bei winterraps (Brassica napus L.) I. genotypische variabilität. J Agron Crop Sci. 169:70–93. doi: 10.1111/j.1439-037X.1992.tb01186.x
- He H, Yang R, Li Y, Ma A, Cao L, Wu X, Chen B, Tian H, Gao J. 2017. Genotypic variation in nitrogen utilization efficiency of oilseed rape (Brassica napus) under contrasting N supply in Pot and field experiments. Front Plant Sci. 8:1358. doi: 10.3389/fpls.2017.01358
- Horst WJ, Behrens T, Heuberger H, Kamh M, Reidenbach G, Wiesler F. 2002. Genotypic differences in nitrogen use-efficiency in crop plants. In: Lynch J.M., Schepers J.S., Ünver I., editor. Innovative soil-plant systems for sustainable agricultural practices, OECD work-shop 2002. Paris: OECD Publications Paris; p. 75–92.
- Jankowski K, Budzyński W. 2007. Response of different breeding forms of winter oilseed rape to date and density of sowing II. seed yield and yield components. Oilseed Crops. XXVIII:195–207. (Reakcja różnych form hodowlanych rzepaku ozimego na termin i gęstość siewu II. Plon nasion i jego składowe).
- Jankowski K, Budzyński WS, Załuski D, Hulanicki PS, Dubis B. 2016. Using fractional factorial design to evaluate the effect of the intensity of agronomic practices on the yield of different winter oilseed rape morphotypes. Field Crop Res. 188:50–61. doi: 10.1016/j.fcr.2016.01.007
- Jaradat AA. 2016. Breeding oilseed crops for sustainable production. Breeding Oilseed Crops for Sustainable Production. 18:421–472. doi: 10.1016/B978-0-12-801309-0.00018-5
- Jenkins PD, Leitch MH. 1986. Effects of sowing date on the growth and yield of winter oil-seed rape (Brassica napus). J Agr Sci–Cambridge. 107:405–420. doi: 10.1017/S0021859600087219
- Jiang C, Shi J, Li R, Long Y, Wang H, Li D, Zhao J, Meng J. 2014. Quantitative trait loci that control the oil content variation of rapeseed (Brassica napus L.). Theor Appl Genet. 127:957–968. doi: 10.1007/s00122-014-2271-5
- Kaczmarek J, Kotecki A, Kotowicz L, Weber R. 2003. Genotype-environmental interaction of yielding winter rape cultivars in PDO experiments. Bulletin of Plant Breeding and Acclimatization Institute. 226/227:395–403. (Interakcja genotypowo-środowiskowa plonowania odmian rzepaku ozimego w doświadczeniach PDO).
- Kessel B, Schierholt A, Becker HC. 2012. Nitrogen use efficiency in a genetically diverse set of winter oilseed rape (Brassica napus L.). Crop Sci. 52:2546–2554. doi: 10.2135/cropsci2012.02.0134
- Lancashire PD, Bleiholder H, Langelüddecke P, Stauss R, Van Den Boom T, Weber E, Witzenberger A. 1991. A uniform decimal code for growth stages of crops and weeds. Ann Appl Biol. 119:561–601. doi: 10.1111/j.1744-7348.1991.tb04895.x
- Leach JE, Stevenson HJ, Rainbow AJ, Mullen LA. 1999. Effects of high plant population on growth of winter oilseed rape (Brassica napus). J Agr Sci. 132:173–180. doi: 10.1017/S0021859698006091
- Li X, Li Q, Yanga T, Nie Z, Chena G, Hu L. 2016. Responses of plant development, biomass and seed production of direct sown oilseed rape (Brassica napus) to nitrogen application at different stages in Yangtze river basin. Field Crop Res. 194:12–20. doi: 10.1016/j.fcr.2016.04.024
- Malagoli P, Laine P, Rossato L, Ourry A. 2005. Dynamics of nitrogen uptake and mobilization in field-grown winter oilseed rape (Brassica napus) from stem extension to harvest-I. Global N flows between vegetative and reproductive tissues in relation to leaf fall and their residual N. Ann Bot. 95:853–861. doi: 10.1093/aob/mci091
- Masclaux-Daubresse C, Daniel-Vedele F, Dechorgnat F, Chardon F, Gaufichon L, Suzuki A. 2010. Nitrogen uptake, assimilation and remobilization in plants: challenges for sustainable and productive agriculture. Ann Bot. 105:1141–1157. doi: 10.1093/aob/mcq028
- Muśnicki C, Gładysiak S. 2008. Periodic evaluation of applied agrotechnic level and the condition of plants on selected rapeseed plantations in Wielkopolska. Oilseed Crops. XXIX:175–184. (Okresowa ocena poziomu stosowanej agrotechniki i stanu roślin na wybranych plantacjach rzepaku w Wielkopolsce).
- Ozolincius R, Stakenas V, Serafinaviciute B. 2005. Meteorological factors and air pollution in Lithuanian forests: possible effects on tree condition. Environ Pollut. 137:587–595. doi: 10.1016/j.envpol.2005.01.044
- Poreta M, Chandrasekar B, van der Hoornd Renier AL, Avice J-Ch. 2016. Characterization of senescence-associated protease activities involved in the efficient protein remobilization during leaf senescence of winter oilseed rape. Plant Sci. 246:139–153. doi: 10.1016/j.plantsci.2016.02.011
- Povilaitis V, Lazauskas S. 2010. Winter wheat productivity in relation to water availability and growing intensity. Žemdirbystė-Agriculture,. 97:59–68.
- Rathke GW, Behrens T, Diepenbrock W. 2006. Integrated nitrogen management strategies to improve seed yield, oil content and nitrogen efficiency of winter oilseed rape (Brassica napus L.): A review. Agr Ecosyst Environ. 117:80–108. doi: 10.1016/j.agee.2006.04.006
- Rummukainen M. 2010. Climate outlook for the baltic sea region. Nordic association of agricultural scientist. NJF Report. 6:12–13.
- Salisbury PA, Barbetti MJ. 2011. Breeding oilseed brassica for climate change. In: Yadav SS, Redden R.J., Hatfield J.S., Lotze-Campen H., Hall A., editor. Crop adaptation to climate change. Chichester, West Sussex, UK: John Wiley & Sons, Ltd; p. 448–463.
- Schuster C, Rathke G-W. 2001. Nitrogen fertilization of transgenic winter oilseed rape. In: Horst W.J, editor. Plant nutrition: food security and sustainability of agro-ecosystems through basic and applied research. Dordrecht: Kluwer Academic Publisher; p. 336–337.
- Shepherd MA, Sylvester-Bradley R. 1996. Effect of nitrogen applied to winter oilseed rape (Brassica napus L.) on soil mineral nitrogen after harvest and on the response of a succeeding crop of winter wheat to nitrogen fertilizer. J Agr Sci–Cambridge. 126:63–74. doi: 10.1017/S002185960008881X
- Sieling K, Kage H. 2008. The potential of semi-dwarf oilseed rape genotype. A review. Agron Sustain Dev. 30:271–279. doi: 10.1051/agro/2009036
- Sierts HP, Geisler G, Leon J, Diepenbrock W. 1987. Stability of yield components from winter oil-seed rape (brassica napus L.). J Agr Crop Sci. 158:107–113. doi: 10.1111/j.1439-037X.1987.tb01153.x
- Sova AV, Vašák J, Soukup J. 1998. Variants of winter rape cultivation technology. Oilseed Crops. XIX:105–112. (Warianty technologii uprawy rzepaku ozimego (Brassica napus L. var. napus)).
- Svecnjak Z, Rengel Z. 2006. Canola cultivars differ in nitrogen utilization efficiency at vegetative stage. Field Crop Res. 97:221–226. doi: 10.1016/j.fcr.2005.10.001
- Ulas A, Schulte A, Erley G, Kamh M, Wiesler F, Horst WJ. 2012. Root-growth characteristics contributing to genotypic variation in nitrogen efficiency of oilseed rape. J Plant Nut Soil Sc. 175:489–498. doi: 10.1002/jpln.201100301
- Velička R, Marcinkeviciene A, Raudonis S, Rimkeviciene M. 2006. Integrated evaluation of rape readiness for overwintering. Acta Agr Scand B-S P. 56:110–116.
- Wang C, Hai J, Yang J, Tian J, Chen W, Chen T, Luo H, Wang H. 2016. Influence of leaf and silique photosynthesis on seeds yield and seeds oil quality of oilseed rape (Brassica napus L.). Eur J Agron. 74:112–118. doi: 10.1016/j.eja.2015.12.008
- Wenda-Piesik A, Kazek M, Ropińska P. 2017. Impact of amino acid biostimulation and microelements fertilization in foliar application on productivity of winter oilseed rape. Fragm Agronom. 34:119–129. (Wpływ aminokwasowego biostymulatora i nawozu mikroelementowego stosowanych nalistnie na produktywność rzepaku ozimego).
- Weymann W, Böttcher U, Sieling K, Kage H. 2015. Effects of weather conditions during different growth phases on yield formation of winter oilseed rape. Field Crop Res. 173:41–48. doi: 10.1016/j.fcr.2015.01.002
- Wielebski F. 2009. Response of different types of winter oilseed rape varieties to crop production systems. I. characteristics of ripening plants of oilseed rape and components of seed yield. Oilseed Crops. XXX:75–90. (Reakcja różnych typów hodowlanych odmian rzepaku ozimego na poziom stosowanej agrotechniki. I. Charakterystyka dojrzewających roślin rzepaku oraz jego plonowanie i układ elementów plonotwórczych).
- Wojanowska T, Sienkiewicz S, Wojtas A. 1993. The influence of increasing nitrogen doses on the yield and chemical composition of winter rape seeds. Oilseed Crops. XVI(1):182–187. (Wpływ wzrastających dawek azotu na plon i skład chemiczny nasion rzepaku ozimego).
- Yusuf RI, Bullock DG. 1993. Effect of several production factors on two varieties of rapeseed in the United States. J Plant Nut. 16:1279–1288. doi: 10.1080/01904169309364612
- Zhao FJ, Evans EJ, Bilsborrow PE, Syers JK. 1993. Influence of sulphur and nitrogen on seed yield and quality of low glucosinolate oilseed rape (Brassica napus L). J Sci Food Agr. 63:29–37. doi: 10.1002/jsfa.2740630106