Abstract
A long-standing issue in connectionist research has concerned the manner in which the human brain contrives to bind values to variables, fillers to roles, and (sensory) features to features. This paper examines the comparative merits of two binding methods whose biological foundations have been empirically confirmed, namely, binding via synchronous firing and binding via conjunctive coding. Within the community of connectionist researchers, it is often asserted that synchronous binding is far more efficient, in terms of the number of neurons required, than conjunctive coding. Moreover, Singer (Neuron, 24, pp. 49–65, 1999) has argued that conjunctive coding is inadequate as a general method of binding, for several cognitively based reasons. This paper argues that the above-cited complaints against conjunctive binding are not decisive. It is further noted that synchronous firing is a surface phenomenon—one that requires an explanation in terms of underlying causes. A model is proposed herein that illustrates one way in which synchronous bindings could be engendered by a substratum that relies, ultimately, upon conjunctive binding nodes. It is demonstrated that such a substratum would require a feasibly small number of conjunctive neurons.
Introduction
A long-standing issue in connectionist research has concerned the manner in which the human brain contrives to bind values to variables, fillers to roles, and (sensory) features to features. Closely related to this issue is the quest for an elegant and efficient solution to ‘variable binding’ in artificial connectionist networks. In this paper, I shall be concerned, ultimately, with biological neural networks, and for this reason the discussion will focus upon the two binding methods whose biological bases have been most directly confirmed. In brief, these two methods are: (a) conjunctive coding, where the binding of two elements is signalled by the cyclical firing of a particular node (realized by one or more neurons) that receives input from other nodes that represent the elements which are to be bound together (or ‘conjoined’); and (b) synchronous firing, where distinct nodes (or distinct sets of neurons) are represented as being bound together when the distinct nodes fire in approximately synchronous cycles.Footnote†
The concepts of conjunctive coding and binding via synchronous firing will be elaborated in following sections, but for the present some observations may be helpful. The use of conjunctive coding as a method for binding values to variables and fillers to roles can be observed in several early connectionist models (cf. Feldman and Ballard Citation1982, Rumelhart et al. Citation1986, Shastri Citation1988) and has been especially valuable in network models of language parsing and/or acquisition (see Cottrell Citation1985, Stevenson Citation1994, Hadley and Hayward Citation1997, Hadley and Cardei Citation1999). Moreover, the binding of sensory features via explicit, conjunctive binding neurons has been empirically verified in studies of the visual cortex of mammals and other cortical regions (Singer Citation2003). Such binding of elements (whether they be features, concepts, variables, or roles) is known to be both reliable and fast. However, binding via conjunctive coding has often been criticized as being combinatorially explosive in the number of nodes (or neurons) required to effect all the bindings possible in a given domain. Unfortunately, this criticism has at times been made in a sweeping form, without regard to the fact that the number of conjunctive nodes required may merely be the simple product of the number of fillers and roles involved (the latter being feasibly sparse in non-perceptual domains). In any case, other criticisms of conjunctive coding have been published, and will be examined in what follows.
In contrast with conjunctive coding, synchronous firing, considered as a binding method, has in recent years attracted considerable notice, both because of its biological basis (cf. Singer Citation1999, Citation2003) and because it (purportedly) avoids the proliferation of nodes attributed to conjunctive coding. Evidence for the role of synchronously firing neurons in sensory feature binding is substantial (Singer Citation1999), and synchronous firing has been used as a basis for variable binding in recent models of inference, including SHRUTI (Shastri and Ajjanagadde Citation1993) and LISA (Hummel and Holyoak Citation1997).
Despite the considerable positive acclaim that synchrony has attracted as a solution to the binding problem, its presumed advantages over conjunctive coding might, for all we actually know, be illusory. I say this because synchronous neural firing cannot be regarded as a fundamental, unanalysable phenomenon. It is well known that the firing of an individual neuron is caused by underlying processes, both chemical and electrical. For this reason, among others, it would be scientifically incoherent to regard the systematic firing of neurons in synchrony as a fundamental, inexplicable phenomenon. Yet, the causes of synchronous firing in non-perceptual realms (which include language comprehension, reasoning, and other forms of higher cognition) are not known at present. The few causal indications that are known are compatible with the following supposition, where variable binding is concerned: apart from spurious coincidences, neurons fire in synchrony only when they are each connected (either directly or indirectly) to explicit conjunctive binding nodes (dub them CBN) whose cyclical firing engenders synchrony in the firing cycles of neurons that the CBNs are connected to. The simplest form of this supposition would require one conjunctive binding node for each pair of synchronously firing nodes. (Of course, alternative suppositions, not involving CBNs, may also be consistent with the known evidence.)
In section 3, it is argued that this (hitherto ill-favoured) supposition may well provide a promising direction for research, within the realm of role–filler binding at the very least (e.g. where thematic roles are bound to concepts that fill those roles). I would emphasize, however, that the entire discussion concerns representational processes within short-term memory, for it is well known that the brain cannot sustain enough distinct synchronous firing patterns (simultaneously) to begin to explain the number of propositions stored in long-term memory (cf. Sougné Citation2001). Regardless of that, it should be borne in mind that until the causes of synchronous firing are understood, we are not in a position to judge the number of neurons involved in the genesis of synchrony firing. In consequence, we are not entitled to assume the greater efficiency of synchronous binding schemes.
In passing, I freely acknowledge that, setting aside claims about relative efficiency, there exist interesting, useful systems that assume synchronous firing as a basic phenomenon for the purpose of modelling some moderately abstract cognitive capacity (e.g. see Shastri and Ajjanagadde Citation1993). It can, of course, be perfectly reasonable to ignore the underlying causes of synchronous firing when one's goal is to model mental processes at levels of abstraction higher than those which will concern us here. Obviously, however, one should not construe the success of such models as evidence that conjunctive coding models are ill-founded. Moreover, when one's goal is to understand the causes of synchronously firing neurons, it is entirely appropriate to delve into lower levels of explanation, where conjunctive binding may well be involved.
The plan for the remainder of this paper is as follows. In section 2, I examine Singer's (Citation2003) tri-fold critique of conjunctive coding as a potential explanation for the binding of sensory features in the realm of perception. It will be argued that the difficulties noted by Singer may not be germane to the phenomena of value–variable binding and filler–role binding, both of which occur in reasoning, language comprehension, and other cognitive domains. (These last two forms of binding are strongly analogous, and hereafter they will, for brevity, simply be referred to as ‘filler–role’ binding.) I shall also consider whether Singer's critique is actually decisive in the domain of sense perception. While an elaboration of Singer's tri-fold critique must await exposition in section 2, here is a glimpse.
| 1. | Within the realm of perception, the number of perceivable combinations of sensory features is too vast to be explained in terms of conjunctive binding; there could not be a separate conjunctive binding node for each such combination. | ||||
| 2. | Where binding is achieved solely via conjunctive neurons, there will be serious unresolved problems whenever the representation of nested relationships between components of complex perceived objects is involved. | ||||
| 3. | The recognition and representation of novel objects and novel relations among features would be highly problematic if conjunctive nodes were the sole binding mechanism. For in this case, ‘rapid reconfiguration of input connections to previously uncommitted cells’ (Singers Citation2003) would appear necessary. | ||||
I wish to emphasize, though, that there may well exist alternative explanations for synchronously firing neurons, which do not require the existence of conjunctive binding nodes. No claims for exclusivity or general superiority are made here. Nevertheless, care is needed to avoid those ‘explanations’ that involve subtle circular reasoning or some form of regress. For example, it will not suffice, in general, to suppose that a ‘value-node’ and a ‘variable-node’ should fire in synchrony only after a direct, weighted link has been trained between those two nodes. For the problem yet arises, how is it that only those nodes that ought to fire in synchrony (given the temporary task at hand) happen to have previously formed the proper trained associations? This is especially problematic in light of the fact that, on one occasion, two nodes ought to fire synchronously, while on another occasion, very soon thereafter, they should not.
Criticisms of conjunctive binding
We turn now to consider three arguments, presented by Singer Citation(2003), to the effect that conjunctive binding (or binding via conjunctive coding) cannot be the sole form of binding employed by the human brain. Before delving into details, I wish to emphasize that, where conjunctive coding is concerned, Singer does not regard all of his points as decisive. Nevertheless, I believe they raise several important issues, which warrant a further exploration of alternative hypothesis. Also, in considering these arguments, it will be helpful to distinguish between their applicability in the domain of sensory feature processing versus other cognitive domains.
The first argument
The first of Singer's arguments is framed entirely in terms of the combinations of features that can occur within intra- and cross-modal sensory processing. Singer rightly emphasizes that the number of such combinations is exceedingly large. Whether the number is so vast as to exceed the number of neurons actually available within the sensory cortices is a question not explicitly addressed by Singer. Moreover, its answer is not obvious. In any case, Singer is by no means alone in assigning great importance to parsimony in this issue. Others, as early as Hinton et al. Citation(1986), have produced similar arguments against conjunctive coding as an explanation for recognition of combinations of sensory features. However, while parsimony may once have been a legitimate issue in the realm of artificial networks (due to small computer memories), its relevance is less clear where the sensory cortices are concerned.
Nevertheless, for argument's sake, I am happy to concede that if the recognition of every single combination of sensory features were to require a dedicated CBN (conjunctive binding node), then some other method of binding must also be at work in the realm of sense perception. (Admittedly, I later suggest a way in which CBNs might provide the underpinnings for recognition even in this realm, although not via a dedicated CBN for every such combination.) In any case, for the present I wish to argue that, contrary to received opinion, the ‘parsimony argument’ may not be decisive in the realm of thought and language, where role–filler binding is widely believed to be involved.
It is of course clearly true that not every combination of concepts that comprise a proposition that a person (P) could think (or entertain, or understand via hearing a sentence) could be represented by a distinct neuron in P's brain. Even though a human can, in practice, entertain only a finite number of such propositions, the number of these expressible by sentences containing 20 words or fewer is absolutely prohibitive, as Fodor and Pylyshyn Citation(1988) have amply demonstrated. Thus, there could not be a CBN corresponding to each combination of concepts involved in each such proposition. Moreover, one may be hard-pressed to find any cognitive researcher who would now advocate this kind of localist approach to the representation of sentence meanings or complete thoughts (expressed propositions).
On the other hand, many researchers working in the realm of sentential meaning (pro-positional) representation embrace the view that mental representations of propositions involve some degree of structure, where constituents of these structures are concepts (fillers) playing particular roles. By means of a humanly feasible number of concepts (or fillers) and a feasible number of roles, one may create a staggering number of complete propositions.
The number of individual concepts that an adult human would typically employ is arguably not much greater than that agent's ‘receptive vocabulary’ (i.e. those words whose meanings are recognized). Admittedly, some vocabulary words have multiple meanings, and so express multiple concepts. However, we can also find distinct words that express the same concept (e.g. ‘tadpole’ and ‘pollywog’). Estimates of the size of the receptive vocabulary of human adults vary, but nearly all estimates indicate that it is rare for even university graduates to possess receptive vocabularies that include more than 30 000 word stems (e.g. ‘frog’ is the stem for ‘frogs’, ‘see’ is the stem for ‘seen’). Indeed, Goulden et al. Citation(1990) placed the estimate for a typical university graduate at 20 000 word families (where a word family contains the word stem and its morphological relatives, including plurals, past tenses, etc.). Arguably, the plurals, past tenses, etc. express concepts that are decomposable into the base concept, connoted by the stem, together with a small number of multi-purpose, modifier concepts that are commonly reused with other stem words (e.g. the concept of ‘plural’ would be such a modifier). One cannot say with certainty, but if we place an upper-bound estimate of 40 000 on the number of concepts known by even a highly literate adult, we would, in all likelihood, not be underestimating. I shall, then, assume the figure of 40 000. (N.B. this figure is assumed for those concepts that are each, respectively, expressed by a given word within a given natural language. Multi-word phrases can, of course, collectively express an even larger number of complex concepts, but such phrases involve multiple roles. The presence of multiple roles would obviously enable the impromptu creation of a very large number of combinations, but my concern is with concepts that would typically bind with one role at a time. In any case, even if one were to accept the very high estimates of, say, Thorpe and Imbert Citation(1989) (in the range of 60 000 –100 000 concepts), this would at worst magnify the required number of CBNs only by a factor of 2.5. As will emerge, this would not create a problem; cf. section 3.3.)
The number of conceptual roles involved in the formation of propositions is a vexed question, and will vary according to which theory or scheme we accept. However, as a first pass, let us consider a simple (though ultimately too simple) scheme inspired by a symbolic encoding method introduced by Smolensky Citation(1990). Following this scheme, we require an upper limit on the number of concepts that could be sequentially expressed by the words in a sentence that a person both hears and understands. Surely, it would be rare for a person to be able to grasp fully any proposition expressed by a spoken sentence containing more than 100 words, but to be very safe, let us set an upper limit of 150 words. Conceptual roles may then be identified with the sequential position that each word in the sentence occupies. It is crucial to note that ‘conceptual role’ is being used here in a special sense. The phrase is chosen merely to convey that, on a given occasion, a concept plays the role of being expressed by a word occupying the Nth position in a given sentence. Thus, a maximum of 150 roles would be needed in the approach under consideration. (Caveat: the individuation of roles in terms of sequence position is due to Smolensky Citation(1990), and no claim is made here for the superiority of this strategy. Rather, I am adopting this approach pro tem, for purposes of illustration.)
Now, let us suppose that each of these 150 roles is locally represented by a unique node, and similarly for each of the 40 000 possible concepts that I have assumed. Suppose also that each possible role-concept pair is connected to a unique binding node. To represent that a particular concept is playing a particular role on an occasion, the corresponding binding node is made reasonably active (i.e. it attains a sufficiently high firing rate). The maximum number of binding nodes required by this scheme is clearly just the product of the number of roles and the number of fillers (or 6 million in this case). Singer, in his discussion of conjunctive coding, identifies binding nodes with single neurons, and cites abundant evidence for the existence of such conjunctive cells within the sensory cortices. However, even if we were to assume that each of 6 million nodes (just mentioned) is realized in the brain by a distinct set of 10 neurons, we would still require just 60 million neurons to serve as binding nodes. When comparable assumptions are made for the 150 roles and 40 000 concept nodes, the total is under 61 million neurons.
Now, 61 million is not an absurd number of neurons to be devoted to the representation of all the possible propositions that a human could, in practice, understand. We must bear in mind, after all, that estimates of the number of neurons in the human cerebral cortex fall in the range of 15 billion (Haug Citation1986), and that much of the processing within the cerebral cortex occurs via parallel means. Moreover, we need not suppose that each binding node is dedicated solely to representing the conjunction of a given role and concept. It may well be that the representation of each such conjunction requires not only the activation of the given binding node, but also the concurrent activation of the given role and concept nodes. When either of the latter two are inactive, the given binding node may be free to participate in other forms of processing. At least, this is true for all that has been shown to the contrary.
It must be acknowledged, however that the scheme just developed does not accord with the kinds of structures that many researchers in mental representation commonly espouse. Although Fodor's (Citation1975) Language of Thought thesis is not universally accepted, there are many who agree that internal representations of propositions must be formed according to some kind of syntax, and that the structure of such representations is more complex than a mere sequence of bound concepts and roles. Among those who are dealing with meaning compositionality in complex sentences, it is often argued that something akin to constituent-oriented, mental tree structures must be involved if crucial aspects of thought compositionality are to explained (see Fodor and Pylyshyn Citation(1988) and Fodor and McLaughlin Citation(1990) for arguments). Of course, many connectionists dispute the Language of Thought approach, and believe that distributed representations will suffice to explain meaning compositionality. Debates on this issue persist, but in this paper I strive to remain neutral (however, see Hadley (Citation2003, Hadley (Citation2004) for a discussion of the merits of distributed representations vis-à-vis systematicity).
Whether or not the Language of [structured] Thought adherents will ultimately prevail, the view that internal representations of propositions involve tree structures, where leaf nodes are associated with thematic roles (such as ‘agent’, ‘action’, ‘instrument’, etc.), remains a theoretical possibility. Certainly, Singer's arguments are not intended to exclude this option, and there are connectionist language interpretation systems that create ‘internal’ parse-tree structures in response to sentential, symbolic input. Three such systems, which all use conjunctive binding nodes (that are triggered by spreading activation), are given in Cottrell Citation(1985), Stevenson Citation(1994) and Hadley and Cardei Citation(1999).
In the remainder of this section, it will be assumed that such tree-based propositional representations are at least a viable possibility. Also, following the lead of many other researchers in the field of mental representation (including those just cited), I shall assume that the kind of parse trees employed in natural language analysis (see O'Grady Citation1997: pp, 136, 175, 271, among others) provide at least a useful analogy for the tree structures of mental representations. Given these premises, I argue that a feasible number of CBNs could provide a basis for the binding of concepts to thematic roles (at leaf nodes) and for the binding of sub-trees to higher levels within a tree. The purpose of these arguments is not only to address issues related to the first of Singer's points, but also to address his second argument, which holds that ‘unresolved problems [for conjunctive binding] arise with the representation of nested relations among the components of composite objects, such as visual scenes or sentences’ (Singer Citation2003). Although my explanations are tailored towards the representations of propositions expressed by sentences, they may well be applicable to ‘nested relations among the components of’ visual scenes, if these nested (recursive) relations in fact provide the basis for hierarchical structures.
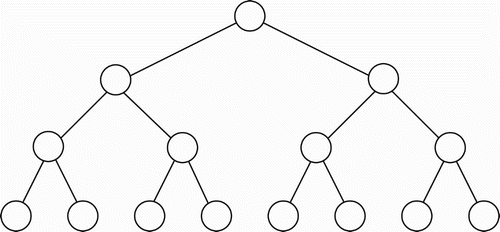
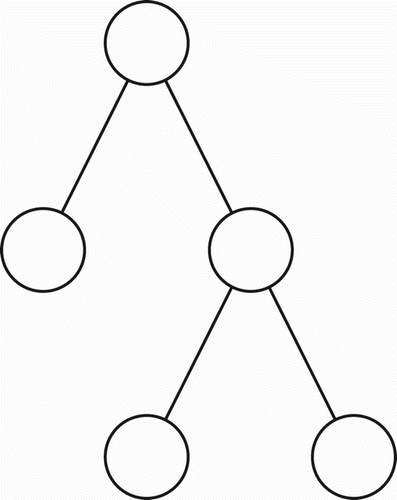
Let us assume, at the outset, that tree-like propositional representations are an assembly of the simplest possible binary sub-trees, one of which is shown in . For simplicity, I refer to each sub-tree of this type as a triplet (it contains three nodes). Note that any complete binary tree structure, such as that shown in , may be viewed as a composite of such triplets, provided the root of one triplet is permitted to coincide with the leaf node of another triplet. Note also that, within modern linguistic theory, it is commonly held that adequate grammars of natural languages will yield binary parse trees (O'Grady Citation1997 p. 248). Thus, a syntactic parse tree for any well-formed sentence of natural language will be binary.
Figure 1. A triplet; the simplest form of binary sub-tree.
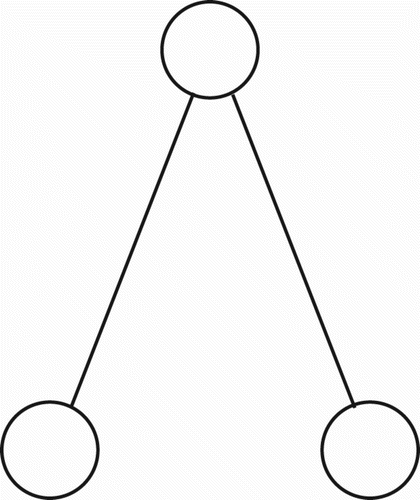
Figure 2. A complete binary tree having eight leaf nodes.
Now, let us further suppose (what is at least possible) that within the human brain there exist (whether innately or otherwise) a plethora of neural structures having the functionally abstract form of a triplet (). That is, although, biologically speaking, such structures would contain finer physical detail, in terms of functionality the structure would perform as a triplet. Also, we may suppose that the ‘leaf’ of one triplet may be connected to the ‘root’ of another triplet by having both the leaf and root nodes make pre-synaptic connections on an intermediate binding node (a CBN). For example, displays a complete binary tree whose internal nodes should all be regarded as virtual; each internal node being implemented by a structure as shown in . The diamond-shaped node in is intended as a CBN whose activation is fed and triggered by spreading activation from the nodes just above and below it. The node above (i.e. A) would in reality be a leaf node of an upper triplet, and the node below (i.e. B) would in reality be the root of a triplet that could attach to node (A) when conditions warrant. Thus, the CBN between nodes A and B provides a coupling mechanism that can be effectively (i.e. sufficiently) activated or not, depending upon the manner in which activation is spread in response to, say, the parsing of a sentence being comprehended (see Stevenson Citation(1994), Hadley and Cardei Citation(1999) and Hadley and Hayward Citation(1997) for explanations of details of such activation spreading and binding).
Figure 3. The implementation of a virtual node. Node A is a leaf of one sub-tree and B is the root of another sub-tree. A and B both connect with a CBN that is represented by a diamond shape.

Now, consider the virtual binary tree shown in . When the internal nodes (X, Y) in that tree are expanded into their implemented forms, we obtain the situation shown in . Assume that on a given occasion, the leftmost CBN in is not effectively active, while the rightmost CBN is effectively active. In that case, from a functional standpoint, the bottom-left triplet in becomes inoperative (functionally detached) and we obtain the virtual tree shown in . In a similar manner, portions of larger, complete binary trees can be functionally absent on a given occasion. In general, the graph of any incomplete binary tree, up to a given depth, can be similarly produced by the mere expedient of slicing out portions of a complete binary tree that has an equal or greater depth. Since this is true, we could conjecture that a general binary tree template, along the lines of the ‘implemented tree’ in , exists (perhaps innately, perhaps not) within the brains of humans who competently use some natural language. Such a tree template would need to be of sufficient depth to ensure that the number of ultimate leaf nodes exceeds, or equals, the maximum number of concepts that a human could apprehend within a single proposition, corresponding to a single sentence. For simplicity, and to accommodate the worst case, I submit that 256 leaf nodes would be adequate. Thus, a complete binary tree, having eight levels below the ultimate root, should suffice for a general template.
Figure 4. A complete binary tree whose internal nodes (X and Y) are shown in their ‘virtual’ form.
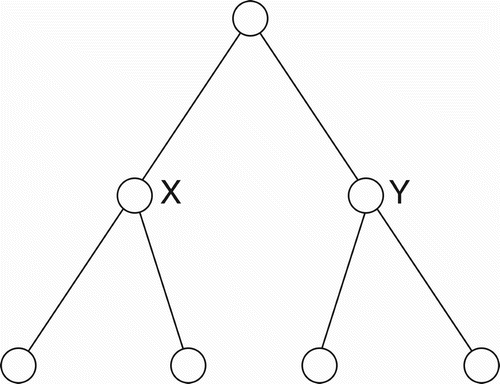
Figure 5. The virtual internal nodes of are here replaced by their ‘implemented versions’. That is, each of X and Y from would actually be a set of three nodes involving a CBN with linkage.
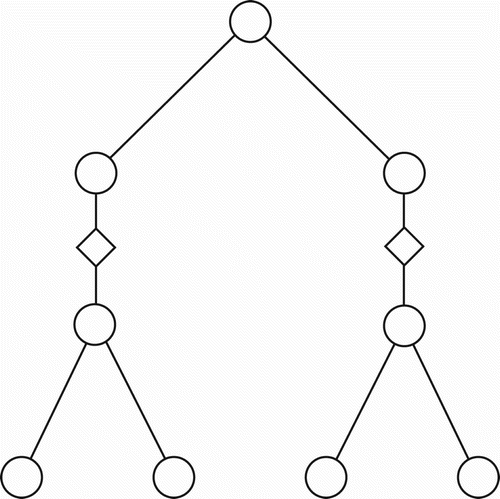
Figure 6. Displayed here is the result of removing the leftmost sub-tree of .
The reader will recall that within ordinary syntactic parse trees, internal nodes possess labels, such as NP (noun phrase). Such labels indicate that internal nodes play roles within the tree. Analogously, I shall regard internal nodes within our template as role nodes. For each such internal node, there could be an attached filler node, which would be the root of some triplet. (Recall that roots of triplets can make attachments, via CBNs, as shown in .) This is not to say that the brain would actually assign labels to nodes in any explicit way, or that labels could not be changed dynamically, if they exist at all.Footnote†
Concerning roles represented by leaf nodes, I would emphasize that the roles being hypothesized are thematic roles, and may be less numerous than the finely grained syntactic roles found in parse trees that grammarians construct. Speaking in terms of types of thematic role, one would be hard-pressed to find a linguistic theory that posited more than 100 thematic roles that a concept or word could play in a sentence. (Indeed, the same could be said of syntactic roles.) It is possible, of course, to individuate the roles that concepts could play in terms other than thematic roles, and some methods of individuation may generate a rather large number of possible roles. However, the roles that concern us ultimately pertain to the structural position that a concept would occupy in the propositional tree representations that are assumed here. For that purpose, I submit that thematic roles serve our purpose quite well. The specific content contributed by a concept towards a proposition is supplied by the concept itself; the role needs only to supply a bare minimum of content.
For example, in the sentence, ‘Mary teaches calculus’, there is no need to individuate the role played by the ‘Mary-concept’ so finely that we are positing a ‘teacher role’. It suffices to view Mary as playing the role of ‘agent’. The notion of teaching is supplied by the verb itself. Specifically, in this case, we could suppose that there exist generic role nodes for agent, action and patient, and that these correspond, in left–right order, to the three leaf nodes shown in . Nodes corresponding to the ‘Mary-concept’ and ‘teaching-concept’ would bind, respectively, with the agent and action role nodes. The integration of the concept and role nodes into a single coherent proposition is ensured by the relative positions of the given role nodes within a tree structure possessing a single root. Given this, there is no need for any trained associations to occur between the ‘teaching-concept’ and any given agent. However, since different types of verb concepts have differing thematic role requirements (as with transitive versus intransitive verbs), it may well be that as the verb concepts are learned, associations are acquired that link the concept node to the appropriate types of trees and leaf nodes to which the concept can bind. For further details on this general approach, see Hadley and Hayward Citation(1997) and Hadley and Cardei Citation(1999).
Returning now to binding issues, and having examined those binding nodes that permit tree structures to be assembled and dismantled, we next consider CBNs that permit individual concepts to become attached to leaf nodes. Recall that at the deepest level of the complete tree template, there are 256 leaf nodes. Each of these 256 nodes should have the capacity to bind with a concept that atomically occurs in a proposition being entertained or comprehended. For example, the proposition expressed by ‘mice chased by cats are often caught’ may involve concepts not explicitly expressed by words in the quoted sentence, but the total number of concepts involved appears to be less than 20. In any case, some comparatively small subset of the 256 leaf nodes would need to bind with the concepts so expressed, and each leaf node must have the capacity to bind (on various occasions) with differing concepts.
For simplicity of counting, and to accommodate the worst case, I shall assume that each leaf node should be capable of binding with each of the 40 000 concepts that I have assumed a human might possess. (In reality, of course, leaf nodes may have typed roles and might bind only with concepts of a certain type.) Thus, given this worst case assumption, each of the 256 leaf nodes will require one CBN for each of the 40 000 concept nodes in question. This means that roughly 10 million CBNs would be required for binding concepts at the deepest level of the tree. Also, depending upon the particular manner in which propositions are ‘constructed’ by the brain, we must allow for the possibility that concept nodes would become bound to tree nodes at shallow levels (since the deletion of sub-trees within the general template could cause an intermediate-level node to become a leaf node). The total number of internal nodes within the general template is 254, and deletion of sub-trees could cause any one of these internal nodes to become a leaf node. Such a leaf node is displayed on the left side of . (Like all other leaf nodes, any ‘intermediate level leaf node’, though permitted to bind with a concept node, is equally free to remain unbound on a given occasion. In this latter case, the leaf node may be regarded as simply absent, from a functional standpoint).
Now, since there is a total of 254 internal nodes in our general tree template, and since we have assumed 40 000 concept nodes, we must add (roughly) an additional 10 million CBNs to the picture (i.e. 254 × 40000). Adding this figure to the 10 million CBNs required for the deepest level leaf nodes (and doing the arithmetic precisely), we discover that a total of 20.4 million CBNs are required, in the worst case, for the binding of concepts to role nodes. An additional 254 CBNs are required (one for each internal node) to allow for the binding together of triplets, as discussed previously, and displayed in .
It appears then, that fewer than 21 million CBNs would suffice for the bindings required to represent even very large propositions. Given the 15 billion (or so) neurons within the cerebral cortex, the number cited does not appear problematic. Moreover, even if we suppose that the brain employs three additional copies of the general tree template, fewer than 84 million CBNs would be required. As noted earlier, the processing functions of CBNs might not be limited to enabling bindings. For all we know, CBNs involved in propositional representations could, on other occasions, be involved in very different sorts of tasks.
Singer's second criticism
As the reader will recall, Singer's second main concern was that ‘unresolved problems [for conjunctive binding] arise with the representation of nested relations among the components of composite objects, such as visual scenes or sentences’ (Singer Citation2003). However, by now it should be clear that, at least in the realm of sentence comprehension and propositional thought, conjunctive binding does not require a prohibitive number of CBNs. Moreover, it is a virtual platitude that tree structures of the kind we are considering can permit the representation of recursive structures within propositions. Also, because each of the 256 ‘ultimate-leaf’ nodes within the conjectured tree template could potentially bind with a substantial number of concepts (probably fewer than 40 000 but more than 50), the total number of representable propositions would be vast indeed.
Concerning the realm of sensory features, the applicability of tree structures is less clear. However, Singer's second critique of conjunctive binding does assume that some nesting of structures is required in the perception of complex situations. Such nesting strongly suggests (not to say, entails) that hierarchical structures are involved, in which case perceptual processes might well involve tree representations of feature combinations. Moroever, the perception of novel intra-modal and cross-modal feature combinations, as cited in Singer's first criticism, might also, for all we presently know, be explicable in terms of tree-based representations that employ CBNs in the manner outlined above. Of course, the precise details of how bindings occur in tree-based representations probably differ substantially from the method that I have outlined. However, given the possibility I have sketched, I submit that Singer's first two arguments do not clearly diminish the prospect that conjunctive coding is the sole method of binding employed in sentence comprehension and thought representation. Even within the realm of sense perception, the explanatory potential of CBNs may exceed Singer's expectations. However, regarding the latter, we must now consider Singer's third criticism of CBNs.
The third argument
Singer's third argument is succinctly summarized in his own words, to wit, ‘it is hard to see how novel objects and hence entirely new relations among features can be recognized and represented, as this would require rapid reconfiguration of input connections to previously uncommitted cells’. It is not entirely clear from his wording whether the ‘entirely new relations among features’ occur solely within the ‘novel objects’ referred to, or whether the ‘new relations’ might also pertain to relationships between two or more novel objects. Singer's use of ‘hence’ would seem to imply the former interpretation, but I shall consider both possibilities.
If the central problem is that of representing any novel object, and this is deemed problematic because a novel combination of known features (some of which are relational features) within an object will, by itself, create novel relationships, then a possible solution has been touched upon above. We have seen that Singer's allusion to nested structures in sense perception introduces the prospect that hierarchical relationships occur in the organization of perceived objects, and this in turn introduces the possibility that some form of ‘perceptual representational trees’ are involved. If we accept the involvement of such trees as a working hypothesis, then matters may resolve straightforwardly. As hypothesized above, each known sensory feature would have the capacity to enter into a binding (via a CBN) with some leaf node of the pre-existing tree structure. Given this, it should be feasible for a novel combination of sensory features to separately, but simultaneously bind with various leaf nodes of the tree. Each time this occurs, we obtain ‘for free’ a representation of novel relations among the sensory features, and the entire tree structure, with its collective bindings, becomes a representation for the novel object. However, although we are still considering the first interpretation of Singer's quoted remark, a further subtlety remains. For, if the ‘novel relations’ in question are presumed to arise not merely because of a novel combination of known features, but because some entirely new, atomic relational feature (such as ‘is-below’) is present, then it is far from clear that any binding method should be expected immediately to accommodate novel relations. It is commonly held, both within the psychology and philosophy of perception, that entirely novel features are not well perceived by humans. Recognition of a novel relational feature involves learning, and improves gradually after repeated exposure. This gradual learning process arguably entails synaptic weight adjustment, which presumably ‘reconfigures’ projections between neurons that immediately respond to the novel feature and other structures which represent the full perception of that feature.
If any reader doubts, however, that synaptic weight adjustment is required for the recognition of novel features (including relational features), there still remains the possibility of innate ‘pre-wiring’. It is certainly true that any set of neurons that immediately responds to a novel relational feature, upon first presentation, will have some pre-existing connections to other neural structures. Given this, it is not wild to suppose that each ‘receptor neuron’ in, say, the visual cortex possesses innate projections to many separate CBNs, where each CBN in turn is connected to some leaf node in a conjectured ‘representation tree’. This supposition at least represents a theoretical possibility that has not been excluded by Singer's arguments. Moreover, if the supposition is granted, then the inclusion of a novel relation into a full representation of a ‘novel object’ could proceed in the straightforward fashion outlined for the case where relational features are not novel.
To summarize, we have now considered the case, introduced by Singer's third criticism, where ‘novel relations’ are interpreted as having arisen within some single novel object. There remains the second interpretation of ‘novel relations’ where an entire situation is novel because separate objects in the situation are related in some fashion that is novel. Yet, this last case is analogous, in relevant respects, to the cases considered above. If we adopt the reasoning presented above, then each distinct object in a novel situation can itself be represented via some perceptual representation tree. The root of each such tree can have many separate CBN connections to various leaf nodes in some higher level tree (T) which is to represent the complete situation. Any ‘novel relation’ that holds between the separate objects in the complete situation will be comprised of feature nodes which can be assumed to possess CBN connections to leaf nodes in that same higher level tree, T. The dynamics of how binding nodes become effectively active are just as hitherto discussed. The considerations that apply to radically novel relations between separate objects are identical to those considered when the relationship existed between features of a single object.
A preliminary causal model for synchronous binding
In this section I present a hypothetical model for how two separate nodes, each of which might be realized by a distinct set of several neurons, could be caused to fire in synchrony. The model involves one winners-take-all (WTA) network, in which CBNs play a key role in the process that engenders synchronous firing. The scope of this model is restricted to instances of filler–role binding, but as we have seen, such binding might well occur at the leaf nodes of tree structures, thereby permitting a vast number of binding combinations to be tokened on differing occasions.
In what follows, it is assumed that a single ‘filler node’ (selected from a finite set of candidate fillers) should have the capacity to bind with a single ‘role node’ (likewise selected from a finite set of candidate roles) on a given occasion. It is also assumed that, within the filler–role domain of applicability, the most active filler node should bind with the most active role node, provided the two nodes are both ‘currently unbound’ (that is, they are both free to bind).Footnote† Examples of connectionist systems that employ this strategy (of binding an unbound filler node to an unbound role node whenever both are the most active nodes possessing appropriate connectivity) include the language learning and parsing system presented in Hadley and Cardei Citation(1999), and the language parsing system of Stevenson Citation(1994). In these systems, a series of bindings can be set sequentially, and can thereafter continue to coexist, subject to activation decay. Likewise, in the model described below, new bindings can become active even when other bindings remain in effect. In addition, all such bindings are subject to activation decay within CBNs. Thus, activation decay permits bindings to be broken in due course.
Hereafter, it is assumed that both role nodes and filler nodes are particular types of concept representations. For example, it seems reasonable to suppose that our understanding of the role of ‘being the agent of some action’ involves having the concept of ‘being an agent’. Also, filler nodes can straightforwardly correspond to concepts. For example, the concept of ‘dogs’ clearly plays the agent role in the proposition ‘dogs chase cats’. Having said this, further elaboration is in order. Among other things, please note that a ‘concept node’ is not being identified with a single neuron. Most, and possibly all, of our concepts involve the representation of multiple features (e.g. ‘frog’ has features corresponding to shape, colour, etc.). Such features, in turn, are widely believed to be representable only when specific sets of neurons become active. For this reason, I shall assume that the active representation of any concept node will likewise require the activation of a significant number of neurons. Thus, each concept node is implemented by a ‘neuron-set’, and nodes are abstractions of such sets.
Moreover, for all that is known to the contrary, it is plausible that within each such neuron-set, there exists a small subset (of perhaps 10 or fewer neurons) that form a reverbatory clique (or assembly) such that:
| a. | every neuron within the clique forms excitatory connections with all others in that clique; | ||||
| b. | every neuron, N, in the total ‘concept neuron-set’, is connected to several members of that clique in a way that permits N to be jointly excited by the activation levels of those clique members. | ||||
In addition, let us assume as a working hypothesis that for each set of neurons, whose joint activations reliably qualify as representing a given concept (whether a role or a filler), there does exist such a clique, containing about 10 neurons. Also, due to conditions (a) and (b) above, it follows that the activation of most (say, 70% or more) members of a concept node's clique will typically lead, via spreading activation, to the activation of all elements of the concept's total neuron-set. In light of this, the following shorthand is adopted: when I describe ‘two nodes as being connected (by a link)’, I am actually saying that within each of the two cliques associated with those abstract nodes, most (70% or more) of the neurons within one of the cliques make afferent (input) connections on most of the neurons in the other clique, and vice versa. Given the multiplicity of connections between the neurons in the respective cliques, and also within each clique, it will be possible for each of the two ‘nodes’ to spread its activation to the other node.
Having said this much, I would remind readers that the model described here is not intended as a biologically accurate account of how synchronous firing is caused. Rather, it is best viewed as a suggestive metaphor that illustrates one possible way in which CBNs might provide underpinnings for synchronous firing. The metaphor displays how such synchrony could be viewed as a symptom of a binding's having occurred, rather than as the mechanism of role–filler binding. Moreover, the model demonstrates that, in principle at least, a feasibly small number of CBNs could provide the required underpinnings.
Initially, for diagrammatic simplicity, the model is presented on a small scale. It will emerge, however, that even when the model is scaled up to accommodate, say, 2000 role nodes and 100 000 filler nodes, the total number of CBNs required would be 200 million, which is clearly within a feasible range (see section 3.3 for further detail on this issue).
displays, abstractly, the overall layout of the approach adopted here. Although the diagram does not display all connections and CBNs involved, the general conjecture is both simple and partially familiar. It is assumed that between each possible pair of filler and role nodes, there resides a unique CBN that is reciprocally linked to the two members of that role–filler pair. illustrates how a single role node is linked to four separate filler nodes via intervening CBNs. Note that if we have N filler nodes and M role nodes, then exactly (N×M) CBNs are needed. Omitted from is the assumption (elaborated presently) that all CBNs involved will reside in a single WTA network.
Figure 7. Role nodes are shown as white circles. Filler nodes are shaded circles. The black diamond-shaped nodes are CBNs. Only four of the 16 CBNs are displayed here.
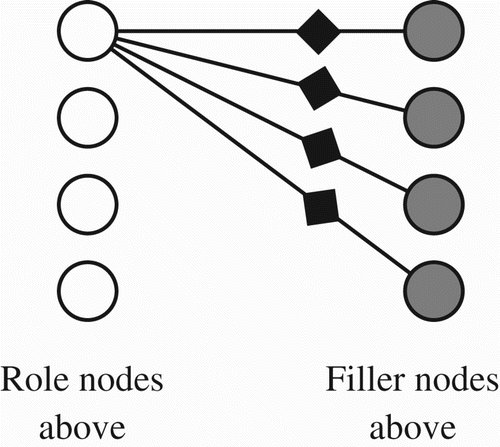
Another noteworthy feature is that, although nodes within the model's diagram are shown in close proximity and spatial regularity, in reality there would be no need for nodes to be in proximity, or to be separated by regular distances. That is, even a perfect biological analogue to the model could incorporate neurons that are widely distributed, although functionally they would need to preserve a connectivity pattern analogous to the one displayed.
Also worth noting is that there is some evidence that mechanisms that actually cause synchronous firing, in cases where the firing is not directly provoked by sensory input, can involve widely distributed cortical connections. As Singer Citation(1999) observed, there are ‘indications that [the causes] are mediated at least in part by cortico-cortical connections’ amongst cells that reside not only ‘in the same area but also cells distributed across different areas and even across the two hemispheres’.
Moreover, synchrony has been observed in the firing of neuron sets located as distantly as the two hemispheres. Given this, it is likely that mediation between the two neuron sets will involve intervening neurons. There is, then, prima facie plausibility, at least, to the notion that synchronous firing would be engendered by intervening CBNs. We now consider one way in which that might occur.
Architectural details of the model
As mentioned earlier, each concept node (whether role or filler) within the model is best viewed as an abstract entity whose implementation contains, as a proper subset, a neuronal clique. In contrast, I shall hereafter assume that each CBN is entirely realized by a clique of about 10 neurons.Footnote† Please bear in mind also that, internally, each clique contains mutually excitatory neurons. Readers may have observed that cliques as conceived here bear a striking resemblance to certain of Hebb's cell assemblies (Hebb Citation1949). Owing to the high degree of intra-connectedness within cliques, and for reasons explained shortly, we may reasonably suppose that all neurons within any clique will rapidly attain effectively synchronous firing rates. Consequently, the firing rate of any abstract node will be identified here with the average firing rate within the corresponding clique. (Prior remarks about connectivity between cliques also apply here.)
Concerning the inducement of synchronous firing, each positive weighted connective link that resides between any pair of neurons (either within a clique, between cliques, or between those neurons that comprise a given ‘concept set’) is assumed to possess a tunable delay factor. As demonstrated in Vibert et al. Citation(1994) and in Sougné Citation2001), such delay factors can be modulated to induce synchronization in the firing of connected neurons. The approach of Vibert et al. is designed to achieve an impressive degree of biological plausibility. Their model is mathematically complex, and the reader is referred to their paper for full details. While Vibert et al. do not present equations for tuning delay factors, they remark that such tuning is possible, and note that many biological factors affect the strength of such delay values. Sougné's (Citation2001) work does present equations for the tuning of delay factors and incorporates a ‘training phase’ for such tuning. I return to this issue in section 3.2.
Returning now to the abstract level depicted in , additional architectural requirements are as follows.
| i. | All CBNs are involved in a single WTA network. Thus (though not displayed in the figure), between each distinct pair of CBN nodes, there exists a reciprocal pair of inhibitory links. All such links are assumed to be equally weighted, with a fixed negative weight of −1. | ||||
| ii. | Following the approach illustrated in Hadley and Hayward Citation(1997) and Hadley and Cardei Citation(1999), each CBN is capable of inhibiting its own capacity to compete with other CBNs. Given this capacity, it is assumed that any CBN that is already actively bound to a role and filler node will refrain from all WTA competitions as long as that specific binding remains active. In Hadley and Hayward Citation(1997), such ‘refraining’ is achieved by modifier links that emanate from each CBN to its own competition links. | ||||
Processing details of the model
Let us now return to lower level implementation details, and consider the activity of postulated ‘neurons’ within cliques, and within concept sets. It has been briefly mentioned that all connective links within cliques and elsewhere possess tunable delay factors. Specifically, it is assumed that all positively valued connective links within the model possess delay values that undergo tuning during a ‘developmental training phase’ which occurs prior to any coherent binding processes. Following the approach of Sougné we may further assume that, during this early training phase, delay tuning occurs whenever it happens, by chance, that random pairs of neurons fire in synchrony for a useful duration of time. By ‘useful duration’ I mean that the synchrony extends long enough for meaningful delay tuning to occur on the connective link between the related pair of neurons. The precise adjustments to the delay value on the link might be made in accord with Sougnéapos;s equation (Sougné Citation2001 p. 110), but in biological reality, the tuning processes may well be more complex than his equation would suggest (as he readily acknowledges).
There is no apparent need, at present, to commit to the precise details of delay tuning processes. Nevertheless, it is reasonable to postulate that, as a consequence of the developmental training phase, the delay factors on all links within cliques, between cliques and within ‘concept sets’ have converged on values that ensure, within acceptable margins of tolerance, that synchronous firing of neurons within a given cluster will rapidly occur, once the majority of such neurons receive initial stimulation. For example, if a given CBN wins a competition with other competing CBNs, the neurons in the winning CBN clique attain high firing rates, and then rapidly converge to virtually synchronous firing.
If the kinds of cliques postulated here were realized in the human cerebral cortex, an additional factor would produce a strong tendency towards the synchronous firing of the neurons involved. For, in these cliques, each neuron receives stimulus from all the others (e.g. from nine other neurons). Any subset of neurons within the clique which happen to be firing in synchrony would be more likely to trigger an immediate firing in the remaining members than would any of the non-synchronously firing neurons present (this characteristic of synchrony is well established; see Singer Citation1999). Given this, coupled with the modulation of delay factors on the connections involved, we have yet another reason for postulating that, within the simulated model, synchronous firing would occur within cliques. Moreover, whenever two cliques are densely interconnected (as is true for a CBN and an attached role or filler clique), and neurons within one of the cliques are firing synchronously, there will be a strong tendency for this synchrony to be conveyed to the attached clique. (Recall that each neuron within a role or filler clique has reciprocal connections with most neurons in the attached CBN clique.)
Having now examined, in a general way, some of the factors that induce synchronous firing, we now consider how, on a particular occasion, the existence of CBNs could cause a pair of role and filler nodes to adopt a synchronous firing pattern. At the outset, we assume that, due either to processes involved in the comprehension of an externally presented sentence, or in the formation of some individual thought, a particular filler node has acquired a higher activation level (firing rate) than all other filler nodes, and similarly, a particular role node has attained a higher activation level than all other role nodes. The firing rates of these two highly active nodes are not assumed to be equal. I do assume, however, that these two most active nodes are currently unbound, that they should become ‘bound’ to each other, and that a sign of this binding is to be synchronous firing. (Recall that there exist connectionist parsing systems (e.g. Hadley and Cardei Citation1999) that illustrate how and why the filler and role nodes that should bind are the most active nodes of their respective types that are available to bind.)
Now, given that we have a highly active role node and filler node (and that other role and filler nodes may be active as well), the following sequence of events is conjectured to occur.
| 1. | (1)]All filler nodes (respectively, role nodes) convey their firing rate (activation level) to each CBN that they are attached to. In consequence, each CBN will receive activation from exactly one filler node and one role node. As a mathematical necessity, the CBN that now receives the greatest activation will be the one that is attached to the most active filler node (Fl) and the most active role node (R1). That particular CBN is currently unbound (not in an active reverbatory coalition) because the attached filler and role nodes were assumed to be unbound. | ||||
| 2. | The CBN just mentioned (call it ‘C1’), as a result of stimulus just received, attains a higher firing rate than all currently unbound CBNs. As a result of this stimulus surge, and the fact that a stimulus was also received by other unbound CBNs, a WTA competition is triggered among all such CBNs. (CBNs already in an active binding will not now compete.) | ||||
| 3. | That CBN, C1, which is connected to the currently most active filler and role nodes (F1 and R1) now wins this WTA competition. Having won the competition, the neurons within C1's clique remain highly active, but the cliques of the losing CBNs fall to activation levels that are well below their firing thresholds. (Thus, each losing CBN is temporarily unable to induce synchrony in its attendant role and filler satellites. Any losing CBN remains in this low activation state until later being restimulated via a strong stimulus from both its attendant role and filler nodes. If and when that occurs, a new WTA competition will ensue among the currently unbound CBNs.) | ||||
| 4. | Since the neurons in C1's (the winner's) clique have high firing rates, and because of the prior tuning of delay factors of all links within the clique, together with the full degree of interconnectedness of the neurons, the members of this clique rapidly assume nearly identical oscillation phases. | ||||
| 5. | Recall that each neuron in C1's clique is connected to most neurons in the attendant cliques of F1 and R1. Likewise, each neuron in the latter two cliques is connected to most neurons in the C1 clique. Thus, even while the neurons in C1's clique are converging towards synchronous firing, activation is also being spread to the F1 and R1 cliques. Because of the prior tuning of delay factors on all links between C1 and F1 (and, respectively, C1 and R1), there will be some pressure on F1 and R1 to achieve synchronous firing with C1. In any event, once synchronous firing is attained within C1, there will be strong pressure for F1's clique (respectively, R1) to enter into synchrony with C1's. This is not only because of the tuned delay factors on links between C1 and F1, but also because each neuron in F1 is receiving multiple (more than five) synchronized stimulus surges from those in C1. The situation is precisely analogous for R1's clique. | ||||
| 6. | Since both F1 and R1 attain close synchrony with C1, the firing frequency of neurons in all three cliques will be approximately identical and in a single, unified phase. Within actual brains, synchronous firing is a matter of degree, and perfect synchrony is rarely observed, if ever. | ||||
In passing, I would remind readers of a remark made early in section 3, namely that bindings created by CBN activity are subject to activation decay. Such decay will occur not only in any CBN that has engendered synchronous firing, but also in the attached pair of role and filler nodes. In this manner, any ‘binding’ that is created can later be broken.
Scaling up the provisional model
By this stage, it has clearly emerged that the number of CBNs required for the overall approach adopted here is on the order of the simple product of the number of roles and fillers involved. Within the ‘propositional tree structures’ described in section 2, role nodes appear both internally and at the leaf level. However, the number of internal nodes is always less the number of leaf nodes, and I have argued that, within the propositional realm, 256 leaf nodes would suffice. I have also argued that a reasonable upper bound on the number of (concept) fillers available would be roughly 40 000. However, let us be extravagant for a moment. Suppose for argument's sake that 1000 leaf (role) nodes are present, and that any of 100 000 filler (concept) nodes could potentially bind with each leaf node. Also assume that each of the 100 000 fillers could potentially bind with an internal node (because an internal node on one occasion could become a leaf on another occasion). Then the total number of CBNs required would not exceed 2 × 1000 × 100,000, which is 200 million. If each CBN is comprised of a clique of 10 neurons, then we require 2 billion neurons to ensure that all potential bindings are possible. Given an estimated 15 billion neurons within the human cerebral cortex, the figure of 2 billion does still seem feasible.
We should bear in mind, however, that this is, by far, an extreme upper bound estimate. For one thing, it is highly unlikely that even a majority of the 100 000 fillers would be appropriate for binding to each role. Moreover, consider the number of distinct propositions that would be representable if 2000 roles were each capable of binding with any one of 100 000 fillers on a given occasion. The total number of combinations (and hence, propositions) would be 100 0002000. This is an inconceivably large number that vastly exceeds the number of propositions a human could ever, in reality, comprehend. It likewise exceeds, by a staggering degree, the number of distinct percepts (sensory patterns) that anyone could ever perceive. Thus, even if the roles involved were sensory in nature, and the fillers perceptual, the figure of 2 billion neurons would seem to be a serious overestimate of the number needed for conjunctive binding purposes.
Summary and conclusions
In the foregoing, I have examined the comparative merits of conjunctive coding and synchronous firing as methods of binding. Particular emphasis has been placed upon the question of efficiency, in terms of the numbers of nodes required by each method. I have argued:
| i. | (i)]that we are not actually in a position to judge the number of nodes required by synchronous binding, because we do not yet know the numerical range of neurons involved in the underlying processes that engender this type of binding, | ||||
| ii. | that the genesis of synchronous firing in non-sensory domains may, for all we know, involve a substratum of CBNs, | ||||
| iii. | that the tri-pronged critique of conjunctive binding, presented in Singer (Citation1999, Singer (Citation2003), is far from decisive in non-sensory domains, where the organization of information may involve tree structures whose binding mechanisms require only a feasible number of CBNs. | ||||
Returning to point (iii), I have argued that even within the domain of sensory feature binding, Singer's case against conjunctive binding may be inconclusive; for, it remains possible that hierarchical organization of feature information is pervasive in sense perception, in which case tree structures, grounded upon CBNs, may provide the binding apparatus for all of the following:
-
(a) the perception of novel objects;
-
(b) the perception of novel relations within and among objects;
-
(c) the perception of nested structures within situations.
Turning to the genesis of synchronous firing and binding, I have, in section 3, presented a connectionist model, grounded upon conjunctive coding, which illustrates how synchronous bindings could be engendered in the realm of roles and fillers. As argued in section 3.3, the approach requires only a feasible number of CBNs. Moreover (contrary to my figures’ proportions), the model is compatible with a widely distributed configuration of nodes, separated by irregular distances. As previously acknowledged, the model is not proposed as biologically realistic. Rather, it is presented as a hypothetical analogue whose details may suggest the direction in which the true explanation of synchronous binding is to be found. In any case, the hypothetical model illustrates an important logical possibility, i.e. that binding via synchronous firing is merely a symptom of a deeper form of binding, one that intrinsically involves the operation of conjunctive binding nodes.
Acknowledgements
I am very grateful to Lokendra Shastri and to the journal's anonymous reviewers for several beneficial criticisms of a prior draft of this paper.
Additional information
Notes on contributors
R. F. Hadley
Email: [email protected]Notes
†Other proposed solutions to the binding problem include: CONPOSIT, by Barnden and Srinivas Citation(1991); Lange and Dyer Citation(1989); and CONSYDERR, by Sun Citation(1992). Each of these solutions possesses merit and features of interest. Whether they also possess biological foundations remains an open question.
†As the paper unfolds, readers may note certain abstract analogies between the approach adopted here and that espoused by van der Velde and de Kamps Citation(2006). As it happens, virtually all of this paper was written well before I had any knowledge of theirs, and my circuitry remains a great deal simpler than theirs. Indeed, van der Velde and de Kamps contend that their methods avoid the use of CBNs. In any case, my overall approach is an extension of my earlier work (Hadley and Hayward Citation1997), which pre-dated the solutions espoused by van der Velde and de Kamps.
†Note that if explicit labels were to be employed, they could conceivably be dynamically changed via a separate set of CNBs that connect to the internal nodes.
† A given node is considered to be the ‘most active’ filler node (respectively, role node) provided it has the highest firing rate among all filler (respectively, role) nodes connected to a given core network.
†Note, therefore, that our use of CBNs would not imply that some single neuron represents the concept of a ‘yellow volkswagon’. In any case, it would require more than the activation of a single CBN to represent a complex concept, for the role and filler being bound by the CBN would also need to be activated.
References
- Barnden , J. A. and Srinivas , K. 1991 . Encoding techniques for complex information structures in connectionist systems . Connection Sci , 3 : 263 – 309 .
- Cottrell , G. Connectionist parsing . Proceedings of the Seventh Annual Conference of the Cognitive Science Society . Mahwah, NJ. pp. 201 – 211 . Lawrence Erlbaum Associates .
- Elman , J. L. , Bates , E. A. , Johnson , M. H. , Karmiloff-Smith , A. , Parisi , D. and Plunkett , K. 1996 . Rethinking Innateness: A Connectionist Perspective on Development , Cambridge, MA : MIT Press .
- Feldman , J. A. and Ballard , D. H. 1982 . Connectionist models and their properties . Cognitive Sci , 6 : 205 – 254 .
- Fodor , J. A. 1975 . The Language of Thought , New York : Crowell .
- Fodor , J. A. and Pylyshyn , Z. W. 1988 . Connectionism and cognitive architecture: a critical analysis . Cognition , 28 : 3 – 71 .
- Fodor , J. A. and McLaughlin , B. P. 1990 . Connectionism and the problem of systematicity: why Smolensky's solution doesn't work . Cognition , 35 : 183 – 204 .
- Goulden , R. , Nation , P. and Read , J. 1990 . How large can a respective vocabulary be? . Appl. Linguist , 11 : 341 – 363 .
- Hadley , R. F. 2004 . On the proper treatment of semantic systematicity . Minds and Machines , 14 : 145 – 172 .
- Hadley , R. F. 2003 . “ Systematicity in connectionist generalization ” . In The Handbook of Brain Theory and Neural Networks , Edited by: Arbib , M. A. 1151 – 1156 . Cambridge, M.A : MIT Press .
- Hadley , R. F. and Cardei , V. C. 1999 . Language acquisition from sparse input with no error feedback . Neural Net , 12 : 217 – 235 .
- Hadley , R. F. and Hayward , M. B. 1997 . Strong semantic systematicity from Hebbian connectionist learning . Minds Machines , 7 : 1 – 37 .
- Haug , H. 1986 . History of neuromorphometry . J. Neurosci. Meth , 18 : 1 – 17 .
- Hebb , D. O. 1949 . The Organization of Behaviour , New York : Wiley .
- Hinton , G. E. , McClelland , J. L. and Rumelhart , D. E. 1986 . “ Distributed representations ” . In Parallel Distributed Processing; Explorations in the Microstructure of Cognition, Vol. I , Edited by: Rumelhart , D. E. , McClelland , J. L. and the PDP Research Group . Cambridge, MA : MIT Press . D.E. Rumelhart, J.L. McClelland and the PDP Research Group, Eds
- Hummel , J. E. and Holyoak , K. J. 1997 . Distributed representations of structure: a theory of analogical access and mapping . Psychol. Rev , 104 : 427 – 466 .
- Lange , T. and Dyer , M. 1989 . High-level inferencing in a connectionist network . Connection Sci , 1 : 181 – 217 .
- O'Grady , W. 1997 . Syntactic Development , Chicago : The University of Chicago Press .
- Rumelhart , D. E. , McClelland , J. L. and the PDP Research Group . 1986 . Parellel Distibuted Processing; Explorations in the Microstructure of Cognition, Vol. I , Cambridge, MA : MIT Press .
- Shastri , L. 1988 . A connectionist approach to knowledge representation and limited inference . Cognitive Sci , 12 : 331 – 392 .
- Shastri , L. and Ajjanagadde , A. 1993 . From simple associations to systematic reasoning: a connectionist encoding of rules, variables, and dynamic bindings using temporal synchrony . Behav. Brain Sci , 16 : 417 – 494 .
- Singer , W. 2003 . “ Synchronization, binding and expectancy ” . In The Handbook of Brain Theory and Neural Networks , Edited by: Arbib , M. A. 1136 – 1143 . Cambridge, MA : MIT Press .
- Singer , W. 1999 . Neuronal synchrony: a versatile code for the definition of relations? . Neuron , 24 : 49 – 65 .
- Smolensky , P. 1990 . Tensor product variable binding and the representation of symbolic structures in connectionist systems . Artif. Intell , 46 : 159 – 216 .
- Sougné , J. P. 2001 . Binding and multiple instantiation in a distributed network of spiking nodes . Connection Sci , 13 : 99 – 126 .
- Stevenson , S. A unified model of preference and recovery mechanisms in human parsing . Proceedings of the Sixteenth Annual Conference of the Cognitive Science Society . Mahwah, NJ. pp. 824 – 829 . Lawrence Erlbaum Associates .
- Sun , R. 1992 . On variable binding in connectionist networks . Connection Sci , 4 : 93 – 124 .
- Thorpe , S. J. and Imbert , M. 1989 . “ Biological constraints on connectionist modeling ” . In Connectionism in perspective , Edited by: Pfeiller , R. , Schreter , Z. , Fogelman-Souile , F. and Steels , L. Amsterdam : Elsevier .
- van der Velde , F. and de Kamps , M. 2006 . Neural blackboard architectures of combinatorial structures in cognition . Behav. Brain Sci ,
- Vibert , J. F. , Pakdaman , K. and Azmy , N. 1994 . Inter-neural delay modification synchronizes biologically plausible neural networks . Neural Net , 7 : 589 – 607 .