Abstract
Tiller demography and contribution to seed yield were studied in first year seed crops of Bromus inermis ‘L⊘far’ and Festuca pratensis ‘Salten’ planted on different dates and with increasing plant densities (A: 15 or 18 Jun/11 plants m−2, B: 15 or 30 July/44 plants m−2, C: 15 August or 10 September/178 plants m−2) in field trials at Landvik, SE Norway, in 1998 and 1999. While the total tiller population in most crops increased until seed harvest, it decreased during panicle elongation in crops of Bromus inermis and Festuca pratensis that had reached 2000 and 3500 tillers m−2 in early spring, respectively. Except for the fact that many of the primary tillers of Bromus inermis died after producing barren stems, tillers formed in August and September had the greatest chance of becoming reproductive and produced the longest stems with the heaviest inflorescences in both species. Most tillers produced in winter or early spring either remained vegetative or died, but spring-emerging tillers contributed up to 30% of the total seed yield in early-established, low-density crops of Festuca pratensis. The results are discussed in relation to former papers suggesting that inter-tiller transfer of flower induction stimuli is possible in some perennial grasses. It is concluded that tillers produced in winter and spring contribute more to seed yield in Festuca pratensis than in Bromus inermis.
Introduction
Many investigations have shown that the optimal plant density of perennial grasses is lower in seed than in forage production. (e.g., Fulkerson, Citation1959; Nordestgaard, Citation1975a, Citation1975b, Nordestgaard, Citation1977, Citation1979; Jonassen, Citation1980; Meijer, Citation1984; Jonassen & Torskenæs, Citation1994; Aamlid, Citation1994; Aamlid & Jonassen, Citation2006). This is true regardless of whether the species has a bunch-type growth habit with only intravaginal tillers, such as Festuca pratensis (Jonassen, Citation1980), or a rhizomatous growth habit allowing colonization of new areas, such as Bromus inermis (Jonassen & Torskenæs, Citation1994). For the former species, Nordestgaard (Citation1975a) argued that 50–100 plants per m2 was an optimal density for seed production. This may well be correct also for Bromus inermis, although the present authors have seen crops planted at only 10 plants per m2 to produce excellent seed yields in this rhizomatous species.
In order to flower and produce seed, Bromus inermis and Festuca pratensis require both primary induction in autumn and secondary induction in spring (Heide, Citation1994). Whereas the primary induction requirement in Bromus inermis is met by 4–6 weeks of short-day (SD) exposure with no response to classical vernalization treatments (Heide, Citation1984), Festuca pratensis shows limited response to photoperiod, but requires an extended period at low temperatures (0–10°C). Heide (Citation1988) determined that this period had to be at least 16 weeks, and although later studies have shown that shorter exposures may be sufficient (Fang et al., Citation2004; Aamlid, Citation2005; Ergon et al., Citation2006), the principle remains that seed crops must be established early enough to produce a sufficient number of strong tillers before photoperiod and/or temperature becomes inductive in autumn. In this context, first year seed yields of Festuca pratensis (Jonassen & Hillestad, Citation1990) and Bromus inermis (Jonassen, Citation1994) have been shown to decline significantly if sowing is delayed after mid-July.
With the possible exception of red fescue (Meijer, Citation1984; Aamlid, Citation2005), individual tillers of most perennial grasses do not have a distinct juvenile stage where they are non-receptive to primary induction stimuli. At least in Lolium perenne, new tillers may be vernalized as buds or while still in the leaf sheaths of their mother tillers (Kleinendorst, Citation1974). Alternatively, new tillers may indirectly receive primary induction stimuli through their vascular connections with parent tillers, as suggested by Havstad et al. (Citation2003, Citation2004) for Dactylis glomerata and Bromus inermis. With regard to seed production, a consequence of such indirect induction is that even tillers emerging in spring/early summer, and therefore not directly exposed to primary inductive conditions, are able to flower and produce seed in the same year. However, Havstad et al. (Citation2004) found no evidence for transfer of florigenic stimuli in a growth chamber study with Festuca pratensis.
In the present trial we established seed crops of Bromus inermis and Festuca pratensis on three different dates during summer, with a low plant density on the earliest date, an intermediate plant density on the intermediate date and a high plant density on the latest date. By doing so, we intended to obtain crops with different structures during the critical period of primary induction in autumn. Our objectives were to study the development and reproductive behaviour of tillers formed during consecutive periods, especially those formed in spring, in seed crops of different structures. Based on earlier studies in controlled environments (Havstad et al., Citation2003, Citation2004) we hypothesized that the chances for tillers produced in winter or spring would be 1) greater in crops planted early at low density than in crops planted later at a higher density, and 2) greater in Bromus inermis than in Festuca pratensis.
Materials and methods
Experimental
Two experiments were carried out at The Norwegian Institute for Agricultural and Environmental Research, Arable Crop Division Landvik (58°N) during 1998–2000. The first experiment was established in 1998 and the second in 1999. Both experiments were discontinued after seed harvested in the first ley year.
In the year of establishment, plants of Festuca pratensis ‘Salten’ (origin 67°N) and Bromus inermis ‘L⊘far’ (origin 61°N) were raised from seed and grown at 20°C and continuous light (150 µm m−2 s−1 from TL-33 fluorescent tubes) in a growth chamber for four weeks after seedling emergence. After establishment, the plants were transplanted into 1.5 m×1.5 m field plots on three different dates and at three different plant densities (crops A, B and C; ). At the time of transplanting, Bromus inermis and Festuca pratensis usually had 1 and 2–3 tillers per plant, respectively. The plots were laid out on silty loam soils with three blocks (replicates) in 1998 and two blocks in 1999. All plots were fertilized with 50 kg N ha−1 as compound NPK-fertilizer (25:2:6), both immediately after the last planting date () and in the following spring (early May). Irrigation was carried out only to ensure rapid establishment after each transplanting. Temperature, rainfall and incident radiation were mostly lower during 1998/1999 than during 1999/2000 (). The ground was mostly frozen and covered with snow from December 1998 until March 1999, but there was no long-lasting frost or snow cover in the winter of 1999/2000.
Table I. Planting dates in 1998 and 1999, plant spacings, plant densities and the number of plants in a central frame used for labelling of tillers and of seed yield determination.
Table II. Temperature, precipitation, and incipient radiation at Landvik during the seed growing seasons 1998–99 and 1999–00, as well as 30-year normal values for temperature and precipitation.
All plant characters were recorded on a 30 cm×30 cm subplot located in the middle of each 1.5 m×1.5 m main plot. Within this central subplot, each newly emerged tiller was tagged, using different coloured wire rings on 15 June, 15 July, 15 August, 2 October, 1 November and 3 May in 1998/99 (no tagging during winter due to frost/snow), and on 18 June, 30 July, 10 September, 10 October, 10 November, 13 January and 28 April in 1999/2000. On the same dates, tillers were recorded as dead if all leaves on the tiller had wilted. On 1 November 1998 and 10 November 1999, at the end of the establishment season, aboveground plant material was cut to a height of 5 cm, removed from the plots, dried, weighed and used to calculate shoot dry weight per tiller.
In the seed harvest year, heading date was recorded for all reproductive tillers when the top of the panicle appeared above the collar of the flag leaf. In addition, stem length was recorded for all reproductive tillers. Within the central subplot, each individual panicle was harvested with a pair of scissors at seed maturity in late July (Festuca pratensis) or early August (Bromus inermis). Panicles tagged on the same date were threshed separately by hand, and the seed yield cleaned on a laboratory air-screen machine (Westrup LALS, Slagelse, Denmark). The seed yield per inflorescence was calculated by dividing total seed yield by panicle number.
Statistical analyses
Separate analyses of variance (PROC GLM; SAS, Citation1990) were performed for each species in each year. The analyses were conducted according to a split plot model with planting date/ plant density (crops A, B or C) as the main factor and tiller emergence periods as subgroups within each main plot. This allowed for a good statistical test of tiller emergence period and the interaction planting date/plant density×tiller emergence period, but comparisons of planting dates/plant densities were hampered by few degrees of freedom. Significant (p% < 5) differences were always identified by Student Neuman Keul multiple comparison tests (SAS, Citation1990). Differences in the fate (reproductive, vegetative or dead) of tillers formed during consecutive periods were evaluated by χ2 homogeneity analyses (PROC FREQ; SAS, Citation1990). Simple correlations coefficients between plant age and tiller density at the end of the establishment season and panicle production and seed yield from tillers produced after the November registration (in the following referred to as late-formed tillers) were calculated by PROC CORR (SAS, Citation1990).
Results
Tiller formation
Regardless of planting date/plant density, the most vigorous formation of new tillers in Bromus inermis occurred during the periods 15 August–2 October 1998 and 10 September–10 October 1999 (). In 1998, this was the case also for Festuca pratensis, but in 1999, crop A and crop B of this species had their main tillering period from 30 July until 10 September (). Tillering after 10 October was generally slow in both species, and in 1999/2000, very few tillers were formed from early November to January, despite the mild winter. By contrast, probably due to resumed tillering in early spring, the density of all crops, and especially of late-established Festuca pratensis, increased from November (1998/1999) or January (1999/2000) to late April/early May. After that, many new tillers were also formed until seed harvest in the first ley year.
Figure 1. Total tiller population and the number of new, dead and reproductive tillers during consecutive periods in seed crops of Bromus inermis established on three different dates and with three different plant densities in 1998–1999 and 1999–2000. a and d: Planted 15 June 1998 or 18 June 1999 at plant spacing 30×30 cm; b and e: planted 15 July 1998 or 30 July 1999 at plant spacing 15×15 cm; c and f: planted 15 August 1998 or 15 September 1999 at plant spacing 7.5×7.5 cm.
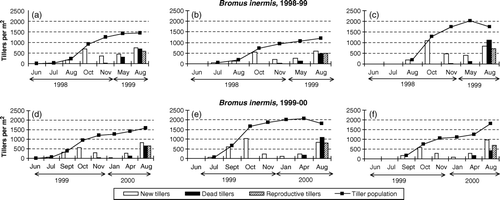
Figure 2. Total tiller population and the number of new, dead and reproductive tillers during consecutive periods in seed crops of Festuca pratensis established on three different dates and with three different plant densities in 1998–1999 and 1999–2000: a and d: Planted 15 June 1998 or 18 June 1999 at plant spacing 30×30 cm; b and e: planted 15 July 1998 or 30 July 1999 at plant spacing 15×15 cm; c and f: planted 15 August 1998 or 10 September 1999 at plant spacing 7.5×7.5 cm.
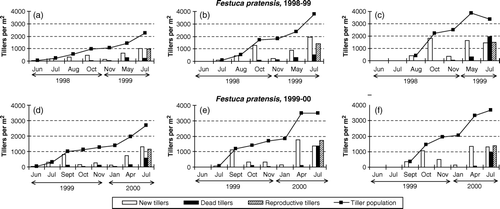
Although higher plant densities were always higher in late than in early plantings, significant differences occurred in tiller density by the end of the establishment season (). In Bromus inermis, the intermediate crop produced the lowest tiller number in 1998/99, but the highest tiller number in 1999/00. In Festuca pratensis, the two late-established crops were significantly denser than the early-established crop in 1998/1999. The average shoot dry weight per tiller at the end of the growing season was always higher in crops established early at a low density than in crops established later at higher densities ().
Table III. Effect of planting date and plant density on tiller density and mean shoot dry weight per tiller at the end of the establishment season (1 November 1998 and 10 November 1999). Means of three and two replicates per treatment in 1998 and 1999, respectively.
Fate of tillers produced during consecutive periods
Regardless of year, planting date/plant density, and species, very few tillers died during the establishment year. Mortality rates during the winter and early spring (April) were mostly higher for early- than for late-established crops ( and ). The highest death rates were always recorded in the seed harvest year, and these rates generally increased with increasing crop density at the onset of the growing season. Crop C of both species in 1998/1999 and crop B of both species in 1999/2000 had a net decrease in total tiller numbers from early May until seed harvest ( and ).
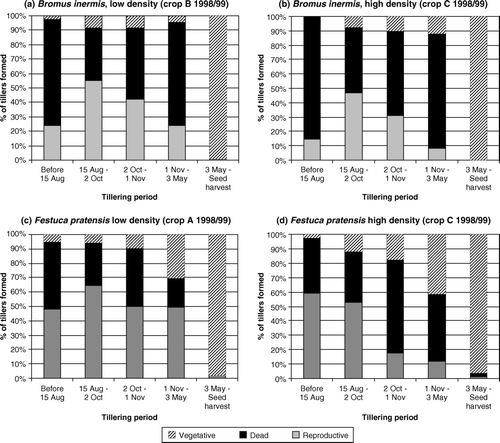
shows the fate of consecutive tiller age groups in crops with the lowest (crop B of Bromus inermis and crop A of Festuca pratensis) and highest (crop C of both species) tiller density during the establishment year 1998/1999. According to χ2 analyses, the proportion of vegetative, reproductive or dead tillers differed significantly (p% < 0.1) among the various age groups. This was the case even if tillers formed after 3 May, which were almost exclusively vegetative, were omitted from the analyses. The figure shows that the chances for tillers, especially those produced from October to May, to become reproductive and contribute to seed yield were higher in low-density than in high-density crops. Except for the high-density crop of Festuca pratensis, tillers formed between 15 August and 2 October always had the highest chance of becoming reproductive. Although a high percentage of the tillers formed in winter/early spring in Festuca pratensis were still vegetative at seed harvest, failure to produce panicles was more often due to tiller death than to tillers remaining vegetative. The high mortality of tillers formed before 15 August in Bromus inermis was usually associated with the elongation of barren stems in the planting year. Moreover, crops A and B of both Bromus inermis and Festuca pratensis typically developed 15–25 inflorescences per m2 even in the planting year; these tillers also contributed to tiller death as they were removed by the end of the growing season.
Figure 3. Percentage of tillers formed in consecutive periods that were reproductive, dead and vegetative in low-density and high-density seed crops of Bromus inermis and Festuca pratensis.
Reproductive tillers
and present the contribution of tillers formed during various periods to the final panicle population in 1999 and 2000, respectively. Reproductive tillers were usually produced earlier in the establishment year in Festuca pratensis than in Bromus inermis, but Festuca pratensis also had a greater percentage of winter/early spring-produced tillers within its reproductive tiller population. As indicated by the significant planting date/plant density×tiller age interactions, the relative contribution of late-formed tillers to the panicle population decreased with later planting/higher planting density for both species in 1998/1999 and for Festuca pratensis in 1999/2000. For Bromus inermis in 1999/2000, the situation was reversed, most probably due to the low density of crop C in this year. On average, for both experimental years and all planting dates/plant densities, 88 and 76% of the reproductive tillers had emerged before early November in Bromus inermis and Festuca pratensis, respectively.
Table IV. Fertile tillers per m2 formed during consecutive periods in seed crops of a) Bromus inermis and b) Festuca pratensis in 1998–99. Percentage of total fertile tiller population is given in parentheses. (crop A: planted on 15 June at 11 plants m−2 ; crop B: planted on 15 July at 44 plants m−2; crop C: planted on 15 Aug at 178 plants m−2 ).
Table V. Fertile tillers per m2 formed during consecutive periods in seed crops of a) Bromus inermis and b) Festuca pratensis in 1999–2000. Percentage of total fertile tiller population is given in parentheses. (crop A: planted on 18 June at 11 plants m−2; crop B: planted on 30 July at 44 plants m−2; crop C: planted on 10 Sep at 178 plants m−2 ).
Days to heading
In both species, late-formed tillers headed later than early-formed tillers. gives average heading dates for the 1999–2000 crops only, but the effects of tiller emergence period were similar in 1998–1999, i.e., about five days delay in heading for tillers produced in spring compared with tillers produced before 15 August. On average, for tiller emergence periods, later planting at higher density caused delayed heading of Bromus inermis by three and five days in 1998/1999 and 1999/2000, respectively, but had no effect on earliness in Festuca pratensis (data not shown). The interactions between plant age/plant density and tiller forming period were not significant.
Table VI. Main effect of tiller forming period on days to heading (after 1 May), stem length and seed weight per panicle in 1999–2000. Means of three planting dates/plant densities in each species.
Stem length
Except for Festuca pratensis in 1998–1999, stems were significantly (9–17 cm) longer on the earliest-established plots (crop A) than on the latest established plots (crop C, data not shown). In Festuca pratensis, stem length decreased consistently with tiller emergence period, but in Bromus inermis, tillers formed after 28 April developed longer stems than tillers produced in late autumn (10 October–10 November) ().
Seed weight per inflorescence
In Festuca pratensis inflorescences from late-formed tillers invariably produced less seed than inflorescences from early-emerging tillers (). In Bromus inermis tillers that had emerged before 10 October produced heavier inflorescences than tillers that had emerged between 10 November and 13 January or after 28 April. The main effects of planting date/ plant density on inflorescence weight were not significant, although there were often tendencies for crop C to produce lighter inflorescences than crops A and B (data not shown).
Seed yield
Regardless of planting date/plant density, most of the seed yield of Bromus inermis in 1999 () and 2000 () was produced on tillers that had emerged in late August and September the previous year. In both years, seed yields of Festuca pratensis were distributed among several age groups in crop A, but concentrated on tillers produced in August and September in crop B and, most notably, in crop C ( and ).
Table VII. Seed yield (kg ha−1) from tillers formed during consecutive periods in seed crops of a) Bromus inermis and b) Festuca pratensis in 1998–1999. Percentage of total seed yield is given in parentheses. (crop A: planted on 15 June at 11 plants m−2; crop B: planted on 15 July at 44 plants m−2; crop C: planted on 15 Aug at 178 plants m−2 ).
Table VIII. Seed yield (kg ha−1) from tillers formed during consecutive periods in seed crops of a) Bromus inermis and b) Festuca pratensis in 1999–2000. Percentage of total seed yield is given in parentheses. (crop A: planted on 18 June at 11 plants m−2; crop B: planted on 30 July at 44 plants m−2; crop C: planted on 10 Sep at 178 plants m−2 ).
Correlations
While inflorescence production from late-formed tillers in Bromus inermis was negatively correlated with plant density the previous autumn (), plant age exerted minor influence on both panicle number and seed yield in this species. In Festuca pratensis, plant age and tiller density in November were mutually correlated and their relationships with seed yield from late-formed tillers were therefore of the same magnitude. Plant age was, however, more influential than tiller density on the number of panicles emanating from spring-formed tillers.
Table IX. Simple correlation coefficients between plant age and tiller density as determined 1 Nov 1998 and 10 Nov 1999, and panicles numbers and seed yield emanating from tillers produced after these dates. Lower left (in italics): Bromus inermis ; upper right: Festuca pratensis. n = 15.
Discussion
The tiller density of early-planted seed crops increased continuously from planting to seed harvest in both species ( and ). However, when the density in April/May reached about 2000 tillers per m2 in Bromus inermis and 3500 tillers per m2 in Festuca pratensis (crop C in 1998/1999 and crop B in 1999/2000), tiller numbers tended to decrease in early summer. A similar decline has previously been described in Festuca pratensis (Langer & Lambert, Citation1959), Bromus inermis (Lamp, Citation1952), and several other grasses (Lambert, Citation1967; Colvill & Marshall, Citation1984). In Phleum pratense, Aamlid (Citation1997) observed a net loss of tillers in May and June only for crops with more than 1200 tillers per m2 in April. The high mortality during tiller elongation in spring/early summer was probably due to an increased demand for nutrients and carbohydrates during stem elongation, accompanied by mutual shading within the canopy (e.g., Mitchell, Citation1953). New tillers which have not yet developed their own adventitious root system and at least three to four leaves are the ones most vulnerable to intertiller competition (Williams, Citation1964; Ong, Citation1978). Consequently, it has previously been observed that tillers formed shortly after establishment contribute a greater percentage of the panicle population in dense than in open seed crops (e.g., Meijer, Citation1984; Aamlid, Citation1996). This is in agreement with the present trials, where as many as 71% of the tillers in Bromus inermis and 42% of tillers in Festuca pratensis that had been tagged in April/May died before seed harvest.
As expected, most of the tillers in Bromus inermis developed into elongated sterile stems after being exposed to long summer days in the year of establishment. Although the crops were mown in early November, very few of these tillers survived and contributed to seed yield ( and ; ). The high mortality of the early-emerging tillers of Bromus inermis supports the findings by Lamp (Citation1952) who studied seed production in several different clones of the same species.
In both species the majority of the reproductive tillers had been formed the previous autumn. This is in agreement with earlier studies in Festuca pratensis (Langer & Lambert Citation1959, Odgaard, Citation1968; Havstad, Citation1998) and Bromus inermis (Lamp, Citation1952). However, in 1998/1999, as much as one-third of the panicles in the early-established/low-density crop A of Festuca pratensis emanated from tillers that had emerged after snow melt in March (b). This result is hard to reconcile with Heide's (Citation1988, Citation1994) description of Festuca pratensis as a species with an extreme (up to 16 weeks) requirement for primary induction and with our earlier findings that little or no intertiller transfer of flower induction stimuli occurred in this species. In agreement with Havstad (Citation1996) who found tillers of Festuca pratensis to have no juvenile stage, one explanation might be that these late-formed tillers were induced already at the bud stage, i.e., while still in the sheaths of their parent tiller (Kleinendorst, Citation1974). However, there are also more recent studies suggesting that the primary induction requirement in Festuca pratensis is not as obligate and extreme as thought previously (Aamlid, Citation2005); at least, there seems to be considerable genetic variation for this character (Fang et al., Citation2004; Ergon et al., Citation2006). As our previous studies were conducted in growth chambers with much lower light intensities than under field conditions (Havstad et al., Citation2004) it might also be that each individual tiller in those studies did not have sufficient resources to sustain the reproductive development of daughter tillers.
While the panicle production capacity of late-formed tillers in our previous experiments with Festuca pratensis seems to be underestimated, the opposite might well be correct for Bromus inermis. In that species, Havstad et al. (Citation2004) found tillers emerging after primary induction to contribute as much as 40% of the total panicle population, as opposed to a maximum of 18% in the present study. The explanation for this discrepancy is probably that the growth chamber studies were conducted with temperatures in the range 9–15°C, which is optimal for short-day induction of Bromus inermis (Heide, Citation1984) but much higher than field temperatures in April in the seed production district of Norway ().
In the present experiment, the effect of plant age on panicle development from late-formed tillers is confounded with the effect of tiller density and, thus, competition level within the various crops. It is, however, interesting to observe that tiller density, but not plant age, was significantly correlated with panicles emanating from late-formed tillers in Bromus inermis, whereas plant age exerted a greater influence in Festuca pratensis (). This can be further substantiated by, in , comparing crops A and C of Bromus inermis (both of similar low tiller density) and crops B and C of Festuca pratensis (both of high tiller density). While, in Bromus inermis, late-formed tillers had a similar chance of becoming reproductive irrespective of whether their parent plants had been planted on 18 June or 10 September, late-formed tillers in Festuca pratensis were more likely to become reproductive if connected to a parent planted on 30 July than on 10 September.
Besides determining whether a tiller becomes reproductive or not, tiller emergence period had a major impact on seed yield per inflorescence (). This is in agreement with Ryle and Langer (Citation1963) who explained inflorescence size as a function of meristem size at the time of spikelet initiation. shows a small variation in heading date for the various tiller classes, and provided this was the case also for initiation date, it is reasonable to assume that the number of primary branches at the onset of reproductive development was directly related to tiller age. As a result of more seeds per inflorescence, tillers formed early (August and September) made a greater relative contribution to seed yield ( and ) than to total panicle population ( and ) in both species.
In conclusion, the results presented for Festuca pratensis in and confirm our first hypothesis–namely, that tillers emerging in winter and spring make a greater contribution to yield in seed crops established early at a low plant density than in crops established later at a higher density. For Bromus inermis, the conclusion is less clear, as tiller density by the end of the growing season seems to have a greater impact than plant age on the reproductive behaviour of late-formed tillers (). Our second hypothesis, that late-formed tillers are more important to seed yield in Bromus inermis than in Festuca pratensis, has to be rejected as the data suggest it is the other way round, at least for well established plants.
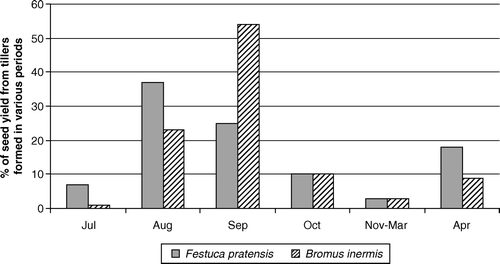
When not using a cover crop, it is usually recommended to establish Norwegian seed crops of Bromus inermis and Festuca pratensis at a seeding rate of ca. 5 kg ha−1 during the first two weeks of July (Jonassen & Hillestad, Citation1990; Jonassen, Citation1994). On the condition that this gives an optimal plant density of ca. 100 plants m−2 (Nordestgaard, Citation1975a), presents a generalized chart of the contribution to seed yield that can be expected from tillers formed during consecutive periods in such crops.
Figure 4. Generalized pattern of the contribution to seed yield of tillers formed during various periods in seed crops of Bromus inermis and Festuca pratensis sown without cover crop during the first two weeks of July in SE Norway.
Acknowledgements
This research was funded by grant no. 121708 from the Norwegian Research Council. We thank Anne A. Steensohn and Åge Susort for excellent technical assistance.
References
- Aamlid , T.S. 1994 . Pure stand establishment of Poa pratensis L. for seed production: sowing rates and sowing methods . Norwegian Journal of Agricultural Sciences , 8 : 277 – 290 .
- Aamlid , T.S. 2005 . A comparison between primary induction requirements in vegetatively propagated ramets and seedlings of Festuca pratensis and Festuca rubra . Acta Agriculturæ Scandinavica , 55 : 299 – 303 .
- Aamlid , T.S. 1996 . Autumn growth and seed-producing capacity of tillers of Poa pratensis ‘Lavang’ and ‘Leikra’ . Norwegian Journal of Agricultural Sciences , 10 : 583 – 592 .
- Aamlid , T.S. 1997 . Nitrogen and moisture inputs to seed crops of timothy (Phleum pratense L.). II. Split applications of nitrogen in the seed harvest year . Journal of Applied Seed Production , 15 : 5 – 16 .
- Aamlid , T.S. & Jonassen , G.H. ( 2006 ). Seed production of a forage and a lawn cultivar of common bent (Agrostis capillaris) as affected by sowing rate . Acta Agriculturae Scandinavica ( Accepted, in press. )
- Colvill , K.E. and Marshall , C. 1984 . Tiller dynamics and assimilate partitioning in Lolium perenne with particular reference to flowering . Annals of Applied Biology , 104 : 543 – 557 .
- Ergon , Å. , Fang , C. , J⊘rgensen , Ø. , Aamlid , T.S. and Rognli , O.A. 2006 . Quantitative trait loci controlling vernalization requirement, heading time and number of panicles in meadow fescue (Festuca pratensis Huds.) . Theoretical and Applied Genetics , 112 : 232 – 242 .
- Fang , C. , Aamlid , T.S. , J⊘rgensen , Ø. and Rognli , O.A. 2004 . Phenotypic and genotypic variation in seed production traits within a full-sib family of meadow fescue (Festuca pratensis Huds.) . Plant Breeding , 123 : 241 – 246 .
- Fulkerson , R.S. 1959 . The effects of seeding rate and row width in relation to seed production in orchard grass, Dactylis glomerata L . Canadian Journal of Plant Science , 39 : 355 – 363 .
- Havstad , L.T. 1996 . Juvenility and flowering in Festuca pratensis Huds. 1. Effects of plant age, cultivar and duration of primary induction treatments . Norwegian Journal of Agricultural Sciences , 10 : 159 – 178 .
- Havstad , LT. 1998 . “ Vegetative development and flowering in meadow fescue (Festuca pratensis Huds.) as affected by plant size and cultivars ” . In Seed production. NJF-report no 121. Seminar no 284 , 68 – 75 . Norway : Sandefjord and Landvik .
- Havstad , L.T. , Aamlid , T.S. , Heide , O.M. and Junttila , O. 2003 . Transfer of florigenic stimuli between tillers in photoperiodically split plants of Dactylis glomerata and Bromus inermis . Physiologia Plantarum , 118 : 270 – 277 .
- Havstad , L.T. , Aamlid , T.S. , Heide , O.M. and Junttila , O. 2004 . Transfer of flower induction stimuli to non-exposed tillers in a selection of temperate grasses . Acta Agriculturæ Scandinavica Section B Soil and Plant Science , 54 : 23 – 30 .
- Heide , O.M. 1984 . Flowering requirements in Bromus inermis, a short-long-day plant . Physiologia Plantarum , 62 : 251 – 256 .
- Heide , O.M. 1988 . Flowering requirements of Scandinavian Festuca pratensis . Physiology Plantarum , 74 : 487 – 492 .
- Heide , O.M. 1994 . Control of flowering and reproduction in temperate grasses . New Phytologist , 128 : 347 – 362 .
- Jonassen , G.H. ( 1980 ). Effect of low plant density on seed yield of meadow fescue (Festuca pratensis Huds.), cocksfoot (Dactylis glomerata L.), red fescue (Festuca rubra L.) and common bent (Agrostis tenuis Sibth.) . Forskning og fors⊘k i landbruket , 31 , 187 – 196 . ( In Norwegian, with English abstract ).
- Jonassen , G.H. ( 1994 ). Establishment of smooth bromegrass (Bromus inermis Leyss.) seed crops with or without cover crop, and on different dates without cover crop . Norsk landbruksforsking , 9 , 51 – 57 . ( In Norwegian, with English abstract ).
- Jonassen , G.H. & Hillestad , R. ( 1990 ). Etablering av fr⊘eng uten dekkvekst . In: Fr⊘avl. NJF seminarium nr 173. Tune Landboskole, Denmark June 18–20 1990 . (pp. 84 – 93 ). ( In Norwegian ).
- Jonassen , G.H. & Torskenæs , E. ( 1994 ). The effects of seeding rates on seed production of smooth bromegrass (Bromus inermis Leyss.) Norsk landbruksforsking , 8 , 315 – 321 . ( In Norwegian, with English abstract ).
- Kleinendorst , A. 1974 . Some effects of vernalization on the reproductive capacity of Lolium perenne L . Netherlands Journal of Agricultural Science , 22 : 6 – 21 .
- Lambert , D.A. 1967 . Competition between plants of cocksfoot (Dactylis glomerata) grown for seed . Journal of the British Grassland Society , 4 : 274 – 279 .
- Lamp , F. 1952 . Reproductive activity in Bromus inermis in relation to phases of tiller development . Botanical Gazette , 113 : 413 – 438 .
- Langer , R.H.M. and Lambert , D.A. 1959 . Ear-bearing capacity of tillers arising at different times in herbage grasses grown for seed . Journal of the British Grassland Society , 14 : 137 – 140 .
- Meijer , W.J.M. 1984 . Inflorescence production in plants and in seed crops of Poa pratensis L. and Festuca rubra L. as affected by juvenility of tillers and tiller density . Netherlands Journal of Agricultural Science , 32 : 119 – 136 .
- Mitchell , K.J. 1953 . Influence of light and temperature on the growth of ryegrass (Lolium spp.). I. Pattern of vegetative development . Physiologia Plantarum , 6 : 21 – 46 .
- Nordestgaard , A. ( 1975a ). Seed rates of meadow fescue (Festuca pratensis Huds.) for seed production . Tidsskrift for Planteavl , 79 , 417 – 428 . ( In Danish, with English abstract ).
- Nordestgaard , A. ( 1975b ). Seed rates of timothy (Phleum pratense) for seed production . Tidsskrift for Planteavl , 79 , 433 – 445 . ( In Danish, with English abstract ).
- Nordestgaard , A. ( 1977 ). Seed rates of perennial ryegrass (Lolium perenne L.) for seed production . Tidsskrift for Planteavl , 81 , 315 – 324 . ( In Danish, with English abstract ).
- Nordestgaard , A. ( 1979 ). Seed rates of cocksfoot (Dactylis glomerata) for seed production . Tidsskrift for Planteavl , 83 , 111 – 122 . ( In Danish, with English abstract ).
- Odgaard , P. ( 1968 ). Forel⊘bige resultater fra unders⊘gelser over skuddannesle hos græsser til fr⊘avl . In: Referat af indlæg ved 2. nordiske seminar vedr⊘rende fr⊘avlsfors⊘g . Årslev , Danmark . (pp. 43 – 56 ). ( In Danish ).
- Ong , C.K. 1978 . The physiology of tiller death in grasses. 1. Influence of tiller age, size and position . Journal of the British Grassland Society , 33 : 197 – 203 .
- Ryle , G.J.A. and Langer , R.H.M. 1963 . Studies on the physiology of flowering of timothy (Phleum pratense L.). II. Influence of temperature and day length on the size of the inflorescence . Annals of Botany , 27 : 233 – 244 .
- SAS . 1990 . SAS/STAT User's Guide. Version 6 , Cary, NC, USA : SAS Institute .
- Williams , R. D. 1964 . Assimilation and translocation in perennial ryegrass . Annals of Botany , 28 : 419 – 425 .