
Abstract
l-ribose isomerase (L-RI) from Cellulomonas parahominis MB426 can convert l-psicose and d-tagatose to l-allose and d-talose, respectively. Partially purified recombinant L-RI from Escherichia coli JM109 was immobilized on DIAION HPA25L resin and then utilized to produce l-allose and d-talose. Conversion reaction was performed with the reaction mixture containing 10% l-psicose or d-tagatose and immobilized L-RI at 40 °C. At equilibrium state, the yield of l-allose and d-talose was 35.0% and 13.0%, respectively. Immobilized enzyme could convert l-psicose to l-allose without remarkable decrease in the enzyme activity over 7 times use and d-tagatose to d-talose over 37 times use. After separation and concentration, the mixture solution of l-allose and d-talose was concentrated up to 70% and crystallized by keeping at 4 °C. l-Allose and d-talose crystals were collected from the syrup by filtration. The final yield was 23.0% l-allose and 7.30% d-talose that were obtained from l-psicose and d-tagatose, respectively.
Graphical Abstract
We described the production of l-allose and d-talose from l-psicose and d-tagatose using immobilized enzyme of recombinant L-RI.
Key words:
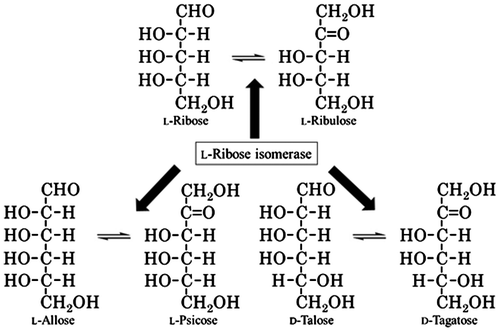
Monosaccharides and their derivatives which hardly exist in nature are called “rare sugars”.Citation1) There is little knowledge of biological and physiological functions of rare sugars so far.Citation2–5) For instance, d-allose, one of rare aldo-hexose, has been reported to have a possibility to be applied for pharmaceutical industry such as inhibitor of ischemia/reperfusion injury.Citation5) l-Fructose and l-xylulose are potential inhibitors of various glycoprotein processes in various cell culture systems.Citation6) Therefore, to investigate them, we are studying on production of various kinds of rare sugars using microorganisms and their enzymes. We focused on l-ribose isomerase (L-RI), which is a very important enzyme to produce various rare sugars. L-RI catalyzes the reversible reactions such as l-ribose ↔ l-ribulose, l-allose ↔ l-psicose, and d-talose ↔ d-tagatose. L-RI has been found in only two types of microorganisms (Fig. ). In our previous paper, we reported that Acinetobacter calcoaceticus DL-28Citation7,8) and Cellulomonas parahominis MB426Citation9) can constitutively produce L-RI. These enzymes were purified, characterized, and over-expressed in E. coli. We reported that L-RI from C. parahominis MB426 possesses a high thermal stability and broad substrate specificity as compared with that from A. calcoaceticus DL-28. Therefore, L-RI of C. parahominis MB426 is demonstrated to be a good candidate for rare sugars production.
Fig. 1. Schematic representation of aldose–ketose interconversion reactions catalyzed by L-RI.
Since l-allose ($680/g, Omicron Biochemical, Inc.) and d-talose ($380/g, Omicron Biochemical, Inc.), which are very expensive rare aldo-hexose, are not commercially available and limited for research works. For instance, methyl 4-O-benzyl-α-l-allo-pyranoside, which is l-allose homolog could be synthesized by organic reaction.Citation10) However, it requires high cost and complicated process. It can only be produced through a series of costly and laborious chemical reactions. Moreover, there are few examples of d-talose synthesis. Therefore, we attempted to produce them using microbes and its enzymes with high efficiency at low cost more. The biological effects of l-allose and d-talose have not been clarified to date, l-allose and d-talose may have great physiological functions.
In the present paper, we described the production of l-allose and d-talose from l-psicose and d-tagatose using immobilized enzyme of recombinant L-RI under the optimized condition, respectively.
Materials and methods
Chemicals
All chemical agents were purchased from Wako Pure Chemicals (Osaka). DAIAION HPA25L resin was purchased from Mitsubishi chemical Co. (Tokyo). l-psicose and d-tagatose was prepared by our laboratory as described previously.Citation11–15)
Culture condition
The L-RI gene of C. parahominis MB426 was inserted in pQE30, and the constructed plasmid was transformed in E. coli JM 109.Citation9) The recombinant E. coli JM109 was cultured in a medium composed of 3.5% polypepton, 2.0% yeast extract, 0.5% NaCl, and 100 μg/ml ampicillin in the 15-L jar fermentor at 30 °C for 12 h. Recombinant L-RI was induced by addition of 1.0 mM IPTG in the culture medium. After 4 h, the cells were harvested by centrifugation at 10,000 rpm for 10 min at 4 °C.
Partial purification of recombinant L-RI
The collected cells were washed twice with 50 mM glycine-NaOH buffer (pH 9.0) and were disrupted by sonication (Branson Sonifier 450, Dunbury, USA) at 4 °C. The cell free extract was suspended with final concentration of 10 mM MnCl2. The cell debris was precipitated by centrifugation at 10,000 rpm for 10 min at 4 °C. The supernatant was collected and added by polyethylene glycol #6000 to a final concentration 5% (w/v), and the mixture was stirred for 30 min at 4 °C. This mixture was centrifuged at 12,000 rpm for 30 min and precipitate was discarded. Polyethylene glycol #6000 was added to a final concentration 15% (w/v). After stirring for 30 min, the precipitate was collected by centrifugation at 12,000 rpm for 30 min and was resuspended in a small quantity of 10 mM glycine-NaOH buffer (pH 9.0). This final solution was used as partially purified enzyme.
Enzyme assay and protein assay
The L-RI activity was assayed by measuring the increase of keto-sugar, l-ribulose, using cysteine-carbazole method.Citation16) One unit of L-RI is defined as the amount of enzyme that converts 1 μmol of l-ribose to l-ribulose in 1 min. The reaction mixture was assayed by HPLC system (Shimadzu, Kyoto) equipped with a refractive index detector (Shimadzu RID-10A) and a separation column (GL-C611, Hitachi, Tokyo) at 60 °C eluted with 10−4 M NaOH at a flow rate of 1.0 ml/min. Protein concentration was measured according to the method of Bradford with bovine serum albumin as a standard.Citation17)
Optimization of L-RI immobilization condition
The partially purified L-RI (1000 units) was immobilized on 1, 2, 3, 4, 5, or 10 g of DIAION HPA25L at 4 °C for 2 days. Immobilization efficiency was calculated with residual activity in the supernatant, which was analyzed by cysteine-carbazole method as described above.
Sugar conversion
Conversion reactions were performed by batch method in conical flask. A reaction mixture contained 50 mM glycine-NaOH buffer (pH 9.0), 10% (w/w) l-psicose and d-tagatose, and the recombinant L-RI at 40 °C. The production of l-allose from l-psicose was performed with 2000 units of the recombinant L-RI for 24 h. The production of d-talose from d-tagatose was performed with 1000 units of the recombinant L-RI for 4 h.
Reusability of immobilized L-RI
After the reaction reached an equilibrium state, the reaction mixture was collected by filtration, and the immobilized L-RI was used for batch reaction again. Several rounds of conversion using immobilized enzymes were performed.
Separation of l-allose and l-psicose, d-talose and d-tagatose
After reaction, the each reaction solution was filtered. The filtrate was deionized with a mixture of DIAION SK1B (H+) (Mitsubishi chemical Co., Tokyo) and Amberlite IRA-411 () (ORGANO Co., Tokyo) ion-exchange resins. Each of the solution contained the substrate and the product was concentrated using a vacuum rotary evaporator to a concentration of 30%. After concentration, the each mixture was applied to a column of DOWEX 50 W-X2 (The Dow Chemical Company, USA) in the Ca2+ form. This column can separate a reaction mixture, which is composed of two main components. The sample was eluted with deionized water and 10 ml of each fraction was collected. The fractions containing the product were pooled and concentrated by evaporation.
Identification of product
The product was identified by HPLC analysis, 13C NMR spectrophotometer (model ALPHA 400, JEOL, Tokyo) at 400 MHz according to the method of Gray, optical rotation with P-1030 (Jasco, Tokyo), and melting point measurements.Citation18)
Results
Immobilization of L-RI
When 1,000 units of L-RI was immobilized with 1, 2, 3, or 4 g of DIAION HPA25L beads, residual activity in the supernatant was confirmed. Since 1,000 units of L-RI was completely bound to 5 or 10 g of DIAION HPA25L beads, the best immobilization condition of L-RI was achieved when 5 g of beads was used. The partially purified L-RI, which was prepared from 3 L culture, had 18,000 U of enzyme activity. Based on the result of the immobilization test, the enzyme could be immobilized with 90 g beads.
Production of l-allose from l-psicose
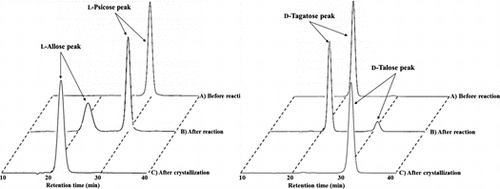
As shown in Fig. (A), when the reaction at 40 °C for 1 day was performed, the equilibrium ratio was about 33 (l-allose): 67 (l-psicose) from 10% (w/w) l-psicose as the substrate. Several cycles of conversion using immobilized enzyme were performed. The immobilized enzyme showed over 33% of product conversion in the 6th cycle (Fig. ). Since at 7th batch, the reaction could not reach an equilibrium state. The final yield of l-allose was 33.0 g from 1 L of 10% (w/w) l-psicose syrup for 7th reactions.
Fig. 2. l-allose (A) and d-talose (B) production by immobilized L-RI.
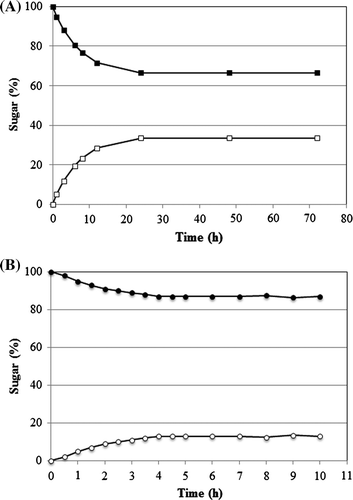
Fig. 3. HPLC of l-allose.
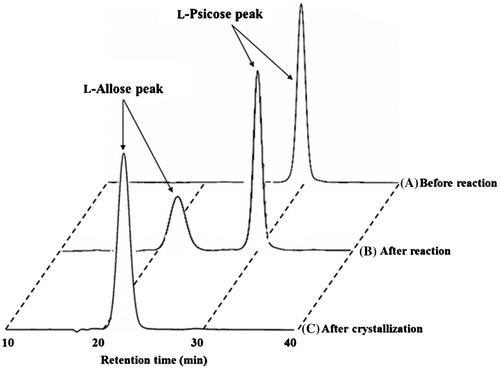
The product, which was expected to be l-allose, was isolated from the reaction mixture and purified according to the method described in Materials and Methods. The separated fraction containing l-allose was analyzed by HPLC, and confirmed the purity of the product. The purified l-allose was concentrated to about 70% (w/w) by evaporation and crystalized at 4 °C. The final yield of l-allose crystal was 23.0 g.
The obtained crystal was measured for an optical rotation property and melting point. The crystal structure was analyzed by 13C NMR spectroscopy. The main signals in the 13C NMR spectra of the isolated product were identical with those of authentic d-allose. The retention time of HPLC was indistinguishable from that of authentic d-allose. The optical rotation of the isolated product was determined to be −13.07° (5.0%, 30 °C, in H2O), whereas that of authentic d-allose was +13.56°. The melting point of the product and authentic d-allose was 131 °C. These results suggested that the product was l-allose.
Production of d-talose from d-tagatose
As shown in Fig. (B), when the reaction for 4 h was performed, the equilibrium rate was about 13 (d-talose): 87 (d-tagatose) from 10% (w/w) d-tagatose as a substrate. When several cycles of conversion were performed, the immobilized enzyme showed over 13% of product conversion from in the 37th cycle (Fig. ). The final yield of batch d-talose was 9.50 g from about 730 mL of 10% (w/w) d-tagatose syrup for 38th reactions.
Fig. 4. HPLC of d-talose.
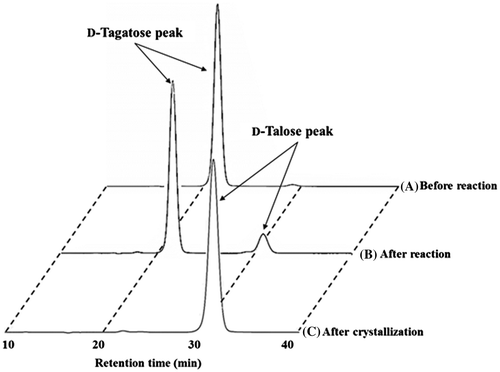
In the same methods as l-allose production, d-talose was isolated and purified. The purified d-talose was concentrated to about 70% (w/w) by evaporation and crystalized at 4 °C. The final yield of d-talose crystal was 7.3 g.
The main signals in the 13C NMR spectra of the isolated product were identical with those of authentic d-talose. The retention time of HPLC was indistinguishable from that of authentic d-talose. The optical rotation and the melting point of the product were identical with authentic d-talose (+22.01°, 127 °C). These results suggested that the product was d-talose.
Discussion
In recent years, not only consumers but also nutritionists and clinicians have shown a great deal of interest in rare sugars. Due to the scarcity in nature and high cost of rare sugar, their utilization is limited. Therefore, the development of mass production process is greatly important for increasing the utilization of these sugars. In this paper, we succeeded in the production of l-allose and d-talose from l-psicose and d-tagatose, respectively, without any significant by-product formation using the immobilized L-RI of C. parahominis MB426. We previously reported the production of d-psicose from d-fructose by d-tagatose 3-epimerase of P. cichorii ST-24.Citation11,12) d-psicose was then reduced to allitol by Enterobacter agglomerans 221e.Citation13) And, l-psicose was obtained from allitol using Gluconobacter frateurii IFO3254.Citation14) d-tagatose was also able to be produced from galactitol.Citation15) We attempted to produce l-psicose and d-tagatose from d-fructose and galactitol, respectively, as mentioned above. Using l-psicose and d-tagatose as a substrate, we produced l-allose and d-talose using the immobilized L-RI. The final yield of l-allose and d-talose crystals could be obtained each other 23.0 and 7.30 g, respectively. It is a novel method of l-allose and d-talose production, and this shows that L-form sugar was able to produce from natural sugar through this method.
Among the previous immobilized methods tested as carriers for immobilization, the highest activity yield was that of immobilized L-RI on DIAION HPA25L beads.Citation19) Thus, DIAION HPA25L beads were used as carriers throughout this study. We prepared immobilized enzyme in order to compare L-RI of C. parahominis MB426 with that of A. calcoaceticus DL-28. The remarkable property is that the immobilized L-RI of C. parahominis can continuously reuse over 150 h at 40 °C, while that L-RI of A. calcoaceticus could only catalyze for 70 h at 30 °C.Citation7) Furthermore, the immobilized L-RI of C. parahominis is more thermostable than that of A. calcoaceticus, this showed that the immobilized L-RI of C. parahominis is more useful in rare sugar production than that of A. calcoaceticus. Moreover, the substrate used was not consumed in this method and the immobilized L-RI of C. parahominis retained almost its original level of activity for a long time. This present information has considerable importance for the possible commercial manufacture of l-allose and d-talose by an enzymatic process.
Authorship
All authors were responsible for the study concept and design. Y.T., and G.T., carried it out. All the authors contributed to the critical revision of the manuscript.
Additional information
Funding
References
- Granström TB, Takata G, Tokuda M, Izumori K. Izumoring: a novel and complete strategy for bioproduction of rare sugars. J. Biosci. Bioeng. 2004;97:89–94.10.1016/S1389-1723(04)70173-5
- Matsuo T, Izumori K. Effect of dietary d-psicose on diurnal variation in plasma glucose and insulin concentration of rats. Biosci. Biotechnol. Biochem. 2006;70:2081–2085.
- Hayashi N, Iida T, Yamada T, Okuma K, Takehara I, Yamamoto T, Yamada K, Tokuda M. Study on the postprandial blood glucose suppression effect of d-psicose in borderline diabetes and the safety of long-term ingestion by normal human subjects. Biosci. Biotechnol. Biochem. 2010;74:510–519.10.1271/bbb.90707
- Sui L, Dong Y, Watanabe Y, Yamahuchi F, Hatano N, Izumori K, Tokuda M. Growth inhibitory effect of d-allose on human ovarian carcinoma cell in vitro. Anticancer Res. 2005;25:2639–2644.
- Hossain MA, Izuishi K, Tokuda M, Izumori K, Maeta H. d-allose has a strong suppressive effect against ischemia/reperfusion injury: a comparative study with allopurinol and superoxide dismutase. J. Hepatobiliary Pancreat Surg. 2004;11:181–189.
- Muniruzzaman S. Inhibition of glycoprotein processing by l-fructose and l-xylulose. Glycobiology. 1996;6:795–803.
- Shimonishi T, Izumori K. New enzyme, l-ribose isomerase from Acinetobacter sp. strain DL-28. J. Ferment. Bioeng. 1996;81:493–497.
- Mizanur RM, Takada G, Izumori K. Cloning and characterization of a novel gene encoding l-ribose isomerase from Acinetobacter sp. strain DL-28 in Escherichia coli. Biochim. Biophys. Acta. 2001;1521:141–145.
- Morimoto K, Terami Y, Maeda Y, Yoshihara A, Takata G, Izumori K. Cloning and characterization of the l-ribose isomerase gene from Cellulomonas parahominis MB426. J. Biosci. Bioeng. 2013;115:377–381.10.1016/j.jbiosc.2012.10.021
- Alonzo DD, Guaragna A, Napolitano C, Palumbo G. Rapid access to 1,6-anhydro-β-l-hexopyranose derivatives via domino reaction: synthesis of l-allose and l-glucose. J. Org. Chem. 2008;73:5636–5639.
- Itoh H, Sato T, Izumori K. Preparation of d-psicose from d-fructose by immobilized d-tagatose 3-epimerase. J. Ferment. Bioeng. 1995;80:101–103.10.1016/0922-338X(95)98186-O
- Takeshita K, Suga A, Takada G, Izumori K. Mass production of d-psicose from d-fructose by a continuous bioreactor system using immobilized d-tagatose 3-epimerase. J. Biosci. Bioeng. 2000;90:453–455.10.1016/S1389-1723(01)80018-9
- Muniruzzaman S, Tokunaga H, Izumori K. Conversion of d-psicose to allitol by Enterobacter agglomerans strain 221e. J. Ferment. Bioeng. 1995;79:323–327.
- Takeshita K, Shimonishi T, Izumori K. Production of l-psicose from allitol by Gluconobacter frateurii IF0 3254. J. Ferment. Bioeng. 1996;81:212–215.
- Muniruzzaman S, Tokunaga H, Izumori K. Isolation of Enterobacter agglomerans strain 221e from soil, a potent d-tagatose producer from galactitol. J. Ferment. Bioeng. 1994;78:145–148.
- Dische Z, Borenfreuod E. A new spectrophotometric method for the detection of keto sugars and trioses. J. Biol. Chem. 1951;192:583–587.
- Bradford MM. A rapid and sensitive method for the quantitation of protein utilizing the principle of protein–dye binding. Anal. Biochem. 1976;72:248–254.
- Que L, Gray GR. Carbon-13 nuclear magnetic resonance spectra and the tautomeric equilibriums of ketohexoses in solution. Biochemistry. 1974;13:146–153.10.1021/bi00698a023
- Bhuiyan HS, Itami Y, Izumori K. Immobilization of l-rhamnose isomerase and its application in l-mannose production from l-fructose. J. Ferment. Bioeng. 1997;84:558–562.