
ABSTRACT
In modern agriculture, plant-derived substances referred to as plant growth and development regulators or biostimulators are increasingly used. The aim of the research was the influence of the applied biostimulators on the concentration of glucosinolates in seeds of three morphotypes of winter oilseed rape. The research was carried out in three growing seasons at the University of Natural Sciences and Humanities in Siedlce (52o03'N i 22o33'E), Poland. The examined factors were: morphotypes of winter oilseed rape: hybrid restored with a traditional type of growth (Konkret), population (Poznaniak), hybrid restored with a semi-dwarf type of growth (PX104), II-four types of biostimulators: 1. Asahi®SL, 2. Silvit®, 3. Tytanit®, 4. control variant. The applied biostimulators had a positive effect on the feed value of the seeds in terms of glucosinolates. In all varieties and years of research, the lowest concentration of these compounds was recorded after the application of the Silvit biostimulator, while in the first year of research after the application of Silvit and in the second year after the application of Tytanit and Asahi SL biostimulators, the same content of these compounds was obtained.
Introduction
Glucosinolates (GSL) are anionic organic compounds classified as secondary plant metabolites, which have a β-D-glucose molecule, sulfonated oxime and a side chain derived from indole (tryptophan), aliphatic (alanine, valine, leucine and methionine) or aromatic (phenylalanine, tyrosine) amino acids (Clarke Citation2010).
Glucosinolates are found in plant species belonging to the Brassicaceae family, Capparaceae family and Caricaceae family. In plants, these compounds exist in the glycoside form, showing no toxicity to animals, plants and pathogens.
Koritsas et al. (Citation1991) emphasises that the concentration of glucosinolates is influenced by environmental factors and signalling molecules related to biotic and abiotic stress responses, and Sang et al. (Citation1984) consider the genotype, climate and growing conditions, including fertilisation and harvest time.
Obtaining high and good quality crops in plant production is primarily the ability to prevent or reduce the occurrence of biotic and abiotic stresses and regenerate the damage they cause. In modern agriculture, plant-derived substances referred to as plant growth and development regulators or biostimulators are increasingly used.
The research hypothesis was adopted in the study that the use of plant growth and development regulators may improve the quality of seeds by reducing the concentration of harmful sulphur compounds called glucosinolates. The aim of the research was to determine the effect of the applied biostimulators on the concentration of glucosinolates in seeds of three morphotypes (population, restored hybrid with traditional and semi-dwarf growth) of winter oilseed rape.
Material and methods
The field experiment was caried out in three growing seasons (2013-2016) at the Agricultural Experimental Station – Zawady (52o03'N and 22o33'E) belonging to the University of Natural Sciences and Humanities in Siedlce, Poland. The experiment was set up in a split-split-plot configuration with three replications. The area of one plot for harvesting was 18 m2. The examined factors were: morphotypes of winter oilseed rape: hybrid restored with a traditional type of growth (Konkret), population (Poznaniak), hybrid restored with a semi-dwarf type of growth (PX104), II – four types of biostimulators:
biostimulator Asahi®SL (sodium ortho-nitrophenol, sodium para-nitrophenol, sodium 5-nitroguaiacol), used in the fall in the 3–5 leaf stage (BBCH 13-15), in spring after the start of vegetation (BBCH 28-30) and two weeks after the second treatment, in doses of 0.60 L/ha,
biostimulator Silvit® (active silicon, potassium oxide, boron – form of pure element, zinc – form of pure element), applied 3 weeks after emergence (BBCH 12-14), in spring after the start of vegetation (BBCH 28-30) and two weeks after the second treatment, at doses of 0.20 L/ha.
biostimulator Tytanit® (titan), used in the fall in the 4–8 leaf stage (BBCH 14-18), in spring after the start of vegetation: beginning of development of side shoots (BBCH 21-36) and in the stage of flower bud development – beginning of flowering (BBCH 50-61), at doses of 0.20 L/ha
control variant – without the use of biostimulators.
The experiment was carried out on soil classified according to WRB FAO (Citation2014) to the group of the Haplic Luvisol, sanded soil, belonging to the very good rye soil complex, valuation class IVb. In the years of the experience, the soil reaction (pH) ranged from 5.68–5.75. The soil was characterised by a low total nitrogen content (average from 0.80–0.90 g/kg), phosphorous content (average from 0.33–0.55 g/kg), potassium content (average from 0.61–0.67 g/kg) and calcium content (average from 0.82–0.85 g/kg) and the average magnesium content (from 0.38–0.46 g/kg) and sulphur content (from 0.11–0.15 g/kg). It has a low abundance in assimilable forms of phosphorous (average from 75.0–80.0 mg/kg) and the average assimilability of potassium (from 200.0–205.0 mg/kg) and magnesium (average from 59.0–61.0 mg/kg).
The phosphorous-potassium fertilisation at the dose of 40.0 kg P/ha and 110.0 kg K/ha with the first dose of 40.0 kg N/ha was used before sowing. Fertilisation was used in the form of Lubofos for rape at the dose of 600.0 kg, i.e. 21.0 kg N/ha, 26.4 kg P/ha, 92.1 kg K/ha, 34.8 kg S/ha, 1.2 kg B/ha. Fertilisation rates were supplemented by 55.9 kg/ha of ammonium nitrate (19.0 kg N/ha), 29.6 kg/ha of triple superphosphate (13.6 kg P/ha) and 29 kg/ha of potassium salt (17.9 kg K/ha). The second nitrogen dose of 100.0 kg/ha was applied in spring before vegetation (BBCH 28-30) using ammonium nitrate at the dose of 255.5 kg/ha (86.9 kg N/ha) and ammonium sulphate at the dose of 62.5 kg/ha (13.1 kg N/ha + 15.0 kg S/ha). The third dose of nitrogen 60.0 kg/ha was applied at the beginning of budding (BBCH 50) using ammonium nitrate at the dose of 176.5 kg/ha (60.0 kg N/ha).
Rape was collected in two stages, in the first and second decade of July: I year: 11.07.2014; II year: 17.07.2015; III year: 14.07.2016.
Winter oilseed rape seed samples were analysed by chemical analysis for the content of glucosinolates (μmol/g) in the dry matter of seeds, which was determined using high pressure liquid chromatography (HPLC) with reversed phase columns and gradient elution according to PN-EN ISO 9167-1.
Research results were statistically analysed by ANOVA. The results of the study were statistically analysed using the analysis of variance. The significance of the sources of variation was tested by the Fischer-Snedecor ‘F’ test, and the assessment of significance of differences at the significance level p <0.05 between the compared averages used Tukey’s multiple intervals. Statistical calculations were made based on our own algorithm written in Excel.
In the years of the experiment, various climatic conditions prevailed (). In the 2014–2015 growing season, the highest annual rainfall was recorded – on average 599.2 mm and the lowest annual air temperature – on average 8.8°C. The 2015–2016 growing season was the warmest and driest. The annual rainfall was 43.8 mm lower than the long-term average, and the average annual air temperature was 1.3°C higher than the average for 1996–2010.
Table 1. Characteristics of weather conditions in the years 2013–2016 (Poland).
Results and discussion
Glucosinolate content (µmol/g) in seeds of the studied cultivars depending on the biostimulators used
On the basis of the conducted own research, it was shown that the bioregulators used reduced the concentration of harmful sulphur compounds in the rape seeds of the cultivars studied, as shown in . On the basis of the conducted own research, after the application of the Silvit biostimulator, the best feed value of seeds was obtained in all morphotypes. After the use of Asahi SL and Tytanit in the population morphotype, the differences in the value of this feature (on average 7.99 µmol/g) were statistically insignificant (). A similar trend was observed in the Konkret (on average 11.08 µmol/g). This is consistent with the results of Sikorska et al. (Citation2018). The authors also obtained the lowest concentration of these compounds in seeds collected from plots sprayed with Silvit biostimulators (on average 8.88 µmol/g), then Tytanit – on average 9.08 µmol/g and Asahi SL – on average 9.65 µmol/g.
Figure 1. Glucosinolate content (GSL) (µmol/g) (interraction cultivars and types of biostimulators used).
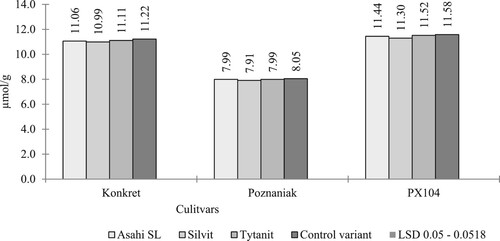
Table 2. Glucosinolate content (GSL) (µmol/g) depending on factors of experience text values marked with the same letter do not differ significantly at α = 0.05 under table.
The beneficial effect of plant growth and development regulators on the content of glucosinolates in rapeseed was demonstrated by Ullah et al. (Citation2012), while Gugała et al. (Citation2020) found an increase in the concentration of harmful sulphur compounds (GSL) after the use of a biostimulator containing free amino acids. Similar results were obtained by Jankowski et al. (Citation2019) after foliar application of macronutrient and micronutrient fertilisers in the fall.
Glucosinolate content (µmol/g) depending on the cultivars used
Own research showed that glucosinolate content depended on the genetic factor (). The seeds of the cultivars studied had an average of 7.98–11.46 µmol/g of these compounds, while El-Beltagi and Mohamed (Citation2010) obtained an average of 4.40–5.95 µmol/g, Broniarz and Stroiwąs (Citation2013) on average from 7.1–16.4 μmol/g, Jankowski et al. (Citation2015) on average 15.25 μmol/g, Sikorska et al. (Citation2018) on average from 8.84–9.84 μmol/g d.m, while in the studies by Bocianowski et al. (Citation2020) the average content in the studied genotypes ranged from 4.13 (for PN66 × PN21) to 8.53 µmol/g (Californium).
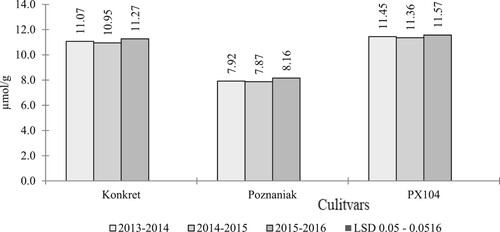
In the authors’ own research the lowest concentration of these compounds was found in the population cultivar Poznaniak, a significantly higher content, on average by 3.12 µmol/g, in the hybrid morphotype restored with a traditional type of growth (Konkret), and the highest half-dwarf hybrid (PX104) (). Similar research results were obtained by Wójtowicz and Jajor (Citation2006), Sikorska et al. (Citation2018) and Gugała et al. (Citation2020). In turn, Ratajczak et al. (Citation2017) found that the seeds of the semi-dwarf hybrid PR45D03 were characterised by a significantly lower content of glucosinolates compared to the hybrid with the traditional type of growth (PR46W31) and the population variety (Californium).
Glucosinolate content (µmol/g) depending on climatic conditions
The own research showed the influence of climatic conditions in the years of the research on the content of glucosinolates in rape seeds (, 2). In the tested morphotypes, the lowest concentration of these compounds was found in the wettest (k=1.71) year of the study, and the highest in the driest and warmest year of the study (). Bouchereau et al. (Citation1996), Ullah et al. (Citation2012), Lääniste et al. (Citation2016) and Sikorska et al. (Citation2018) who found that drought stress during flowering increased the concentration of these compounds.
Based on the authors’ own research, it was shown that in all growing seasons the seeds of the population cultivar Poznaniak had the lowest concentration of these compounds, while the highest was found in the semi-dwarf hybrid PX104 ().
Figure 2. Glucosinolate content (GSL) (µmol/g) (interraction years and types of biostimulators used).
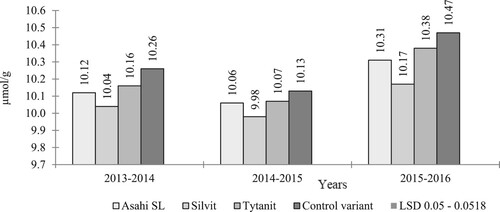
In all growing seasons, the applied biostimulators reduced the content of harmful sulphur compounds (). The most significant reduction in the value of this feature compared to the control variant was noted after the application of Silvit. In the first year of research, after the use of Asahi and Tytanit, the seeds of the studied varieties showed the same value of this feature. A similar tendency was noted in the second year of the study, and in the third year the differences between these biostimulators were small and amounted to an average of 0.07 µmol/g. Summing up, the applied biostimulators had a positive effect on the feed value of seeds in terms of glucosinolate (GSL) content. In all the tested cultivars and the years of research, the lowest concentration of these compounds was recorded after the application of the Silvit biostimulator, while in the first year of research after the application of Silvit and in the second year after the application of Tytanit and Asahi SL biostimulators, the same content of these compounds was obtained in rapeseed. Among the cultivars studies, regardless of the biostimulator used, the population morphotype was distinguished by the best seed feed value.
Figure 3. Glucosinolate content (GSL) (µmol/g) (interraction years and cultivars).
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
Notes on contributors
Anna Sikorska
Anna Sikorska is a doctor of agricultural sciences in agronomy discipline. Her research interests are biostimulants influence on arable crops and winter rape. She is co-author of over 50 scientific and popular science works.
Marek Gugała
Marek Gugała is a professor of agricultural sciences in the agronomy discipline. His research interests are potato agriculture and the influence of biostimulators on arable crops. He is author of over 400 scientific and popular science works and co-author of over 20 inventions, including patents.
Krystyna Zarzecka
Krystyna Zarzecka is a professor of agricultural sciences in the agronomy discipline. Zarzecka's research interest is potato agrotechnics
References
- Bocianowski J, Liersch A, Nowosad K. 2020. Genotype by environment interaction for alkenyl glucosinolates content in winter oilseed rape (Brassica napus L.) using additive main effects and multiplicative interaction model. Curr Plant Biol. 21:100137. doi:10.1016/j.cpb.2020.100137.
- Bouchereau A, Clossais-Besnard N, Bensaoud A, Leport L, Renard M. 1996. Water stress effects on rapeseed quality. Eur J Agron. 5:19–30. doi:10.1016/S1161-0301(96)02005-9.
- Broniarz J, Stroiwąs A. 2013. Rapeseed. In: J. Broniarz, A. Stroiwąs, editor. Descriptive list of agricultural plant varieties. (In Polish.). Słupia Wielka, Poland: Research Center for Cultivar Testing; p. 22–25.
- Clarke B. 2010. Glucosinolates, structures and analysis in food. Anal Methods. 2:310–325.
- El-Beltagi HEDS, Mohamed AA. 2010. Variations in fatty acid composition, glucosinolate profile and some phytochemical contents in selected oil seed rape (Brassica napus L.) cultivars. GRASASYACEITES. 61(2). ABRIL-JUNIO, 143-150, ISSN: 0017-3495. doi:10.3989/gya.087009.
- Gugała M, Sikorska A, Zarzecka K. 2020. The effect of fertilization with sulphur, boron, and amino acids on the content of glucosinolate in winter rape seeds. Agronomy. 10(4):1–9. DOI: 10.3390/agronomy10040519.
- Jankowski KJ, Budzyński WS, Kijewski Ł, Zając T. 2015. Biomass quality of Brassica oilseed crops in response to sulfur fertilization. Agron J. 107:1377–1391. DOI: 10.2134/agronj14.0386.
- Jankowski KJ, Sokólski M, Szatkowski A. 2019. The effect of autumn foliar fertilization on the yield and quality of winter oilseed rape seeds. Agronomy. 9(12):849. doi:10.3390/agronomy9120849.
- Koritsas VM, Lewis JA, Fenwick GR. 1991. Glucosinolate response of oilseed rape, mustard and kale to mechanical wounding and infestation by cabbage stem flea beetle (Psyliodes chysocephala). Ann Appl Biol. 118:209–221. DOI: 10.1111/j.1744-7348.1991.tb06099.x.
- Lääniste P, Eremeev V, Mäeorg E, Jõudu J. 2016. Effect of sowing date on oil, protein and glucosinolate concentration of winter oilseed rape (Brassica napus L. Agronomy Res. 14(2):1384–1395.
- Ratajczak K, Sulewska H, Szymańska G. 2017. New winter oilseed rape varieties – seed quality and morphological traits depending on sowing date and rate. Plant Prod Sci. 20(3):262–272. DOI: 10.1080/1343943X.2017.1304809.
- Sang JP, Minchinton IR, Johnstone PK, Truscott RJW. 1984. Glucosinolate profiles in the seed, root and leaf tissue of cabbage, mustard, rapeseed, radish and Swede. Can J Plant Sci. 64:77–93. DOI: 10.4141/cjps84-011.
- Sikorska A, Gugała M, Zarzecka K, Kapela K. 2018. The effect of biostimulants on the glucosinolate content in winter oilseed rape (Brassica napus L.) seeds. Plant Soil Environ. 64:7–12. doi:10.17221/642/2017-PSE.
- Skowera B. 2014. Changes of hydrothermal conditions in the Polish area (1971−2010). Fragmenta Agronomica. 31(2):74–87. (in Polish).
- Ullah F, Bano A, Nosheen A. 2012. Effects of plant growth regulators on growth and oil quality of canola (Brassica napus L.) under drought stress. Pak J Bot. 44(6):1873–1880.
- Wójtowicz M, Jajor E. 2006. Effect of nitrogen fertilization on chemical composition of seeds of five winter oilseed rape cultivars. Rośliny Oleiste - Oilseed Crops. XXVII(1):31–43.
- World reference base for soil resources. 2014. International soil classification system for naming soils and creating legends for soil. FAO. World Soil Resources Reports, 106. Rome. Field experiment, http://www.fao.org.