
Abstract
Two aroma compounds of volatile thiols, 2-furanmethanethiol (2FM) and ethyl 2-mercaptopropionate (ET2MP), were formed in five types of Japanese soy sauce during fermentation by yeast. The concentrations of 2FM and ET2MP in the soy sauce samples increased during alcoholic fermentation. The concentrations of 2FM and ET2MP were higher in the soy sauce fermented by Zygosaccharomyces rouxii than in that fermented by Candida versatilis. The enantiomers of ET2MP were separated by gas chromatography in a capillary column. The average enantiomeric ratio of ET2MP in the soy sauce was approximately 1:1. 2FM was formed by yeast in a medium prepared from cysteine and furfural, and cysteine is considered the key precursor of 2FM by yeast in soy sauce.
Graphical Abstract
Two important aroma compounds of volatile thiols, 2-furanmethanethiol etc., were formed in Japanese soy sauce during fermentation by yeast.
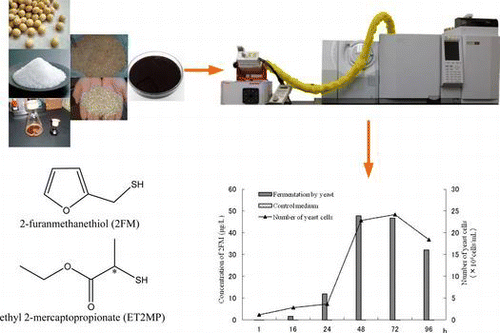
The aroma of soy sauce is one of the most important factors determining its quality and marketability to consumers. More than 300 flavor components have been confirmed in Japanese soy sauce.Citation1,Citation2) 4-Hydroxy-2(or 5)-ethyl-5-(or 2)-methyl-3-(2H)-furanone (HEMF) has a strong, sweet, caramel-like aroma that is very important.Citation3,Citation4) We have reported the mechanism of HEMF by yeast during the aging of moromi,Citation5–Citation7) and we consider that there is likely to be another important compound of soy sauce in the unidentified odorant formed by the Maillard reaction and/or yeast. We have reported the identification of 2-furanmethanethiol (2FM), benzenemethanethiol, and Ethyl 2-mercaptopropionate (ET2MP) in raw soy sauce and heat-treated soy sauce, and we reported that these volatile thiols were very important aroma components of heat-treated soy sauce.Citation8)
2FM, which has an intense, roasted, coffee-like aroma, was first identified in roasted coffee. It has been reported to be an important aroma component in such foods as roasted meatCitation8–Citation10) and white wine.Citation11) The odor threshold was reported to be 0.4 ng/L in a model hydro-alcoholic solution by Tominaga et al.Citation12) Shibamoto and RussellCitation13) have reported the formation of 2FM from furural in the presence of cysteine by an amino-carbonyl reaction, and Blanchard et al.Citation14) have reported that 2FM was formed from furfural in white wine by yeast. Ohata et al.Citation15) have reported that 2FM is a very important component of miso aroma, and that 2FM can be generated by yeast during the fermentation process. We think that the formation of 2FM in raw soy sauce is related to the alcoholic fermentation of yeast, because 2FM was identified in raw soy sauce in our previous study.Citation8)
ET2MP is a compound used in flavor chemistry. It has a tropical fruit-like aroma. It has been identified in cashew apple juice,Citation16) strawberries, and camembert cheese.Citation17) ET2MP can be formed by yeast during the alcoholic fermentation of soy sauce, as it has been identified in raw soy sauce.Citation8) ET2MP clearly has two different enantiomers due to one asymmetrical carbon atom at position 2. Separation of these two enantiomers of ET2MP is very important in studying the mechanism of ET2MP in soy sauce, because only one of the enantiomers is formed by yeast.
Many thiols have been isolated from cooked foods as important aroma compounds due to their odors, and low odor thresholds.Citation18,Citation19) Many thiols have been formed by heating,Citation20) but yeast must play an important role in the formation of 2FM and ET2MP in soy sauce.
Materials and methods
Materials and chemicals
Three types of raw soy sauce and corresponding heat-treated samples were analyzed in order to separate the enantiomers of ET2MP. The samples were provided by the Experimental Station of Miyagi Miso-Shoyu Industy Cooperative (Miyagi, Japan).
2FM, cysteine, and dichloromethane were purchased from Sigma–Aldrich (Tokyo). ET2MP was purchased from Tokyo Chemical Industry (Tokyo), and 4-methoxy-2-methyl-2-mercaptobutane (an internal standard) was purchased from Oxford Chemicals (Bound Brook, NJ). d-Glucose, sodium chloride, potassium dihydrogenphosphate, magnesium sulfate heptahydrate, 2-furfural, and methionine were purchased from Kanto Chemical (Tokyo). Yeast extract was purchased from Becton, Dickinson (Bound Brook, NJ), and protease, glutaminase, pectinase, α-amylase, and glucoamylase were purchased from Amano Enzyme (Nagoya, Japan).
Micro-organisms
Zygosaccharomyces rouxii RS1, C. versatilis, and Tetragenococcus halophilus, provided by Iwate Industrial Research Institute, were chosen for the preparation of soy sauce samples. Z. rouxii 061 was used to confirm the formation of 2FM. This yeast has the ability to form large amounts of HEMF as an aroma compound in soy sauce. It was provided by the Experimental Station of Miyagi Miso-Shoyu Industry Cooperative (Miyagi, Japan).
Soy sauce samples
We focused on the formation of 2FM and ET2MP by yeast in this study. A mixture of koji and saline solution was heated to inactivate enzymes and micro-organisms. Six types of soy sauce samples were prepared under various micro-organism conditions in six tanks. A mixture of 2.4 kg of koji and 3.6 L of a boiled saline solution (22.5%, w/w) was heated to 80 °C and then cooled with water to 22 °C, except for one type of soy sauce sample. Table shows the fermentation conditions for the six types of soy sauce samples. Type I was prepared as control from the mixture without sterilizing in order to make evident the generation of volatile thiols by Aspergillus. Soy sauce types II–VI were prepared from the sterilized mixtures. As Table shows, enzymes (protease, glutaminase, pectinase, α-amylase, and glucoamylase), which had the same activity in koji of type I, were added to prepare soy sauce types II–VI. Z. rouxii RS1, C. versatilis, and T. halophilus were added to prepare type II, Z. rouxii RS1 was added to prepare type III, C. versatilis was added to prepare type IV, and T. halophilus was added to prepare type V. No micro-organism was added to prepare type VI as control. All of the soy sauce samples were fermented at 25 °C for 15 weeks, and then were pasteurized by heating at 85 °C for 30 min. The water-soluble nitrogen content, pH value, and glucose concentration for each soy sauce were evaluated by the standard method for soy sauce analysis to determine the normal fermentation of these samples without any microbial contamination.
Table 1. Fermentation conditions for the soy sauce samples.
Analysis of volatile thiols in the soy sauce samples
2FM and ET2MP in the six types of soy sauce samples after fermentation for 0, 2, 4, 6, 8, 11, 15 weeks and in the heat-treated soy sauce samples after ferments 15 weeks were assayed by the method described in our previous paper.Citation8) Three analyses were carried out on each sample. The significance of volatile thiols concentrations in heat-treated types I–VI were assessed by Student’s t test.
ET2MP enantiomers in the soy sauce
The ET2MP enantiomers were assayed with a Shimadzu GC–MS 2010 instrument equipped with a column of InertCap CHIRAMIX (0.25 mm × 30 m, 0.25 μm film thickness). The oven was programmed at 55 °C for 1 min, rising to 100 °C at 0.7 °C/min. The mass spectrometer was operated at EI 70 eV in SIM mode, with selection for ions of m/z 61, 88, and 134.
Olfactory descriptors of the enantiomers
A descriptive analysis of the ET2MP enantiomers was conducted by gas chromatography–olfactometry (GC–O) with an ET2MP standard alcoholic solution. The concentration of this solution was determined to be 78.6 μg/L, which was the maximum level of ET2MP in the heat-treated soy sauce samples in our previous study, by 5,5′-dithio-bis (2-nitrobenzoic acid) by the method of Tominaga and DubourdieuCitation21) The ET2MP standard solution was diluted stepwise with ethanol from 1:2 to 1:16, and 2 μL of each diluted solution was analyzed with a Shimadzu GC-17A instrument equipped with an InertCap CHIRAMIX column. The oven was programmed for 70 °C for 1 min, rising to 100 °C at 0.7 °C/min. GC–O was performed thrice for each sample by four panelists who responded with odor descriptions of the ET2MP enantiomers.
Media for the formation of 2FM
The medium for the starter culture was prepared from 5 mL of raw soy sauce, 2.5 g of glucose, and 5 g of sodium chloride dissolved in 50 mL of ultra-pure water.Citation7) The sample base mediumCitation5) consisted of 5 g of glucose, 10 g of sodium chloride, 1 g of potassium dihydrogenphosphate, 0.5 g of magnesium sulfate heptahydrate, and 0.5 g of a yeast extract, all in 100 mL of ultra-pure water adjusted to pH 5.2 with sodium hydroxide. Each medium was sterilized after it was prepared.
Z. rouxii 061 was incubated at 27 °C for 3 d in the medium for the starter culture, and then yeast cells were collected. The collected cells were washed with a buffer containing 10 mL of 1 M Tris–hydrochloric acid at pH 7.4, 2.48 g of sodium chloride, and 0.123 g of magnesium sulfate heptahydrate dissolved in distilled water made up to 500 mL with distilled water and sterilized. The base medium contained 9 mM (87.6 mg/100 mL) furfural and 10 mM cysteine as precursor compounds for 2FM. The concentration of the yeast in the base medium was adjusted to 106 cells/mL, and then the base medium was fermented at 27 °C.
In order to clarify the relationship between the formation of 2FM and the concentrations of the precursor compounds, media were prepared with various concentrations of furfural and cysteine. The base medium containing 9 mM furfural was prepared with and without increasing quantities of cysteine (5 mM and 10 mM). The base medium, containing 10 mM cysteine, was prepared with increasing quantities of furfural (4.5 mM and 9 mM). Control media that did not contain yeast were obtained by adding the corresponding concentrations of furfural and cysteine. Aroma concentrates were prepared from the supernatants of media that had been centrifuged to remove yeast cells. The concentration of 2FM in each medium was assayed as described above for the soy sauce samples. Two analyses were carried out on each medium, and two replicate experiments were performed for each medium.
Results and discussion
Quantification of 2FM and ET2MP in the soy sauce samples
Evaluation of the pH values, water-soluble nitrogen contents, and glucose concentrations in the soy sauce samples during the fermentation process indicated that five types (I, II, III, IV, and VI) had been successfully fermented, while type V was contaminated by micro-organisms during fermentation (data are not shown). Thus it was impossible to consider the relationship between the formation of 2FM or ET2MP and the lactic acid bacteria.
The concentrations of 2FM in the five types of soy sauce samples during the fermentation process are shown in Table . The changes in the concentration of 2FM in types I and VI were similar during fermentation. 2FM is generated in foods during heating by the Maillard reaction. No micro-organism was added to type VI after sterilization, and hence we think that the presence of 2FM in type VI was generated only by a slow Maillard reaction process. We also think that 2FM in type I was likewise generated by a Maillard reaction, and that Aspergillus is unlikely to have played a role in the formation of 2FM in the soy sauce.
Table 2. Concentrations of 2FM and ET2MP in the five types of soy sauce during fermentation (μg/L)Table Footnotea.
The concentrations of 2FM in types II and III were higher than in types I and VI. It appears that 2FM was not only generated by a slow Maillard reaction, but also by yeast during the fermentation of these soy sauce types. The quantity of 2FM generated by Z. rouxii RS1 was higher than by C. versatilis, since the concentration of 2FM in type III was higher than in type IV. In addition, the same level of 2FM concentration was generated in types II and III. These results suggest that Z. rouxii RS1 played a major role in the formation of 2FM during the fermentation of the soy sauce.
The concentrations of ET2MP in the five types of soy sauce during fermentation are shown in Table . The formation of ET2MP in type I started from the sixth week. No yeast or Aspergillus was added to type VI, and no ET2MP was generated at any time during fermentation. The concentrations of ET2MP in types II, III, and IV increased during fermentation. The increased ET2MP concentration in type II was similar to that in type III, while the concentration of ET2MP in type III was higher than that in type IV. We assume that Z. rouxii RS1 played a major role in ET2MP formation during soy sauce fermentation.
After the fermentation process, the five types of soy sauce samples were pasteurized under the same conditions as used in the production process for commercial soy sauce (heating at 85 °C for 30 min). The concentrations of 2FM and ET2MP in each type of soy sauce were increased by heat treatment (Table ), although 2FM and ET2MP were not formed during heat treatment of the koji mixture for pasteurization before fermentation. The results of the t test suggested that the 2FM concentrations in types I–VI were significantly different from each other, except for types II and III, types IV and VI. The concentrations of 2FM in heat-treated types II and III were significantly higher than those in types I and VI (p < 0.05). In addition, the 2FM concentration in type III was significantly higher than that in type IV (p < 0.01). The assessment results for ET2MP concentrations in types I–VI were almost identical to those of 2FM concentrations. These results suggest that precursor compounds of 2FM and ET2MP were formed during fermentation by yeast, and that their concentrations increased during heating. Hence, we think the fermentation played a very important role in the formation of 2FM and ET2MP.
ET2MP enantiomers in the soy sauce
Separating the enantiomers of ET2MP in the soy sauce was important in order to confirm the formation of ET2MP by yeast, because yeast was likely to form only one of the enantiomers.
Fig. shows the GC–MS trace for the aroma concentrate from heat-treated soy sauce by analysis in the InertCap CHIRAMIX column. The two enantiomers of ET2MP were separated successfully. Table shows the average enantiomeric ratio of ET2MP in the raw soy sauce and the heat-treated soy sauce, which was confirmed to be about 1:1.
Fig. 1. Chromatographic peaks of ET2MP enantiomers from a heat-treated soy sauce in SIM mode produced by overlapping selected ions (m/z 61, 88, and 134).
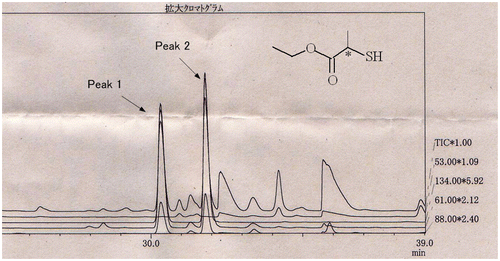
Table 3. Enantiomeric ratioTable Footnotea of ET2MP in three samples of soy sauce.
We could not directly determine the configuration of the ET2MP molecule, but the odor quality of the enantiomers was evaluated with a racemic mixture of commercial ET2MP. This mixture solution exhibited a tropical fruit-like aroma, and the odor quality of the enantiomers changed when the standard solution was diluted, as shown in Table . Both enantiomers exhibited a rubber-like note at a concentration of 78.6 μg/L, but peak 1 exhibited a pineapple-like note and peak 2 an onion-like note when the standard solution was diluted to 9.8 μg/L. Our previous study found the average level of ET2MP in heat-treated soy sauce to be about 20 μg/L.Citation8) Hence, we conclude that one of the ET2MP enantiomers contributed a fruity note and the other one a sesame-like note to the aroma of the heat-treated soy sauce.
Table 4. Olfactory descriptors of the ET2MP enantiomers.
Ethyl esters are usually produced by yeast metabolism during the fermentation of moromi.Citation22) We think that the formation of ET2MP in soy sauce is related to yeast fermentation, but the average enantiomeric ratio in the soy sauce was approximately 1:1. Although yeast played an important role in the formation of ET2MP in the soy sauce, the role of yeast was considered to be only part of the story. It was therefore necessary to do further experiments, in order to clarify the mechanism of the formation of ET2MP by yeast in soy sauce.
Formation of 2FM by yeast
Furfural is a major dehydration product of pentose by the Maillard reaction. Furfural has also been reported to play an important role in the formation of 2FM in that reaction,Citation23,Citation24) but 2FM is an unstable compound that readily oxidizes to a disulfide.Citation25) Kumazawa and MasudaCitation26) have reported that the residual ratio of 2FM in a citric/Na2HPO4 buffer solution decreased with increasing pH values after heating. In addition, Muench et al.Citation27) have reported that the formation concentration of 2FM from glucose and cysteine showed a maximum at pH 5.0. Huynh-Ba et al.Citation28) have reported that baker’s yeast catalyzed the transformation of a cysteine–furfural conjugate into 2FM at pH 8.0, while Blanchard et al.Citation14) have reported that 2FM was formed by yeast from furfural and cysteine in white wine at pH 3.5. A pH value of 4.7 for soy sauce is therefore advantageous for generating 2FM by yeast during alcoholic fermentation, and the results of our experiments with soy sauce samples confirmed that 2FM can be generated by yeast. We think that furfural is formed during the heat treatment of soybeans and wheat used for making koji.
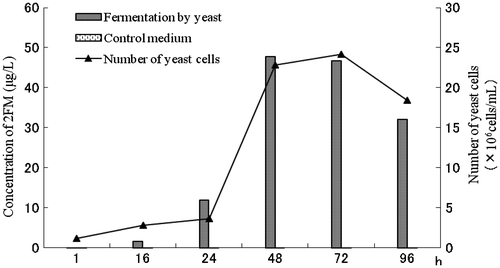
The concentrations of 2FM in the media during fermentation are shown in Fig. . The concentration of 2FM increased to about 50 μg/L until the yeast cells increased to a level of 107 cells/ml. The 2FM concentration in the control medium, which did not contain yeast, was only 0.2 μg/L, and is considered to have been formed by the Maillard reaction. These results suggest that furfural and cysteine were the precursor compounds of 2FM, and that the role of yeast in 2FM formation during fermentation was very important.
Fig. 2. Formation of 2FM from furfural and cysteine in the fermentation and control medium.
Note: Standard deviation over the average concentration was ±0.1 μg/L.
Methionine and furfural were added to the medium for fermentation, but no 2FM was detected (data not shown), indicating that methionine was not a precursor in the biogeneration of 2FM.
Key precursor compound for 2FM generation
The concentrations of 2FM formed during fermentation were considerably affected by the amounts of precursor compounds added to the medium. The effectiveness of furfural and cysteine as precursor compounds of 2FM was hence compared according the concentration of 2FM in media prepared with differing amounts of furfural or cysteine.
As Table shows, 2FM was not be detected in the presence of only a single precursor compound in fermentation medium. This result suggests that both the thiol group of cysteine and the carbon group of furfural were necessary for the generation of 2FM by yeast. The maximum concentration of 2FM was 47.7 μg/L when 10 mM cysteine and 9 mM furfural were added to the fermentation medium, but the maximum concentration of 2FM decreased to 9.5 μg/L when 10 mM cysteine and 4.5 mM furfural were added to the fermentation medium, while the concentration of 2FM decreased to 2.8 μg/L when 5 mM cysteine and 9 mM furfural were added to it. These results suggest that cysteine played a more important role than furfural in generating 2FM in the fermentation medium. No 2FM detected in the control medium without yeast when 10 mM cysteine and 4.5 mM furfural were added, but 2FM was detected when 5 mM cysteine and 9 mM furfural added. Hence, we conclude that furfural played a more important role than cysteine in the control medium without yeast in generating 2FM by the Maillard reaction. These results imply that the formation mechanism for 2FM by yeast was different from the Maillard reaction.
Table 5. Influence of furfural (FL) and cysteine on the amount of 2FM (μg/L) in the fermentation medium.
According to a study by Huynh-Ba et al.Citation28), the yield of 2FM from a cysteine–furfural conjugate by β-(C-S) lyase was zero at under pH 6.0. We think that the generation of 2FM in our media was not related to β-(C-S) lyase, because all of the media were adjusted to pH 5.2 before fermentation, and the average pH value was 4.6 at the end of fermentation. Blanchard et al.Citation14) have reported that the quantity of 2FM produced was proportional to the amount of cysteine added to the medium. Our results for the generation of 2FM in the soy sauce media are in accord with the data of Blanchard et al. The concentration of 2FM in our media was greatest at a high concentration of cysteine. Hence, we conclude that thiol groups were transferred from cysteine by metabolism with yeast, and that 2FM was generated by a reaction between furfural and the thiol group. It is necessary to clarify the role of yeast in the reaction between the carbonyl group of furfural and the thiol group in further study.
In conclusion, 2FM and ET2MP were generated in the fermentation of five types of soy sauce. The concentrations of 2FM and ET2MP in the soy sauce samples fermented by yeast (Z. rouxii RS1 and C. versatilis) were distinctly higher than in the soy sauce fermented by Aspergillus or in the control soy sauce sample to which no micro-organisms were added. Hence, we conclude that yeast played an important role in the formation of 2FM and ET2MP in the soy sauce.
The optical isomers of ET2MP were separated by GC–MS analysis in order to clarify the role of yeast in ET2MP formation. We could not directly determine the configuration of the ET2MP molecule, but the odor quality of the enantiomers was determined with a commercial ET2MP racemic mixture. One of the enantiomers exhibited a fruity note, and the other one, a sesame-like note. The average enantiomeric ratio of ET2MP in the soy sauce was approximately 1:1. Thus, although yeast played an important role in the formation of ET2MP in the soy sauce, it was not the only factor.
Furfural and cysteine as precursor compounds of 2FM were added to the medium for fermentation by yeast. 2FM was generated in the fermentation medium, but could not be detected in the control medium without yeast. Thus, furfural and cysteine were important precursors in the biogeneration of 2FM by yeast. The concentration of 2FM in the fermentation medium increased with increasing concentrations of cysteine. Hence, we conclude cysteine is the key precursor in the biogeneration of 2FM by the reaction between furfural and the thiol group formed by cysteine metabolism of yeast.
Acknowledgments
We express our thanks to Dr Motoko Ohata and Dr Eisuke Takahisa for their advice and contributions to this study. We are indebted to the Experimental Station of the Miyagai Miso-Shoyu Industrial Cooperative for kindly providing soy sauce samples.
Related Research Data
References
- Yokotsuka T, Sasaki M. Fermented protein foods in the Orient: shoyu and miso in Japan. In: Wood BJ, editor. Microbiology of fermented foods. London: Blackie Academic and Professional; 1998. p. 351–415.
- Nakadai T. Koryo. 1998;200:69–80. Japanese.
- Nunomura N, Sasaki M, Asao Y, Yokotsuka T. Agric. Biol. Chem. 1976;40:491–495.
- Nunomura N, Sasaki M, Asao Y, Yokotsuka T. Agric. Biol. Chem. 1976;40:485–490.
- Sugawara E, Sakurai Y. Biosci. Biotechnol. Biochem. 1999;63:749–752.
- Sugawara E, Ohata M, Kanazawa T, Kubota K, Sakura Y. Biosci. Biotechnol. Biochem. 2007;71:761–763.
- Ohata M, Kohama K, Morimitsu Y, Kubota K, Sugawara E. Biosci. Biotechnol. Biochem. 2007;71:407–413.
- Meng Q, Kakuta T, Sugawara E. Food Sci. Technol. Res. 2012;18:429–436.
- Reichstein T, Staudinger H. GB Patent 260, 960. 1926 Oct 15.
- Garbusov V, Rehfeld G, Wölm G, Golovnja RV, Rothe M. Nahrung. 1976;20:235–241.
- Tominaga T, Blanchard L, Darriet P, Dubourdieu D. J. Agric. Food Chem. 2000;48:1799–1802.
- Tominaga T, Guimertau G, Dubourdieu D. J. Agric. Food Chem. 2003;51:1016–1020.
- Shibamoto T, Russell GF. J. Agric. Food Chem. 1977;25:109–112.
- Blanchard L, Tominaga T, Darriet P, Dubourdieu D. J. Agric. Food Chem. 2001;49:4833–4835.
- Ohata M, Tominaga T, Dubourdieu D, Kubota K, Sugawara E. J. Agric. Food Chem. 2009;57:2481–2485.
- Maciel ML, Hansen TJ, Aldinger SB, Labows JN. J. Agric. Food Chem. 1986;34:923–927.
- Alain M, Henry-Eric S, Pascal B. J. Agric Food Chem. 2008;56:4674–4680.
- Kerscher R, Grosch W. J. Agric. Food Chem. 1998;46:1954–1958.
- Tominaga T, Niclass Y, Frérot E, Dubourdieu D. J. Agric. Food Chem. 2006;54:7251–7255.
- Hofmann T, Schieberle P. J. Agric. Food Chem. 1998;46:235–241.
- Tominaga T, Dubourdieu D. J. Agric. Food Chem. 2006;54:29–33.
- Higuchi T, Aoki T, Uchida K. J. Ferment. Bioeng. 1991;71:163–167.
- Parliment T, Stahl H. Dev. Food Sci. 1995;37:805–813.
- Tressl R, Silwar R. J. Agric. Food Chem. 1981;29:1078–1082.
- Hofmann T, Schieberle P, Grosch W. J. Agric. Food Chem. 1996;44:251–255.
- Kumazawa K, Masuda H. J. Agric. Food Chem. 2003;51:2674–2678.
- Münch P, Hofmann T, Schieberle P. J. Agric. Food Chem. 1997;45:1338–1344.
- Huynh-Ba T, Matthey-Doret W, Fay LB, Bel Rhlid R. J. Agric. Food Chem. 2003;51:3629–3635.