
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
This work aimed to evaluate the skin anti-photoaging properties of ginsenoside Rb3 (Rb3), one of the main protopanaxdiol-type ginsenosides from ginseng, in HaCaT keratinocytes. The skin anti-photoaging activity was assessed by analyzing the levels of reactive oxygen species (ROS), pro-matrix metalloproteinase-2 (proMMP-2), pro-matrix metalloproteinase-9 (proMMP-9), total glutathione (GSH), and superoxide dismutase (SOD) activity as well as cell viability in HaCaT keratinocytes under UV-B irradiation. When HaCaT keratinocytes were exposed to Rb3 prior to UV-B irradiation, Rb3 exhibited suppressive activities on UV-B-induced ROS, proMMP-2, and proMMP-9 enhancements. On the contrary, Rb3 displayed enhancing activities on UV-B-reduced total GSH and SOD activity levels. Rb3 could not interfere with cell viabilities in UV-B-irradiated HaCaT keratinocytes. Rb3 plays a protective role against UV-B-induced oxidative stress in human HaCaT keratinocytes, proposing its potential skin anti-photoaging properties.
Graphical abstract
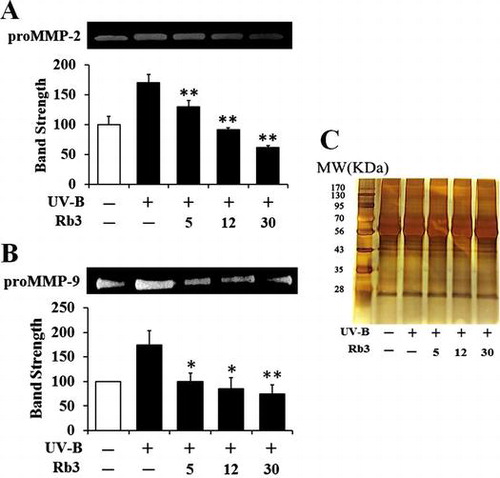
Effects of Rb3 on the UV-B-induced proMMP-2 (A) and proMMP-9 (B) activities in conditioned media of HaCaT keratinocytes. In (C), the equal loading was shown.
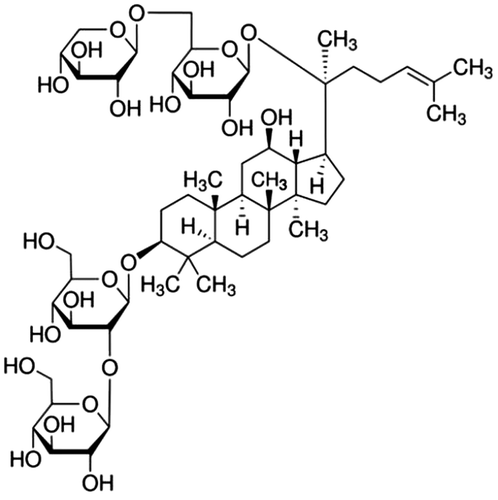
Ginseng, the dried root of Panax ginseng Meyer, has been used as a popular traditional medicine a long time, especially in Oriental countries such as Korea, China, and Japan. In recent times, as its popularity has increased all over the world, its pharmacological efficacies have drawn interest nowadays more than ever before. Most of the known pharmacological activities of ginseng are attributed to its peculiar ingredients called ginsenosides. Ginsenosides, also known as ginseng saponins, are categorized into four types, such as protopanaxadiol (PPD)-, protopanaxatriol (PPT)-, oleanolic acid-, and ocotillol-type ginsenosides, based upon the chemical structures of their aglycones called sapogenins.Citation1) A PPD-type ginsenoside Rb3 (Rb3, Fig. ) is one of the major active ingredients of total 150 ginsenosides identified from ginseng.Citation1)
Fig. 1. The chemical structure of ginsenoside Rb3 (Rb3).
A variety of pharmacological properties of Rb3 in purified form are currently ascertained. Rb3 ameliorates myocardial ischemia–reperfusion injury in rats through alleviating the decrease of malondialdehyde content and the decrease of superoxide dismutase (SOD) activity.Citation2) Rb3 plays a cardioprotective role against myocardial ischemia–reperfusion injury induced by oxygen and glucose deprivation followed by reperfusion in mouse cardiac myoblast cells via the suppression of JNK-mediated NF-κB activation, ROS generation, and expression of MMP-2 and MMP-9.Citation3) Rb3 was similarly shown to possess a cardioprotective effect against isoproterenol-induced myocardial injury and heart function impairment due to its antioxidant activity which alleviates the increase of malondialdehyde content and the decrease of SOD and catalase activities.Citation4) Rb3 inhibits angiotensin II-induced proliferation of cultured vascular smooth muscle cells through suppressing angiotensin II-induced G0/G1 to S phase transition and attenuating expression of mRNAs of c-fos, c-jun, and c-myc, which suggests its preventive effect on cardiovascular diseases.Citation5) Rb3 exhibits a neuroprotective property through depressing the basal synaptic transmission in correlation with the activation of the GABAA receptor as a ligand-gated ion channel consisting of chloride channel complex.Citation6) Rb3 exerts antidepressant-like effects which are assessed using forced swim test, tail suspension test, and learned helplessness procedure, and its effects are based upon biochemical variations in brain-derived neurotrophic factor and monoamine neurotransmitters, including 5-hydroxytryptamine, dopamine, and norepinephrine.Citation7) Until recently, its beneficial effects on human skin cells have not been clearly assessed.
Skin photoaging is defined as premature aging due to repeated exposure to UV radiation from the sun and artificial UV sources. UV-B (280–315 nm) radiation, regarded as one of well-known causes of skin photoaging, evokes a photo-oxidation reaction which disrupts the antioxidant status of skin cells and increases the cellular ROS level.Citation8–11) In epidermal keratinocytes, UV-B radiation induces the immediate generation of superoxide radical which is sequentially converted to other ROS species, such as hydrogen peroxide and hydroxyl radical.Citation12)
Matrix metalloproteinases (MMPs), a large and complex family of zinc-dependent endopeptidases, are capable of degrading essentially all components of extracellular matrix.Citation13) Skin photoaging is closely linked with the deterioration of dermal extracellular matrix due to increased MMP expression and decreased collagen synthesis under oxidative stress.Citation14,15) Collagen degradation, considered as a cause of aging in both naturally aged and photoaged skins, is associated with the induction of MMPs which are secreted from epidermal keratinocytes and dermal fibroblasts.Citation16) UV radiation, including UV-B, induces production of MMPs and then gives rise to damages to the skin, leading to skin photoaging.Citation17)
In the present work, the skin anti-photoaging properties of Rb3 was assessed by evaluating the levels of ROS, pro-matrix metalloproteinase-2 (proMMP-2), and pro-matrix metalloproteinase-9 (proMMP-9) as well as some antioxidant components, such as total glutathione (GSH) and SOD, in HaCaT keratinocytes under UV-B irradiation. This work focused on proMMP-2 and proMMP-9, based on the previous finding that their expression and activities were enhanced in UV-B-irradiated HaCaT keratinocytes.Citation15)
Materials and methods
Chemicals
Rb3 (purity ≥98%) was obtained from Ambo Institute (Seoul, Korea). Bovine serum albumin (BSA), gelatin, Bradford reagent, sodium dodecyl sulfate (SDS), 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT), 2′,7′-dichlorofluorescein diacetate (DCFH-DA), dihydrorhodamine 123, 5,5′-dithiobis (2-nitrobenzoic acid) (DTNB), glutathione reductase (GR), reduced GSH, NADPH, cytochrome c, xanthine, and xanthine oxidase were from Sigma-Aldrich Chemical Co. (St. Louis, MO, USA). Fetal bovine serum (FBS), Dulbecco’s modified Eagle’s medium (DMEM), and penicillin–streptomycin were from HyClone Laboratories Inc. (Logan, UT, USA). Cell lysis buffer was purchased from Promega Korea (Seoul, Korea). All other chemicals used in the present work were of the highest grade commercially available.
Cell culture
An immortalized HaCaT keratinocyte cell line (ATCC, Manassas, VA, USA) was grown in DMEM containing 10% heat-inactivated FBS, 100 units/mL penicillin, and 100 μg/mL streptomycin in a humidified atmosphere with 5% CO2 at 37 °C. Prior to the treatments, 1 × 105 HaCaT keratinocytes were typically seeded on 24-well plates and grown overnight, washed twice with 1 mL phosphate-buffered saline (PBS), and replaced with 1 mL FBS-free medium. After UV-B irradiation without or with Rb3 pretreatment, the mammalian cells were cultured under the same growth conditions described above.
UV-B irradiation
An ultraviolet lamp (peak, 312 nm; model VL-6 M, Vilber Lourmat, Marine, France) was used as a UV-B source. A radiometer (model VLX-3 W, Vilber Lourmat, Marine, France) with a sensor (bandwidth, 280–320 nm; model CX-312, Vilber Lourmat, Marine, France) was used to monitor a radiation intensity. The HaCaT keratinocytes used in the present work were irradiated with 70 mJ/cm2 UV-B radiation at 25 °C.
Preparation of cellular lysate
Adherent cells were washed twice with PBS and stored on ice for 5 min. The cells were obtained by scraping off the bottom of the dish with a cell scraper and centrifuged at 15,000 rpm for 10 min. The cell pellets were resuspended in cell lysis buffer [25 mM Tris-phosphate (pH 7.8), 2 mM 1,2-diaminocyclohexane-N,N,Nv,Nv-tetraactic acid, 2 mM dithiothreitol, 10% glycerol, 1% Triton X-100] and stored for 30 min on ice. Cellular lysates were taken after centrifugation at 15,000 rpm for 15 min.
Protein contents in cellular lysates were determined according to the procedure of BradfordCitation18) using BSA as a standard.
Determination of intracellular reactive oxygen species (ROS)
To fluorometrically determine intracellular ROS in HaCaT keratinocytes, a redox-sensitive fluorescent probe DCFH-DA, which generates the fluorescent 2′,7′-dichlorofluorescein (DCF; λexcitation = 485 nm, λemission = 530 nm) upon enzymatic reduction and subsequent oxidation by ROS, was used as previously described.Citation19) After the treatment with Rb3 and/or 20 µM DCFH-DA for 30 min at 37 °C, the cells were twice washed with 1 mL FBS-free medium. The cells are resuspended in 1 mL FBS-free medium and irradiated with 70 mJ/cm2 UV-B radiation. The intracellular ROS levels were immediately quantitated by Multi-Mode Microplate Reader (Synergy™ Mx, BioTek Instruments, Winooki, VT, USA).
In confocal microscopic analysis,Citation19) the cells were treated with Rb3 and/or 20 µM dihydrorhodamine 123 for 30 min at 37 °C, irradiated with 70 mJ/cm2 UV-B radiation, and immediately analyzed using Confocal Laser Scanning Microscope (Fluoview-FV300, Olympus, Tokyo, Japan).
Cell viability assay
To determine the cellular survivals of HaCaT keratinocytes in the presence of Rb3, the cell viability was determined using MTT assay which is used to assess metabolic activity.Citation20) Cells were treated with Rb3 for 30 min. After removing the medium by suction, the cells were treated with 5 µg/mL MTT in medium for 4 h. The cells were then lysed with dimethyl sulfoxide, and the amount of formazan, generated from the reduction of MTT by the mitochondria of living cells, was quantitated by the absorbance at 540 nm.
Gelatin zymography
The gelatinolytic activities of proMMP-2 and proMMP-9 in conditioned medium were determined using zymographic analysis as previously described.Citation21) Cells were further incubated for 24 h at 37 °C, and twice washed with 1 mL PBS. The cells in 1 mL FBS-free medium were treated with Rb3 for 30 min and irradiated with 70 mJ/cm2 UV-B radiation. The conditioned medium, taken from the irradiated culture incubated for 24 h at 37 °C, was fractionated on 10% (w/v) SDS–PAGE gel impregnated with 1 mg/mL gelatin under a non-reducing condition. The proteins in the gel were renatured by shaking with 2.5% Triton X-100 at room temperature for 30 min, which was repeated two times, and incubated in incubation buffer (50 mM Tris buffer, pH 7.8, 5 mM CaCl2, 0.15 M NaCl, 1% Triton X-100) for 24 h. After the gel was stained with 0.1% Coomassie Brilliant Blue R-250, gelatin-degrading enzyme activities were convinced as clear zones against a blue background. proMMP-2 and proMMP-9 activity bands were identified in accordance with its molecular mass, which was estimated by molecular mass markers.
Western blotting analysis
In order to detect proMMP-2 in conditioned medium and proMMP-9 in cellular lysate, Western blotting analyses were performed using anti-MMP-2 (ALX-210–753, Enzo Life Sciences, Farmingdale, NY, USA) and anti-MMP-9 (3852S, Cell Signaling Technology, Danvers, MA, USA) antibodies as primary antibodies. GAPDH, as an internal standard, in cellular lysate was detected using anti-GAPDH antibody (LF-PA0212, AbFrontier, Seoul, Korea). Conditioned medium and cellular lysate were run on 10% (w/v) SDS–PAGE and electrotransferred to PVDF membranes. The membranes were blocked with blocking buffer (2% BSA in 1× TBS-Tween 20), probed with primary antibody overnight at 4 °C, incubated with secondary antibody (goat anti-rabbit IgG-pAb-HRP-conjugate; ADI-SAB-300, Enzo Life Sciences, Farmingdale, NY, USA) for 1 h at room temperature, and developed using an enhanced West-save up™ (AbFrontier, Seoul, Korea).
Quantitation of total GSH
As previously described,Citation22) total GSH contents in cellular lysates were quantitated using an enzymatic recycling assay based on GR. The reaction mixture (200 μL), containing 175 mM KH2PO4, 6.3 mM EDTA, 0.21 mM NADPH, 0.6 mM DTNB, 0.5 units/mL GR, and cellular lysate, was incubated at 25 °C. Absorbance at 412 nm was monitored using a microplate reader. Total GSH was reported as μg/mg protein.
Determination of SOD activity
As previously described,Citation23) total SOD activity in cellular lysates was spectrophotometrically determined as reduction of cytochrome c with xanthine/xanthine oxidase system. The reaction mixture (200 μL) contained 50 mM phosphate buffer (pH 7.4), 0.01 units/mL xanthine oxidase, 0.1 mM EDTA, 1 μM catalase, 0.05 mM xanthine, 20 μM cytochrome c, and cellular lysate. A change in absorbance was monitored at 550 nm.
Statistical analysis
The results were represented as mean ± SD. Differences between experimental groups were analyzed using one-way ANOVA followed by post hoc Tukey HSD test for multiple comparisons. A p value less than 0.05 was considered statistically significant.
Results
Suppressive effect of Rb3 on UV-B-induced ROS elevation
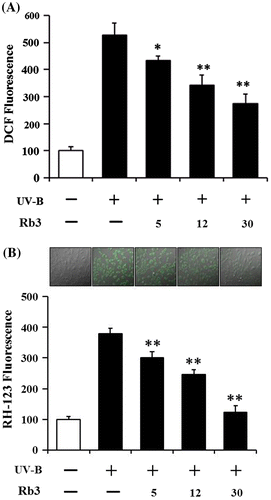
HaCaT keratinocytes were exposed to varying concentrations (0, 5, 12, or 30 μM) of Rb3 prior to the irradiation with 70 mJ/cm2 UV-B radiation. As shown in Fig. (A), the UV-B irradiation alone, in the absence of Rb3 pretreatment, could cause about 5.3-fold enhancement in the ROS level over that in the non-irradiated control cells. Rb3 attenuated the UV-B-induced ROS elevation in a concentration-dependent fashion (Fig. (A)). Rb3 at the concentrations of 5, 12, and 30 μM reduced the UV-B-induced ROS enhancement to 82.3, 65.1, and 52.2% of that from the UV-B irradiation alone, respectively (Fig. (A)). As shown in Fig. (B), this attenuating effect of Rb3 was also confirmed using confocal microscopic analysis. In the confocal microscopic analysis, the UV-B irradiation alone, in the absence of Rb3 pretreatment, could give rise to about 3.8-fold elevation in the ROS level over that in the non-irradiated control cells (Fig. (B)). Rb3 at the concentrations of 5, 12, and 30 μM diminished the UV-B-induced ROS enhancement to 79.4, 65.2, and 32.6% of that from the UV-B irradiation alone, respectively (Fig. (B)). Taken together, Rb3 displays a suppressive effect on the UV-B-induced ROS elevation in HaCaT keratinocytes.
Fig. 2. Effect of Rb3 on reactive oxygen species (ROS) elevation in HaCaT keratinocytes under irradiation with 70 mJ/cm2 UV-B radiation. HaCaT cells were subjected to fresh medium with the indicated concentrations (0, 5, 12, or 30 μM) of Rb3 for 30 min prior to the irradiation. The intracellular ROS level was quantitated using DCFH-DA in a microplate fluorometer (A) and dihydrorhodamine 123 in confocal microscopic analysis (B).
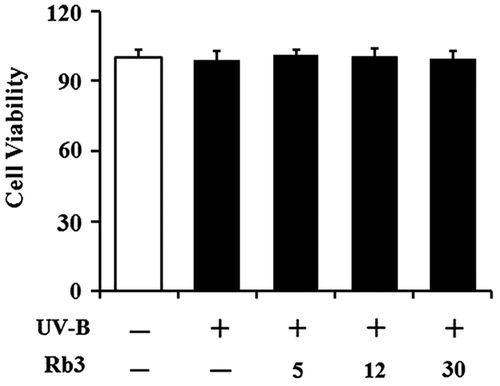
No cytotoxicity on cellular viability
To detect whether UV-B radiation at the used intensity and Rb3 at the used concentrations exhibit cytotoxicity on HaCaT keratinocytes or not, their effects on the viabilities of HaCaT keratinocytes were determined using MTT assay. As shown in Fig. , UV-B irradiation alone could not make a change in the viability of HaCaT keratinocytes, and their viabilities remained to be similar to that of the non-irradiated cells. Rb3 was not able to alter the viability of HaCaT keratinocytes under UV-B irradiation (Fig. ). In brief, neither UV-B radiation nor Rb3, at the used conditions, exhibits cytotoxicity on HaCaT keratinocytes.
Fig. 3. Effect of Rb3 on cellular viability in HaCaT keratinocytes under irradiation with 70 mJ/cm2 UV-B radiation.
Diminishing effect on UV-B-induced proMMP-2 and proMMP-9 elevation
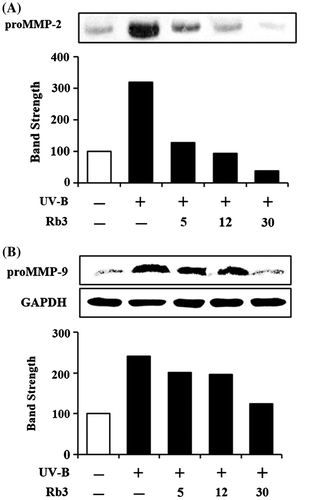
Since UV radiation is capable of enhancing production of certain MMP family members, such as MMP-9, MMP-2, and MMP-1, chronic exposure to UV radiation impairs normal architecture of the skin, leading to skin photoaging.Citation24) Rb3 attenuated the UV-B-induced proMMP-2 gelatinolytic activity in HaCaT keratinocytes in a concentration-dependent manner (Fig. (A)). Rb3 could similarly diminish the UV-B-induced proMMP-9 gelatinolytic activity in conditioned medium in a concentration-dependent fashion (Fig. (B)). Since Rb3 was able to diminish the UV-B-induced proMMP-2 and proMMP-9 gelatinolytic activities, the effects of Rb3 on proMMP-2 and proMMP-9 protein levels were examined using Western blotting analysis. As shown in Fig. (A), the UV-B radiation alone enhanced the proMMP-2 protein levels in conditioned medium, and Rb3, at the concentrations of 5, 12, and 30 μM, could reduce the UV-B-induced proMMP-2 protein levels in conditioned medium in a concentration-dependent manner. proMMP-9 protein levels in cellular lysates were also changed in a similar manner. The UV-B-induced proMMP-9 protein levels were concentration-dependently attenuated by Rb3 (Fig. (B)). Collectively, Rb3 is capable of down-regulating the UV-B-induced proMMP-2 and proMMP-9 in HaCaT keratinocytes through attenuating secretion of proMMP-2 and proMMP-9 proteins.
Fig. 4. Effect of Rb3 on the gelatinolytic activities of proMMP-2 (A) and proMMP-9 (B) in the conditioned media obtained from HaCaT keratinocyte cultures under irradiation with 70 mJ/cm2 UV-B radiation.
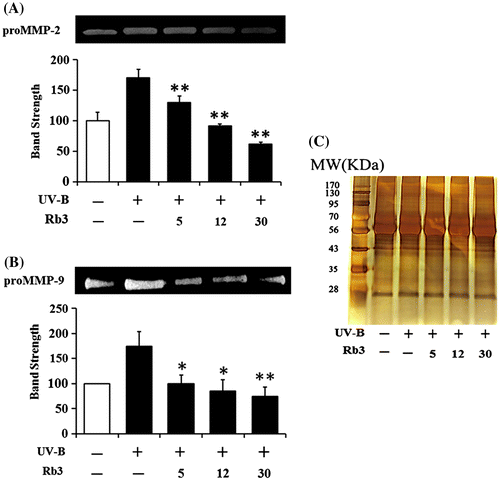
Fig. 5. Effect of Rb3 on proMMP-2 (A) protein levels in conditioned medium and proMMP-9 protein levels (B) in cellular lysate.
Enhancing effect of Rb3 on UV-B-reduced total GSH
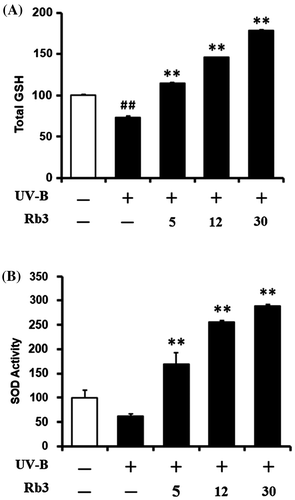
Consistently with a previous report,Citation25) total GSH contents were significantly diminished in HaCaT keratinocytes irradiated with 70 mJ/cm2 UV-B radiation (Fig. (A)). Pretreatment with 5, 12, and 30 μM Rb3 enhanced the total GSH levels by 1.6-, 2.0-, and 2.4-fold, respectively, compared to that of the irradiated HaCaT keratinocytes that did not receive the treatment (Fig. (A)). In brief, these findings indicate that the GSH-enhancing effects of Rb3 might possibly combat the deleterious effects of UV-B irradiation.
Fig. 6. Effect of Rb3 on total glutathione (GSH, (A)) and superoxide dismutase (SOD, (B)) activity levels in cellular lysates prepared from HaCaT keratinocyte cultures under irradiation with 70 mJ/cm2 UV-B radiation.
Enhancing effect of Rb3 on UV-B-reduced total SOD activity
Total SOD activity tended to be diminished by irradiation with 70 mJ/cm2 UV-B radiation in HaCaT keratinocytes (Fig. (B)). This diminishment could be overcome by the pretreatment with Rb3 (Fig. (B)). Rb3 at 5, 12, and 30 μM could enhance SOD activities by 2.7-, 4.1-, and 4.6-fold, respectively, compared to that of the UV-B-irradiated HaCaT keratinocytes without the reception of Rb3 treatment (Fig. (B)). Taken together, Rb3 is capable of enhancing SOD activity diminished under UV-B irradiation in HaCaT keratinocytes.
Discussion
As humans are getting older, their skins usually undergo photoaging which is mainly caused by repeated exposure to UV radiation, in addition to chronological aging. UV-A and UV-B radiation from sunlight is believed to be responsible for approximately 90% of skin photoaging.Citation26,27) They disturb cellular redox homeostasis systems in skin cells, and bring about an oxidative stress caused by imbalanced pro-oxidant processes and antioxidant defense systems, which results in skin photoaging. UV-B radiation induces ROS elevation, which causes oxidative damage in epidermal and dermal cells, and severe oxidative stress may give rise to apoptotic or necrotic death in skin cells.Citation28) An elevation in intracellular ROS levels, leading to oxidative stress, is thought to trigger an initiating step of skin photoaging.
Enhanced ROS levels due to UV radiation bring about production and secretion of MMPs, including MMP-1, MMP-2, MMP-3, MMP-9, and MMP-13, in epidermis and dermis, which contribute to skin damage and photoaging.Citation29,30) Some MMPs are generated as latent precursor forms (proMMPs). For example, MMP-2 (64 kD) is secreted as proMMP-2 (72 kD) and located on the cell surface and needs activation to exert its catalytic function.Citation31) After proMMP-2 is recruited to the cell surface by interacting with tissue inhibitor of metalloproteinase-2 (TIMP-2) bound to membrane type 1 MMP (MT1-MMP) by forming a ternary complex, it is activated on the cell surface by free MT1-MMP closely located to the ternary complex.Citation32) MT1-MMP is currently known to be the most common physiological activator of proMMP-2.Citation33) Type I collagen stimulates proteolytic activation of constitutively secreted proMMP-2 on the cell surface.Citation34) UV-A activates MMP-1 through the involvement of oxidative stress under which ROS are excessively produced and/or antioxidant functions are depleted in the skin cells.Citation17) Since skin photoaging is initiated by elevation of ROS level and/or MMP activity, blocking of their elevation has been suggested to be one of strategies to protect skin photoaging. Since skin photoaging is also a leading cause of skin cancer, its blocking may significantly reduce skin cancer risk. Several extracts of natural origin and natural constituents have already shown to have such blocking activities. For example, an extract of Butea monosperma flowers diminishes the secretion of MMP-1, MMP-2, MMP-9, and MMP-10 in UV-B-treated normal human epidermal keratinocytes (NHEKs).Citation35)
In the present work, the UV-B irradiation at the intensity of 70 mJ/cm2 significantly enhanced the ROS levels in HaCaT keratinocytes, which was detected using both fluorometric and confocal microscopic analyses. Together with the previous findings, the current finding reconfirms that the cellular damages caused by UV-B radiation are mediated by intracellular ROS. Although Rb3 was identified to scavenge the UV-B-induced ROS in HaCaT keratinocytes in this work, mechanism(s) responsible for the ROS-scavenging effect of Rb3 currently remain unknown. One possibility is that Rb3 suppresses the cellular procedure(s) responsible for the production of ROS caused by UV-B radiation. Another possibility is that Rb3 directly interacts with ROS although seemingly improbable. A third possibility is that Rb3 induces or activates antioxidant components which can scavenge ROS. This third possibility might be indirectly supported by the previous finding that Rb2, one other PPD-type ginsenoside, activates the SOD1 gene through enhancing the specific binding of transcription factor AP2 to the proximal promoter.Citation36) It could be also supported by the enhancing effects of Rb3 on total GSH and SOD activity, identified in this work. In the present work, when cultured HaCaT keratinocytes were subjected to UV-B radiation, proMMP-2 and proMMP-9 activities and secretion were supposedly enhanced, and the UV-B-induced proMMP-2 and proMMP-9 was found to be attenuated by Rb3 in a concentration-dependent manner. Although the precise mechanism(s) on the attenuating properties of Rb3 on proMMP-2 and proMMP-9 elevation in UV-B-irradiated HaCaT keratinocytes remain elusive, scavenging of ROS by Rb3 would be a preceding step. The attenuated ROS levels, caused by Rb3, might subsequently suppress UV-B-induced proMMP-2 and proMMP-9 enhancements in HaCaT keratinocytes, implying a plausible skin anti-photoaging property of Rb3.
In this work, we chose to measure proMMP-2 and proMMP-9, the precursor forms of MMP-2 and MMP-9, at both activity and protein levels to detect the suppressive effects of Rb3 on the MMPs in the UV-B-irradiated HaCaT keratinocytes. Based on the previous finding that UV-B irradiation significantly induces proMMP-2 and proMMP-9 in HaCaT keratinocytes,Citation15) the elevation of proMMP-2 and proMMP-9 productions is supposed to be involved in the UV-B-induced skin photoaging. This might be based on the fact that elevations in the production and subsequent secretion of proMMP-2 and proMMP-9 by UV-B irradiation in keratinocytes leads to the enhancements in their gelatinolytic activities in conditioned medium. Compared to the proMMP-2 and proMMP-9 activity bands, the MMP-2 and proMMP-9 activity bands appeared very weak, which could be sometimes negligible or even absent under our zymographic conditions (data not shown). This finding could confirm that proMMP-2 and proMMP-9 possess the partial gelatinolytic activities. It might also mean that the proMMP-2 and proMMP-9 activity levels are comparable to their total proMMP-2/MMP-2 and proMMP-9/MMP-9 activity pools, respectively. The corresponding results could be obtained from the Western blotting analysis. These results strongly suggest that UV-B irradiation up-regulates both proMMP-2 and proMMP-9 at the production levels, rather than at their activation stages. They also made us detect proMMP-2 and proMMP-9, the predominant forms, rather than MMP-2 and MMP-9, to evaluate the suppressive effects of Rb3 on the induction of proMMP-2 and proMMP-9 in the UV-B-irradiated HaCaT keratinocytes. However, the effects of Rb3 on the activation of proMMP-2 and proMMP-9 would be very interesting in the future works.
GSH is a principal small-molecular antioxidant which plays central roles in maintaining cellular redox homeostasis and protecting against oxidative stress and injury. UV-B radiation-induced GSH depletion in HaCaT keratinocytes mimics the pathogenesis of several cutaneous disorders. Decreased activity of γ-glutamylcysteine synthetase, a rate-limiting enzyme in GSH biosynthesis, and diminished cysteine uptake through the functional inhibition of the system Xc(-), a cysteine transporter on the cell membrane,Citation25) are implicated in pathogenesis. Indeed, inactivation of the cysteine transporter system is suggested to be a major contributor to UV-B-induced diminishment of GSH levels in HaCaT keratinocytes.Citation25) GSH depletion was also shown in a commercially available reconstituted human epidermis model of UV-B radiation.Citation37) When mouse lenses are exposed to UV-B radiation, simultaneous attenuation in both ATP and GSH levels, which could be prevented by caffeine, was observed.Citation38) In the aquatic worm tubifex, UV-B radiation induces production of singlet oxygen, superoxide anions, and hydroxyl radicals, and diminishes levels of GSH, DNA, RNA, and protein.Citation39) But, the GSH-enhancing effects of ginsenosides have not been clearly detected in the experiments using UV-B irradiation.
Eukaryotes produce three different SOD isozymes which regulate intracellular superoxide anion () levels by catalyzing the dismutation of
to H2O2; Cu/ZnSOD in cytosol, MnSOD within mitochondrial matrix, and an extracellular form similar to Cu/ZnSOD.Citation40) Based upon immunohistochemical studies, it was revealed that Cu/ZnSOD is chiefly expressed in keratinocytes.Citation41) Cu/ZnSOD exhibits a crucial defensive role against UV-B-induced injury of HaCaT keratinocytes.Citation42) In the present work, Rb3 was found to enhance SOD activity decreased in keratinocytes under UV-B-induced oxidative stress, which might partly support its skin anti-photoaging properties.
The results presented herein imply that Rb3 overcomes GSH and SOD depletion caused by exposure to UV-B radiation. Although the mechanisms of these overcoming activities remain unknown, enhancement of antioxidant components, such as GSH and SOD, might be an initiating event that leads to scavenging of ROS in epidermal keratinocytes. This finding partly proposes that the skin anti-photoaging activities of Rb3 are based upon activation or induction of antioxidant components which can abolish ROS.
HaCaT cells, as the human transformed and immortalized keratinocytes, have been known to be different from NHEKs in several molecular aspects. NHEKs are more resistant to hydrogen peroxide-induced oxidative stress than HaCaT cells.Citation43) Disruption of the constitutive NF-κB activity, which is activated by UV-B radiation and inflammatory tumor necrosis factor-α, in HaCaT cells gives rise to alterations in NF-κB signaling that are more consistent with the NF-κB signaling pathway in NHEKs.Citation44) The protection mediated by p38 which is markedly activated by UV radiation results from the cytoplasmic retention of p53 following its phosphorylation and stabilization, which subsequently its nuclear functions, in NHEKs, while p53 remains confined to the nucleus and cell survival is not affected by inhibition of p38 in HaCaT cells expressing a mutated form of p53 that probably affects its configuration.Citation45) Plant polyphenols or their combination with UV-A radiation or lipopolysaccharide induces strong up-regulation of stress responses in HaCaT cells but not in NHEKs.Citation46) A differential inducibility of the cell antioxidant networks in HaCaT cells and NHEKs results in a differential cellular level of ROS, explaining their different radiosensitivities.Citation47) However, the anti-apoptotic and antioxidant effects of 5,7,3′,4′-tetrahydroxyflavanone (eriodictyol) was similarly confirmed in UV-induced cell death of both HaCaT cells and NHEKs.Citation48) Considering differences or similarities between HaCaT cells and NHEKs, the skin anti-photoaging effect of Rb3 may need to be confirmed in NHEKs in the future approaches.
In conclusion, the potential skin anti-photoaging activity of Rb3 was recognized in HaCaT keratinocytes exposed to UV-B radiation. Rb3 suppresses the UV-B-induced intracellular ROS levels and production and secretion of proMMP-2 and proMMP-9, while it enhances the UV-B-depleted total GSH levels and SOD activity. Rb3 might elicit its skin anti-photoaging property through the down-regulation of UV-B-induced proMMP-2 and proMMP-9 based upon an ROS-dependent mechanism, which results from restoration of total GSH and SOD activity depleted under UV-B irradiation. Elucidation of precise mechanism(s) on these effects of Rb3 will require further experimental approaches. Rb3 would be a plausible candidate used as a natural resource for manufacturing anti-photoaging cosmetics which may have similar or enhanced activities with fewer side effects.
Author contributions
C.L. and K.K. designed the research and wrote the paper. S.O. and Y.O. performed the experiments. I.R. analyzed the data.
Disclosure statement
No potential conflict of interest was reported by the authors.
Funding
This work was supported by Korea Healthcare Technology R&D Project, Ministry of Health & Welfare, Republic of Korea [grant number A103017].
Related Research Data
References
- Shi Y, Sun CJ, Zheng B, Li Y, Wang Y. Simultaneous determination of nine ginsenosides in functional foods by high performance liquid chromatography with diode array detector detection. Food Chem. 2010;123:1322–1327.
- Shi Y, Han B, Yu X, Qu S, Sui D. Ginsenoside Rb3 ameliorates myocardial ischemia-reperfusion injury in rats. Pharm. Biol. 2011;49:900–906.
- Ma L, Liu H, Xie Z, et al. Ginsenoside Rb3 protects cardiomyocytes against ischemia-reperfusion injury via the inhibition of JNK-mediated NF-κB pathway: a mouse cardiomyocyte model. PLoS One. 2014;9:e103628.
- Wang T, Yu XF, Qu SC, Xu HL, Sui DY. Ginsenoside Rb3 inhibits angiotensin II-induced vascular smooth muscle cells proliferation. Basic Clin. Pharmacol. Toxicol. 2010;107:685–689.
- Wang T, Yu X, Qu S, Xu H, Han B, Sui D. Effect of ginsenoside Rb3 on myocardial injury and heart function impairment induced by isoproterenol in rats. Eur. J. Pharmacol. 2010;636:121–125.
- Jiang S, Miao B, Song X, Jiang Z. Inactivation of GABA(A) receptor reduces ginsenoside Rb3 neuroprotection in mouse hippocampal slices after oxygen-glucose deprivation. J. Ethnopharmacol. 2011;133:914–916.
- Cui J, Jiang L, Xiang H. Ginsenoside Rb3 exerts antidepressant-like effects in several animal models. J. Psychopharmacol. 2012;26:697–713.
- Birch-Machin MA, Swalwell H. How mitochondria record the effects of UV exposure and oxidative stress using human skin as a model tissue. Mutagenesis. 2010;25:101–107.
- Barresi C, Stremnitzer C, Mlitz V, et al. Increased sensitivity of histidinemic mice to UVB radiation suggests a crucial role of endogenous urocanic acid in photoprotection. J. Invest. Dermatol. 2011;131:188–194.
- Ikehata H, Ono T. The mechanisms of UV mutagenesis. J. Radiat. Res. 2011;52:115–125.
- Peres PS, Terra VA, Guarnier FA, Cecchini R, Cecchini AL. Photoaging and chronological aging profile: understanding oxidation of the skin. J. Photochem. Photobiol. B. 2011;103:93–97.
- Aitken GR, Henderson JR, Chang SC, McNeil CJ, Birch-Machin MA. Direct monitoring of UV-induced free radical generation in HaCaT keratinocytes. Clin. Exp. Dermatol. 2007;32:722–727.
- Curran S, Murray GI. Matrix metalloproteinases in tumour invasion and metastasis. J. Pathol. 1999;189:300–308.
- Chae S, Piao MJ, Kang KA, et al. Inhibition of matrix metalloproteinase-1 induced by oxidative stress in human keratinocytes by mangiferin isolated from Anemarrhena asphodeloides. Biosci. Biotechnol. Biochem. 2011;75:2321–2325.
- Kim MS, Oh GH, Kim MJ, Hwang JK. Fucosterol inhibits matrix metalloproteinase expression and promotes type-1 procollagen production in UVB-induced HaCaT cells. Photochem. Photobiol. 2013;89:911–918.
- Lee YM, Kim YK, Kim KH, Park SJ, Kim SJ, Chung JH. A novel role for the TRPV1 channel in UV-induced matrix metalloproteinase (MMP)-1 expression in HaCaT cells. J. Cell. Physiol. 2009;219:766–775.
- Pygmalion MJ, Ruiz L, Popovic E, et al. Skin cell protection against UVA by sideroxyl, a new antioxidant complementary to sunscreens. Free Radical Biol. Med. 2010;49:1629–1637.
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976;72:248–254.
- Royall JA, Ischiropoulos H. Evaluation of 2′,7′-dichlorofluorescin and dihydrorhodamine 123 as fluorescent probes for intracellular H2O2 in cultured endothelial cells. Arch. Biochem. Biophys. 1993;302:348–355.
- Freshney RI. Culture of animal cells: a manual of basic technique. 4th ed. New York (NY): Wiley-Liss Press;1994.
- Kleiner DE, Stetler-Stevenson WG. Quantitative zymography: detection of picogram quantities of gelatinases. Anal. Biochem. 1994;218:325–329.
- Nakagawa K, Saijo N, Tsuchida S, et al. Glutathione-S-transferase pi as a determinant of drug resistance in transfectant cell lines. J. Biol. Chem. 1990;265:4296–4301.
- Lee YY, Kim HG, Jung HI, et al. Activities of antioxidant and redox enzymes in human normal hepatic and hepatoma cell lines. Mol. Cells. 2002;14:305–311.
- Quan T, Qin Z, Xia W, Shao Y, Voorhees JJ, Fisher GJ. Matrix-degrading metalloproteinases in photoaging. J. Invest. Dermatol. Symp. Proc. 2009;14:20–24.
- Zhu M, Bowden GT. Molecular mechanism(s) for UV-B irradiation-induced glutathione depletion in cultured human keratinocytes. Photochem. Photobiol. 2004;80:191–196.
- Jenkins G. Molecular mechanisms of skin ageing. Mech. Ageing Dev. 2002;123:801–810.
- Rittié L, Fisher GJ. UV-light-induced signal cascades and skin aging. Ageing Res. Rev. 2002;1:705–720.
- Rabe JH, Mamelak AJ, McElgunn PJ, Morison WL, Sauder DN. Photoaging: mechanisms and repair. J. Am. Acad. Dermatol. 2006;55:1–19.
- Fisher GJ, Wang ZQ, Datta SC, Varani J, Kang S, Voorhees JJ. Pathophysiology of premature skin aging induced by ultraviolet light. N. Engl. J. Med. 1997;337:1419–1428.
- Lee YM, Kang SM, Lee SR, et al. Inhibitory effects of TRPV1 blocker on UV-induced responses in the hairless mice. Arch. Dermatol. Res. 2011;303:727–736.
- Ellenrieder V, Alber B, Lacher U, et al. Role of MT-MMPs and MMP-2 in pancreatic cancer progression. Int. J. Cancer. 2000;85:14–20.
- Nagase H. Cell surface activation of progelatinase A (proMMP-2) and cell migration. Cell Res. 1998;8:179–186.
- Sato H, Takino T. Coordinate action of membrane-type matrix metalloproteinase-1 (MT1-MMP) and MMP-2 enhances pericellular proteolysis and invasion. Cancer Sci. 2010;101:843–847.
- Morley ME, Riches K, Peers C, Porter KE. Hypoxic inhibition of human cardiac fibroblast invasion and MMP-2 activation may impair adaptive myocardial remodelling. Biochem. Soc. Trans. 2007;35:905–907.10.1042/BST0350905
- Krolikiewicz-Renimel I, Michel T, Destandau E, et al. Protective effect of a Butea monosperma (Lam.) Taub. flowers extract against skin inflammation: antioxidant, anti-inflammatory and matrix metalloproteinases inhibitory activities. J. Ethnopharmacol. 2013;148:537–543.10.1016/j.jep.2013.05.001
- Kim YH, Park KH, Rho HM. Transcriptional activation of the Cu,Zn-superoxide dismutase gene through the AP2 site by ginsenoside Rb2 extracted from a medicinal plant, Panax ginseng. J. Biol. Chem. 1996;271:24539–24543.
- Meloni M, Nicolay JF. Dynamic monitoring of glutathione redox status in UV-B irradiated reconstituted epidermis: effect of antioxidant activity on skin homeostasis. Toxicol. In Vitro. 2003;17:609–613.
- Varma SD, Hegde KR, Kovtun S. UV-B-induced damage to the lens in vitro: prevention by caffeine. J. Ocul. Pharmacol. Ther. 2008;24:439–444.
- Misra RB, Babu GS, Ray RS, Hans RK. Tubifex: a sensitive model for UV-B-induced phototoxicity. Ecotoxicol. Environ. Saf. 2002;52:288–295.
- Frank S, Kämpfer H, Podda M, Kaufmann R, Pfeilschifter J. Identification of copper/zinc superoxide dismutase as a nitric oxide-regulated gene in human (HaCaT) keratinocytes: implications for keratinocyte proliferation. Biochem. J. 2000;346:719–728.
- Kobayashi T, Saito N, Takemori N, et al. Ultrastructural localization of superoxide dismutase in human skin. Acta Derm. Venereol. 1993;73:41–45.
- Sasaki H, Akamatsu H, Horio T. Protective role of copper, zinc superoxide dismutase against UVB-induced injury of the human keratinocyte cell line HaCaT. J. Invest. Dermatol. 2000;114:502–507.
- Liu L, Xie H, Chen X, et al. Differential response of normal human epidermal keratinocytes and HaCaT cells to hydrogen peroxide-induced oxidative stress. Clin. Exp. Dermatol. 2012;37:772–780.
- Lewis DA, Hengeltraub SF, Gao FC, Leivant MA, Spandau DF. Aberrant NF-κB activity in HaCaT cells alters their response to UVB signaling. J. Invest. Dermatol. 2006;126:1885–1892.
- Chouinard N, Valerie K, Rouabhia M, Huot J. UVB-mediated activation of p38 mitogen-activated protein kinase enhances resistance of normal human keratinocytes to apoptosis by stabilizing cytoplasmic p53. Biochem. J. 2002;365:133–145.
- Pastore S, Lulli D, Potapovich AI, et al. Differential modulation of stress-inflammation responses by plant polyphenols in cultured normal human keratinocytes and immortalized HaCaT cells. J. Dermatol. Sci. 2011;63:104–114.
- Isoir M, Buard V, Gasser P, Voisin P, Lati E, Benderitter M. Human keratinocyte radiosensitivity is linked to redox modulation. J. Dermatol. Sci. 2006;41:55–65.
- Lee ER, Kim JH, Kang YJ, Cho SG. The anti-apoptotic and anti-oxidant effect of eriodictyol on UV-induced apoptosis in keratinocytes. Biol. Pharm. Bull. 2007;30:32–37.10.1248/bpb.30.32