
Abstract
In order to investigate the difference in their characteristic floral scents between Prunus mume Siebold & Zucc. and the related Prunus species, their headspace volatiles and endogenous extraction were analyzed by gas chromatography–mass spectrometry. The efficiency of substrate utilization of the flowers was studied by incubating them with different alcohol substrates. Our results indicated that benzyl acetate is a dominant compound influencing the characteristic floral scent of P. mume. An alcohol substrate concentration of 4 mmol L−1 and a reaction time of 2 h were constituted the reaction condition for catalysis of exogenous alcohol substrates by the flowers. Under these conditions, Prunus sibirica exhibited the highest utilization efficiency for benzyl alcohol substrate while the utilization efficiency of Prunus persica was the lowest. Comparative analysis of several alcohol substrates indicated that the flowers of the tested species had selective specificity for benzyl alcohol substrates.
Graphical Abstract
Prunus species exhibited different utilization efficiency on benzyl alcohol, when the flowers were fed with sufficient amount of substrates.
Prunus mume Siebold & Zucc. is an important ornamental and fruit tree species that produces strong floral fragrance. Other plants of the Prunus species lack aromatic flowers but possess strong resistance to coldness. Hybridizing P. mume with cold-resistant plants of the Prunus species is one approach for selecting cold-resistant cultivars of P. mume.Citation1) In recent years, studies on the floral scent of P. mume have achieved significant breakthroughs. For instance, research on the volatile compounds in P. mume has revealed that benzyl acetate is likely to be one of its major aromatic constituents.Citation2,3) These findings have set the foundation for further study on the difference in floral scent between P. mume and related cold-resistant plants of the Prunus species.
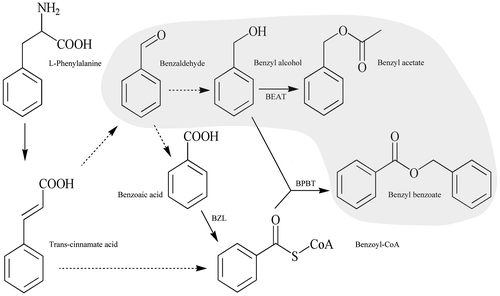
With increase in the number of research on plant secondary metabolism, the mechanisms for biosynthesis of aromatic compounds are increasingly becoming clear.Citation4) In particular, the metabolic pathway of aromatic compounds in Petunia hybridaCitation5−Citation7) has received considerable attention in recent years. The principal aromatic compounds in P. hybrida are benzenoid compounds. The metabolic pathway by which benzaldehyde is reduced to benzyl alcohol and converted to benzyl acetate is an important aspect of metabolism of benzenoid compounds (Fig. ). Preliminary research on biosynthesis of benzyl acetate has revealed that the enzyme acetyl-CoA:benzyl alcohol acetyltransferase is involved in floral scent production in Clarkia breweri.Citation8) The biosynthesis pathways of aromatic compounds involve a variety of intermediate products, some of which are not volatile or have poor volatility.Citation9) Research on the volatilization and endogenous contents of aromatic compounds and their precursors is crucial for revealing the difference in floral scent between related species.Citation10)
Fig. 1. The metabolic pathway leading to synthesis of benzyl acetate and benzyl benzoate found in floral scents of various plants. Solid lines indicate established biochemical reactions and broken lines indicate possible steps for which enzymes have not yet been characterized.
Notes: BEAT, acetyl-CoA:benzyl alcohol acetyltransferase; BPBT, benzoyl-CoA:benzyl alcohol benzoyltransferase; BZL, benzoate:CoA ligase. Volatile compounds are shown with a light gray background.
Biosynthesis of volatile esters in plants involves its catalysis by the alcohol acyltransferase, which is dependent on acyl coenzyme A.Citation11) A common function of alcohol acyltransferase is to transfer the acyl group of acyl coenzyme A to various alcohol substrates.Citation12) The functions of a series of alcohol acyltransferases have been characterized by feeding live plants with different exogenous alcohol substrates.Citation13,14) Since alcohol acyltransferase exhibits substrate preference,Citation15) plants metabolize selective alcohol substrates and their dominant ester compounds are volatilized.Citation16,17)
The objective of our study was to reveal the underlying basis of difference in floral scent between P. mume and related plants of the Prunus species, and to determine appropriate conditions for the breeding of aromatic Prunus plants. P. mume and related plants of the Prunus species that blossom in early spring in North China were used as study materials. The headspace volatilization amount and endogenous contents of benzaldehyde, benzyl alcohol and benzyl acetate of the flowers were investigated. Furthermore, the ester yields of Prunus species flowers treated with different exogenous alcohol substrates were examined. Finally, the selective specificity of different Prunus species to metabolize exogenous alcohol substrates was determined.
Materials and methods
Plant materials
Study materials were obtained from the campus of Beijing Forestry University and consisted of the following: P. mume Siebold & Zucc. “Fenhong”, strong aroma; Prunus armeniaca L. “Caoxing”, no aroma; Prunus sibirica L., light aroma; Prunus cerasifera Ehrh. “Pissardii”, no aroma; Prunus persica Batsch “Duplex”, no aroma. The experiments were conducted from February to May 2013. For every species, the shoots of 3–5 selected plants were cut. The shoots from the outside of canopy with flower buds were placed into a beaker containing ultrapure water and cultured in a climate-controlled growth chamber at 20 °C.Citation18) The light intensity was 60 μmol m−2 s−1 and the photoperiod was 12L/12D.
Collection of volatile compound
Volatile compounds were collected using the static headspace adsorption method.Citation19) The adsorption tube (CAMSCO, Houston, USA) filled with Tenax GR adsorbent (60/80 mesh; Alltech, Deerfield, IL) was pretreated using the thermal desorption instrument. Under the condition of 270 °C, N2 was introduced for 2 h to eliminate impurities from the adsorbent. When the buds of every Prunus species blossomed fully, approximately 10 flowers were picked and weighed before putting into a gas sampling bag (Ted- 050, Plastic Film Corp, USA). The bag was vacuumed using a mini pump (GSP- 300FT- 2, Gastec Corporation, Japan), then filled with 3 L clean air using activated carbon tower and placed in a culture box for 1 h at 20 °C in light. Finally, the air in the gas sampling bags was trapped into the adsorption tube at the rate of 200 mL min−1 using a mini pump. Gas sampling bags without flowers were used as controls. The procedure was repeated 3–4 times. After sampling, the sorbent tubes were kept at −20 °C until analysis.
Collection of endogenous extract
Endogenous volatile compounds in the flowers were extracted with ethyl acetate as the extraction solvent.Citation20) A total of 0.50 g of full bloom flowers was collected for each species. The flower samples were frozen in liquid nitrogen and ground to powders with a mortar and pestle. The powders were extracted using 1.5 mL ethyl acetate and anhydrous sodium sulfate was added to remove water.Citation21) Benzyl propionate was added as the internal standard. These steps were repeated three times to obtain triplicates for each sample. The obtained solution was preserved in a −20 °C refrigerator for further testing.
Catalysis of alcohol substrates by the flowers of 5 related Prunus species
Authentic standards (Sigma, USA) of hexanol, cis-3-Hexen-1-ol, benzyl alcohol, phenethanol, and cinnamyl alcohol were each diluted to 4 mmol L−1. For each alcohol substrate, 5 mL of the diluted solution was added to a 10 mL centrifuge tube. Full bloom flowers of each species (P. mume, P. armeniaca, P. sibirica, P. cerasifera and P. persica) totaling a weight of 0.50 g were weighed precisely and added to the centrifuge tube. All flowers were submerged into the solution. With the cover on, the centrifuge tube was placed in a culture box at 20 °C for 2 h. At the end of this incubation step, the flowers were immediately taken out of the solution. The residue solution was then dried on filter paper and quickly frozen in liquid nitrogen.Citation13) Solvent extraction was conducted by the same method as that used for collection of endogenous extracts from flowers.
ATD-GC/MS analysis
The ATD (Auto Thermal Desorber, TurboMatrix 650, PerkinElmer, USA) working conditions were as follows: the carrier gas was He; the primary thermal desorption temperature was 260 °C, and the time was 10 min; the cold trap temperature during the primary thermal desorption was −25 °C; the secondary thermal desorption temperature was 300 °C and the rate of temperature rise was 40 °C s−1; the valve temperature was 230 °C; the temperature of the GC/MS pipe was 250 °C; the gas split was 4.6%. The GC (Clarus 600, PerkinElmer, USA) conditions were as follows: the chromatographic column was DB-5 Low Bleed/MS column (30 m × 0.25 mm × 0.25 μm), the carrier gas was He; the starting temperature of the programmed temperature rise was 40 °C and was maintained for 2 min; then the temperature was raised to 180 °C at the rate of 4 °C min−1 and maintained for 3 min. Then the temperature was raised to 220 °C at the rate of 20 °C min−Citation1. The MS (Clarus 600T, PerkinElmer, USA) conditions were as follows: the ionization mode was EI; the electron energy was 70 eV; the scan range for the mass-to-charge ratio (m/z) was 29–500 amu; the GC/MS interface temperature was 250 °C; the ion source temperature was 220 °C.
Analysis of endogenous extracts was conducted using the same equipment and GC/MS analysis conditions described above. The samples were injected using a micropipettor. The temperature at the injection entrance was 250 °C. The GC was used in the non-split mode, the injection volume was 1 μL and the solvent delay was 3 min.
Quantitative determination of volatile compounds and endogenous extracts
The external standard method was used for quantification of volatile compounds. Authentic standards (Sigma, USA) of benzaldehyde, benzyl alcohol and benzyl acetate were each diluted to different concentration with ethyl acetate. For each diluted solution, One microlitre aliquot was added to an adsorption tube and incubated at 60 °C for 15 min.Citation22) The same ATD-GC/MS conditions as those used for the volatile compound analysis were applied. Standard curves were drawn based on the results. The linear regression equation was used for quantification of volatile compounds. The calculation for endogenous compounds was performed by adding benzyl propionate as the internal standard.Citation23)
Data processing
Turbomass version 5.4.2 (PerkinElmer) was used for the calculation of peak area. The search library was the NIST08 mass spectral library. The relative contents of the components were calculated by the peak area normalization method. Statistical analysis was performed using SPSS 18.0.0 (Version 18.0.0, IBM SPSS, IBM Corporation, Armonk, NY).
Results
Relative contents of benzaldehyde, benzyl alcohol and benzyl acetate in the headspace volatiles from the flowers of different species
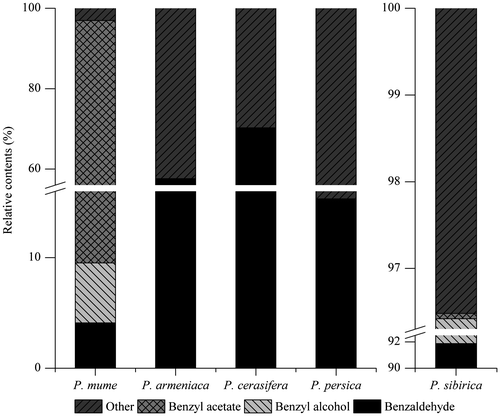
The volatile compounds in different species were different (Fig. ). Benzaldehyde, benzyl alcohol and benzyl acetate were all detected in the flowers of P. mume and P. sibirica. However, no benzyl alcohol and benzyl acetate were detected in the other three species. Benzyl acetate represented 90.36% of the volatile compounds in P. mume and only 0.06% in P. sibirica. Benzaldehyde was the common volatile compound detected in all five species. Its proportions were higher than 57.60% in P. armeniaca and P. cerasifera and the highest at 91.88% in P. sibirica. Thus, in comparison with related plants of the Prunus species, benzyl acetate was the dominant and characteristic volatile compound in P. mume.
Fig. 2. The relative contents of benzaldehyde, benzyl alcohol and benzyl acetate in the headspace volatile compounds from the flowers of 5 related plants of the Prunus species.
Notes: “Other” represented the total volatile compounds excluding these three compounds.
Relative contents of benzaldehyde, benzyl alcohol and benzyl acetate in endogenous extracts from the flowers of different species
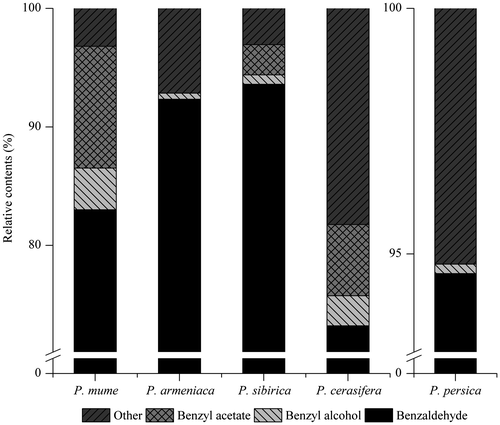
Fig. shows the relative contents of benzaldehyde, benzyl alcohol and benzyl acetate in ethyl acetate extracts. Benzyl alcohol was the common compound in the five related Prunus species, while benzyl acetate was not detected in P. armeniaca and P. persica. In terms of relative contents of the compounds, benzaldehyde was the main compound in the extracts. It constituted 73.22 and 94.60% of the extracted compounds in P. cerasifera and in P. persica, respectively. The relative content of benzyl alcohol was very low in P. armeniaca and P. persica at only 0.51 and 0.19%, respectively. In P. mume, the benzyl alcohol content was 3.51%. The relative content of benzyl acetate in P. mume was the highest among the five species at 10.28%. Thus, benzaldehyde was detected in all the tested species, while benzyl alcohol and benzyl acetate were found in large quantity only in some of the species.
Fig. 3. The relative contents of benzaldehyde, benzyl alcohol and benzyl acetate in ethyl acetate extracts from the flowers of 5 related plants of the Prunus species.
Notes: “Other” represented the total extract compounds excluding these three compounds.
Quantitative analysis of benzaldehyde, benzyl alcohol and benzyl acetate in headspace volatiles and in endogenous extracts of different species
Table shows the results of our quantitative analysis of the three volatile compounds from headspace volatiles and endogenous extracts of the five plant species. Benzaldehyde content in the endogenous extract of P. sibirica was 864.38 μg g−1, which was 7.5 times that in P. cerasifera and significantly higher than that in the other species. Benzyl acetate content in P. mume was significantly higher than those in P. sibirica and P. cerasifera. Among the headspace volatiles, benzaldehyde content was the highest in P. sibirica at 605.18 ng g−1 h−1. The volatilization amount of benzaldehyde content in P. persica was the lowest (2.96 ng g−1 h−1). The volatilization amounts of benzyl alcohol and benzyl acetate in P. mume were both high. The volatilization amount of benzyl acetate in P. mume was 4,846.80 ng g−1 h−1, but over 10,000 times lower in P. sibirica at 0.39 ng g−1 h−1. The difference between the volatilization amount and the content in endogenous extracts was significant. The content of benzaldehyde in the endogenous extract was 9.49 × 104 times that of the volatilization amount in P. persica. The benzyl acetate content in the endogenous extract of P. mume was 18.14 times the volatilization amount, while in P. sibirica, the difference reached 6.03 × 104 times. These results indicated that benzyl acetate had a stronger tendency to be volatile than benzaldehyde, and the volatilization efficiency of benzyl acetate was related to their endogenous amounts.
Table 1. Quantitative analysis of benzaldehyde, benzyl alcohol and benzyl acetate in headspace volatiles and in endogenous extracts of different species.
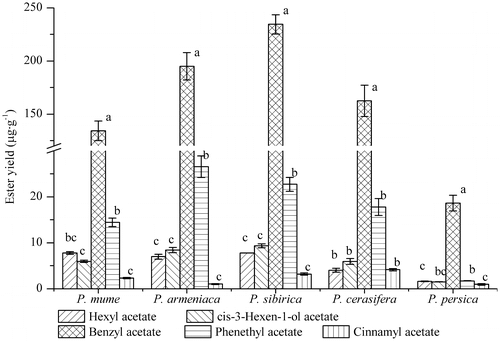
Ester yields of flowers of different plant species with different alcohol substrates
The ester yields of flowers of different plant species with different alcohol substrates are shown in Table . Under the same concentration, reaction time and temperature, the catalysis efficiency of alcohol substrates varied with the type of flower. P. mume and P. sibirica had the highest catalysis efficiency on hexanol. The latter had an ester yield reaching 7.80 μg g−1, with its product yield of hexyl acetate 4.84 times that of P. persica. The cis-3-hexen-1-ol acetate content was 5.98 μg g−1 in P. mume, which was significantly lower than that in P. armeniaca and P. sibirica. The yields of benzyl acetate from benzyl alcohol substrate catalyzed by P. armeniaca, P. sibirica and P. cerasifera flowers all exceeded that by P. mume flower. Notably, the benzyl acetate yield of reactions catalyzed by P. sibirica flowers was 12.58 times those by P. persica flowers. The yield of phenethanol substrate was the highest when catalyzed by the flowers of P. armeniaca, while that of cinnamyl alcohol was the highest when catalyzed by P. cerasifera. These experiments revealed that the catalysis efficiencies of P. persica flowers on all the substrates were significantly lower than those of other species. On the other hand, the catalysis efficiencies of P. sibirica flowers on most substrates were significantly higher than those of other species.
Table 2. Ester yields of flowers of different plant species with different alcohol substrates.
Selective specificity for alcohol substrates of flowers from different species
Five alcohol substrates, each containing different carbon chain structure, were used in this study. Hexanol and cis-3-Hexen-1-ol have straight chains and the other three have aromatic rings with different length of carbon chains.Citation24) Fig. indicates that different species had selective specificity for certain benzyl alcohol substrates. Benzyl acetate yields from benzyl alcohol were all higher than those from other alcohol substrates. High catalysis efficiency was also found for phenethanol. The ester yields of phenethanol with P. armeniaca and P. sibirica flowers were significantly higher than those of all other substrates except benzyl alcohol. No significant difference between catalysis efficiencies of hexanol, cis-3-Hexen-1-ol and cinnamyl alcohol was found for the plant species we tested. Thus, our results show that the flowers of the tested species exhibit strong selective specificity for certain alcohol substrates. Except P. persica, all of the plant species could efficiently and selectively catalyze benzyl alcohol to produce large amounts of benzyl acetate.
Fig. 4. The selective specificities for alcohol substrates of flowers from different species.
Notes: The Y axis represents the ester yield (μg g−1) with ethyl acetate as the solvent. The error bars show standard deviation. The different lowercase letters in the same group represent significant difference in Duncan’s multiple range test (P < 0.05).
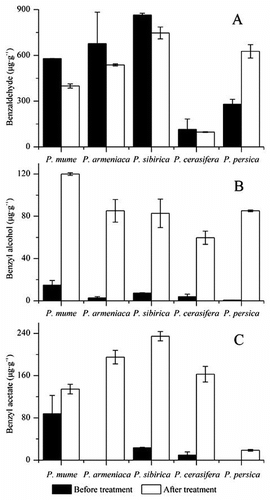
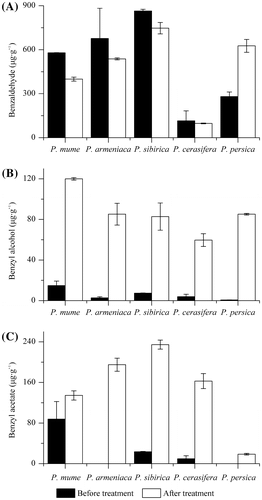
The contents of benzaldehyde, benzyl alcohol and benzyl acetate before and after treatment
Fig. shows the contents of benzaldehyde, benzyl alcohol and benzyl acetate in the flowers of different species before and after benzyl alcohol treatment. The content of benzyl alcohol was far more than that of endogenous benzyl alcohol before treatment (Fig. (B)). This result indicated that sufficient amount of benzyl alcohol substrates was obtained in the reaction system. When the flowers of Prunus species were fed with benzyl alcohol substrates, benzyl acetate were produced in all the reaction systems (Fig. (C)). The yields of benzyl acetate catalyzed by P. mume, P. sibirica and P. cerasifera flowers all exceeded the content of endogenous benzyl acetate before treatment. Fig. (A) shows that the content of benzaldehyde in the four species is decreased after benzyl alcohol treatment except P. persica. Interestingly, the content of benzaldehyde was increased after benzyl alcohol treatment in P. persica. Meanwhile, small amount of benzyl acetate was synthesized in the flower of P. persica after treatment.
Fig. 5. The contents (μg g−1) of benzaldehyde (A), benzyl alcohol (B) and benzyl acetate (C) in the flowers of different species before and after benzyl alcohol treatment.
Note: The error bars show standard deviation.
Discussion
Plants can produce volatile small molecular compounds in blossom. The amount and the aromatic features of the volatile compounds determine the aromatic characteristics of the plants.Citation10) Benzyl acetate is a dominant volatile compound in P. mume, with its relative content reaching 90.36%. Quantitative analysis of volatile compounds indicated that the volatilization amount of benzyl acetate in P. mume was 4,846.80 ng g−1 h−1. A large amount of benzyl acetate was volatilized into the air, which may be the basis for the aroma sensed by humans.Citation25) Benzyl acetate has a strong fruit and jasmine aroma.Citation26) Previous studies on C. breweriCitation27) and Silene conoideaCitation28) indicated that benzyl acetate is involved in the formation of the characteristic aroma of these plants. Compared with the flowers of P. mume, those of P. armeniaca, P. cerasifera and P. persica are not aromatic. Interestingly, benzyl acetate was not detected in the headspace volatiles of the 3 non-aromatic plant species. Furthermore, olfactory evaluation of benzyl acetate authentic standards (data not shown) confirmed that benzyl acetate is a dominant aromatic compound in P. mume.
Before volatilization, the aromatic compounds are present in different tissues of the flower.Citation29,30) Quantitative analysis of the endogenous extracts and the headspace volatiles of different plant species indicated that only a small amount of compounds was emitted from the flower in 1 h and most of the compounds remained in the floral tissue. This observation suggests that the emission of stored volatile compounds from floral tissues may be controlled, perhaps by an excretory process regulated at the cellular level.Citation31) When the content of biosynthesized volatile compound is far beyond the limit of the cellular restrictive ability, the compound will be emitted from the flowers.
Benzyl alcohol was detected in the endogenous extracts of all 5 related plants of the Prunus species (Table ). However, benzyl acetate was not detected in P. armeniaca and P. persica, which had significantly lower benzyl alcohol content compared to P. mume. In the presence of sufficient amount of benzyl alcohol substrates, all the tested plants produced benzyl acetate, indicating that the amount of substrates was an important factor influencing the ester yields. Notably, P. mume, P. sibirica and P. cerasifera were found to be able to naturally synthesize benzyl acetate (Table ). However, the amount of benzyl acetate synthesized endogenously was far lower than that synthesized after feeding them with more benzyl alcohol substrates (Fig. (C)). Thus, our results indicated that with sufficient amounts of benzyl alcohol substrate, all plants had the ability to produce large amounts of benzyl acetate. The catalysis efficiency of P. persica on benzyl alcohol was far lower than that of other species. A possible explanation for this is the absence of a high-efficiency alcohol acyltransferase for benzyl alcohol in the flowers of P. persica. P. persica is phylogenetically remote from P. mume.Citation32,33) The contribution of differences in the genetic sequence of alcohol acyltransferase in these two plants to the observed difference in their catalysis efficiency needs further investigation.
P. persica, P. armeniaca, P. sibirica and P. cerasifera are important ornamental trees that blossom in early spring in Northern China. They have similar flower shapes but lack the special aroma of P. mume. Our results show that benzyl acetate is the dominant compound determining the characteristic floral scent of P. mume. Furthermore, production of benzyl acetate is either low or absent in the flowers of Prunus plants that are non-aromatic. Our experiments indicate that, with sufficient benzyl alcohol, benzyl acetate can be abundantly biosynthesized in the flowers of some non-aromatic species. Benzyl alcohol is produced by the reduction of benzaldehyde (Fig. ), which is an intermediate product in the metabolic pathway of benzenoid compounds.Citation5,34) The end product of benzaldehyde in these metabolic pathways is a determinant of the characteristic floral scent of Prunus plants. Although a large amount of benzaldehyde exists in the endogenous extracts of various Prunus species (Fig. ), as in the case of Petunia hybridCitation35), the non-aromatic species appear to lack the molecular apparatus for reducing benzaldehyde to benzyl alcohol or part of the benzyl alcohol is converted to other products due to the existence of competitive pathways. Moreover, the increase of benzaldehyde content after treatment in P. persica indicated that the conversion of benzaldehyde to benzyl alcohol was reversible.Citation5) Further investigation is necessary to confirm these. Nevertheless, our results provide new insights into the difference in floral scent between P. mume and related plants of Prunus species, as well as theoretical support for the breeding of aromatic Prunus species.
Funding
This research was funded by the Special Project for Cultivating and Developing of the Scientific and Technological Innovation Base (Z131105002813013), the State Plan for High-Tech Research of China (2013AA102607) and Special Fund for Beijing Common Construction Project (2013).
Notes
Abbreviation: GC–MS, gas chromatography–mass spectrometry.
Related Research Data
References
- Chen J. Chinese mei flowers cultivars. Beijing: China Forestry Publishing House; 2010.
- Jin H, Chen J, Jin Y. Comparison of different cultivars of Prunus mume major gas ingredients. Acta Hortic. Sin. 2005;32:1139–1140.
- Zhao Y, Pan H, Zhang Q, Pan C, Cai M. Dynamics of fragrant compounds from Prunus mume flowers. J. Beijing For. Univ. 2010;32:201–206.
- Schwab W. Biosynthesis of plant-derived flavor compounds. Plant J. 2008;54:712–732.
- Boatright J, Negre F, Chen X, Kish CM, Wood B, Peel G, Orlova I, Gang D, Rhodes D, Dudareva N. Understanding in vivo benzenoid metabolism in petunia petal tissue. Plant Physiol. 2004;135:1993–2011.
- Hoballah ME, Stuurman J, Turlings TC, Guerin PM, Connetable S, Kuhlemeier C. The composition and timing of flower odour emission by wild Petunia axillaris coincide with the antennal perception and nocturnal activity of the pollinator Manduca sexta. Planta. 2005;222:141–150.
- Verdonk JC, Haring MA, van Tunen AJ, Schuurink RC. ODORANT1 regulates fragrance biosynthesis in petunia flowers. Plant Cell Online. 2005;17:1612–1624.
- Dudareva N, D Auria JC, Raguso RA, Pichersky E. Acetyl-CoA:benzylalcohol acetyltransferase – an enzyme involved in floral scent production in Clarkia breweri. Plant J. 1998;14:297–304.
- Lavy M, Zuker A, Lewinsohn E, Larkov O, Ravid U, Vainstein A, Weiss D. Linalool and linalool oxide production in transgenic carnation flowers expressing the Clarkia breweri linalool synthase gene. Mol. Breeding. 2002;9:103–111.
- Pichersky E, Dudareva N. Scent engineering: toward the goal of controlling how flowers smell. Trends Biotechnol. 2007;25:105–110.
- D Auria JC. Acyltransferases in plants: a good time to be BAHD. Curr. Opin. Plant Biol. 2006;9:331–340.
- Dudareva N, Pichersky E. Biochemical and molecular genetic aspects of floral scents. Plant Physiol. 2000;122:627–633.
- Aranovich D, Lewinsohn E, Zaccai M. Post-harvest enhancement of aroma in transgenic lisianthus (Eustoma grandiflorum) using the Clarkia breweri benzyl alcohol acetyltransferase (BEAT) gene. Postharvest Biol. Technol. 2007;43:255–260.
- Guterman I, Masci T, Chen X, Negre F, Pichersky E, Dudareva N, Weiss D, Vainstein A. Generation of phenylpropanoid pathway-derived volatiles in transgenic plants: rose alcohol acetyltransferase produces phenylethyl acetate and benzyl acetate in petunia flowers. Plant Mol. Biol. 2006;60:555–563.
- Beekwilder J, Alvarez-Huerta M, Neef E, Verstappen FW, Bouwmeester HJ, Aharoni A. Functional characterization of enzymes forming volatile esters from strawberry and banana. Plant Physiol. 2004;135:1865–1878.
- Aharoni A, Keizer LC, Bouwmeester HJ, Sun Z, Alvarez-Huerta M, Verhoeven HA, Blaas J, van Houwelingen AM, De Vos RC, van der Voet H. Identification of the SAAT gene involved in strawberry flavor biogenesis by use of DNA microarrays. The Plant Cell Online. 2000;12:647–662.
- Dexter R, Qualley A, Kish CM, Ma CJ, Koeduka T, Nagegowda DA, Dudareva N, Pichersky E, Clark D. Characterization of a petunia acetyltransferase involved in the biosynthesis of the floral volatile isoeugenol. Plant J. 2007;49:265–275.
- Shirasu M, Fujioka K, Kakishima S, Nagai S, Tomizawa Y, Tsukaya H, Murata J, Manome Y, Touhara K. Chemical identity of a rotting animal-like odor emitted from the inflorescence of the titan arum (Amorphophallus titanum). Biosci., Biotechnol., Biochem. 2010;74:2550–2554.
- Kong Y, Sun M, Pan HT, Zhang QX. Composition and emission rhythm of floral scent volatiles from eight lily cut flowers. J. Am. Soc. Hortic. Sci. 2012;137:376–382.
- Akakabe Y, Kusunoki A, Tanaka R, Kanetsune Y. A comparison of volatile components of setomi with its parent cultivars. Biosci. Biotechnol. Biochem. 2010;74:659–662.
- Oyama-Okubo N, Ando T, Watanabe N, Marchesi E, Uchida K, Nakayama M. Emission mechanism of floral scent in Petunia axillaris. Biosci., Biotechnol., Biochem. 2005;69:773–777.
- Proffit M, Johnson SD. Specificity of the signal emitted by Figs. to attract their pollinating wasps: comparison of volatile organic compounds emitted by receptive syconia of Ficus sur and F. sycomorus in Southern Africa. S. Afr. J. Bot. 2009;75:771–777.
- Dötterl S, Wolfe LM, Jürgens A. Qualitative and quantitative analyses of flower scent in Silene latifolia. Phytochemistry. 2005;66:203–213.
- Gonda I, Bar E, Portnoy V, Lev S, Burger J, Schaffer AA, Tadmor Y, Gepstein S, Giovannoni JJ, Katzir N, Lewinsohn E. Branched-chain and aromatic amino acid catabolism into aroma volatiles in Cucumis melo L. fruit. J. Exp. Bot. 2010;61:1111–1123.
- Firestein S. How the olfactory system makes sense of scents. Nature. 2001;413:211–218.
- Bauer K, Garbe D, Surburg H. Common fragrance and flavor materials. Weinheim: Wiley-VCH; 2006.
- Raguso RA, Pichersky E. Floral volatiles from Clarkia breweri and C. concinna (Onagraceae): recent evolution of floral scent and moth pollination. Plant Syst. Evol. 1995;194:55–67.
- Jürgens A, Witt T, Gottsberger G. Flower scent composition in night-flowering Silene species (Caryophyllaceae). Biochem. Syst. Ecol. 2002;30:383–397.
- El-Sharkawy I, Manríquez D, Flores FB, Regad F, Bouzayen M, Latché A, Pech J. Functional characterization of a melon alcohol acyl-transferase gene family involved in the biosynthesis of ester volatiles. Identification of the crucial role of a threonine residue for enzyme activity*. Plant Mol. Biol. 2005;59:345–362.
- Kondo M, Oyama-Okubo N, Ando T, Marchesi E, Nakayama M. Floral scent diversity is differently expressed in emitted and endogenous components in Petunia axillaris lines. Ann. Bot. 2006;98:1253–1259.
- Altenburger R, Matile P. Further observations on rhythmic emission of fragrance in flowers. Planta. 1990;180:194–197.
- Mowrey BD, Werner DJ. Phylogenetic relationships among species of Prunus as inferred by isozyme markers. Theor. Appl. Genet. 1990;80:129–133.
- Shimada T, Haji T, Yamaguchi M, Takeda T, Nomura K, Yoshida M. Classification of mume (Prunus mume Sieb. et Zucc.) by RAPD assay. J. Jpn Soc. Hortic. Sci. 1994;63:543–551.
- Clark DG, Pichersky E, Verdonk J, Dudareva N, Haring M, Klahre U, Schuurink R. Petunia. New York, NY: Springer; 2009. Chapter 3, Benzenoids dominate the fragrance of petunia flowers; p. 51–69.
- Orlova I, Marshall-Colón A, Schnepp J, Wood B, Varbanova M, Fridman E, Blakeslee JJ, Peer WA, Murphy AS, Rhodes D. Reduction of benzenoid synthesis in petunia flowers reveals multiple pathways to benzoic acid and enhancement in auxin transport. The Plant Cell Online. 2006;18:3458–3475.