Abstract
Variable binding is a difficult problem for neural networks. Two new mechanisms for binding by synaptic change are presented, and in both, bindings are erased and can be reused. The first is based on the commonly used learning mechanism of permanent change of synaptic weight, and the second on synaptic change which decays. Both are biologically motivated models. Simulations of binding on a paired association task are shown with the first mechanism succeeding with a 97.5% F-Score, and the second performing perfectly. Further simulations show that binding by decaying synaptic change copes with cross talk, and can be used for compositional semantics. It can be inferred that binding by permanent change accounts for these, but it faces the stability plasticity dilemma. Two other existing binding mechanisms, synchrony and active links, are compatible with these new mechanisms. All four mechanisms are compared and integrated in a Cell Assembly theory.
1. Introduction
Symbol systems have been enormously successful and it has been proposed that, at least at some level, humans are symbol processors (Newell Citation1990). Whether humans are symbol processors or not, they can effectively use rules, and symbolic systems, such as ACT (Anderson and Lebiere Citation1998), have been very successful as models of human cognition. This success is probably due to the rule based or at least rule-like behaviour of humans in a wide range of tasks such as natural language processing.
Unfortunately, symbolic systems also have problems with brittleness (Smolensky Citation1987). The symbols are not grounded (Harnad Citation1990) and it is difficult or impossible to learn new symbols that are not just some combination of existing symbols (Frixione, Spinelli, and Gaglio Citation1989).
These and other problems provided motivation for the rise of connectionism, particularly in the 1980s. Connectionist systems are particularly good at learning, and thus may be able to learn new symbols. If the systems learn from an environment, the newly learned symbols might even have semantic content grounded in that environment.
However, early connectionist systems were criticised for their inability to perform symbolic processes (Lindsey Citation1988). In particular, they were criticised for their lack of compositional syntax and semantics (Fodor and Pylyshyn Citation1988).
Variable binding offers an answer to these criticisms. A good variable binding solution allows for the implementation of rules; connectionist primitives can be combined, and variables instantiated as constants. If this can be done so that the result has compositional syntax and semantics, the criticism will have been answered.
For a binding mechanism to be functional, it must be able to support a range of binding behaviours (Section 2.1). Binding by synchrony (Malsburg Citation1981) is a well explored mechanism that is functional, but it can only support a limited number of bindings. Similarly, binding by active links (van der Velde and de Kamps Citation2006) has also been explored and is functional. Both mechanisms are restricted to active bindings, that is, the bindings must be continuously supported by neural firing, and when that firing ceases so do the bindings. This may limit the effectiveness of a neural system, particularly as it relates to composition (Section 6.2).
After some background for reader orientation, binding by synaptic change is introduced. This comes in two forms, binding by short-term potentiation (STP) and binding by compensatory long-term potentiation (LTP). Simulations that indicate these mechanisms are functional are described, in particular showing bindings can be formed and erased, that bindings can overlap, that a large number of bindings can be supported simultaneously, and that they can provide compositional syntax and semantics. It is shown that the four binding mechanisms, two existing and the two novel synaptic change mechanisms, are not mutually exclusive, and one system could use all four mechanisms. Ramifications for memory formation speed and duration are also explored along with other issues in the discussion and conclusion.
2. Background
Humans behave as if they have compositional syntax and semantics, so if systems based solely on neural models are to duplicate human behaviour, they too must exhibit compositional syntax and semantics behaviour. One way for neural systems to exhibit compositional syntax and semantics is by variable binding.
A good cognitive model should have compositional syntax and semantics (Fodor and Pylyshyn Citation1988). Standard symbolic cognitive architectures have this compositionality, but it is more difficult for connectionist models to exhibit it.
Compositional semantics means that the semantics of a complex thing includes the semantics of that thing's constituents. So sentence 1.
Pat loves Jody.
Sentence 1
Includes the semantics of Pat, love, and Jody. Compositional syntax means that the syntactic structure of complex things affects the underlying semantics. For example, the semantics of sentence 1 is different from the semantics of sentence 2.
Jody loves Pat.
Sentence 2
So the semantics of a sentence must be more than the sum of its parts.
Variable binding can be used to solve these problems in a neural system by binding the semantics of constituents in a syntax sensitive way. Sentence 1 could be represented by a case frame (Fillmore Citation1968) for love where the actor slot is bound to Pat, and the object slot to Jody.
2.1. The variable binding problem
The variable binding problem is a key neural network problem that involves combining representations. It is also called the binding problem (Malsburg Citation1986), and the dynamic binding problem (Shastri and Aijanagadde Citation1993).
Perhaps the simplest variable binding problem is binding the features of an object. This is required when a new object is presented. If an object is composed of features, then when an object is presented, its features need to be bound together. One classic example is the red-square problem. If the system is presented with two objects, a red-square and a blue-circle, it can relatively easily activate the internal representation of all four of these features. The question is, how does the system know which pairs are bound.
A system can use a solution based on existing objects. For example, if there are two sets of 100 features that can be bound, the problem can be solved by having 10,000 stored bindings, but this number will grow exponentially with the number of features, and the number of potential combinations. This solution is just a form of auto-associative memory that is open to the problem of exponential growth and thus combinatorial explosion. However, the features being bound into an object do not need to be a variant of an existing object, but can be a combination that is novel for the system.
Another example of this problem is binding parts into a whole, such as binding elements of a square lattice into rows or columns (Usher and Donnelly Citation1998). A third variant of this problem is the what-where problem. If a system can recognise multiple objects simultaneously and their locations, how does it know where each thing is and which things are in each location. This is an example of the above problem; in this case, location is one of the features, so one variant is the left-square right-circle problem.
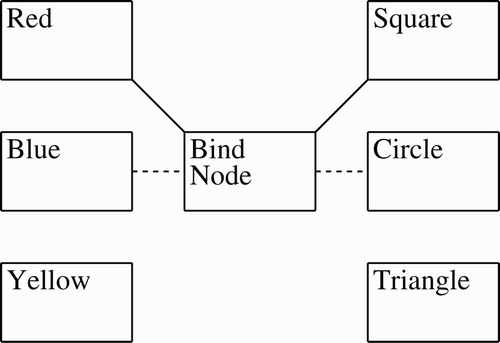
Furthermore, unlike the standard associative memory task, binding features of an object has the associated difficulty of erasing the binding. After some time, red and square are no longer bound, and both may be bound to some other concept, for example red-triangle. This reuse problem is also a question of binding duration. As long as the binding persists, it can be used, but once it stops working, it can be reused for a new binding (Section 2.2). This paper is mainly concerned with bindings that are formed and then later erased so they can be reused. is an example of this. Here each box refers to a group of neurons with the outer six boxes referring to concepts (e.g. Red and Circle) and the centre box acting as a binding node. An initial binding of Red and Square, represented by the solid lines, is later replaced by the binding of Blue and Circle, represented by the dashed lines.
Figure 1. Idealised binding with bind node: initial binding of red and square is later replaced by blue binding with circle.
Another standard problematic example is filling in frames (Shastri and Aijanagadde Citation1993; Henderson Citation1994; Jackendoff Citation2002; van der Velde and de Kamps Citation2006). An example of this would be a verb frame (Fillmore Citation1968). For example the verb move might take an actor, object and location. In the sentence Pat moved the ball to the door. Pat would be the actor, the ball the object, and to the door the location. When processing a sentence, the system would have to fill in the details by binding these objects to these slots. Perhaps frames are a common task for systems that use variable binding due to the compositional syntax and semantics problems mentioned by early critics of connectionist systems (Fodor and Pylyshyn Citation1988). Frames are a flexible knowledge representation format (Schank and Abelson Citation1977); they are a relational structure where data is used to fill in structures with variables. The basic frames are templates that need to be instantiated, and reused. Erasing the original's filler is one mechanism that can enable reuse. Moreover, if properly implemented, frames give compositional semantics.
Rules are another important case where variable binding is needed. Firstly, rule based systems are Turing complete (Hopcroft and Ullman Citation1979), so a neural implementation of rules would be Turing complete. This is not particular surprising as others have shown other connectionist systems to be Turing complete (Siegelmann and Sontag Citation1991). Secondly, rules are widely used as a means of modelling human cognition (Laird, Newell, and Rosenbloom Citation1987; Anderson and Lebiere Citation1998), so rules are important for cognitive modelling. An example rule would be if X gave Y to Z, then Z possesses Y. Finally, sequences are important and can be implemented by rules and by connectionist systems. For example, one system uses dynamic connections to learn sequences (Feldman Citation1982). These learned sequences are then automatically forgotten by a process of connection weight decay.
Unification is a more complex form of variable binding. This is done by symbolic systems such as language processing systems (Shieber Citation1986) and logic programming. There are a range of unification approaches, and complex structures such as directed acyclic graphs may be combined (unified). It is a complex form of pattern matching. This can lead to a case where a structure may be illegally combined with a subset of itself, known as the occurs check (Browne and Sun Citation1999). Unification in neural systems may incorporate soft constraints making the system more flexible (Hofstadter Citation1979; Kaplan, Weaver, and French Citation1990). For instance, there may be a grammar rule that combines a noun phrase and a verb phrase and requires that they agree in number; a soft constraint may allow the same rule to apply, in some circumstances, when they do not agree in number, and this rule could be used to recognise ungrammatical sentences.
A problem that is closely related to variable binding is Hetero-associative memory, which refers to the association of an input with an output. This is roughly what Smolensky (Smolensky Citation1990) refers to as variable binding, which differs from the term as used in this paper because hetero-associative memories are permanent or extremely long-lasting. Perhaps this difference is the basis of the term dynamic binding. To avoid confusion, in this paper, variable binding will only refer to the case where a binding can be erased and reused.
Hetero-associative memory is a common and well understood form of memory (Willshaw, Buneman, and Longuet-Higgins Citation1969). Here items are combined, and each is linked to that combination. Presentation of one enables the system to retrieve the combined representation. Of course restrictions can be placed on the inputs, and several features may be needed to activate the full set of items (Furber, Bainbridge, Cumpstey, and Temple Citation2004). Standard neural models can account for this problem using standard Hebbian learning rules to implement a form of LTP (Gerstner and van Hemmen Citation1992) for permanent synaptic change. However, this work is not easily extended to associative memories like semantic nets (Quillian Citation1967). The problem here is that one memory needs to be associated with another, yet the two must remain separate. One excellent graph theoretic approach to this problem deals with biological constraints on connectivity and activation (Valiant Citation2005). Both hetero-associative memory and associative memory, though related, are different from variable binding (but see Section 6.3).
2.2. Properties of binding mechanisms
Different binding mechanisms have different properties. This paper proposes that three properties are particularly important. These properties are:
(1) Persistence of binding. | |||||
(2) Number of bindings supported. | |||||
(3) Speed to bind. | |||||
Others have discussed the number of bindings property (e.g. Shastri Citation2006; van der Velde and de Kamps Citation2006), but persistence of binding and speed to bind are not typically discussed. This may be due to other work on binding being almost exclusively based on bindings being supported by neural firing (Section 6.2).
Persistence of binding has already been mentioned. Hetero-associative memories (Section 2.1), as typically modelled, persists forever. At the other extreme, binding via synchrony only persists as long as at least one of the bound items is firing, and binding by active links persists as long as the binding node is firing. This leaves a wide range of times that a binding might persist.
The number of bindings supported refers to how many entirely independent bindings, or distinct entities, can be supported simultaneously. One mechanism might be based on reusable binding nodes. Each node might be used to support one binding, and there are as many bindings as nodes. has one binding node that can support any of the nine possible bindings of one colour and one shape. A second, or third, node could be added to support another. The solution of forming a dedicated binding node for each possible binding is impractical because it would require an exponential number of nodes, so the nodes must be reusable. Therefore, in the case of verb frames (Fillmore Citation1968), each slot of each verb might be a binding node. The slot fillers could be simple nouns, or they could consist of other verbs, in for example the case of sentential complements, to allow an arbitrary degree of complexity. Of course complex noun phrases would also need binding slots. With active links (van der Velde and de Kamps Citation2006) each binding node is represented by a circuit, and these can be combined to form verb frames. Binding via synchrony does not use nodes but has a limited number of bindings that a system can store (Sections 2.4.1 and 6.1).
Finally, time to bind is an important consideration. How long must items be coactive before they can be bound? Binding via synchrony is very fast and can occur within tens of ms (Wennekers and Palm Citation2000). The binding via LTP mechanism proposed below (Sections 3.1 and 4.2) takes much longer.
2.3. Cell assemblies and learning
A cell assembly (CA) is the neural basis of a symbol (Hebb Citation1949). A CA is a subset of neurons that have high mutual synaptic strength enabling neurons in the CA to fire persistently after external stimulation ceases. In the simulations discussed in this paper, a small subset of all the neurons represents a symbol. If many of the neurons in the CA are firing, the symbol is active.
CAs give a sound answer to the neural representation of two types of memory, long-term memory and short-term (or working) memory. The firing of many neurons in a CA is the neural implementation of short-term memory; this high frequency and persistent firing makes the CA active.
The red-square problem can be restated in terms of CAs. There is a CA each for red, blue, square, and circle. When a red-square and a blue-circle are presented, all four base CAs are active. is an example of this problem. In this example, each cell represents a neuron with circles representing neurons that fire in a given period. The relevant rows are labelled with all neurons in a row representing the appropriate feature, and CAs are represented by orthogonal sets of neurons. In this case some, but not all, of the relevant neurons are firing. Somehow the pairs must be bound, so that the system can ascertain, for example, the colour of the square, and this binding should only persist for a relatively small amount of time.
Figure 2. Sample neural firing pattern for red-square and blue-circle.

A CA is formed by a process of synaptic modification, and typically, this synaptic modification is modelled as a form of LTP and long-term depression (LTD). CAs are long-term memories with Hebbian learning rules providing the link between long- and short-term memories (Hebb Citation1949; O'Neill, Senior, Allen, Huxter, and Csicsvari Citation2008). When neurons co-fire, they become more likely to fire together because their mutual synapses are strengthened (Hebb Citation1949), and eventually, this can lead to the formation of a CA. Hebbian learning is local; it occurs between two neurons that are connected and takes information based solely on these neurons. Typically the synaptic weight is increased when both the pre-synaptic and post-synaptic neurons fire. For all but the simplest forms of Hebbian learning, there is an associated form of forgetting that is, somewhat oddly, called anti-Hebbian learning. Here, if one neuron fires and the other does not, the synaptic weight is decreased (White, Levy, and Steward Citation1988), preventing the weight from growing without limit. There is significant biological evidence for Hebbian learning (Miyashita Citation1988; Brunel Citation1996; Messinger, Squire, Zola, and Albright Citation2005). Moreover, as this learning is based on pairs of neurons, biological experiments are relatively simple, so there is good reason to believe that some sort of Hebbian learning does occur in brains.
None the less, the precise mechanisms that are used by biological systems are not entirely clear. There are a range of Hebbian learning algorithms that follow the above definition, but differ from each other; none account for all biological data, and the biological data is far from complete.
The simplest rule merely increases the synaptic weight when both neurons co-fire. There is no anti-Hebbian rule, and the weight may be clipped at some value (Sompolinsky Citation1987) to prevent it growing without limit.
Timing is also important to learning. The Hebbian rule involves the firing of neurons at the same time. In a model that uses continuous time, the same time requires some degree of flexibility. Work on Spike Timing Dependent Plasticity (Gerstner and Kistler Citation2002) adds another dimension to the complexity of Hebbian rules. In these rules, precise timing dynamics are important with the order of neural firing affecting whether the change in synaptic weight is positive or negative.
The interaction between learning and firing leads to a complex dual dynamics (Hebb Citation1949). Once a CA is learned, it is hard to forget because any activation of it strengthens its intra-CA connections; this is a form of the stability plasticity dilemma (Carpenter and Grossberg Citation1988; Fusi, Drew, and Abbott Citation2005). Similarly, it is difficult to do anything with a CA until it has formed.
Hebbian learning rules are the most widely accepted model of the mechanism used by the brain to form CAs, the neural basis of concepts. Binding is not necessarily related to Hebbian learning, but if CAs, once formed, can be appropriately bound, then the resulting system can have compositional semantics and syntax. It then remains to ask what mechanisms can be used to bind CAs together?
2.4. Solutions to the problem
The mechanism that is most commonly used in neural simulations of variable binding is synchrony (Malsburg Citation1981). A lesser used mechanism is active links (van der Velde and de Kamps Citation2006), and both require neural firing to maintain the binding.
2.4.1. Binding via synchrony
Binding via synchrony requires neurons that are bound together to fire together. Therefore, if two neurons are bound, they might fire at times X, X+0.2, X+0.5, X+0.8, and X+1. For example, the neurons might fire at 0.1, 0.3, 0.6, 0.9, and 1.1; and then repeat the pattern at 1.5, 1.7, 2.0, 2.3, and 2.5. Of course there is some room for variation, and the binding usually applies to a much larger number of neurons than two.
A good example of this is SHRUTI, a non-neural connectionist mechanism (Shastri and Aijanagadde Citation1993). In this model, different sets of concept nodes are bound together by firing at roughly the same time. Rules can be instantiated in the nodes, and these can continue to propagate the bindings to new items. SHRUTI has been used to develop, among other things, a syntactic parser (Henderson Citation1994). Here synchrony is used to bind slots and fillers. Unfortunately, the system only allows 10 bindings, so only relatively simple sentences can be processed.
There is significant evidence for synchronous firing in biological neural systems (Eckhorn et al. Citation1988; Abeles, Bergman, Margalit, and Vaddia Citation1993; Bevan and Wilson Citation1999). Some really convincing evidence that synchronous firing is used for biological binding is provided by a study that shows how binding is facilitated by a stimulus that is presented synchronously (Usher and Donnelly Citation1998).
There are several simulated neural models of binding via synchrony (e.g. Bienenstock and Malsburg Citation1987; Wennekers and Palm Citation2000). Networks of spiking neurons are used to segment a visual scene into different objects based on the firing timing of neurons associated with those objects (Knoblauch and Palm Citation2001); a scene with a triangle and a square is presented, and neurons associated with the square fire together and the triangle neurons fire together, but at different times from the square neurons. Spiking neurons are also used to parse simple text (Knoblauch, Markert, and Palm Citation2004) using binding via synchrony.
One major problem with binding via synchrony is the number of bindings that it supports (Section 2.2). The connectionist SHRUTI parser (Henderson Citation1994) is limited to 10 bindings, and Shastri and Ajjanagadde suggest that this limit is about 10 (Shastri and Aijanagadde Citation1993). All bound items must fire in roughly the same pattern, but to handle variations within neural behaviour, this pattern must be somewhat flexible. Similarly, items that are bound differently must fire in a different pattern. For example, the neurons in red and square must fire in roughly the same pattern, while the neurons in blue must fire in a pattern that is different from red. As these firing patterns must occur in relatively brief time scales (∼33 ms), and they must be relatively flexible, there are only a restricted number of bindings that can be maintained simultaneously. It is not entirely clear how many bindings biological neural systems allow, but as more bindings exist, there is an increased likelihood that closely related patterns will coalesce thus incorrectly combining sets of bound items.
2.4.2. Binding via active links
A more recent approach to the binding problem creates active neural circuits to support the binding (van der Velde and de Kamps Citation2006). Both primitives and binding nodes are represented by neural circuits, similar to CAs. The binding is selected by active primitives and is maintained by neural firing in the binding node. Like binding by synchrony, the binding stops once firing stops in the binding node and stopping the binding circuit erases the binding. Binding can persist beyond firing in the primitives.
Effective simulations of natural language processing and vision have been demonstrated. This is a promising mechanism for variable binding. The active neural circuit solution is similar to an older connectionist solution called dynamic connections (Feldman Citation1982). Dynamic connections are used to store bindings that are activated by a pair of inputs, and then persist for a considerable period. The persistence automatically decays allowing the node to be reused later.
2.4.3. Binding via LTP
Another option is to bind by changing synaptic weights. An earlier version of the work presented in this paper used a fatiguing leaky integrate and fire (fLIF) neural model to implement rules to count from one number to another (Huyck and Belavkin Citation2006). A Hebbian learning rule is used to change synaptic weights permanently as a form of LTP.
Sougne provides an interesting blend between binding by changing synaptic weights and binding by synchrony (Sougne Citation2001). The changing synapses regulate synchrony by modifying delays on connections.
Unfortunately, a general binding solution based on LTP faces the stability plasticity dilemma (Carpenter and Grossberg Citation1988). The dilemma is how is it possible to add new knowledge without disrupting existing knowledge in a neural net (Lindsey Citation1988). With binding, base CAs would need to be stable, bindings would need to be plastic, and new CAs would still need to be formed. Thus any system that allowed a LTP based binding to be erased could have the problem of erasing the base CAs that are being bound.
2.4.4. Other connectionist binding mechanisms
One standard mechanism is to create a new binding element for each possible binding. As mentioned earlier (Section 2.1), this has the problem of combinatorial explosion. This combinatorial explosion might be addressed by use of hierarchically allocated binding nodes (Hadley Citation2007) using prespecified roles. For natural language parsing, this requires millions of nodes, but the brain has billions of neurons, so this is plausible.
Another connectionist mechanisms for binding is merely to combine the bound representations, but this leads to systems that have problems with compositional syntax. An example is Tensor Product binding (Smolensky Citation1990) which forms a type of cross product of the variables that are being bound.
While some work has been done on binding via synaptic change in neural systems, most neural binding work has been done using synchronous firing. Some non-neural connectionist work is relevant to the problem. However, the possibility of binding via synaptic change is an under-explored area.
3. Binding via LTP and STP
There is strong evidence that distinct features that co-occur in a particular object cause synchronous neural firing (Eckhorn et al. Citation1988; Abeles et al. Citation1993; Usher and Donnelly Citation1998). While this appears to be solid evidence for binding via synchrony, it is not conclusive proof. Synchronous firing may simply be an emergent property of the neural representation of the new object as it is an emergent property of standard long-term CAs (Wennekers and Palm Citation2000). Assuming there is binding by synchrony, it still has a problem with capacity and a problem with duration of binding.
It is not entirely clear how many bindings can be maintained by a network at any given time, but each binding must have its own unique pattern of synchrony (Section 2.4.1). Natural language processing may require many bindings as do other tasks such as object recognition. Since CAs cross brain areas (Pulvermuller Citation1999), orthogonalising domains (e.g. vision and language) is not a viable solution; that is, the brain can not be partitioned into areas where bindings are distinct so that binding frequencies can simultaneously support multiple distinct bindings.
Also, the synchronous binding only persists as long as the CAs are active. Once they stop, the binding is lost. While it is not entirely clear how long memories persist, there is a wide range of times over which a binding might persist.
Even if binding via synchrony occurs in the brain, this does not mean that there are not other types of binding. A different mechanism for binding, as is shown below, is change in synaptic weights. There are at least two variants of known biological synaptic weight change, LTP and STP.
3.1. Binding via LTP
One possible solution to the binding problem is permanent synaptic change; biologically this is LTP and LTD. Objects are bound using synaptic weight change, and these weight changes remain until future learning erases them.
For LTP to be able to solve the variable binding problem, the binding must be able to be erased. The mechanism then faces the stability plasticity dilemma (Carpenter and Grossberg Citation1988). If the same mechanism is used to form the initial memories and to do the binding, something else must prevent the initial memories from being erased when the bindings are erased.
3.2. Binding via STP
Most simulation work that involves learning relies on LTP. However there is another type of learning, STP, and there is extensive evidence that STP occurs in biological neural systems (Buonomano Citation1999; Hempel, Hartman, Wang, Turrigiano, and Nelson Citation2000). It is still a type of Hebbian learning, based on the firing behaviour of the neurons a synapse connects, so that co-firing increases the synaptic weight. However, unlike LTP, the change is not permanent.
Some have proposed that STP provides support for LTP (Kaplan, Sontag, and Chown Citation1991). That is, in the initial stage of CA formation, short-term connection strength adds activation to the nascent CA that supports the co-firing that provides impetus for LTP. More recently, short-term connection strength has been proposed as another basis of working memory (Fusi Citation2008; Mongillo, Barak, and Tsodyks Citation2008). This contradicts the basic idea of active CAs as the basis of working memory, but the two proposals may be compatible.
Another use for STP is for binding. In this case, the base memories are bound using STP. As the STP is automatically erased, so is the associated binding. This paper is the first to describe the use of STP in simulations of binding.
Note that the four binding mechanisms, synchrony, active links, compensatory LTP and STP, are not mutually exclusive. Section 5.4 shows synchronous firing behaviour alongside binding via LTP and STP, and describes how all four mechanisms could be combined in a single system.
4. Simulating binding with LTP and STP
To show that the STP and compensatory LTP binding mechanisms function, simulations of a simple paired association task, similar to the red-square problem (Section 2.1), are described. These and all the simulations described in this paper, use the same basic fLIF neural model.
4.1. fLIF model
The neural model that is used for the simulations described in this paper is an extension of the standard leaky integrate and fire (LIF) model which is in turn an extension of the integrate and fire (IF) model. A similar model (Chacron, Pakdaman, and Longtin Citation2003) has been shown to account for inter-spike intervals under various input conditions better than the standard LIF model. The IF model, commonly called the McCulloch Pitts neuron (McCulloch and Pitts Citation1943), has a long standing history and is quite simple. Roughly, neurons are connected by uni-directional synapses. A neuron integrates activity from the synapses connected to it, and if the activity surpasses a threshold, the neuron fires sending activity to the neurons it connects to. Connections may be excitatory or inhibitory; excitatory connections adding activity from the post-synaptic neuron and inhibitory connections subtract activity. LIF models are more biologically faithful than simple IF models (Churchland and Sejnowski Citation1992). In the IF model, if a neuron does not fire, it loses all its activity. In the LIF model, a neuron retains a portion of that activity making it easier to fire later. Typically, the neuron loses all its activity when it fires (Maass and Bishop Citation2001). All of these models are less complex and less accurate than Hodgkin Huxley models (Hodgkin and Huxley Citation1952) and other compartmental models (Dayan and Abbott Citation2005) which are extremely faithful to biology, breaking each neuron into several compartments and modelling interactions on a fine time grain (<1 ms).
The simulator runs in discrete steps with every neuron being modified in each step, and activity being collected in the next. The network of neurons can be broken into a series of subnets. Each neuron has two variables associated with it, and an array of synapses, and each subnet has four constants associated with all its neurons.
The two variables associated with each neuron i are fatigue F i and activation A i . As neurons fire, activation is passed to neuron i and is accumulated in A i .
The first constant is the firing threshold, θ. A neuron i fires if
If a neuron does not fire, some of its activation leaks away. This leak, or decay, is the second constant D where D>1. Ignoring external input and assuming i did not fire at t−1, activation of neuron i at time t is
When neuron i fires, it sends activation (or inhibition) along its synapses to other neurons according to the strength of each synapse, so neuron j receives activation according to synaptic strength w
ij
. The neuron is an integrator, so it accumulates activity from the synapses connected to it. Therefore, given P
j
, the prior activation of neuron j, either 0 or EquationEquation (2), the activation at time t+1 is
These equations describe an LIF model (Maass and Bishop Citation2001). The fatigue variable is incremented by the third constant F c in a cycle when the neuron fires, and is decremented by the fourth constant F r in a cycle when the neuron does not fire. This makes it more difficult for neurons to fire the longer they are firing. Fatigue is a property of biological neurons (Kaplan et al. Citation1991).
The model has a loose link with time in biological neurons. The model does not incorporate conductance delays or refractory periods, and these behaviours all happen in under 10 ms, so each given cycle can be considered to be roughly 10 ms. Consequently, each neuron emits at most one spike per 10 ms. of simulated time, and the timing precision is at most 10 ms. This is a shortcoming of the model, but enables efficient simulation of hundreds of thousands of neurons on a standard PC.
The model also has some degree of topological faithfulness. The Hopfield Net (Hopfield Citation1982) has been a popular system for modelling brain function (Amit Citation1989), but it requires neurons to be well connected and connections to be bi-directional. Neither constraint is biologically accurate. However, one key point that these and other attractor nets (e.g. Rumelhart and McClelland Citation1982; Ackley, Hinton, and Sejnowski Citation1985) show is that attractor states are important; an attractor state is where roughly the same neurons and only those neurons fire in each cycle. This is a key point of CAs (Section 2.3).
The system uses neurons that are either inhibitory or excitatory but not both. While there is some debate over the biological behaviour, this follows the strict constraint of Dale's Law (Eccles Citation1986). In the simulations described in this section, the ratio is 4 excitatory to 1 inhibitory neuron as is claimed in the mammalian cortex (Braitenberg Citation1989).
The connectivity of the network, and subnets is also important. Like the mammalian brain, excitatory neurons are likely to connect to neurons that are nearby. The network is broken into a series of rectangular subnets. As distance is relevant, the topology of each subnet is toroidal (the top is adjacent to the bottom, and sides are adjacent to each other, like folding a piece of paper into a donut) to avoid edge problems. In the simulations described in this section, excitatory neurons also have one long distance axon with several synapses. So a neuron connects to nearby neurons and to neurons in one other area of the subnet. These connections are assigned randomly, so each new subnet is extremely unlikely to have the same topology as another subnet with the same number of neurons. EquationEquation (4) is used for connectivity.
4.2. Simulating binding by compensatory LTP
The first set of simulations being reported in this paper involve binding via permanent changes of synaptic strength. This involves a compensatory Hebbian learning mechanism (Huyck Citation2004) that makes permanent changes to increase a synapse's strength, akin to LTP, and permanent changes that decrease the strength, akin to LTD. The simulation also makes use of spontaneous neural activation, a known biological phenomenon (Amit and Brunel Citation1997), to support erasing bindings.
The gross topology is shown in . There are three subnets called the letter subnet, the number subnet, and the binding subnet. The letter and number subnets are trained to contain 10 CAs each. Both nets consist of 1600 neurons and the binding subnet has 400. The binding subnet has spontaneous neural firing (see below) to enable erasing. As the base subnets do not have spontaneous firing, their CAs, once learned, are much more stable.
Figure 3. Topology of intra-subnet connections in the compensatory LTP binding simulation: each neuron in the base subnets connect to the bind subnet, and each neuron in the bind subnet connects to the base subnets.
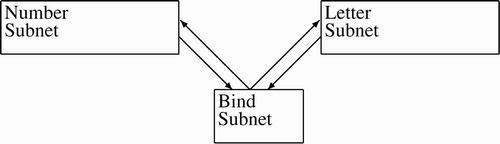
In addition to the intra-subnet connection, each bind neuron has 15 connections to both the other subnets. The neurons of the base subnets, letter and number, have 16 connections to the bind subnet and all inter-subnetwork connections are randomly assigned. The initial weights are initialised to a number close to 0.
The compensatory learning mechanism is another type of Hebbian learning. It forces the total synaptic strength leaving a neuron towards the desired weight, W B . Elsewhere (Huyck Citation2007), this learning mechanism has been used to learn hierarchical categories where categories share neurons. Compensatory learning is biologically plausible because the overall activation a neuron can emit is limited. Since a neuron is a biological cell, it has limited resources, and synaptic strength may well be one such resource.
The compensatory rule modifies the correlatory learning rules to include a goal total synaptic weight W
B
. EquationEquation (5) is the compensatory increase rule and EquationEquation (6)
is the compensatory decreasing rule; that is, EquationEquation (5)
is a Hebbian rule and EquationEquation (6)
an anti-Hebbian rule. W
B
is a constant which represents the desired total synaptic strength of the pre-synaptic neuron, and W
i
is the current total synaptic strength. R is the learning rate, which is 0.1. P is a constant and must be greater than 1. The larger it is, the less variance the total synaptic weight has from W
B
. P, W
B
, and R are constants associated with a particular subnet. When the two neurons co-fire there is an increase in synaptic weight corresponding to EquationEquation (5)
. If the pre-synaptic neuron fires and the post-synaptic neuron does not fire, the weight is decreased according to EquationEquation (6)
.
A summary of the value of the constants used in the first simulation can be found in . These values were determined by exploration of the parameter space via simulation. The parameter space, including topology, is practically infinite. This particular location is almost certainly not optimal, but does show solid results. An understanding of the dynamics of CA activation and formation is essential to select these parameters; this includes knowledge of various tradeoffs between parameters such as reducing firing threshold is similar to increasing synaptic strength. To a lesser extent, biological constraints also help in directing the search. For instance, excitatory synaptic weight is in the range of 0–1, and it is known that several neurons are needed to cause another to fire (Abeles Citation1991) so the threshold θ is much greater than that. In one study of anaesthetised guinea pigs, simulated models accounted for spiking behaviour when decay was roughly D=1.25 (Lansky, Sanda, and He Citation2006).
Table 1. Network constants.
During the entire run, there is spontaneous activation in the binding net. Spontaneous neural firing is a property of biological neurons (Abeles et al. Citation1993; Amit and Brunel Citation1997; Bevan and Wilson Citation1999), and it has been proposed as a mechanism for weakening and even erasing memories (Huyck and Bowles Citation2004).
In this simulation, some neurons may be spontaneously activated. This is modelled by the selection of a random number 0≤r<1 for each neuron in each cycle. If the r<0.03 the neuron is spontaneously active. Therefore, roughly 3% of neurons in the bind subnet fire spontaneously each cycle.
The simulation first learns the base number and letter CAs, then one of each is randomly selected to be bound. This is a simple paired association task similar to the task performed in earlier connectionist simulations (Feldman Citation1982) and those done in psychological experiments (e.g. Sakai and Miyashita Citation1991). Once bound, the binding is tested, followed by a test for an unbound letter and number. The binding is then erased by spontaneous activation; and the tests are rerun. For measurement, this binding, testing, erasing, and retesting process is repeated 10 times on each of 10 different networks.
The base CAs are learned by merely presenting components of them. As both the base nets consist of 1600 neurons, they can be divided into 10 orthogonal CAs of 160 neurons each. Fifty randomly selected neurons of a particular CA are selected and presented for 10 cycles. This is akin to clamping, but these neurons are given units of activation. After fatigue has accumulated they may not fire. After the 10 cycles of activation, the network is allowed to run for 40 more cycles. It is then reset with all activation and fatigue zeroed. Then a new CA is presented. Each set of 50 cycles of activation, run-on, and short-term variable resetting is called an epoch.
Each base CA is presented in a rotation so that all CAs are presented once every 1000 cycles. The complete training phase is 20,000 cycles so that each base CA is presented 20 times. Note that spontaneous activation in the bind net continues throughout this time.
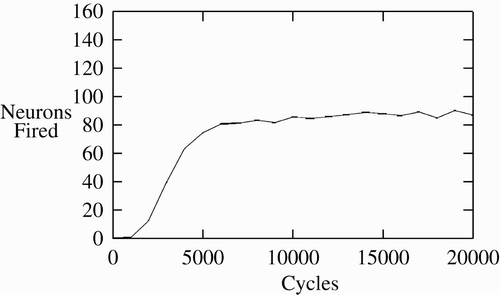
shows the CA formation process. A network is created with synaptic weights near 0. It is then trained, and at the 45th cycle of each training epoch, the number of neurons in the presented CA is measured. This is averaged over the presentation of each of the 20 base CAs, and over 10 networks. The number of neurons outside the desired CA firing was also measured, but was always zero. This shows a rapid increase in persistence, neurons firing towards the end of each training epoch, followed by a gradual increase after the 5000th cycle. Note, the maximum number of neurons that could be firing is 160, but fewer are firing due to fatigue. By cycle 20,000, the base CAs are quite persistent.
Figure 4. Neurons firing per cycle indicating CA formation.
After the training phase, the epoch duration is lengthened to 1000 cycles for the binding phase. A randomly selected letter CA and a randomly selected number CA are presented simultaneously. In a system that accepted visual input, both items would be presented simultaneously as in a paired association task. In this simulation, 50 neurons from both CAs are selected at random and presented for 10 cycles. As the CAs are already formed, these almost always persist for the duration of the binding epoch.
As ever, the bind subnet is spontaneously activated during this phase. Throughout this period the synaptic weights between the subnets gradually increase. When binding is successful, neurons in the bind subnet fire due to input from the active number and letter CA. This in turn causes the inter-subnet synapses to increase. In essence, a new CA is being formed and it includes neurons from all three subnetworks.
It is crucial that two CAs in the base subnets are simultaneously active. This is similar to the mechanism used for node activation by dynamic connections (Feldman Citation1982). Along with the spontaneously active bind neurons, these base neurons provide sufficient activation to fire some of the neurons in the bind subnet. Firing these base neurons causes the mutual synaptic strength between them and the base neurons to increase leading to further neural firing in the bind subnet. By the end of the binding epoch, a CA has been formed that includes the binding neurons, and this composite CA can be reactivated at any time over a significant period of time.
In the second epoch, the bound number is presented, and in the third, the bound letter is presented. When successful, this leads to activation of the binding CA and the opposite base CA. This further reinforces the inter-subnet synaptic strengths, improving the binding.
In the fourth epoch a randomly selected unbound number is presented, and an unbound letter is presented in the fifth. The correct result here is that no neurons in the opposite subnetwork fire.
The synaptic strength from the binding subnet that supports the binding is being reduced during the test unbound phase, but four further epochs of no base presentation are run to allow the binding to be sufficiently erased. The synapses from the binding subnet that support the binding move rapidly towards zero due to the application of compensatory learning rules (EquationEquations (5) and Equation(6)
) caused by spontaneous firing.
Synapses from the bind subnet to the base subnets are erased during the period of no presentation. During this period, neurons in the bind subnet fire, but no neurons in the base subnets fire. Consequently, the weights are reduced towards 0.
However, the synapses to the bind subnet from the base subnets are not changed during the testing of unbound items or during the period of no presentation. Instead, these synapses are reduced by the compensatory learning mechanism during the next two test epochs (epochs seven and eight).
The synaptic weights from neurons in the base subnets to the bind subnet do not change between the last binding test, and the first bind retest. Why then does the presentation of the here to fore bound item not cause the bind subnet to activate as it had done during the presentation in the second and third epochs?
Firstly, there are fewer neurons firing in the just bound item. This is due to the loss of intra-subnet synaptic strength during the binding. Secondly, there is little initial feedback from the bind node since its neurons no longer have much synaptic weight to the recently bound item. During this initial phase, the synaptic weights in the just bound item are changing. The weights to the bind node are being reduced while the weights within the just bound item are increasing. There is only a small part of the parameter space where this difficult task can be solved (Section 4.4).
Finally, there are four tests to assure that the binding has been erased. The formerly bound number and letter CAs are presented, followed by the formerly tested unbound number and letter.
For each network, this series of tests was run 10 times. It was run on a total of 10 networks. When the testing epoch length was 1000 cycles, 192/200 or, 96%, of the binding tests were successful, and 595/600, or 99.2%, tests of unbound CAs were successful. These measurements can be combined using a standard F-2 . The F-score is 97.5%.
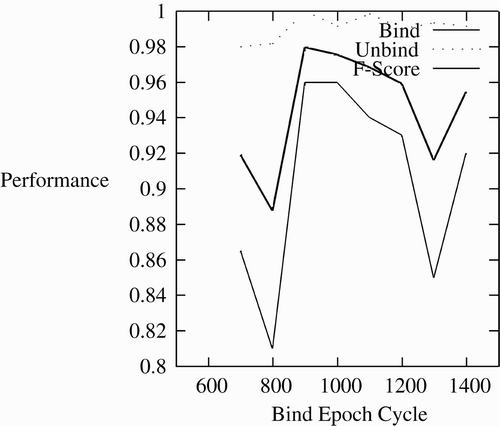
The length of the binding period is important. Substantial variations from the binding period of 1000 cycles causes decreased performance. shows this. Performance is best around 1000 cycles, and trails off when it is shorter or longer.
Figure 5. Effect of binding period on binding.
It is important that the base CAs must be formed and solid before binding occurs. They need to be solid so that they can fully participate in the binding process. This solidity is supported by a low firing threshold (θ=4) and a low decay rate (D=1.5); together these enable rapid recruiting of new neurons in a few presentations and high activity in the formed CA.
During base training, the binding area should not form a CA. That is, no single CA should be able to recruit many neurons from the binding area. Instead, two base CAs are needed to recruit neurons in the binding area. Consequently, little activity should be retained in the binding area (D=5), and it should be difficult to fire a neuron in the binding area (θ=7). As the binding area needs to be quickly recruited when two CAs are active, the total synaptic strength is high (W B =28) so that the connections to the other areas and within can be quickly formed. This differentiation between systems (binding vs. bound) may be supported neurally by different neural types or neural pathways. It is a hallmark of neural processing, that different neurons behave differently.
None the less, a similar system could probably be developed with all subnets having similar or even identical parameters. The difference in threshold could be removed with a corresponding change in synaptic weights. The total synaptic strength would see a corresponding reduction, though it would still be different between subnets. This could probably be compensated by changing the number of neurons. Changing the decay rate would be more difficult because the bind subnet can not retain much activity. A plausible solution would be to include an inhibitory system for the subnet that would inhibit neurons in the bind subnet on each cycle and thus eliminate the effect of a small amount of activity over many cycles. This has not been implemented, but it is likely that such a system could be developed.
This simulation fits into a rather small part of the parameter space. This is largely due to the rather precise way that the synapses from the base CAs to the binding subnet are erased. There is no spontaneous activation in the base subnets so the connections remain the same during the erase epochs. However, during the binding epoch, the synapses between neurons in the base CAs being bound have their strength taken by the synapses to the binding net. The loss of the feedback from the binding net after the erase epochs is enough to prevent the activation of the binding net when the bound base CA ignites.
Since this is so precise, minor changes to parameters cause a rapid decrease in performance. Changing the number of synapses from each base neuron to the bind neurons from 16 to 15 gives Bound/Unbound/F-Score results of 87%/95.5%/91.1%, and changing the number from 16 to 17 gives B/U/F results of 78.5%/94.8%/85.9%. Similarly, changing the base nets’ desired total synaptic strength (W B ) from 21 to 20 gives B/U/F results of 45%/99.2%/61.9%, and changing it from 21 to 22 gives results of 80.5%/89.8%/84.9%. Changing parameters individually is a form of gradient descent search; while gradient descent is not the best way to find an optimal place in the space, it can help to find local minima.
This is a particularly difficult binding simulation because there is no spontaneous activation in the base nets to facilitate erasing the binding. However, the lack of this spontaneous activation allows those CAs to persist indefinitely. Additionally, binding still works quite effectively.
4.3. Simulating binding by STP
Another option to implement variable binding by synaptic modification is to change the basic mechanism of synaptic change. LTP and LTD require the synaptic weight to remain unchanged until there is another application of one of the rules. Since synaptic change is caused by neural firing, the synaptic weights will remain unchanged until the appropriate neurons fire.
Another option is to have the weights automatically revert to zero over time. A rule that did this would be akin to STP. Note that the rule is still Hebbian in nature, changing the synaptic weight based solely on the firing behaviour of the two neurons that a synapse connects, but in this case, the weight also changes towards 0 when there is no firing.
The binding via STP simulations reported below are identical to the binding via compensatory LTP simulations (Section 4.2) except the bind subnet is removed, neurons are replaced by neurons that learn via both LTP and STP, and the binding epochs are 50 cycles. The bind subnet was provided to localise erasing of bindings; with STP the bindings are automatically erased at the neural level.
For STP, the simulation uses a new type of model neuron, termed a fast-bind neuron. The basic properties remain the same (Section 4.1), but some of the synapses leaving these neurons change their weights based on a different mechanism that accounts for STP.
The learning rule for fast-bind synapses that was used in these simulations is the simplest type of Hebbian learning. For each fast-bind synapse, if the pre-synaptic neuron fires in the same cycle as the post-synaptic neuron, the strength increases by the learning constant, which is 0.1. The weight is clipped at 1.
The rule for reducing synaptic weight is equally simple. If the neuron does not fire in a cycle, all fast-bind synapses leaving it have their weight decreased by a constant k (in this case k=0.004 which was selected to assure the binding persisted for roughly 250 cycles after last use). Therefore, a maximally weighted synapse, will return to 0 after 250 cycles of inactivity. Similarly, a minimally weighted synapse will go to 1 after 10 cycles of pre and post-synaptic co-firing.
The topology of the number and letter subnets is the same as in the LTP simulations, with 80% excitatory and 20% inhibitory, and inhibitory neurons have no fast-bind synapses. Each neuron has two fast-bind synapses to neurons in each CA in the opposite subnet, and those neurons are randomly selected.
The constants of the letter and number nets are the same as those in the LTP experiment; these are shown in . The training length is the same, 20,000 cycles, and the procedure is the same. The testing patterns are the same: binding epoch, two bind test epochs, two unbound test epochs, four epochs with no presentation, then two more tests of the formerly bound CAs, and two tests of the unbound CAs.
When the epoch lengths are 50 cycles, the system performs perfectly over 10 bindings on each of 10 nets. That is, all 100 bindings were successful, and all associated 100 erasings were successful. The Bound/Unbound/F-Score results are 100%/100%/100%. The bindings only need 10 cycles to be fully established, and as they are given 50, they are firmly established. Similarly, only 250 cycles are needed for the bindings to be fully erased. As there are two unbound test epochs, and four non-presentation epochs after the binding, there are 300 cycles of erasing, so erasing is also perfect.
4.4. Performance of LTP vs. STP
It has been significantly simpler to use binding by STP than to use binding by compensatory LTP. The portion of the parameter space that has been explored, where binding via compensatory LTP functions acceptably, is quite small. This has required the use of relatively precise topologies, precise training and use regimes, and spontaneous activation has been used only in the Bind subnet to support erasing. On the other hand, binding by STP works in a much larger range of conditions, and no exploration was done as the parameters for the LTP experiment were used. The manipulation of learning and forgetting weights allows for a corresponding manipulation of bind and unbind times (Section 5.2). Consequently, the next section discusses simulations using binding by STP to account for crosstalk and compositionality.
Compensatory LTP should be able to account for these phenomena, but complex training regimes may be needed, so at this juncture it seems unwise to describe further LTP simulations. The basic problem with binding by compensatory LTP along with erasing by spontaneous activation is that it faces the stability plasticity dilemma. Some memories are stable, the items being bound, and some are not, the bindings. It is difficult for the same mechanism to account for both. Formation of bindings is slow and they persist for a long time, just like CAs, so it may be better to view binding by compensatory LTP as a form of associative memory. However, this provides a new way of addressing the stability plasticity dilemma that is more fully discussed in Section 6.3.
The above simulations use fLIF neurons, but binding by compensatory LTP and STP should both be applicable to other neural systems. Spiking models are particularly appropriate (e.g. Maass and Bishop Citation2001). The rules may force breaking of the constraints of some attractor nets (e.g. Hopfield Net connections would no longer be bidirectional), but this is not incompatible from a simulation perspective (Amit Citation1989). Continuous value output neural models (e.g. Rumelhart and McClelland Citation1982) should also be compatible with binding via STP. It is not entirely clear how spontaneous activation would be implemented in these models, but compensatory learning should still work. It is also not clear how these mechanisms would apply to connectionist systems that do not have a close relationship to biological neurons like multi-layer perceptrons (Rumelhart and McClelland Citation1986).
The binding by compensatory LTP and binding by STP models that are presented in this paper are examples of classes of learning algorithms. The compensatory LTP mechanism was chosen because a compensatory mechanism eases recruitment of new neurons to a CA, binding, and supports erasing. The STP mechanism was chosen because of its simplicity. Ultimately, it is hoped that the neurobiological basis of neural learning will be sufficiently illuminated to say which algorithms are used for memory formation and variable binding in the biological system. Until then, an exploration of different binding algorithms and their use in large systems to simulate complex behaviour may be a good way to explore alternative neural binding mechanisms.
5. Further evaluation of binding by STP
In the binding by compensatory LTP simulation (Section 4.2), a binding node was used. In the STP simulation (Section 4.3) no explicit binding node was used, but implicitly, each CA was a binding node so that 20 bindings could be supported. This required that each CA was connected to each CA in the opposite subnet, and this would require a geometric growth in synapses as the number of base CAs grew linearly. The use of binding nodes can make growth of synapses grow linearly as the base CAs grow linearly with each base CA connecting to the binding node. Of course, it is also possible to have many binding nodes to support multiple bindings at a given time. How do multiple bindings interact and how many can be supported?
5.1. Crosstalk
In this section, a system that stores multiple bindings is described. Storing these bindings could lead to problems of cross talk, but none are seen. The simulation combines both STP and LTP on a single neuron with specific synapses devoted to each. The gross topology is similar to that of , but in this experiment there are multiple binding nodes.
There are four CAs in the letter subnet, four in number and four in bind. The letter and number CAs consist of 160 neurons each and the bind CAs have 100. All excitatory neurons have synapses leaving them that are modified by the compensatory LTP rule and synapses that are modified by the STP rule. The intra-subnet connections are the same as in the experiment described in Section 4.2 and all of these are modified by compensatory LTP.
Each neuron also has connections outside of the subnet and these are governed by the STP rule. Each neuron in the letter and number subnets has two connections to a randomly selected neuron in each CA in the bind subnet, and each of the bind neurons had three connections to each CA in the other subnets. This means that each neuron received roughly the same number of fast bind inputs as those in Section 4.3.
As in Sections 4.2 and 4.3, the base CAs were trained for 20 epochs of 50 cycles each. This formed stable CAs, and there was no spontaneous activation. The constants were the same as those for the base subnets in (θ=4, D=1.5, F c =1.0, F r =2.0,W B =21, and P=1.3).
Bindings were set by a single epoch of 50 cycles of presentation of one letter, one bind, and one number CA. Initially this was A0, B1, C2, and D3 each with a unique binding node.
Testing followed immediately with the numbers being presented in order. At the end of 50 cycles, the net was reset and the next number presented. On 100 nets, 400 of 400 correct bind and letter CAs fired in cycle 49 and no other neurons in those subnets fired. As expected, a random one to one binding (e.g. A1, B2, C3, D0 each with a unique binding node) faired as well.
This test means that bindings are set and then allowed to be maintained without activation for 150 cycles. With automatic synaptic reduction set at 0.004 (k=0.004) for each cycle when the pre-synaptic neuron does not fire, the synaptic weights return to zero after 250 cycles of inactivity. The simulation is run with a 50 cycle rest after the last binding, for a total of 200 cycles between the last cycle of each binding and each test. On 100 nets, none of the letter CAs have neurons firing, though 21 of the 400 bind nodes have some firing. The simulation was run with a 100 cycle rest after the bindings are set, and indeed the weights have returned to 0 and no firing was found in the bind and letter subnets.
The bindings are not formed simultaneously. So simultaneous presentation of red-square and blue-circle to the visual channel could not readily form two separate bindings. An attentional mechanism might be used with one object being attended to first and bound, followed by the second. Alternately, a different mechanism, e.g. active links, could be used to solve this problem (Section 5.4).
One common problem with binding is the presentation of two overlapping bindings, e.g. a red-triangle, and a red-square. This has been called the problem of two (Jackendoff Citation2002). This has been solved by a separate binding node for each pair (van der Velde and de Kamps Citation2006); elsewhere, this binding has been modelled with a computer simulation of CAs (deVries Citation2004) to account for psychological evidence.
The simulation was modified so that A0, B1, C0, and D1 were presented, each with a unique binding node. When the letter was presented the correct number CA was highly active with no incorrect neurons firing for each of the 400 presentations on 100 tests. This shows that the binding by STP addresses the problem of two.
Another test was done by presenting the number. When 0 was presented either A, C, or both could ignite; and B, D, or both could ignite for 1. On 100 runs, when 2 or 3 were presented, no letter neuron fired. Of the 200 positive tests, both of the bound letter CAs had over 100 neurons fire 158 times, between 10 and 100 fired in one and the other was over 100 21 times, and in 21 tests fewer than 10 neurons fired in one while the other was near peak. This means that usually both of the bound CAs ignited, but occasionally, due to competition, only one did.
As described in Section 4.1, each subnet is set up as a competitive subnetwork, with inhibitory neurons that connect randomly within the subnet. In this case, each inhibitory neuron had 60 synapses. Fewer synapses lead to less competition, and more synapses to more competition. With 30 synapses on one hundred runs, both letter CAs fired on each of the 200 tests, though on two tests less than 100 neurons fired in one CA. With 90 synapses on 100 runs on all 200 tests only one was active and the other had less than ten neurons firing. Note that an inappropriate neuron was never seen firing. Therefore, with ambiguous bindings, behaviour is dependent on the extent of competition.
5.2. Capacity
In some sense, an exploration of the number of bindings that can be simultaneously supported by STP is unnecessary. It is obvious that different orthogonal bindings can be independently supported. For instance, filling in the topology for with values from the simulations of Section 5.1 means that each orthogonal binding set can be represented by six base CAs of 160 neurons, and one binding CA of 100 neurons, or 1060 neurons. Therefore, the brain has a capacity for billions of these orthogonal bindings, though it is extremely doubtful that the brain has anything like that many orthogonal bindings.
Note that orthogonalising for synchrony is not the same as orthogonalising for STP binding nodes. With STP, CAs can be involved with multiple bindings simultaneously without being active, and there is no constraint on how many orthogonal bindings it can be in and be active. With synchrony, if a CA is in multiple distinct bindings it has to fire in synchrony with all of them.
None the less it is interesting to see how many potentially overlapping bindings, as in the experiments in Section 5.1, can be held simultaneously. Using the same method as in Section 5.1, one binding can be set at a time, and parameters can be varied to expand from the four bindings supported there. For simulations with extra CAs, an equal number of letter, number, and bind CAs are added. shows a range of behaviour of simulations. The labels in the figure refer to binding weight reduction k and bind durations with the 0.004/50 referring to the first simulations of Section 5.1 that support four bindings. The other lines refer to different settings of k and bind durations that allow more bindings to be supported.
Figure 6. Duration of bindings via STP varying by reduction rate and time to bind.
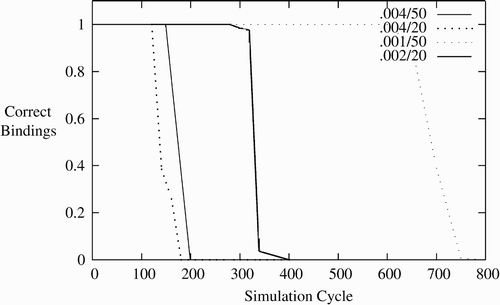
First, the synaptic weight reduction parameter k can be reduced from 0.004. As it is reduced, bindings will last longer and thus more can be set. In the 0.004/50 line of the maximum theoretical duration of an inactive binding is 250 cycles as all of the synaptic weights will have returned to 0. In the simulations, there are two synapses per neuron per CA, so several neurons need to be active to cause firing and the bindings will not last for the full 250 cycles. More synapses would cause this binding to persist longer, but could still not persist beyond 250 cycles. The 0.001/50 line represents a synaptic weight reduction parameter of k=0.001. This extends the maximum duration to 1000 cycles, though again this may not be reached. Practically, this performs entirely effectively to 650 cycles, and with 50 cycles to bind, this supports 13 bindings.
Similarly, reducing bind time increases the bindings that can be maintained. With a learning weight of 0.1, 10 cycles are the minimum to fully bind. The 0.004/20 line in the figure represents a bind epoch of 20 cycles. This has the same maximum duration of 250 cycles, but more bindings can be supported over this time. There is theoretical limit of 10 bindings, but 7 are maintained perfectly.
Reduced bind time and smaller synaptic weight reduction combine multiplicatively. The 0.002/20 line in theoretically supports 20 bindings, four times two for the synaptic weight reduction parameter times 50/20 for the bind time. Practically it is supporting 15 perfectly effectively. A further set of simulations was run with 0.001/20 (not shown in figure). This shows 30 bindings being supported perfectly.
There is evidence that STP can last over 30 s (Varela, Sen, Gibson, Abbott, and Nelson Citation1997), which is 3000 cycles in the model. With 10 presentations to bind, 300 overlapping bindings can theoretically be supported simultaneously following the above binding setting mechanism.
5.3. Compositionality
The binding by STP mechanism supports frames, and thus supports compositional semantics. A simulation based on four subnetworks described in binds successfully over 98% of the time.
Figure 7. Gross topology of the simulation of binding with frames. The rule subnet inhibits the slots of the frames that are not active, and the slots are bound to the appropriate verbs and nouns via STP.
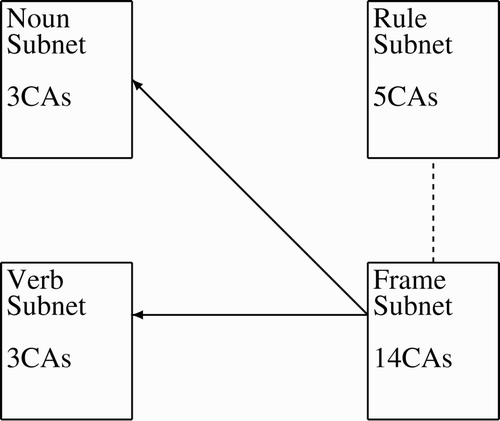
The four subnets are the Verb, Noun, Rule, and Frame subnets. The Verb and Noun subnet consist of three CAs each of 160 neurons each representing a word; the Rule subnet of five CAs each of 800 neurons each representing a rule; and the Frame net consists of 14 CAs each of 100 neurons which represent two frames each of seven slots. The constants were again the same as those for the base subnets in (θ=4, D=1.5, F c =1.0, F r =2.0,W B =21, and P=1.3).
As in the earlier simulations, connectivity within each subnet was distance biased with 80–20 excitatory to inhibitory neurons. In the Frame subnet this was extended with synapses that learn via the STP rule. Each frame consisted of seven slots, so the simulation has two frames. The base slot was connected to the frame's other slots, and, as in the simulations from Section 5.1, each of the neurons had two fast bind synapses to each of the appropriate CAs, along with the existing synapses. The sentential complement slot had fast-bind synapses within the Frame subnet (see below).
Connectivity between the subnets was from the Frame subnet to the Verb and Noun subnets, represented by the arrows in ; and from the Rule subnet to the Frame subnet, represented by the dashed line. Each frame consisted of seven slots: base, base verb, actor, object, location, instrument, and sentential complement. The actor, object, and location slots had connections to each of the nouns, and the base verb slot had connections to each of the verbs. Each of the excitatory neurons had two synapses to each of the appropriate CAs, and these synapses are modified by the STP rule. The sentential complement slot was also connected to the base slot of the other frame in the same fashion as the other slots were connected to nouns and verbs. The instrument slot was not used in this simulation.
The rules inhibited the frame slots that were incompatible. Each inhibitory neuron had 15 connections to each of those slots, and the rule CAs had 800 neurons to provide sufficient inhibition to prevent those slots from igniting even when bound.
As in the earlier simulations the net was trained by 20 presentations of 50 cycles for each of the base CAs. As the rule CAs were 800 neurons, 400 neurons were presented during training instead of 50 for CAs in the other nets.
The relevant binding parameters are 20 cycles and k=0.004. Binding was done by frames that correlated to the sentences Jody loves Pat., Pat loves Jody., Pat went to the store, and Jody said Pat went to the store. This was done by presenting the appropriate rule, slot and filler. For example, the verb love, the first frame's base slot, the first frame's base verb slot, and the start VP rule were presented for 20 cycles. For the first three sentences this was three presentations; for Jody loves Pat:
(1a) the first frame's base and base verb, verb love, and the start VP rule; | |||||
(1b) the first frame's base and actor slot, noun Jody, and add actor rule; | |||||
(1c) the first frame's base and object slot, noun Pat, and add object rule. | |||||
The second sentence inverted the actor and object; the third used the verb go and replaced the object rule and slot with location, and used the noun store. For the fourth sentence there were seven presentations:
(4a) the first frame's base and base verb, verb said, and the start VP rule; | |||||
(4b) the first frame's base and actor slot, noun Jody, and add actor rule; | |||||
(4c) the first frame's base and scomp, and add scomp rule; | |||||
(4d) the first frame's scomp, second frame's base, and add scomp rule; | |||||
(4e) the second frame's base and base verb, verb went and the start VP rule; | |||||
(4f) the second frame's base and actor slot, noun Pat, and add actor rule; | |||||
(4g) the second frame's base and location slot, noun store, and add location rule. | |||||
The complete test was done in three phases. The first phase bound the slots for Jody loves Pat. into the first frame. There was then a period of erasing of 250 cycles. The second phase bound the slots for Pat loves Jody. into the first frame and the slots for Pat went to the store. into the second frame. There was then another period of erasing followed by the slots for Jody said Pat went to the store. being bound into the first and second frame.
Testing followed the phases before erasing. Testing was done by presenting the base frame slot and the rule. The simulation was run on 10 different nets and each net did all three phases 10 times. The correct binding was considered to have occurred if more than 10 neurons in the correct node were firing in the 19th cycle after presentation; no incorrect binding was considered to have occurred if no other neuron in the appropriate net fired. For any given run, there were 14 possible correct bindings, and 13 possible incorrect bindings (the sentential complement could not have gotten the wrong base frame as both should be active). All of the correct bindings were formed and in 1290 of the 1300 runs no incorrect bindings occurred. This gives a Bound/Unbound/ F-Score result of 100%/99.2%/99.6%. Note that the failures that occurred all occurred within one net towards the end of the run, and were based on the base frame slot CAs recruiting each other via LTP.
A sentence is represented by a verb frame that has slots that are dynamically filled. Pat loves Jody. includes the semantics of Pat, love and Jody, and is different from Jody loves Pat. The simulation shows the difference between these two sentences and shows that frames can be implemented by STP. Similarly, the simulation of the semantic representation of Pat went to the store. shows that extra slots can be added seamlessly, and that multiple sentences can be stored simultaneously.
The phenomena is recursive. The simulation of the sentence Jody said Pat went to the store. shows that verb frames can be slot fillers. There is no theoretical limitation to the depth from a psycholinguistic standpoint. From a simulation standpoint, reactivation of bindings might be necessary during parsing to support the bindings, but Section 5.2 shows how 300 bindings might be stored without recourse to separation.
CAs are associative structures but frames are relational. This difference is bridged, above, by fast-bind connections. Initially, the frame is represented by the base slot, and the remaining slots are inactive. As slots are filled, the base slot and the particular slots are coactive; STP causes them to be bound so the base slot will activate the bound slots, but not the unbound slots. In the test, the unbound slots are explicitly activated via external activation.
A more sophisticated framing mechanism has been used in a natural language parser (Huyck Citation2009). This parser uses frames for both Noun and Verb phrases because both can have others as components. Rules no longer suppress frames, but instead activate particular slots in combination with existing activation. The parser is stackless and follows other psycholinguistic models (Lewis and Vasishth Citation2005).
When multiple rules are applicable because of simultaneous activation, competition via inhibition selects the rule to apply. For instance, when parsing a simple sentence like I saw. two items are active the NP I and the VP saw. Two rules are also applicable the AddActor rule and the AddObject rule. The VP is more active since it has been more recently activated, so the AddActor rule wins and is applied. Once a slot is bound, it is marked as bound (neurally) and cannot be rebound. In more complex sentences, several frames can be simultaneously active. In Pat said go to the store yesterday. The frames VP1 said actor-Pat scomp VP2, VP2 go loc-to-store, PP1 to-store, and NP3 yesterday are all simultaneously active; the NP1 Pat frame is inactive since it can no longer be modified. The rule that adds yesterday as the time of VP1 will activate the appropriate slot in that frame and the binding will be complete; the other two frames VP2 and PP1 are already bound. The rule causes the binding, but the binding persists after the rule ceases firing.
It is fair to note that during parsing of a sentence, multiple constituents may be simultaneously active. Only the appropriate items must be used to fill the appropriate slots. Binding by STP has now been used in two parsers: a stack based parser (Huyck and Fan Citation2007), and a memory based parser (Huyck Citation2009). In the stack based parser, the appropriate items are selected by activating them off of the stack, while other items on the stack are dormant.
In the memory based parser all active items are active, but binding sites are activated via rules. The item being bound has particular neurons that are associated with it being bound, and these are only activated by the rule. The slot that is being filled has connections to the neurons for all possible fillers with synapses that learn via STP. As only one slot and one filler are activated by a particular rule, only they are bound. Therefore, if a particular PP is being set as the instrument of a particular verb, the PP's neurons for being bound are active while no other filler has those associated neurons active; the verb's instrument slot is active and only that slot is active. The binding is completed, and the PP has a feature (represented by neurons) set that shows it has been bound. It may still remain active, but will no longer be used as a filler.
5.4. Combining binding mechanisms
Variable binding is a complex problem and is needed for a wide range of behaviour. Consequently, a system that could use a range of binding mechanisms would be more flexible than one that was limited to one mechanism. Fortunately, all four mechanisms, binding by compensatory LTP, binding by STP, binding by active links, and binding by synchrony are compatible.
The above binding by STP and by compensatory LTP experiments exhibit synchronous firing behaviour. For example, shows the firing behaviour of neurons in one run of the binding by compensatory LTP simulation described in Section 4.2. This shows a section of one binding epoch. The x axis shows the number of neurons firing in a subnetwork, and the y axis shows the cycle. Initially, the number and letter CAs are firing in different cycles. As the strength of the binding node grows, its neurons fire more frequently, and all three subnets begin to fire in synchrony; the firing is so closely correlated that the dotted number line disappears in the figure as it is covered by the letter line. The number of neurons firing in the base CAs oscillates, while the number firing in the binding CA oscillates while growing. This shows a strongly correlated firing pattern between the CAs.
Figure 8. Firing of neurons showing synchronisation while binding.
shows that items bound by STP fire synchronously. Here one letter is bound to one number as in the simulations in Section 5.1. The number is presented which leads to the activation of the bind CA and then of the letter CA. The firing patterns quickly synchronise.
Figure 9. Firing of neurons showing synchronisation of items bound by STP.
The above fLIF neural model has been used to implement several systems including the Cell Assembly roBot version 1 (CABot1) agent (Huyck Citation2008). CABot1 is an agent in a video game that assists the user and is implemented entirely in fLIF neurons. It consists of vision subnets, planning subnets, an action subnet, a control subnet, and parsing subnets (Huyck and Fan Citation2007). The parsing subnets take the user's commands in natural language and parse them into semantic frames where the slots are filled via binding by STP. It is a stack-based system and the stack also binds by STP. The semantic result then leads to goals being set within the agent. Goals are context dependent, so a command like Turn toward the pyramid. needs to bind the goal to the location of the pyramid. This is done dynamically in a fashion similar to active links.
Similarly, a second parser has been developed that uses binding by STP for the stack and binding by compensatory LTP to fill the semantic frames. This indicates that these two variable binding mechanisms can be combined.
Referring back to associative memory (Section 2.1), both STP and synchrony have been proposed as mechanisms for supporting associative memory formation. There has been solid simulation work in the support of hetero-associative memory formation by synchrony (Shastri Citation2002; Gunay and Maida Citation2006). This avoids the stability plasticity dilemma by making bindings plastic and forgettable and hetero-associative memories permanent. It has also been proposed that short-term connection strength can be used to support long-term memory formation (Kaplan et al. Citation1991). Finally, there have been simulations that show active links also support long-term memory formation (van der Velde and de Kamps Citation2006).
6. Discussion
This paper has shown how two mechanisms for binding by synaptic change function. It has shown that one, STP, is capable of handling cross-talk and accounting for compositional semantics, and has inferred that the other mechanism, compensatory LTP, can too. Consequently, these new binding mechanisms can account for the problems described in Section 2.1. Elsewhere, it is shown how the earlier binding mechanisms, synchrony (Shastri and Aijanagadde Citation1993) and active links (van der Velde and de Kamps Citation2006), can solve these problems.
Since all four binding mechanisms are capable of binding, how do they differ? Below, each mechanism is evaluated on three important binding properties.
6.1. Binding properties
Both binding by compensatory LTP and binding by STP as described in this paper have values associated with the properties of Section 2.2. gives a qualitative overview of these values and those associated with binding by synchrony and by active links. The first column refers to the duration of the binding, the second to the number of different bindings that can be supported, and the third to speed to bind.
Table 2. Binding property values by method.
The persistence of binding for synchrony, and for active links is based solely on neural firing. With synchrony, the binding persists while the bound items fire. With active links, the binding persists while the binding node is firing.
With binding by STP, the binding lasts as long as there is synaptic support for it. In the above simulations using binding by STP, synaptic weights are reduced by 0.004 each cycle they are not increased. Therefore, the weights are completely erased in 250 cycles, and may be effectively erased in less; this equates to 2.5 s.
The persistence of binding by compensatory LTP is more difficult to calculate. In Section 4.2, 6000 cycles (four erase epochs and two unbound test epochs) were used to erase the binding, or 60 s. Spontaneous activation in the bind subnet leads to the connections from the subnet being erased. However, strong CAs can remain relatively stable under spontaneous activation due to the relative stability of compensatory LTP. When there is spontaneous activation of a small number of neurons, there are many more applications of anti-Hebbian learning than of Hebbian learning. Therefore, the total synaptic weight, W i , is significantly below the goal weight W B . This means that application of the anti-Hebbian rule changes the weights very little, and makes the original weights surprisingly stable.
The number of separate bindings differs between the four binding mechanisms. It is not clear how many bindings can be supported by synchrony, but one simulation sets the limit at 10 (Henderson Citation1994). At the other extreme, binding by compensatory LTP supports a practically unlimited set of bindings. In the first simulation, there is only one binding, but more could easily be modelled. Binding by LTP supports a number of bindings on the order of the number of neurons. (As the number of synapses leaving a neuron is bounded by a constant, the bits per synapse is constant, and these represent the memory of the system, memory is limited to O(n) bits where n is the number of neurons (Shannon Citation1948). Repeating the experiment from Section 4.2 on orthogonal bindings would give O(n) bindings.) There is no other practical limit for the number of bindings except perhaps time to erase. The binding by STP mechanism that was used in the above simulations also supports a practically unlimited number of binding nodes, though again time is a factor. Section 5.2 shows that simultaneous support for 40 bindings is straight forward. Of course, there can be multiple orthogonal sets of these bindings with, for instance, colour and object, and verb and object, being bound. This would lead to a set of bindings on the order of the number of neurons.
For compensatory LTP, the values regarding time to bind are quite clear. The fLIF model equates one cycle with 10 ms. Therefore, in Section 4.2, it takes roughly 1000 cycles to bind, so roughly 10 s.
Compared with this, binding via STP is quite rapid. In the simulations in Sections 4.3 and 5 the learning rate is set to 0.1 and the weight is clipped at 1; so binding happens in 10 cycles, and this equates to times about 100 ms. This contradicts the statement ‘it is unlikely that there exist mechanisms that support widespread structural changes and growth of new links within’ hundreds of ms (Shastri and Aijanagadde Citation1993). There is biological evidence of STP based on short bursts of spikes that persist for seconds to minutes (Hempel et al. Citation2000).
There is a vast range of evidence for synaptic changes of short duration (see Zucker and Regehr Citation(2002) for a review), and there are a wide range of behaviours, including different behaviours for neurons in different portions of the brain (Castro-Alamancos and Connors Citation1997). Evidence shows that short-term synaptic change can persist from under a second to over 30 (Varela et al. Citation1997). It has been shown that as few as 10 spikes at 50 Hz can lead to STP of synapses (Tecuapetla, Crillo-Reid, Bargas, and Galarraga Citation2007). In the simulations described in this paper, that would be 10 sets of neural firings in alternating cycles. For all that is known to the contrary, it is possible that the relevant form of rapid binding could be implemented by synaptic change. Bursts of 100 Hz firings for as little as 300 ms. leads to STP that endures for tens of minutes (Schulz and Fitzgibbons Citation1997).
It should also be noted that the time courses of the binding by STP and binding by compensatory LTP are affected by the constants, topologies, and presentation mechanics. The above simulations provide example time courses.
Binding by synchrony can occur in tens of ms (Wennekers and Palm Citation2000). As active links take only a few neural firings to form a binding, they too should occur on the order of tens of ms (van der Velde and de Kamps Citation2006).
A related property is the number of items per binding. The compensatory LTP mechanism limits this, but binding by STP and binding by synchrony do not. Binding by active links allows the developer to program this.
6.2. Maintaining binding by firing vs. by synapses
It has been stated that ‘the number of dynamic bindings expressed via some form of activity (e.g. synchrony) will be comparable with the number of ignited (fired) CAs’. If bindings are maintained by neural firing, this is the case, so it is the case for both binding by synchrony and active links. However, if binding is done by synaptic modification, CAs do not need to be active to remain bound; consequently, synaptic modification allows a much larger range of bindings to be supported.
If all of the bound items remain active, as in synchrony, or all of the binding nodes remain active, as in active links, a large number of items are active. This can lead to problems of crosstalk. These can be addressed programmatically, but it is clearly useful to be able to deactivate CAs and retain bindings.
Furthermore, maintaining a binding created by synaptic change, requires fewer neurons firing, and neural firing is biologically expensive (Attwell and Laughlin Citation2001; Aiello and y Rita Citation2002). Maintaining bindings by firing is thus biologically expensive. It costs a lot of energy.
Therefore, binding by firing may be useful, but it comes at a cost. However, binding by synaptic change has to pay much less.
6.3. Binding and memory
The three properties, speed to bind, number of bindings supported, and speed to unbind are also issues of general memory formation. Recall that CAs give an explanation for short-term memory (CA activation and persistence) and long-term memory (stable state CA formation based on LTP). CA activation happens quickly (<20 ms), but does not last long (seconds). CAs form more slowly, perhaps over days, but last much longer, perhaps years. CA activation and CA formation are akin to speed to bind as all involve a memory formation. The cessation of a CA firing, and the loss of a stable state are akin to a binding being erased as all involve the loss of memory.
While there is some debate as to whether memories are lost or not, it is largely accepted that as time passes, memories become less accessible (Klatzky Citation1980). shows the amount of memory that can be accessed as time progresses by different neural memory processes. This figure is meant to be a qualitative guide of the process indicating that as time passes fewer memories from a particular time can be accessed. At the left of the figure, CA activation (CAA) does not last long, but in a given period (say an hour) many memories can be used. On the right, CA formation (CAF) shows that memories last a long time, but not many things (relative to the number of CAs accessed) can be stored. Without binding, this leaves the middle ground empty; how can something be forgotten after only a day? Binding fills in this middle area. Many items may be bound by synchrony (BSyn) and by active links; there are fewer than the CAs that are active, and they can persist longer as only one of the base CAs is needed to keep the binding. Binding by STP (BSTP) probably occurs less frequently because it requires a modification of longer duration, but it persists longer than binding by neural firing. Finally, binding by compensatory LTP (or any LTP) has fewer items bound, but persists longer yet. Therefore, over a given hour, 1000 CAs might activate, 100 sets of CAs might be bound via synchrony, 20 bound by STP, 10 bound by LTP and two new CAs might be created. The active CAs would persist for 1 min, the synchronous bindings for two, the bindings by STP for 5 min, the bindings by LTP for 2 h, one new CA might last for a month and the other for 10 years.
Figure 10. Memory hierarchy: different binding mechanisms provide a possible answer for the wide range of memory duration CAA, CA activation; BSyn, binding via synchrony; AL, binding via active links; BSTP, binding by STP; BLTP, binding by LTP; CAF, CA formation.
These memory mechanisms use and are influenced by the dual dynamics of CA activation and CA formation. One good example of the complexity of these dual dynamics is the erasing of the binding by compensatory LTP in Section 4.2. The weights from the bound letter to the bind subnet are not changed during erasing. When the letter is presented after erasing, the synaptic weights to the bind subnet are high, but they go down rapidly; there is a decline because the neurons in the letter CA are firing and the bind neurons are not, and the decline is rapid because the total synaptic strength is high. This rapid decline completes the erasing. The dynamics also have an effect on the stability of existing CAs and formation of new CAs.
Biological neural systems are always learning (Churchland and Sejnowski Citation1992), and there is always spontaneous firing. Under these conditions, CAs must activate relatively frequently to keep their mutual synaptic strength high. It does not seem reasonable that all CAs are activated relatively frequently. The relative stability of compensatory LTP bindings with spontaneous activation provides some hope that this problem may be resolved, but it has not yet been since the system either is stable without spontaneous activation, or plastic with, but in neither case both.
Binding by synchrony, active links, and STP have a lesser effect on CA stability and plasticity, but they still have an effect. They have less of an effect because they are not based on long-term synaptic change. They still have an effect because they cause the simultaneous firing of neurons in CAs, and this will lead to increased permanent synaptic weight between the bound CAs. This might lead to the CAs recruiting each other, so that they no longer can be independently active.
Binding by compensatory LTP can now be looked at as an associative memory mechanism. CAs that are frequently bound may become more related but, perhaps due to topology, may not recruit each other. Other options for resolving stability problems include modified spontaneous activation mechanisms, subassemblies, and learning rules involving fatigue. In the simulations described in this paper, spontaneous activation is purely random; this might be modified to make neurons fire when they have not fired for a long time, and these neurons might co-fire based on their last activity. Subassemblies are merely sets of neurons that do not persist, but can be activated by spontaneous activation leading to synaptic support. Finally, if synaptic weights only changed significantly when neurons were fatigued, spontaneous activation would have little effect on them. These mechanisms are, of course, speculative.
Binding by compensatory LTP, and to a lesser extent the other binding mechanisms, provides a window into the stability plasticity dilemma of associative memory. It is relatively easy to model the indefinite storage of memories as once stored all memories are stable. When the memory store is large, this may cause no obvious problems. However, access to all memories can not be retained, and access to psychological memories is lost on a range of scales. Perhaps binding by compensatory LTP will provide an answer to how memories can be forgotten after days or years.
7. Conclusion
Binding is an important problem because a solution to it allows a system to have compositional syntax and semantics. This composition is necessary for a system to model the full range of human behaviour. If the particular problems of binding features in an object, frames, and rules can be solved, then a system can be built that is compositional.
This paper has introduced a new variable binding mechanism, binding by STP and made use of the relatively novel variable binding by compensatory LTP. Simulations have shown that these mechanisms, like synchrony and active links, can bind features in an object, and implement rules and frames. Simulations have shown that binding by STP also solves the problem of two and that binding by LTP should be able to.
Binding by STP is fast to bind, persists beyond the activity of the bound CAs, is relatively easy to engineer, and works consistently. Binding by compensatory LTP works, but faces the stability plasticity dilemma. It is slower to bind and the bindings persist longer. Neither of these mechanisms faces a combinatorial explosion to bind items, and both can support a very large number of bindings.
Binding via compensatory LTP and by STP can be used together and with the earlier defined binding mechanisms, binding via synchrony and binding by active links, to complement each other. They each have different behaviours on time to bind, time to erase, and capacity. Along with CA activation and CA formation, these binding mechanisms give a wide range of memory formation and retention behaviour.
Together, these mechanisms allow for a sophisticated use of compositional syntax and semantics in a simulated neural system. This will support the development of complex symbol processing agents from simulated neurons bridging the gap between subsymbolic and symbolic systems.
Acknowledgements
This work was supported by EPSRC grant GR/R13975/01 and EP/D059720. Thanks go to Dan Diaper, Thomas Wennekers, Pieter DeVries, Stephen Kaplan and the SESAME group, and anonymous reviewers for comments on this article.
References
- Abeles , M. 1991 . Corticonics: Neural Circuits of the Cerebral Cortex , Cambridge , , UK : Cambridge University Press .
- Abeles , M. , Bergman , H. , Margalit , E. and Vaddia , E. 1993 . Spatiotemporal Firing Patterns in the Frontal Cortex of Behaving Monkeys . Journal of Neurophysiology , 70 ( 4 ) : 1629 – 1638 .
- Ackley , D. , Hinton , G. and Sejnowski , T. 1985 . A Learning Algorithm for Boltzmann Machines . Cognitive Science , 9 : 147 – 169 .
- Aiello , G. and y Rita , P. B. 2002 . The Cost of an Action Potential . Journal of Neuroscience Methods , 103 ( 2 ) : 145 – 149 .
- Amit , D. 1989 . Modelling Brain Function: The World of Attractor Neural Networks , Cambridge , , UK : Cambridge University Press .
- Amit , D. and Brunel , N. 1997 . Model of Global Spontaneous Activity and Local Structured Activity During Delay Periods in the Cerebral Cortex . Cerebral Cortex , 7 : 237 – 252 .
- Anderson , J. and Lebiere , C. 1998 . The Atomic Components of Thought , Mahwah , NJ : Lawrence Erlbaum .
- Attwell , D. and Laughlin , S. 2001 . An Energy Budget for Signaling in the Gray Matter of the Brain . Journal of Cereberal Blood Flow Metabolism , 21 ( 10 ) : 1133 – 1145 .
- Bevan , M. and Wilson , C. 1999 . Mechanisms Underlying Spontaneous Oscillation and Rhythmic Firing in Rat Subthalamic Neurons . Neuroscience , 19 : 7617 – 7628 .
- Bienenstock , E. and Malsburg , C. V.D. 1987 . A Neural Network for Invariant Pattern Recognition . Europhysics Letters , 4 ( 1 ) : 121 – 126 .
- Braitenberg , V. 1989 . “ Some Arguments for a Theory of Cell Assemblies in the Cerebral Cortex ” . In Neural Connections, Mental Computation , Edited by: Nadel , L. , Cooper , L. , Culicover , P. and Harnish , R. Cambridge , MA : MIT Press .
- Browne , A. and Sun , R. 1999 . Connectionist Variable Binding . Expert Systems , 16 ( 3 ) : 189 – 207 .
- Brunel , N. 1996 . Hebbian Learning of Context in Recurrent Neural Networks . Neural Computation , 8 : 1677 – 1710 .
- Buonomano , D. 1999 . Distinct Functional Types of Associative Long-term Potentiation in Neocortical and Hippocampal Pyramidal Neurons . Neuroscience , 19 ( 16 ) : 6748 – 6754 .
- Carpenter , G. and Grossberg , S. 1988 . The Art of Adaptive Pattern Recognition by a Self-organizing Neural Network . IEEE Computer , 21 : 77 – 88 .
- Castro-Alamancos , M. and Connors , B. 1997 . Distinct Forms of Short-term Plasticity at Excitatory Synapses of Hippocampus and Neocortex . Proceedings of the National Academy of Sciences USA , 94 : 4161 – 4166 .
- Chacron , M. , Pakdaman , K. and Longtin , A. 2003 . Interspike Interval Correlations, Memory, Adaptation, and Refractoriness in a Leaky Integrate-and-fire Model with Threshold Fatigue . Neural Computation , 15 : 253 – 278 .
- Churchland , P. and Sejnowski , T. 1992 . The Computational Brain , Cambridge , MA : MIT Press .
- Dayan , P. and Abbott , L. 2005 . Theoretical Neuroscience: Computational and Mathematical Modeling of Neural Systems , Cambridge , MA : MIT Press .
- deVries , P. 2004 . Effects of Binding in the Identification of Objects . Psychological Research , 69 : 41 – 66 .
- Eccles , J. 1986 . Chemical Transmission and Dale's Principle . Progress in Brain Research , 68 : 3 – 13 .
- Eckhorn , R. , Bauer , R. , Jordan , W. , Brosch , M. , Kruse , W. , Munk , M. and Reitboeck , H. 1988 . Coherent Oscillations: A Mechanism of Feature Linking in the Visual Cortex? . Biological Cybernetics , 60 : 121 – 130 .
- Feldman , J. 1982 . Dynamic Connections in Neural Networks . Biological Cybernetics , 46 : 27 – 39 .
- Fillmore , C. 1968 . “ The Case for Case ” . In Universals in Linguistic Theory , Edited by: Back , E. and Harms , R. New York : Holt, Rinheart and Winston Inc .
- Fodor , J. and Pylyshyn , Z. 1988 . Connectionism and Cognitive Architecture: A Criticial Analysis . Cognition , 28 : 3 – 71 .
- Frixione , M. , Spinelli , G. and Gaglio , S. 1989 . Symbols and Subsymbols for Representing Knowledge: A Catalogue Raisonne . Proceedings of the Eleventh International Joint Conference on Artificial Intelligence , : 3 – 7 .
- Furber , S. , Bainbridge , W. , Cumpstey , J. and Temple , S. 2004 . Sparse Distributed Memory Using n-of-m Codes . Neural Networks , 17 ( 10 ) : 1437 – 1451 .
- Fusi , S. 2008 . A Quiescent Working Memory . Science , 319 : 1495 – 1496 .
- Fusi , S. , Drew , P. and Abbott , L. 2005 . Cascade Models of Synaptically Stored Memories . Neuron , 45 : 599 – 611 .
- Gerstner , W. and Kistler , W. 2002 . Mathematical Formulations of Hebbian Learning . Biological Cybernetics , 87 : 404 – 415 .
- Gerstner , W. and van Hemmen , J. 1992 . Associative Memory in a Network of Spiking Neurons . Network , 3 : 139 – 164 .
- Gunay , C. and Maida , A. 2006 . Using Temporal Binding for Hierarchical Recruitment of Conjunctive Concepts Over Delayde Lines . Neurocomputing , 69 : 317 – 367 .
- Hadley , R. 2007 . Synchronous vs. Conjunctive Binding: A False Dichotomy? . Connection Science , 19 ( 2 ) : 111 – 130 .
- Harnad , S. 1990 . The Symbol Grounding Problem . Physica D , 42 : 335 – 346 .
- Hebb , D. O. 1949 . The Organization of Behavior , New York : John Wiley and Sons .
- Hempel , C. , Hartman , K. , Wang , X. , Turrigiano , G. and Nelson , S. 2000 . Multiple Forms of Short-term Plasticity at Excitatory in Rat Medial Prefrontal Cortex . Journal of Neurophysiology , 83 : 3031 – 3041 .
- Henderson , J. 1994 . Connectionist Syntactic Parsing Using Temporal Variable Binding . Journal of Psycholinguistic Research , 23 ( 5 ) : 353 – 379 .
- Hodgkin , A. and Huxley , A. 1952 . A Quantitative Description of Membrane Current and Its Application to Conduction and Excitation in Nerve . Journal of Physiology , 117 : 500 – 544 .
- Hofstadter , D. 1979 . Godel, Escher and Bach: An Eternal Golden Braid , New York : Basic Books .
- Hopcroft , J. and Ullman , J. 1979 . Introduction to Automata Theory, Languages and Computation , Reading , MA : Addison-Wesley .
- Hopfield , J. 1982 . Neural Nets and Physical Systems with Emergent Collective Computational Abilities . Proceedings of the National Academy of Sciences USA , 79 : 2554 – 2558 .
- Huyck , C. 2004 . Overlapping Cell Assemblies From Correlators . Neurocomputing , 56 : 435 – 439 .
- Huyck , C. 2007 . Creating Hierarchical Categories Using Cell Assemblies . Connection Science , 19 ( 1 ) : 1 – 24 .
- Huyck , C. 2008 . Cabot1: A Videogame Agent Implemented in fLIF Neurons . IEEE Systems, Man and Cybernetics Society , : 115 – 120 .
- Huyck , C. 2009 . A Psycholinguistic Model of Natural Language Parsing Implemented in Simulated Neurons . Cognitive Neurodynamics , : 1 – 14 . DOI: 10.1007/s11571-009-9080.6
- Huyck , C. and Belavkin , R. 2006 . Counting With Neurons: Rule Application With Nets of Fatiguing Leaky Integrate and Fire Neurons . Proceedings of the Seventh International Conference on Cognitive Modelling , : 142 – 147 .
- Huyck , C. and Bowles , R. 2004 . Spontaneous Neural Firing in Biological and Artificial Neural Systems . Journal of Cognitive Systems , 6 ( 1 ) : 31 – 40 .
- Huyck , C. and Fan , Y. 2007 . Parsing With fLIF Neurons’, in . IEEE Systems, Man and Cybernetics Society , : 35 – 40 .
- Jackendoff , R. 2002 . Foundations of Language: Brain, Meaning, Grammar, Evolution , Oxford , , UK : Oxford University Press .
- Kaplan , S. , Weaver , M. and French , R. 1990 . Active Symbols and Internal Models: Towards a Cognitive Connectionism . AI and Society , 4 : 51 – 71 .
- Kaplan , S. , Sontag , M. and Chown , E. 1991 . Tracing Recurrent Activity in Cognitive Elements (TRACE): A Model of Temporal Dynamics in a Cell Assembly . Connection Science , 3 : 179 – 206 .
- Klatzky , R. 1980 . Human Memory: Structures and Processes , Santa Barbara , CA : W. H. Freeman and Co .
- Knoblauch , A. and Palm , G. 2001 . Pattern Separation and Synchronization in Spiking Associative Memories and Visual Areas . Neural Networks , 14 : 763 – 780 .
- Knoblauch , A. , Markert , H. and Palm , G. 2004 . An Associative Model of Cortical Language and Action Processing’, in . Proceedings of the Ninth Neural Computation and Psychology Workshop , : 79 – 83 .
- Laird , J. , Newell , A. and Rosenbloom , P. 1987 . Soar: An Architecture for General Cognition . Artificial Intelligence , 33 ( 1 )
- Lansky , P. , Sanda , P. and He , J. 2006 . The Parameters of the Stochastic Leaky Integrate-and-fire Neuronal Model . Journal of Computational Neuroscience , 21 : 211 – 223 .
- Lewis , R. and Vasishth , S. 2005 . An Activation-based Model of Sentence Processing as Skilled Memory Retrieval . Cognitive Science , 29 ( 3 ) : 375 – 419 .
- Lindsey , R. 1988 . Can This Treatment Raise the Dead . Behavioral and Brain Sciences , 11 ( 1 ) : 41 – 42 .
- Maass , W. and Bishop , C. 2001 . Pulsed Neural Networks , Cambridge , MA : MIT Press .
- Malsburg , C. V.D. 1981 . “ The Correlation Theory of Brain Function ” . Dept. of Neurobiology, Max-Planck-Institute for Biophyscial Chemistry . Technical Report
- Malsburg , C. V.D. 1986 . “ Am I Thinking Assemblies? ” . In Brain Theory , Edited by: Palm , G. and Aertsen , A. 161 – 176 . Heidelberg : Springer-Verlag .
- McCulloch , W. and Pitts , W. 1943 . A Logical Calculus of Ideas Immanent in Nervous Activity . Bulletin of Mathematical Biophysics , 5 : 115 – 133 .
- Messinger , A. , Squire , L. , Zola , S. and Albright , T. 2005 . Neural Correlates of Knowledge: Stable Representation of Stimulus Associations Across Variations in Behavioural Performance . Neuron , 48 : 359 – 371 .
- Miyashita , Y. 1988 . Neuronal Correlate of Visual Associative Long-term Memory in the Primate Temporal Cortex . Nature , 335 : 817 – 820 .
- Mongillo , G. , Barak , O. and Tsodyks , M. 2008 . Synaptic Theory of Working Memory . Science , 319 : 1543 – 1546 .
- Newell , A. 1990 . Unified Theories of Cognition , Cambridge , MA : Harvard University Press .
- O'Neill , J. , Senior , T. , Allen , K. , Huxter , J. and Csicsvari , J. 2008 . Reactivation of Experience-dependent Cell Assembly Patterns in Hippocampus . Nature Neuroscience , 11 : 209 – 215 .
- Pulvermuller , F. 1999 . Words in the Brain's Language . Behavioral and Brain Sciences , 22 : 253 – 336 .
- Quillian , M. 1967 . Word Concepts: A Theory of Simulation of Some Basic Semantic Capabilities . Behavioral Science , 12 : 410 – 30 .
- Rumelhart , D. and McClelland , J. 1982 . An Interactive Activation Model of Context Effects in Letter Perception: Part 2. The Contextual Enhancement and Some Tests and Extensions of the Model . Psychological Review , 89 ( 1 ) : 60 – 94 .
- Rumelhart , D. and McClelland , J. 1986 . Parallel Distributed Processing , Cambridge , MA : MIT Press .
- Sakai , K. and Miyashita , Y. 1991 . Neural Organization for the Long-term Memory of Paired Associates . Nature , 354 : 152 – 155 .
- Schank , R. and Abelson , R. 1977 . Scripts, Plans, Goals and Understanding , Hillsdale , NJ : Lawrence Erlbaum .
- Schulz , P. and Fitzgibbons , J. 1997 . Differing Mechanisms of Expression for Short- and Long-term Potentiation . Journal of Neurophysiology , 78 : 321 – 334 .
- Shannon , C. 1948 . A Mathematical Theory of Communication . Bell System Technical Journal , 27 : 379 – 423 .
- Shastri , L. 2002 . A Computationally Efficient Abstraction of Long-term Potentiation . Neurocomputing , 44–46 : 33 – 41 .
- Shastri , L. 2006 . Comparing the Neural Blackboard and the Temporal Synchrony Based S\MakeUppercasehruti Architectures . Behaviour and Brain Science , 29 : 84 – 86 .
- Shastri , L. and Aijanagadde , V. 1993 . From Simple Associations to Systematic Reasoning: A Connectionist Representation of Rules, Variables, and Dynamic Bindings Using Temporal Synchrony . Behaviour and Brain Science , 16 : 417 – 494 .
- Shieber , S. 1986 . An Introduction to Unification-Based Approaches to Grammar , Stanford , CA : Center for the Study of Language and Information .
- Siegelmann , H. and Sontag , E. 1991 . “ On the Computational Power of Neural Nets ” . SYCON . Technical Report
- Smolensky , P. 1987 . Connectionist AI, Symbolic AI, and the Brain . Artificial Intelligence Review , 1 : 95 – 109 .
- Smolensky , P. 1990 . Tensor Product Variable Binding and the Representation of Symbolic Structures in Connectionist Systems . Artificial Intelligence , 46 : 159 – 216 .
- Sompolinsky , H. 1987 . The Theory of Neural Networks: The Hebb Rule and Beyond’, in . Proceedings of Heidelberg Colloquium on Glassy Dynamics , : 485 – 527 .
- Sougne , J. 2001 . Binding and Multiple Instantiation in a Distributed Network of Spiking Neurons . Connection Science , 13 : 99 – 126 .
- Tecuapetla , F. , Crillo-Reid , L. , Bargas , J. and Galarraga , E. 2007 . Dopaminergic Modulation of Short-term Synaptic Plasticity at Striatal Inhibitory Synapses . Proceedings of the National Academy of Sciences USA , 104(24) : 10258 – 10263 .
- Usher , M. and Donnelly , N. 1998 . Visual Synchrony Affects Binding and Segmentation in Perception . Nature , 394 : 179 – 182 .
- Valiant , L. 2005 . Memorization and Association on a Realistic Neural Model . Neural Computation , 17 : 527 – 555 .
- van der Velde , F. and de Kamps , M. 2006 . Neural Blackboard Architectures of Combinatorial Structures in Cognition . Behavioral and Brain Sciences , 29 : 1 – 72 .
- Varela , J. , Sen , K. , Gibson , J. , Abbott , L. and Nelson , S. 1997 . A Quantitative Description of Short-term Plasticity at Excitatory Synapses in Layer 2/3 of Rat Primary Visual Cortex . Journal of Neuroscience , 17 : 7926 – 7940 .
- Wennekers , T. and Palm , G. 2000 . “ Cell Assemblies, Associative Memory and Temporal Structure in Brain Signals ” . In Time and the Brain: Conceptual Advances in Brain Research , Edited by: Miller , R. Vol. 2 , 251 – 274 . Amsterdam : Harwood Academic Publishers .
- White , G. , Levy , W. and Steward , O. 1988 . Evidence that Associative Interactions Between Afferents During the Induction of Long-term Potentiation Occur Within Local Dendritic Domains . Proceedings of the National Academy of Sciences USA , 85 : 2368 – 2372 .
- Willshaw , D. , Buneman , O. and Longuet-Higgins , H. 1969 . Non-holographic Associative Memory . Nature , 222 : 960 – 962 .
- Zucker , R. and Regehr , W. 2002 . Short-term Synaptic Plasticity . Annual Review of Physiology , 64 : 355 – 405 .