
ABSTRACT
Surface-dwelling beetles offer ecosystem services in agro-ecosystems including biological pest control and organic matter decomposition. This study was conducted to investigate beetle community response to six years of tillage, crop residue application, crop rotation and soil fertility amendments (low fertiliser, high fertiliser, manure, manure + low fertiliser and manure + 60 kg N ha−1) treatments. Reduced tillage increased beetle abundance by 4.6% relative to soil inversion tillage. Crop residue application increased the abundances of Platynus assimilis, Zophosis boei and total beetles by two to three times. Meanwhile, Renatiella reticulata increased by 628.6% in high fertiliser compared to manure + 60 kg N ha−1, manure + low fertiliser and the control. In crop rotation + crop residue retention plots, application of sole manure at 10 t ha−1 increased the Shannon index by 360–365% relative to manure + 60 kg N ha−1 and manure + low fertiliser. Based on the findings in this study, it can be concluded that reduced tillage, retention of crop residues on the soil surface and application of manure increase surface-dwelling beetle abundance and diversity.
Introduction
Farming systems that combine no-tillage, plant residue retention on the soil surface and legume-based crop rotation have the potential to conserve biodiversity and therefore enhance beneficial ecosystem services such as biological pest control and nutrient cycling. Conservation agriculture (CA), a technology that has been widely promoted in the world including sub-Saharan Africa (SSA), has minimum mechanical soil disturbance, permanent plant residue retention and diversity through crop rotation as its key tenets (Jat et al. Citation2012). Meanwhile, conventional farming systems such as soil inversion tillage, high mineral fertiliser application, frequent weeding and plant residue removal contribute greatly to loss in soil biodiversity and the associated services (Liebman and Gallant Citation1997).
Macro-arthropods, such as surface-dwelling beetles, are found across most farmland habitats and contribute greatly to soil biodiversity, making them an integral part of agricultural ecosystems (Diekotter et al. Citation2010; Holopainen and Helenius Citation1992). Although some of their herbivorous species are serious agricultural pests, the majority offer some key beneficial services that can be summed up into biological pest control and nutrient cycling. Their role in regulating ecological processes in agroecosystems, particularly in the temperate regions such as Europe, the United States of America and Australia, has been widely documented (Stinner and House Citation1990; Kromp Citation1999; Soane et al. Citation2012).
Macro-arthropods function on two of the broad levels of organisation of the soil food web (Lavelle et al. Citation1995). Firstly, they act as litter transformers through humidifying ingested plant debris, improving its quality as a substrate for microbial decomposition and fostering the growth and dispersal of microbial populations. Secondly, they also serve as ecosystem engineers that physically modify the habitat directly or indirectly, regulating the availability of resources to other species (Jones et al. Citation1994; Johnson et al. Citation2016). In the soil, this entails altering soil structure, organic matter decomposition and hydrology. In stress conditions, some ecosystems engineers such as dung beetles, Bubas bison, have been found to enhance soil water retention and increase plant growth by magnitudes of up to 10 and 280%, respectively (Johnson et al. Citation2016). Beetles often feed on plants and fungi as well as animal and plant debris which in turn improves soil fertility (Diekotter et al. Citation2010). Previous research has shown that beetles help to control weeds by foraging on weed seeds in the soil, reducing the need for herbicides in cropping systems (Labruyere et al. Citation2016; Jowett et al. Citation2020). Surface-dwelling beetles are among predators of many insects and can therefore be beneficial biological control agents for crop pests including dipterans, lepidopterans, aphids and tenebrionids (Kergunteuil et al. Citation2016). The biological control of pests using arthropods provides a potential alternative to chemical pesticide application and also serves as a sustainable option for long-term pest management (Boyetchko et al. Citation2009).
Mechanical soil disturbance through tillage alters micro-habitats of surface-dwelling beetles by changing the community structure of weeds, influencing the amount of plant litter as well as destroying the refuge and nesting sites (Stinner and House Citation1990). Tillage also affects beetle populations directly by causing mechanical injury and mortality, or indirectly, by modifying their habitat and altering prey availability(Luff Citation1987; Liebman and Gallant Citation1997). Tillage intensity can alter habitat structure and negatively affect activity-density, species richness and community composition of arthropods through provision of shelter and floral food resources (Holland and Luff Citation2000).
The retention of plant residues on the soil surface as practiced in CA creates a microhabitat having a balanced supply of moisture that supports soil life, conserves soil biodiversity and therefore enhances beneficial functional services in agroecosystems (Gill et al. Citation2011). Surface residues act as a food resource for some detritivorous beetle species and also provide suitable refuge as well as nesting sites for both predator and prey species. Meanwhile, the use of crop rotation provides temporal and spatial plant diversity, supporting a broad spectrum of herbivorous arthropods and therefore enhancing soil biodiversity (Steinmann and Dobers Citation2013).
It has been suggested that optimum soil fertility management should be practised in order to realise the full benefits of CA (Vanlauwe et al. Citation2014). This can be achieved through the addition of either organic or inorganic soil fertility amendments (SFA), or both, for improving physical and chemical properties of the soil as well as providing nutrients for crop growth (Adediran et al. Citation2005). Soil amendments may affect soil-dwelling organisms through bottom-up effects on the soil food web. Provision of adequate soil nutrients increases plant biomass production, providing food resources for herbivorous arthropods which are in turn available as prey for predators. In most developing countries such as those in SSA where CA has been widely promoted, most smallholder (SH) farmers are financially constrained and cannot afford the cost of inorganic fertilisers (Druilhe and Barreiro-Hurlé Citation2012). For those who own livestock, SH farmers usually apply animal manure either alone or in combination with some inorganic fertilisers, albeit in small quantities. The use of organic and inorganic SFA in SH farming systems has been shown to produce reasonable crop yields (Mugwira et al. Citation2002; Sileshi et al. Citation2019). However, the interactive role of CA principles and SFA in regulating surface beetle abundance and diversity as well as cropping system performance in SSA has received little attention. The objective of this study was to determine the effects of six years of repeated application of the three CA principles and SFA on surface-dwelling beetle communities and maize grain yield.
Materials and methods
Site description
Data for the study were collected from a six-year-old CA-based field experiment during the 2018/2019 cropping season at Hunyani farm owned by Chinhoyi University of Technology (17°20′S, 30°14′E), 110 km north-west of the capital city of Zimbabwe, Harare. Hunyani farm is located in a subtropical environment with an altitude of 1140 m. Rainfall is unimodal, received between October and May and ranging between 800 and 1000 mm per annum. Mean maximum temperatures are 27 and 15°C during summer and winter, respectively. The soils are Chromic Luvisol (WRB Citation2014) with a pH (1M CaCl2) of 6.0, and high silt and fine sand content making it prone to surface capping (Kodzwa et al. Citation2020). The major crops that are grown by farmers in the area are maize (Zea mays L.), soyabean (Glycine max L.) and tobacco (Nicitiana tabacum L.) during summer and spring wheat (Triticum aestivum L.) in winter.
Experimental design and treatments
The field experiment was established in December 2013 and all treatments were repeatedly applied on the same plots during successive years. The experiment was arranged as a split-split plot design. Respectively, the main-, sub- and sub-subplot factors were: (1) tillage system (reduced tillage: RT, and conventional tillage: CT), (2) crop residue retention (with (R) and without (NR, bare plots) surface residue retention), and (3) crop rotation (with (ROT) and without (MONO) rotation). Soil fertility amendment (SFA) treatments (high fertiliser: HF, low fertiliser: LF, cattle manure: M, cattle manure + low fertiliser: MLF, cattle manure + high fertiliser: MHF, and, control with no SFA: NF) were randomly allocated among each sub-subplot (). The factors and treatments were replicated three times. Reduced tillage involved making planting basins using hand held hoes, each basin measuring about 15 × 15 × 15 cm, length × width × depth, respectively (Mazvimavi and Twomlow Citation2009). For CT plots, all of the aboveground plant biomass was removed before mouldboard ploughing and discing to simulate smallholder farming systems where the crop residues are used as livestock feed. Meanwhile, crop residues were completely removed from NR but were retained in R plots under RT. To have a fully randomised experiment, crop residues were applied soon after ploughing on R treatments of CT plots. A maize-soyabean cropping sequence was followed during successive agricultural seasons in ROT while continuous maize was grown in MONO plots. For SFA, a compound inorganic fertiliser (7% N, 14% P and 7% K) was applied before planting to supply all the P and K, and 30% N requirements of the crop in treatments that required inorganic fertiliser application. This was followed by top dressing with ammonium nitrate fertiliser (34.5% N) when the crop reached 4–6 weeks after crop emergence to supply the remaining 70% of the N requirements. In plots that required organic SFA treatments, well-rotted cattle manure was applied at a rate of 10 t ha−1 before planting. Further details of SFA treatments are shown in . Sub-subplots were 8 × 7 m length and width, respectively, and all measurements were taken from the 8 central rows after discarding two rows from either side of the sub-subplot and four plants from either side of each row.
Table 1. Fertiliser treatments that were used on a long-term experiment at Hunyani farm, Chinhoyi, Zimbabwe, from 2013/2014 to 2018/2019 cropping season.
Agronomic practices
Maize was planted at a spacing of 90 × 50 cm, inter- and intra-row, respectively. Four seeds were planted at each plant position and then thinned to 2 plants per position within the first week of crop emergence to achieve a target population of 44,444 plants ha−1. In ROT plots, during the soyabean cropping year, the crop was planted at a spacing of 45 cm (inter row) × 5 cm (intra-row) to achieve 444,444 plants ha−1. Since there were no known negative effects of chemical herbicides on surface-dwelling beetles, chemical weed control was used. Glyphosate was used as a post-emergence herbicide to kill weeds that emerged before planting. A pre-emergence herbicide, metolachlor, was used to control annual weeds. However, when weeds emerged during the cropping cycle, hand hoeing was used to destroy the weeds and keep the plots weed-free.
Data collection
Beetles were sampled using un-baited pitfall traps placed within each sub-sub-subplot plot at three stages of maize crop growth (tasseling: VT, silking: R1 and physiological maturity: R6). Pitfall trapping is a method that is cheap, simple and effective and is widely used in sampling surface dwelling arthropods although it has some drawbacks (Greenslade Citation1964; da Silva et al. Citation2008). The sampling was done between January and April 2019, exactly five years after the establishment of the experiment. The 2018/2019 cropping season signified the end of the medium-term CA-based experiment as the site was converted back to a conventional tillage system in the following year. Plastic jars with a volume of 1000 cm3 and a brim diameter of 13 cm were used as pitfall traps. One pitfall trap was set up at a random position within the four central rows of each sub-subplot, and buried so that the upper edge was level with the ground surface. Each trap was half-filled with a mixture of 20% alcohol and 80% water to collect and preserve fauna samples in the field. Pitfall traps were left in place to collect beetles for seven days, and then they were closed using plastic sheets for 14 days to rest them before the next seven-day sampling period. Resting and closing of the traps at the end of the seven-day sampling period was done to avoid continuous trapping of arthropods. During each seven-day sampling period, pitfall catches were collected from traps on the third and then seventh day by passing the trap contents through a 1 mm sieve. Levels of the alcohol: water solution in the pitfall traps were topped up to make sure that the traps were half full throughout the sampling period. The short time interval between emptying of pitfall catches was meant to reduce deterioration of the beetle specimens since the alcohol solution of 20% used in the traps was regarded to be weak. The beetle specimens were then stored in vials filled with 70% ethanol and 30% water waiting for further processing. Total beetle catches from each plot for a particular crop growth stage were pooled together to form one sample. Beetle species were identified using taxonomic keys (Triplehorn Citation1965; Lindroth Citation1968).
At the end of the season, maize grain was harvested after physiological maturity. The maize cobs from each plot were shelled and then grain was weighed, and a subsample taken for moisture determination. Maize grain yield was then expressed in kg ha−1 after correcting for the standard moisture content required for stored grain of 12.5% (Tandzi and Mutengwa Citation2020).
Data analysis
Shannon diversity index, Pielou’s evenness index and species richness were estimated for each plot using Paleontological Statistical package (PAST) version 3.14 (Hammer et al. Citation2001). Beetle abundance data was Log(x+2.5) transformed to achieve normality of data distribution and homogeneity. The general analysis of variance (ANOVA) procedure was performed using Genstat statistical package 10th edition (VSN-International Citation2011) to determine the effects of tillage system, plant residue application, crop rotation and SFA together with all their interactions on beetle abundance, species diversity, species richness and evenness as well as maize grain yield. For each crop growth stage, beetle species whose total abundances across all plots fell below 10 were not used for the ANOVA tests. However, all species were included in the estimation of diversity parameters, regardless of their abundance. Where significant differences were detected, mean separation was by ±SED (Standard error of differences, P ≤ .05).
Results
Surface-dwelling beetle community characterisation
A total of 728 beetle individuals from 43 species were collected throughout the sampling period. However, only 13 species had abundances that were 10 and above and were included in statistical tests to determine the effects of treatments on beetle abundance. Meanwhile, all the 43 beetle species were used in determination of beetle community diversity in the studied cropping system. Although the majority of the beetle species were observed during the VT stage, crop growth stage seemed to have profound effects on the temporal distribution of surface-dwelling beetle species. For example, Platynus assimilis and Copris lunaris were observed during the silking (R1), Renatiella reticulata and Graphipterus cordiger were found during the physiological maturity (R6) while Zophosis boei occurred during both the R1 and R6 stages ( and ).
Table 2. Mean (in parentheses) and Log(x + 2.5) transformed abundance of individual and total beetle species in response to two tillage systems during the VT, R1 and R6 growth stages of maize at Hunyani farm, Chinhoyi, Zimbabwe, during the 2018/2019 cropping season. Data was transformed using log10(x+2.5).
Table 3. Mean (in parentheses) and Log(x + 2.5) transformed abundance of individual and total beetle species in response to crop residue application during VT, R1 and R6 growth stages of maize at Hunyani farm, Chinhoyi, Zimbabwe, during the 2018/2019 cropping season. Data was transformed using log10(x+2.5).
Effects of CA and SFA on individual beetle abundance
During the R1 and R6 stages of the maize crop, tillage had a significant effect on total beetle abundance (P = .013, F = 76.94, DF = 1) and Graphipterus cordiger abundance (P = .034, F = 27.79, DF = 1), respectively (). In particular, for these respective maize growth stages, Graphipterus cordiger abundance declined by 2.5% while total beetle abundance increased by 4.6% in RT relative to CT. However, for all other beetle species, there was no significant effect (P > . 05) of tillage across the three maize growth stages.
Crop residue application showed a significant effect on P. assimilis (P = .038, F = 9.23, DF = 1) and total beetle abundance (P = .040, F = 8.95, DF = 1); Z. boei (P = .027, F = 11.77, DF = 1) as well as G. cordiger (P = .006, F = 27.79, DF = 1) during the VT, R1 and R6 stages of maize growth, respectively (). During VT, total and P. assimilis abundances were two and three fold higher in crop residue treated than bare plots. Similarly, plots that had crop residues retained on the soil surface contained 8% more Z. boei individuals than bare plots during the R1 stage. In contrast, a 2.5% decline in the abundance of G. cordiger was observed in crop residue treated plots during physiological maize crop maturity, relative to those without crop residues. The main effects of crop rotation and SFA on beetle species abundance were not significant (P > .05) throughout the three maize growth stages.
During the VT stage, there was a significant interaction of tillage system, crop residue application and SFA on Dromica limpompoipana (P < .001, F = 4.69, DF = 5; ). For RT, SFA and crop rotation had no effect (P > 0.05) on D. limpompoipana. However, in CT, D. limpompoipana abundance was higher in M-60 relative to LF and M under mono-cropping. Moreover, for this tillage system, all the manure treated plots (M, M-60 and MLF) as well as the control under the maize-soyabean crop rotation had fewer D. limpompoipana than LF plots.
Table 4. Mean (in parentheses) and Log(x + 2.5) transformed abundance of Dromica limpompoipana, total and Renatiella reticulata in response to tillage, crop residue application, crop rotation and soil fertility amendment at Hunyani farm, Chinhoyi, Zimbabwe, during the 2018/2019 cropping season.
Significant interactions (P = .007, F = 3.46, DF = 5) of crop rotation × SFA on total abundance were observed during the R1 stage. In the mono cropping system, HF resulted in a decline in total beetle abundance relative to manure treated plots (M and MLF, ). On the other hand, for ROT, HF plots contained 2–3 times more total beetles compared to LF and M-60. Whereas crop residue application had no effect on P. assimilis abundance in mono cropped plots, abundance of this species showed a tendency to increase in crop residue treated plots under maize/soyabean rotation ().
Table 5. Mean (in parentheses) and Log(x + 2.5) transformed abundance of Platynus assimilis, Zophosis boei and Graphipterus cordiger in response to tillage, crop residue application, crop rotation and soil fertility amendment at Hunyani farm, Chinhoyi, Zimbabwe, during the 2018/2019 cropping season.
During the R6 stage, significant interactions of tillage system × SFA (P = .018, F = 2.91, DF = 5), crop rotation × SFA (P = .020, F = 2.85, DF = 5) and crop residue application × crop rotation × SFA (P = .029, F = 2.65, DF = 5) on Renatiella reticulata; and tillage system × crop residue application (P = .006, F = 27.79, DF = 1) on G. cordiger were observed. With respect to CT, except for LF, soil fertility amended plots contained fewer R. reticulata than the untreated control (). On the other hand, HF resulted in a 628.6% increase in R. reticulata compared to M-60, MLF and the control. While no effects of SFA were evident under mono cropping, it was observed that except for LF, application of SFA in a maize/soyabean cropping sequence resulted in reduced R. reticulata abundance relative to the untreated control (). Furthermore, SFA and crop rotation had no effect on R. reticulata abundance when crop residues were removed from the plot (). However, when crop residues were applied and continuous maize was grown (MONO), HF resulted in more R. reticulata individuals than M, M-60 and the control. For crop residue application + maize/soyabean rotation, all SFA treatments except M-60 and LF resulted in reduced R. reticulata abundance compared to the control. Graphipterus cordiger showed no response to crop residue application in RT, but was significantly reduced when crop residues were applied on CT plots (). Meanwhile, Z. boei almost doubled in crop residue treated plots under MONO but showed no response to crop residue application in ROT (). All other interactions were not significant (P > .05).
Effects of CA and SFA on the diversity of surface-dwelling beetles
Crop residue application had a significant effect on beetle Shannon diversity (P = .044, F = 8.37, DF = 1), richness (P = .014, F = 17.61, DF = 1) and evenness (P = .021, F = 13.66, DF = 1). The Shannon and richness indices were almost double in crop residue treated plots compared to plots where no crop residues were applied (Shannon = 0.29 vs. 0.13, richness = 1.24 vs. 0.76, ). However, tillage, crop rotation and SFA had no significant effect (P > .05) on beetle species richness, evenness and diversity. There was a significant interaction (P = .053, F = 2.21, DF = 5) of crop rotation, crop residue application and SFA. Specifically, the application of sole manure at 10 t ha−1 resulted in a 360-365% increase in the Shannon index relative to LF and MLF ().
Figure 1. Effect of crop residue application, crop rotation and soil fertility amendment on beetle diversity (Shannon index) in maize at Hunyani farm, Chinhoyi, Zimbabwe, during the 2018/2019 cropping season. Error bars are ± standard error of difference (SED) for the comparison of soil fertility amendment and crop rotation means within and across crop residue application treatments. Mono = mono cropping with maize, Rotation = maize/soyabean rotation, HF = soil high fertiliser, LF = low fertiliser, M = cattle manure, MLF = cattle manure + low fertiliser, M-60 = cattle manure + 60 kg of N ha−1 MHF = cattle manure + high fertiliser.
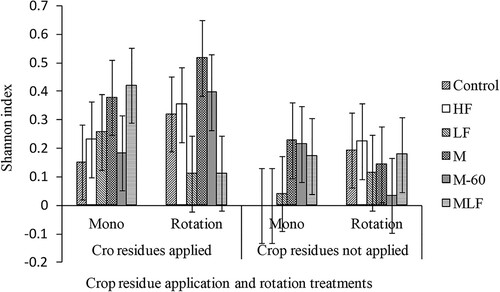
Table 6. Tillage, crop residue application and soil fertility amendment effects on beetle diversity during the maize growth cycle at Hunyani farm, Chinhoyi, Zimbabwe, during the 2018/2019 cropping season.
Maize grain yield and season quality
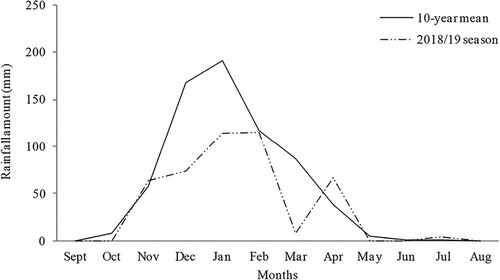
Compared to the 10-year average (670 mm), annual rainfall for 2018/2019 cropping season (470 mm) was 33.4% lower, with an extended mid-season dry spell between January and April 2019 (). Tillage and SFA had no significant effect (P > .05) on the maize grain yield. However, there was a significant effect (P < .05) of crop residue application on maize grain yield (). In particular, maize grain yield almost trebled in crop residue treated plots (2471 kg ha−1) compared to plots that had no crop residues (902 kg ha−1).
Figure 2. Ten-year mean (2007/2008–2017/2018) and 2018/2019 season rainfall for Hunyani farm, Chinhoyi, Zimbabwe.
Discussion
Beetle community abundance
The results of the present study () agree with the findings of a recent study in Zimbabwe which found species such as Zophosis boei to be among the most dominant surface-dwelling beetles in maize fields (Mashavakure, Mashingaidze, Musundire, Gandiwa, et al. Citation2019). Most of the beetles that were observed were beneficial, that is predators, e.g. Platynus assimilis and detritivores e.g. Renatiella reticulata and Z. boei. These findings, therefore, suggest that under the soil and environmental conditions of this specific study, surface-dwelling beetles are among soil biota that may be important in driving biological processes in cropping systems. As shown in , this study further revealed that reduced tillage (RT) increases surface-dwelling beetle abundance compared to conventional tillage (CT). These results corroborate findings of several previous researchers who assert that RT supports higher populations of soil dwelling beetle communities (Kromp Citation1999; Shrestha and Parajulee Citation2010; Soane et al. Citation2012; Mashavakure, Mashingaidze, Musundire, Gandiwa, et al. Citation2019; Mashavakure, Mashingaidze, Musundire, Nhamo, et al. Citation2019). Contrary to CT which causes habitat destruction and food depletion, in RT systems, the soil habitat structure is preserved and this provides stable breeding, hiding and ambush sites for the surface-dwelling beetle communities.
Retention of crop residues on the soil surface increased the abundances of beetle groups such as P. assimilis, Z. boei and total beetles (). These results are in agreement with Dudás et al. (Citation2016) and Gill et al. (Citation2011) who found larger populations of ground beetle and other surface insect communities in plant residues treated than untreated plots. Crop residue mulch helps to moderate microhabitat conditions such as soil moisture and temperature, making them more favourable for beetle survival and reproduction. Surface residues also provide food for detrivorous and shelter for predatory surface insects. However, the role of crop residues in regulating beetle populations was not quantified in the present study. Interestingly, Graphipterus cordiger seemed to be negatively affected by CA principles namely, namely RT and crop residue application (R) as shown in . Shrestha and Parajulee (Citation2010) found that some surface-dwelling beetle species are positively affected by soil inversion tillage. A possible explanation for the positive response of G. cordiger to soil inversion tillage could be that key life stages of this particular species do not coincide with tillage operations, enabling it to evade mortality due to direct impacts of habitat structural disturbance.
Contrary to Brust and King (Citation1994) and in agreement with O’rourke et al. (Citation2008), we found no effect of crop rotation on surface-dwelling beetle communities. This is probably related to differences in soil and climatic conditions from which the reported studies were conducted. The lack of beetle community response to soil fertility amendment (SFA) seem to agree with Mashavakure et al. (Citation2018), Mashavakure, Mashingaidze, Musundire, Gandiwa, et al. (Citation2019) and Mashavakure, Mashingaidze, Musundire, Nhamo, et al. (Citation2019) who recently found no effect of fertiliser treatment on agrobiont macroinvertebrates namely nematodes, spiders and surface-dwelling beetles. In these studies, the authors found that some beetles including Z. boei were associated with the cold dry season, hence in the present study, their presence during the later part of the cropping season probably signals the commencement of population build up as the winter season approaches.
Effects of treatment interactions on beetle communities
There is evidence from this present study that abundances of surface-dwelling beetles can be increased by application of manure alone or in combination with an inorganic fertiliser (sole manure: M, manure + 60 kg of N ha−1: M-60, and manure + low rate of inorganic fertiliser: MLF) in mono cropping systems (MONO), and inorganic fertiliser (low rate of inorganic fertiliser: LF and high rate of inorganic fertiliser: HF) in maize/soyabean rotation systems as shown in . These results concur with the findings of Sadej et al. (Citation2012) who found higher beetle abundance and diversity after application of organic manure. The application of organic manure may positively influence the beetles’ capability of overwintering, burrowing, oviposition and also availability of prey. This is so because soils which contain high organic carbon which is the primary content of soil humus are readily populated by a variety of soil organisms. Animal manure may be an essential food substrate for soil dwelling beetle communities and therefore support higher populations of these important macroinvertebrates (Abdallah et al. Citation2016). In cereal/legume rotation systems (ROT), biological processes such as biological nitrogen fixation are important in providing essential soil constituents including organic carbon and therefore, such habitats can still support sufficient populations of beetles without any need for manure application. Meanwhile, we observed that some beneficial beetle species like P. assimilis in ROT when crop residues were retained on the soil surface. Again crop residues provide essential substrate for refuge as well as food for prey populations.
It is noteworthy that a detritivorous beetle species, R. reticulata, exhibited the most prominent response to treatment interactions (). Evidence from the present study suggests that in farming systems where either RT or MONO + crop residue retention is practised, application of sole inorganic fertiliser at a high rate (HF) is important in increasing the populations of R. reticulata. It can be argued that RT and crop residue application create conditions for high biological activities that provide adequate amounts of soil carbon and such systems will not require amendment with organic fertiliser such as manure. Meanwhile ROT + crop residue retention have the twin benefits of increased soil carbon and nitrogen due to BNF. This system will therefore not benefit from either manure application or high nitrogen application. For detritivorous species such as Z. boei, the results suggest that farmers who practice mono-cropping can increase beetle population and activity by application of crop residues on the soil surface (). As argued earlier on, higher abundance of beetles in crop residue treated plots may be indirectly related to high moisture and improved soil physical properties caused by the addition of organic material thus encouraging survival and population build up of surface dwelling macro-invertebrates.
Surface-dwelling beetle diversity
Crop residue application on the soil surface was most important in influencing surface dwelling beetle diversity, with higher diversity estimates in crop residue treated than bare plots (). These results were in agreement with Dudás et al. (Citation2016) who found that the application of organic mulch significantly increased the diversity of carabid beetles. Of the three CA principles, R + ROT resulted in the highest surface-dwelling beetle diversity when 10 t ha−1 of manure were applied (). It can be argued that this combination provided favourable soil conditions such as soil moisture, soil carbon and stable sites to support diverse species of beetles.
Our present study has potential limitations. Firstly, the study was conducted during a drier year, that is total rainfall received during the cropping season was 66.6% of the 10-year average precipitation for the same site. Drier conditions are known to be associated with reduced biological activity including macro-arthropod activity. This could be the reason for fewer beetle catches during the four-month sampling period. However, it is noteworthy that treatment effects on surface-dwelling beetles were in agreement with previous findings by researchers in other geographical regions. This suggests that the findings of this present study can be a reliable basis for drawing meaningful conclusions and recommendations for management despite these limitations.
Maize grain yield
As illustrated in , tillage, crop rotation and SFA showed no effect on maize grain yield. However, application of crop residues on the soil surface increased maize grain yield. This was most likely because in a dry year like the 2018/2019 season, surface residue application is more important in regulating crop yield than mechanical soil disturbances (Kodzwa et al. Citation2020; Mashavakure, Mashingaidze, Musundire, Gandiwa, et al. Citation2019).
Table 7. Tillage, crop residue application and soil fertility amendment effects on maize grain yield (kg ha−1) at Hunyani farm, Chinhoyi, Zimbabwe, during the 2018/2019 cropping season.
The present study revealed that (1) reduced tillage and crop residue application had positive effects on surface-dwelling beetle abundance. Crop rotation and soil fertility amendment (SFA), particularly when applied alone, had no effect on surface-dwelling beetle abundance. (2) Organic SFA in systems in which mono cropping as well as crop residue removal were practised, and inorganic SFA in maize/soyabean rotation as well as crop residue treated systems were found to increase surface-dwelling beetle abundance. Finally, (3) when some two conservation agriculture (CA) principles, crop residue application and crop rotation, were used, applications of well-rotted cattle manure profoundly increased surface-dwelling beetle diversity. We, therefore, recommend that farmers who practise CA should use organic SFA in order to enhance surface-dwelling beetle diversity and realise some potential benefits such as biological pest control and nutrient cycling. Further studies should be focused on understanding the specific ecological roles of the beetle species that were observed in this study.
Acknowledgements
The authors are grateful to the field assistant at Hunyani farm, Ms Chipo Chirimuuta and the rest of the staff at the farm, for their assistance in trial management and data collection. Nilton Mashavakure conceptualised this present study, supervised the implementation of the experiment, supervised data collection, analysed the data and prepared the manuscript. Nyaradzo M. Chomufana implemented the experiment and collected the data. Kumbirai Musiyiwa supervised the implementation of the experiment, supervised data collection and contributed to the preparation of the manuscript. Justice Nyamangara conceptualised the global research from which the data for this present study were collected and contributed to the preparation of the manuscript.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
Notes on contributors
Nilton Mashavakure
Nilton Mashavakure Holds a PhD in agro-ecology and is a senior academic and researcher of sustainable farming systems. Currently coordinator of Crop Research in the University.
Nyaradzo M. Chomufana
Nyaradzo M. Chomufana Is a graduate with a BSc in Crop Science and Technology.
Kumbirai Musiyiwa
Kumbirai Musiyiwa Holds a PhD in Soil Science. Is a senior academic, research and current deputy Dean of the School of Agciultural Sciences and Technology.
Justice Nyamangara
Justice Nyamangara Hold a PhD in Soil Science, Founding Vice Chanellor of Marondera University of Agrcicultural Sciences and Technology, senior research and acedemic.
References
- Abdallah M, Mwatawala MW, Kudra AB. 2016. Abundance and dispersal of Heteronychus arator (Coleoptera: Scarabaeidae) in maize fields under different fertilizer treatments. SpringerPlus. 5:2–9.
- Adediran JA, Taiwo LB, Akande MO, Sobulo RA, Idowu OJ. 2005. Application of organic and inorganic fertilizer for sustainable maize and cowpea yields in Nigeria. J Plant Nutr. 27:1163–1181. DOI: 10.1081/PLN-120038542.
- Boyetchko SM, Bailey KL, De Clerck-Floate RA. 2009. Current biological weed control agents- their adoption and future prospects. Praire Soils Crops. 2:38–45.
- Brust GE, King LR. 1994. Effects of crop rotation and reduced chemical inputs on pests and predators in maize agroecosystems. Agric Ecosyst Environ. 48:77–89.
- da Silva PM, Aguiar CAS, Niemela J, Sousa JP, Serrano RM. 2008. Diversity patterns of ground beetles (Coleoptera: Carabidae) along a gradient of land-use disturbance. Agric Ecosyst Environ. 124:270–274.
- Diekotter T, Wamser S, Wolters V, Birkhof K. 2010. Landscape and management effects on structure and function of soil arthropod communities in winter wheat. Agric Ecosyst Environ. 137:108–112.
- Druilhe Z, Barreiro-Hurlé J. 2012. Fertilizer subsidies in sub-Saharan Africa. Rome: Food and Agriculture Organization of the United Nations, Agricultural Development Economics Division (ESA). ESA Working Paper No. 12-04.
- Dudás P, Gedeon C, Menyhárt L, Ambrus G, Tóth F. 2016. The effect of mulching on the abundance and diversity of ground beetle assemblages in two Hungarian potato fields. Columella: J Agric Environ Sci. 3:45–53.
- Gill HK, Mcsorley R, Branham M. 2011. Effect of organic mulches on soil surface insects and other arthropods. Fla Entomol. 94:226–232.
- Greenslade PJM. 1964. Pitfall trapping as a method for studying populations of Carabidae (Coleoptera). J Anim Ecol. 33:301–310.
- Hammer Ø, Harper DAT, Paul DR. 2001. Past: paleontological statistics software package for education and data analysis. Palaeontol Electron. 4:1–9.
- Holland JM, Luff ML. 2000. The effects of agricultural practices on Carabidae in temperate agroecosystems. Integr Pest Manag Rev. 5:109–129.
- Holopainen JK, Helenius J. 1992. Gut contents of ground beetles (Col.: Carabidae) and activity of these and other epigeal predators during an outbreak of Sitobion avenae. (Hom. Aphididae). Acta Agric Sand Sect B. 42:57–61.
- Jat RA, Wani SP, Sahrawat KL. 2012. Conservation agriculture in the semi-arid tropics: prospects and problems. Adv Agron. 117:191–273.
- Johnson SN, Lopaticki G, Barnett K, Facey SL, Powell JR, Hartley SE. 2016. An insect ecosystem engineer alleviates drought stress in plants without increasing plant susceptibility to an above-ground herbivore. Funct Ecol. 30:894–902.
- Jones CG, Lawton JH, Shachack M. 1994. Organisms as ecosystem engineers. Oiskos. 69:373–386.
- Jowett K, Milne AE, Garrett D, Potts SG, Senapathi D, Storkey J. 2020. Above- and below-ground assessment of carabid community responses to crop type and tillage. Agric For Entomol. 23:12397.
- Kergunteuil A, Bakhtiari M, Formenti L, Xiao Z, Defossez E, Rasmann S. 2016. Biological control beneath the feet: A review of crop protection against insect root herbivores. Insects. 7:70.
- Kodzwa JJ, Gotosaa J, Nyamangaraa J. 2020. Mulching is the most important of the three conservation agriculture principles in increasing crop yield in the short term, under sub humid tropical conditions in Zimbabwe. Soil Tillage Res. 197:104515.
- Kromp B. 1999. Carabid beetles in sustainable agriculture: a review on pest control efficacy, cultivation impacts and enhancement. Agic Ecosyst Environ. 74:187–228.
- Labruyere S, Bohan DA, Biju-Duval L, Ricci B, Petit S. 2016. Local, neighbor and landscape effects on the abundance of weed seed-eating carabids in arable fields: a nationwide analysis. Basic Appl Ecol. 17:230–239.
- Lavelle P, Lattaud C, Trigo D, Barois I. 1995. Mutualism and biodiversity in soils. Plant Soil Environ. 170:23–33.
- Liebman M, Gallant ER. 1997. Many little hammers: ecological management of crop-weed interactions. In: L. E. Jackson, editor. Ecology in agriculture. San Diego (CA): Academic Press; p. 291–393.
- Lindroth CH. 1968. The ground-beetles (Carabidae, excl. Cicindelinae) of Canada and Alaska Lund, Sweden.
- Luff ML. 1987. Biology of polyphagous ground beetles in agriculture. Agric Zool Rev. 2:237–278.
- Mashavakure N, Mashingaidze AB, Musundire R, Gandiwa E, Muposhi VK, Thierfelder C, Nhamo N, Bere T, Akhtar SS. 2018. Short-term impacts of tillage and fertilizer treatments on soil and root borne nematodes and maize yield in a fine textured cambisol. J Nematol. 50:329–342.
- Mashavakure N, Mashingaidze AB, Musundire R, Gandiwa E, Thierfelder C, Muposhi VK. 2019. Beetle and maize yield response to plant residue application and manual weeding under two tillage systems in northern Zimbabwe. Appl Soil Ecol. 144:139–146.
- Mashavakure N, Mashingaidze AB, Musundire R, Nhamo N, Gandiwa E, Thierfelder C, Muposhi VK. 2019. Soil dwelling beetle community response to tillage, fertilizer and weeding intensity in a sub-humid environment in Zimbabwe. Appl Soil Ecol. 135:120–128.
- Mazvimavi K, Twomlow S. 2009. Socioeconomic and institutional factors influencing adoption of conservation agriculture by vulnerable households in Zimbabwe. Agric Syst. 101:20–29.
- Mugwira LM, Nyamangara J, Hikwa D. 2002. Effects of manure and fertilizer on maizeat a research station and in a smallholder (peasant) area of Zimbabwe. Commun Soil Sci Plant Anal. 33:379–402.
- O’rourke ME, Liebman M, Rice ME. 2008. Ground beetle (Coleoptera: Carabidae) assemblages in conventional and diversified crop rotation systems. Environ Entomol. 37:121–130.
- Sadej W, Kosewska A, Sadej W, Nietuoski M. 2012. Effects of fertilizer and land-use type on soil properties and ground beetle communities. Bull Insectology. 65:239–246.
- Shrestha RB, Parajulee MN. 2010. Effect of tillage and planting date on seasonal abundance and diversity of predacious ground beetles in cotton. J Insect Sci. 10:1–17.
- Sileshi GW, Jama B, Vanlauwe B, Negassa W, Harawa R, Kiwia A, Kimani D. 2019. Nutrient use efficiency and crop yield response to the combined application of cattle manure and inorganic fertilizer in sub-Saharan Africa. Nutr Cycling Agroecosyst. 113:181–199.
- Soane BD, Ball BC, Arvidsson J, Basch G, Moreno F, Roger-Estrade J. 2012. No-till in northern, western and south-Western Europe: a review of problems and opportunities for crop production and the environment. Soil Tillage Res. 118:66–87.
- Steinmann H-H, Dobers ES. 2013. Spatio-temporal analysis of crop rotations and crop sequence patterns in northern Germany: potential implications on plant health and crop protection. J Plant Dis Prot. 120:85–94.
- Stinner BR, House GJ. 1990. Arthropods and other invertebrates in conservation tillage agriculture. Annu Rev Entomol. 35:299–318.
- Tandzi LN, Mutengwa CS. 2020. Estimation of maize (Zea mays L.) yield per harvest area: appropriate methods. Agron Sustain Dev. 10:29. DOI:10.3390/agronomy10010029.
- Triplehorn CA. 1965. Revision of Diaperini of America north of Mexico with notes on extralimital species (Coleoptera: Tenebrionidae). Proc U.S. Natl Mus. 117:349–457.
- Vanlauwe B, Wendt J, Giller K, Corbeels M, Gerard B, Nolte C. 2014. A fourth principle is required to define conservation agriculture in sub-Saharan Africa: The appropriate use of fertilizer to enhance crop productivity. Field Crops Res. 155:10–13.
- VSN-International. 2011. Genstat for Windows. 10th ed. Hemel, Hempstead: VSN-International.
- WRB. 2014. World reference base for soil resources: International soil classification system for naming soils and creating legends for soil maps. World Soil Resources Reports.