Abstract
A modified potometer consisting of a water-filled glass tube mounted in parallel with a metric scale and connected to a fruit was used to study water uptake into detached fruits of sweet cherry. Results describing quantities of imported water from some introductory studies were in the same range as in previously published studies conducted with alternative methods. Water uptake was measured in fruits of four cultivars kept in air of approximately 50±5% RH followed by immersion in water 1–2 h into the experimental period. This caused a significant decrease in water uptake. However, the fruits continued to accumulate water at a reduced rate. When the fruits were dried, the water uptake immediately increased to the same level as before the immersion in water. The fruit skin of two cultivars was exposed to normal air after being ruptured with four shallow cuts to simulate cuticular fractures or shallow cracks. These two treatments did not give statistically different water uptake through the pedicel, although there was a tendency to an increased rate of water uptake in fruits with ruptured fruit skin.
Introduction
Potometry is an experimental method of studying water absorption by whole plants or parts of plants by placing the plant or plant part in a closed container equipped with a graduated tube allowing volumetric measurement of water uptake. During potometric experiments the plant or plant part may be exposed to different environmental conditions (Kramer, Citation1959).
A problem using plant parts in potometric systems is the callus-forming ability of the sieve tube elements; callus is usually formed in abundant amounts within 20–30 min after cutting the plant part. The callus effectively blocks transport of solutes in the sieve tubes (Eschrich, Citation1975).
In situ measurements of water uptake by stone fruits are difficult; Christensen (Citation1973) measured fruit weight of detached cherry fruits to study fruit expansion, Lilleland & Newsom (Citation1934) and Kozlowski (Citation1968) measured fruit volume changes in attached fruits, and similarly Lang & Thorpe (Citation1989) in grapes. These methods did not, however, take into consideration that part of the total water import to the fruit which evaporates from the fruit surface. Appropriate methods for directly measuring quantitatively the water uptake into fruits have accordingly been lacking.
Potometry is an alternative experimental method with which to measure water uptake by plant parts, and is used for measurement of overall water management in plants (Scholander et al., Citation1955, Citation1957). Potometry as a method of studying water flow and the factors which influence it might therefore contribute to a better understanding of water interactions in sweet cherry fruits in different environments. In addition, the method is simple and inexpensive.
This paper presents and discusses potometry as a method of studying water flow through the fruit pedicel into detached sweet cherry fruits. Measurements of water uptake through sweet cherry pedicels in fruits immersed in water to simulate rain, and in fruits with injured fruit surface to simulate cracks or cuticular fractures, are described.
Materials and methods
The potometric apparatus
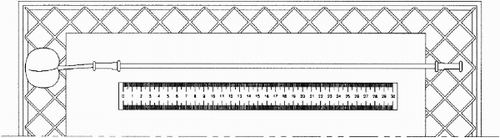
The potometric apparatus was assembled from a capillary glass tube with internal diameter 0.6±0.1 mm mounted parallel to a metric scale on a rectangular piece of cardboard. In both ends of the glass tube was mounted a piece of rubber tubing (5 cm long, internal diameter 2.5 mm) with back-folded ends (). The potometer was partly filled with water by adding vacuum pressure from one end with a syringe coincidentally with submerging the other end into distilled water until a water meniscus appeared in the glass tube. A dye solution (NIGROSIN®) was added to the water to make the water meniscus better detectable. A fruit was mounted in the waterfilled end of the potometer and the vacuum pressure syringe was then removed. The apparatus was placed horizontally onto a grate. Readings of the water meniscus position were made every 15 min for 5–6 h. When necessary, the potometer was refilled within the experimental period by adding water into the fruit-end rubber tubing using a syringe with a thin needle.
Fig. 1 Schematically shown potometric apparatus modified to measure water uptake in cherry fruit pedicels. A fruit is mounted in the end of a waterfilled capillary glass tube through a rubber tubing. The glass tube is mounted parallell to a metric scale on a rectangular piece of cardboard. The apparatus is placed horizontally onto a grate and readings are made every 15 min.
Accumulated water W f per fruit pedicel at any time of measurement was calculated by:
Plant material
Fruits grown in the experimental field at Ullensvang Research Centre, Lofthus, Norway on trees receiving standard treatments regarding irrigation, fertilization and plant protection spray applications were used.
Sweet cherry fruits with a pedicel end (2 cm) were cut from the tree under water to avoid entry of air into the vessel elements and brought immediately to the laboratory. The watercovered pedicel ends were mounted in the waterfilled end of the potometer. A drop of liquid, fast-drying glue was placed in the mounting connection to seal it and the fruit was dried carefully with tissue paper. Standard conditions such as morning picked fruits similar in appearance regarding size and ripeness and approximately standardized environmental conditions were applied. The surrounding air had 50±5% relative humidity (RH) during the experimental period (Hanna Instruments HI 9065 Thermo Hygrometer™).
Introductory water uptake studies
Water uptake in samples of detached fruits was measured in an introductory study. Subsequently, water uptake in pedicels of intact fruits was compared to the corresponding uptake in pedicels from which the fruits were removed. To detect any leaching from the potometer, a control potometer with a clamp closing the rubber was included.
The two experiments were run over a five-day period (July 18–22 1994) and fruits of ‘Merton Glory’ were used. The temperature in the laboratory varied from +25°C in the morning to +28°C in the afternoon all days.
Water uptake of fruits immersed in water and with ruptured fruit skin
Fruits of ‘Merton Glory’ and ‘Ulster’ harvested on July 25, 26, 27 1994 and July 28, 29 1994, respectively were mounted in the potometric apparatus. Two hours into the experimental period the fruits were exposed to a water-saturated environment by covering the fruits with constantly wet tissue paper. After two hours the fruits were carefully dried with tissue paper. Water uptake was measured throughout a 6-h experimental period. The experiment was repeated with fruits of ‘Sunburst’ (August 14) and ‘Rainier’ (August 15 and 18) in 1997.
Fruits of ‘Van’ and ‘Sunburst’ harvested in 1994 on August 9 and 25, respectively, were similarly mounted in the potometric apparatus. Shortly into the experimental period the fruits were ruptured with 4 razor blades (1–2 mm into the fruit skin) to simulate cracks or cuticular fractures. Intact fruits in normal air served as controls in all experiments.
All the experiments are summarised in with cultivars, date of experiment, number of fruits and the order of applied treatment.
Table 1. Two years of experiments with potometric water uptake by sweet cherry fruit by experiment, cultivar, date of experiment, number of fruits and order of applied treatments. Air is normal air of approximately 50±5% RH and immersed in water means that the fruits were covered with continously wet tissue paper. When the fruit skin was ruptured it was cut with 4 razor blades 1–2 mm into the fruit flesh
Single fruits were used as replicates in all the experiments, numbers varying from 3–10 fruits. The relationship between variables was analysed by linear regression and analysis of variance (ANOVA) using the statistical package Minitab®.
Results
Introductory water uptake studies
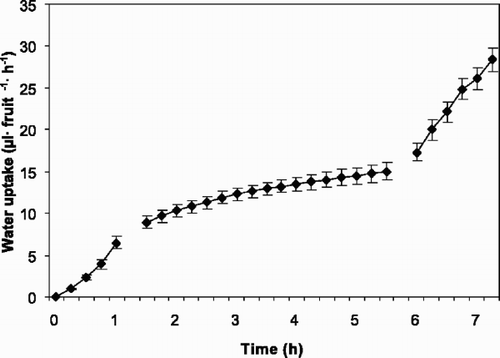
Net water accumulation into intact fruits was in the range 9.2–11.6 μl fruit−1 h−1, similar to the results presented in the upper part of . Accumulated water (y) in mean of 3 fruits expressed over a 5-h period (x) fitted well to a linear model, with the regression equation y=–0.54+0.18x, r 2=99.8, P=0.001.
Fig. 2 Potometrically measured accumulated water uptake through sweet cherry fruit pedicels with and without fruits of ‘Merton Glory’ (1994) for 5 h.
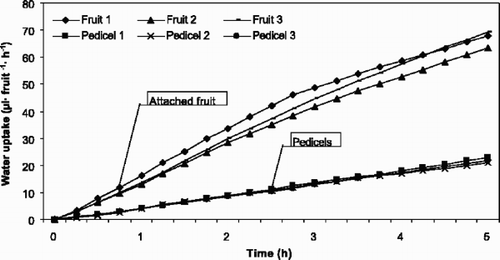
Similar, but slightly greater water accumulation (12.7–13.5 μl fruit−1 h−1) was observed in the pedicels of intact fruits in the comparative experiment. Through pedicels from which the fruits were removed, however, significantly less flow was measured (4.2– 4.6 μl fruit−1 h−1), indicating an average reduction of 67% ().
In all the controls, where a clamp closed the potometer at one end, only negligible water loss was observed, indicating no water wastage in the potometric experiments reported.
Water uptake of fruits immersed in water and with ruptured fruit skin
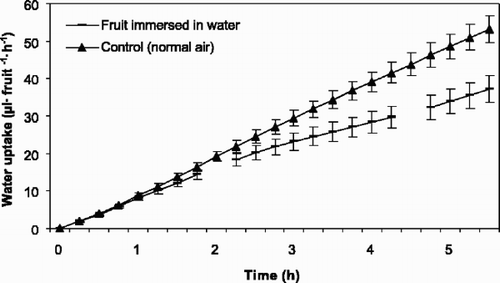
The rate of water uptake through the pedicel decreased significantly (P≤0.000) when fruits of ‘Merton Glory’ were immersed in water (). The pedicel absorbed, before immersion in water, an average of about 2.5 μl fruit−1h−1 more than after immersion.
Fig. 3 Accumulated water uptake through the pedicels of fruit of sweet cherry ‘Merton Glory’ (1994) kept in air and interrupted by fruit immersion in water for 3 h. The first break in line is when the fruits were immersed in water, while the second break is when the fruits were dried. Vertical bars represent standard error of the means (SE) and each point represents 10 fruits. Regressions describing the data are: (▴) y=–0.69+9.90x, r 2=0.865; (-) y=–0.14+8.21x, r 2=0.840; y=0.98+5.06x, r 2=0.523 and y=3.91+1.54x, r 2=0.090 before, during and after immersion, respectively.
The rate of water uptake decreased also in fruits of ‘Ulster’, ‘Rainier’ and ‘Sunburst’ ( –), although not significantly for ‘Ulster’ (P=0.197, P=0.044, and P=0.000, for the three cultivars respectively) when immersed in water. The decrease was on average 4.0, 4.9 and 5.0 μl fruit−1 h−1 for the three cultivars, respectively.
Fig. 4 Accumulated water uptake through the pedicel of fruits of sweet cherry ‘Ulster’ (1994) kept in air and interrupted by fruit immersion in water for 3 h. The first break in line is when the fruits were immersed in water, while the second break is when the fruits were dried. Vertical bars represent standard error of the means (SE) and each point represents 5 fruits. Regression describing the data are: (▴) y=−1.10+8.32x, r 2=0.984; (-) y=0.18+8.15x, r 2=0.911; y=0.64+4.69x, r 2=0.954 and y=−0.52+5.75x, r 2=0.971 before, during and after immersion, respectively.
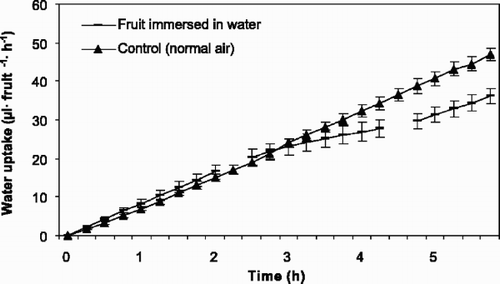
Fig. 5 Accumulated water uptake through the pedicel of fruits of sweet cherry ‘Rainier’ (1997) kept in air and interrupted by fruit immersion in water for 3 h. The first break in line is when the fruits were immersed in water, while the second break is when the fruits were dried. Vertical bars represents standard error of the means (SE) and 9 fruits were immersed in water in addition to 5 control fruits. Regression describing the data are: (▴) y=0.79+6.17x, r 2=0.923; (-) y=0.11+8.08x, r 2=0.536; y=0.98+3.38x, r 2=0.648 and y=0.13+4.30x, r 2=0.334 before, during and after immersion, respectively.
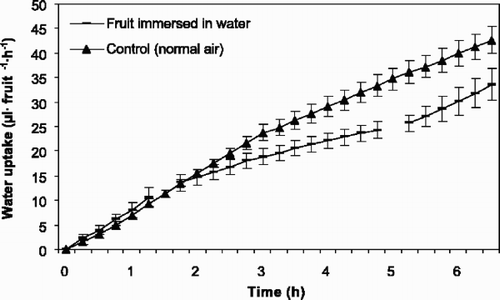
Fig. 6 Accumulated water uptake through the pedicel of fruits of sweet cherry ‘Sunburst’ (1997) in air and interrupted by fruit immersion in water for 4 h. The first break in line is when the fruits were immersed in water, while the second break is when the fruits were dried. Vertical bars represents standard error of the means (SE) and each point represents 9 fruits. Regression describing the data are: y=−0.53+6.55x, r 2=0.780; y=0.82+1.43x, r 2=0.553 and y=0.11+9.64x, r 2=0.792 before, during and after immersion, respectively.
Linear regression fitted well for regression equations describing the curves, although there might be a small curvature on the graphs. All the regression coefficients were significant except when fruits of ‘Merton Glory’ were dried after immersion in water.
The rate of water uptake through the pedicels of fruit of ‘Van’ and ‘Sunburst’ () did not increase statistically (P=0.065 and P=0.299, respectively) when the fruit surface was ruptured. Fruits with ruptured fruit skin absorbed, however, on average 2.4 μl fruit−1 h−1 and 1.4 μl fruit−1 h−1 more than control fruits for the two cultivars, respectively.
Table 2. Potometrically measured water uptake through the pedicel (μl fruit−1 h−1) and regression equations for the relationship between water uptake through the pedicel (y) and time (x) in two cultivars of sweet cherries with ruptured fruit skin (4 cuts with razor blades) and control fruits with intact fruit skin, 1997. Linear regression model y=a+bx
Discussion
Introductory water uptake studies
The net water import by single, healthy fruits was found to be in the range 9.2–13.5 μl fruit−1 h−1 in the two experimental series. Results from previous fruit expansion studies (Lilleland & Newsom, Citation1934; Kozlowski, Citation1968; Christensen, Citation1973) were comparable to the present studies if approximately converted to corresponding figures, as discussed by Sekse (Citation1998).
The strong reduction in water accumulation measured in pedicels where fruits were removed would be expected for either of two reasons, or both; a reduction of the transpiring surface and a removal of a possible sink effect in the intact fruit. The flow through stems lacking fruit was solely that which transpired from the pedicel surface (no drop formed on the stub during the experimental period). If differences in surface area were the sole explanation for the water uptake reduction, the difference in water uptake should have been significantly larger. This indicated that sweet cherry fruits, even detached, acted as sinks for water supply through their pedicels.
Cutting the plant material in water appeared to prevent any possible air blocking of the transport system (Scholander et al., Citation1957), since the net water uptake was comparable to results obtained by other researchers, as discussed above. Formation of callus that may block sieve cells or the collapse of such cells are other possible problems in potometric experiments (Eschrich, Citation1975; Salisbury & Ross, Citation1991). In the present experiment any blocking of callus or collapse, if it occurred, must have taken place within very short time after cutting (15 min), since no break in the curve expressing accumulated water occurred within the experimental period. However, some kind of obstruction, although negligible, might have occurred, and have been responsible for the slight decline in the uptake during the experimental period, indicated by a slight curvature of the lines expressing the accumulated water in all the experiments.
Water uptake of fruits immersed in water and with ruptured fruit skin
The water uptake through the pedicel was strongly affected when the fruits were immersed in water. Immersing the fruits resulted in less water import to the fruits. This could be expected since the transpiration most probably decreased to a pronounced extent; but the fruits actually continued to take up water to some extent, and this accumulation may have been a part of the cracking mechanisms by causing an increase in the internal fruit turgor.
Turgor pressure build-up in the fruit is a normal part of fruit growth (Coombe, Citation1976), and Sekse (Citation1995a) demonstrated evidence for influence of turgor pressure on the development of fruit cuticular fractures by altered water supply to the tree, but this build-up of turgor pressure has been questioned (Christensen, Citation1972). The results of the presented experiments might support the theory that turgor pressure inside the sweet cherry fruit is an acting force in the fruit cracking process (Yamamoto et al., Citation1990; Sekse, Citation1995b), but further research is necessary. The potometric system used can be a useful method in later studies of the effects of water uptake on cracking.
When the fruit skin was ruptured, fruits did not stastistically accumulate more water through the pedicel, although there was a tendency to that effect (). This tendency is probably a result of increased transpiration through the cuts. Experiments with fruits with cuticular fractures have proved that such fractures are pathways for water uptake (Hovland & Sekse, Citation2003). Fruits of Japanese Persimmon (Diospyros kaki Thunb.) had a pronounced increase in the concentration of soluble solids in the fruit flesh directly under cracks in the skin (Iwanami et al., Citation2002). This is probably an effect of a similarly increased water loss due to the fractures resulting in a concentration of the sugar content.
Conclusions
A modified potometric method of measuring water uptake through pedicels of detached cherry fruits has been described. It allowed measurements of water uptake into sweet cherry fruits; both into regular fruits, into fruits treated differently prior to the experiment and into fruits exposed to different conditions within the experimental period. Any blocking of the stem vessels due to callus formation was negligible over a 5-h experimental period. Fruits immersed in water had less water uptake through the pedicel compared to fruits in air of 50±5% RH, although they continued to take up water at a lower rate. Fruits with a ruptured fruit surface did not statistically increase water uptake when exposed to air of 50±5% RH compared to fruits without such ruptures.
Acknowledgments
Aksel Selberg is thanked for making the drawing. The Norwegian Research Council granted 6 months sabbatical leave for Lars Sekse at Lincoln University, New Zealand during which the method described was partly developed.
Additional information
Notes on contributors
Kari Louise Hovland *
Hovland, K. L. and Sekse, L. (Norwegian Crop Research Institute, Ullensvang Research Centre, N-5781 Lofthus, Norway). Water uptake through sweet cherry (Prunus avium L.) fruit pedicels: influence of fruit surface water status and intact fruit skin.Notes
Hovland, K. L. and Sekse, L. (Norwegian Crop Research Institute, Ullensvang Research Centre, N-5781 Lofthus, Norway). Water uptake through sweet cherry (Prunus avium L.) fruit pedicels: influence of fruit surface water status and intact fruit skin.
References
References
- Christensen , JV . 1972 . Cracking in cherries. IV: Physiological studies of the mechanisms of cracking . Acta Agric. Scand. , 22 : 153 – 162 .
- Christensen , JV . 1973 . Cracking in cherries. VI. Cracking susceptibility in relation to the growth rhythm of the fruit . Acta Agric. Scand. , 23 : 52 – 54 .
- Coombe , BG . 1976 . The development of fleshy fruits . Ann. Rev. Plant. Physiol. , 27 : 507 – 528 .
- Eschrich W 1975 Sealing systems in phloem In: Zimmermann, M. H. & Milburn, J. A. (eds.) Encyclopaedia of Plant Physiology. New Series Vol. 1. Transports in Plants I. Phloem Transport, Springer Verlag Berlin pp. 39–56
- Hovland , KL and Sekse , L . 2003 . The development of cuticular fractures in fruits of sweet cherries can vary with cultivar and rootstock . J. Am. Pom. Soc. , 57 : 58 – 62 .
- Iwanami , H , Yamada , M and Sato , A . 2002 . A large increase of soluble solids concentration at shallow concentric skin cracks in Japanese persimmon . Sci. Hort. , 94 : 251 – 526 .
- Kozlowski , TT . 1968 . Diurnal changes in diameters of fruits and tree stems of Montmorency cherry . J. Hort. Sci. , 43 : 1 – 15 .
- Kramer JP 1959 Transpiration and the water economy of plants In: Steward, F. C. (ed.) Plant Physiology Vol. II pp. 607–730 Academic Press New York
- Lang , A and Thorpe , MR . 1989 . Xylem, phloem and transpiration flows in grape: application of a technique for measuring the volume of attached fruits to high resolution using Archimedes’ principle . J. Exp. Bot. , 40 : 1068 – 1078 .
- Lilleland , O and Newsom , L . 1934 . A growth study of the cherry fruit . Proc. Am. Soc. Hort. Sci. , 32 : 291 – 299 .
- Salisbury FB Ross CW 1991 Transport in the phloem In: Plant Physiology, Wadsworth Publishing Company Belmont California pp. 161–188
- Scholander , PF , Love , WE and Kanwisher , JW . 1955 . The rise of sap in tall grapevines . Plant Phys. , 30 : 93 – 104 .
- Scholander , PF , Ruud , B and Leivestad , H . 1957 . The rise of sap in a tropical liana . Plant Phys. , 32 : 1 – 6 .
- Sekse , L . 1995a . Cuticular fracturing in fruits of sweet cherry (Prunus avium L.) resulting from changing soil water contents . J. Hort. Sci. , 70 : 631 – 635 .
- Sekse , L . 1995b . Fruit cracking in sweet cherries (Prunus avium L.). Some physiological aspects: a mini-review . Sci. Hort. , 63 : 135 – 141 .
- Sekse , L . 1998 . Fruit cracking mechanisms in sweet cherries (Prunus avium L.): a review . Acta Hort. , 468 : 637 – 648 .
- Yamamoto , T , Hosoi , K and Watanabe , S . 1990 . Relationship between the degree of fruit cracking of sweet cherries and the distribution of surface stress of the fruit analysed by a newly developed system . J. Jap. Soc. Hort. Sci. , 59 : 509 – 517 .