
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Here, we firstly reported to generate panels of anti-idiotypic single-chain variable fragments (scFvs) of Bacillus thuringiensis Cry2Aa toxin by chain shuffling method. Light- and heavy-chain-shuffled libraries based on two parental clones were constructed. The signal of the light-chain-shuffled library was 4.2-fold higher than the heavy one. Solely after one round panning, eight mutants were rapidly isolated from the chain-shuffled libraries. The apparent affinity of the best clones was 2.65 × 106 M−1, a 2.59-fold improvement over the parental clone. The binding abilities between anti-idiotypic scFvs with brush border membrane vesicle of Plutella xylostella larvae were also determined. This work demonstrated that chain shuffling could rapidly generate panels of antibodies with similar binding abilities but of the distinct idiotypic composition. The serials of mutants can be used to map the binding epitope of the receptors in target insects instead of Cry2Aa toxin.
Introduction
Anti-idiotypic antibody is a useful tool for understanding the molecular mimicries with many applications. It has been produced to different antigens and used widely to react with cellular receptors, including hormone receptors (Lan et al., Citation2017; Lan, Zheng, Khan, & Li, Citation2015), virus receptors (Huang & Campadelli-Fiume, Citation1996; Kiyohara et al., Citation2009), harpin receptor (Chen et al., Citation2008) and chimeric immune receptors (Cheung, Guo, Modak, & Cheung, Citation2003).
The standard method for the generation of anti-idiotypic antibodies is the conventional hybridoma techniques. However, due to the idiotope (the antigen-binding region) of an antibody which is used to produce anti-idiotypic antibodies was only a small part of the whole antibody. The positive rate of hybridoma-secreting anti-idiotypic antibodies is much less than common antibodies. In addition to hybridoma techniques, the selection of anti-idiotypic antibodies by phage display is an alternative way. This method could rapidly isolate antibodies even without immunization (Tu et al., Citation2012). However, for the same reason, the isolation of anti-idiotypic antibodies from naïve library often results in none or few positive clones (Goletz et al., Citation2002).
Chain shuffling is a mutagenesis method, which allows the recombination of a heavy or light chain of a specific antibody with a complementary variable domain library (Irving, Kortt, & Hudson, Citation1996). Since the shuffling approaches mimic somatic hypermutation and could avoid the deleterious mutagenesis in the framework regions of antibodies, they may be better than random or site-directed mutagenesis to maintain the functional antibodies (Ness et al., Citation2002). This will be beneficial when rapidly generating panels of antibodies with related specificity but distinct idiotypic composition from a limited parental clone, and it has many applications (Kang, Jones, & Burton, Citation1991).
Chain shuffling could also optimize the paring of the heavy chain and light chain to improve the functions of antibodies. Depending on the antibodies, chain shuffling can result in the loss of binding affinity or generation of several new active combinations, some of which have improved affinity (Brockmann, Citation2010). The affinity improvement in some recent reports has been 1.4–15 folds for heavy-chain shuffling (Kramer, Citation2002; Marks et al., Citation1992; Rojas et al., Citation2004; Shi et al., Citation2010) and 2.1–185 folds for light-chain shuffling (Fitzgerald, Leonard, Darcy, Danaher, & O’Kennedy, Citation2011; Osbourn et al., Citation1996). In addition to the affinity, fine specificities and recognition of the epitope can also be changed by chain shuffling (Lu et al., Citation2003) or can overcome well-recognized protein-expression problems in bacteria (Rojas et al., Citation2004).
In previous work, we only isolated two anti-idiotypic scFvs (B10, F2) of Bacillus thuringiensis Cry2Aa toxin from Tomlinson I library (Wu, Citation2015). To obtain more functional antibodies with similar specificity and optimized paring of heavy and light chains, these two parental clones were shuffled with variable heavy- and light-chain repertoires of the same un-panned library.
Material and methods
Chemical and reagents
Two anti-idiotypic scFv clones in E. coli TG1 (labeled B10 and F2) were isolated from Tomlinson I library by our laboratory. Polyclonal antibodies of Cry2Aa toxin were produced from New Zealand white rabbits by our laboratory. Tomlinson I library (a naïve scFv library constructed based on a human framework with side-chain diversity), pIT2 vector, E. coli TG1 were purchased from Source Bioscience (UK). T4 DNA ligase, NotI and NcoI were obtained from New England Biolabs (USA). 2× Taq PCR master mix and 2× pfu PCR master mix were purchased from DBI (Germany) and Dongsheng Biotechnology (China), respectively. DNA gel extraction spin kits, 6-well plate and 96-well plate were purchased from Corning (USA). Skim milk powder was obtained from Solabio (China). HRP-conjugated anti-M13 monoclonal antibodies were purchased from GE Healthcare (USA). All reagents were of analytical grade.
Construction of chain-shuffled libraries
The variable heavy region (VH) and light region (VL) of B10, F2 or un-panned Tomlinson I library were amplified by PCR with 2× Taq PCR master mix. For the amplification of VH, primer 1 and primer 2 were used and for the case of VL, primer 3 and primer 4 were used (). The PCR cycling conditions were as follows: 1 cycle, 5 min at 95°C; 30 cycles, 1 min at 94, 1 min at 55°, 1 min at 72°C; 1 cycle, 10 min at 72°C.
Table 1. Sequence of the PCR primers used for construction of chain-shuffled librariesa.
The purified VH and VL PCR products were connected to form a full-length scFv gene in a strand overlap extension (SOE)-PCR reaction by using 2× pfu PCR master mix. The SOE-PCR programme was 1 cycle, 5 min at 95°C; 7 cycles of 1 min, 94°C, 1 min at 55°C, 1 min at 72°C, and then primer 1 and primer 4 were added to the reaction vessel, the SOE-PCR was continued with the following programme: 30 cycles, 1 min at 94°C, 1 min at 55°C, 1 min at 72°C; 1 cycle, 10 min at 72°C.
The SOE-PCR products and pIT2 (vector) were digested with Not I and Nco I at 37°C for 4 h and were purified by using DNA gel extraction spin kits. The digested DNA and pIT2 were ligated together by T4 DNA ligase at 16°C for 18 h. Then, the ligation products were transformed into TG1. The transformed cells were cultured in 50-mL 2× TY (containing 100 μg/mL ampicillin and 1% glucose) at 37°C, 250 RPM until the mid-exponential growth was achieved and subsequently rescued by the addition of M13KO7 helper phage. The culture was incubated in a 37°C water bath for 30 min and was spinned at 3000 × g, 10 min. The pellet was suspended in 100 mL 2×TY (containing 100 μg/mL ampicillin, 50 μg/mL kanamycin and 0.1% glucose) and shaking at 30°C, 250 rpm overnight. The next day, the phage was precipitated according to the protocol of Tomlinson (I + J) library (de Wildt, Mundy, Gorick, & Tomlinson, Citation2000; Ossysek et al., Citation2015).
Panning of chain-shuffled libraries
For the panning steps, a six-well plate was coated with 1 mL of 100 μg/mL polyclonal antibodies of Cry2Aa toxin in a carbonate–bicarbonate buffer saline (CBS, 50 mmol/L, pH 9.6) at 4°C overnight. The plate was washed three times with phosphate-buffered saline (PBS, 50 mmol/L, pH 7.4) containing 0.05% v/v Tween-20 and blocked by completely filling with 2% skim milk powder in PBS (MPBS) and incubating the plates at room temperate for 2 h. After washing the plate three times, 1 mL of 107 cfu/mL phage from the heavy- or light-chain-shuffled library in 2% MPBS were added to the well and incubated at room temperature for 2 h under gentle shaking. The subsequent washing steps, elution, multiplication of the phage, polyclonal and monoclonal phage ELISA were done according to the protocols of Tomlinson (I + J) library.
Ranking clones by competitive phage ELISA
The optimal working concentration of coating antigen (polyclonal antibodies of Cry2Aa toxin) and the dilution times of phage supernatant for each clone were determined by checkerboard titration (Liu et al., Citation2017). The 96-well plates were coated with the optimal working concentration of polyclonal antibodies of Cry2Aa toxin in CBS. The next day, the plates were washed three times with PBS containing 0.05% v/v Tween-20 (PBST) by automated microplate washer and blocked by 2% MPBS (200 μL/well) and incubated for 1 h at 37°C. Following the incubation, 50 μL of varying concentrations of polyclonal antibodies of Cry2Aa toxins were incubated with 50 μL of the optimal dilution of scFvs supernatants and added to each well. Following an incubation for 1 h at 37°C, the plates were washed, 100 μL/well HRP-conjugated anti-M13 monoclonal antibodies (1:5000 diluted by PBS) were added and incubated for 1 h at 37°C. Following a washing step, 100 μL/well of a tetramethylbenzidine solution (120 μL of 10 mg/mL TMB-DMSO and 30 μL of 0.65% (v/v) H2O2 diluted with 11.85 mL of citrate-sodium citrate buffer (CPBS, 100 mmol/L, pH5.5)) was added. The reaction was stopped after 15 min by adding 50 μL/well of 2 mol/L H2SO4, and the absorbance was read by a microplate reader at 450 nm.
Dose–response curves were analysed by using software Sigmaplot version 12.0. The ratios of ELISA mean absorbance values (n = 3) of antibody-binding response in the presence of free antigen to the absence of antigen (Y, B/B0) were plotted against the logarithm of free antigens concentrations (X) and fitted to the following four-parameter logistic equation (Liu et al., Citation2007).
where A is the high asymptote, B is the slope, C is the antigen concentration that was required to inhibit 50% maximal binding (IC50), and D is the low asymptote.
Brush border membrane vesicle preparation and ELISA binding analysis
Brush border membrane vesicles (BBMVs) were prepared from midgut tissue of fourth instar Plutella xylostella larvae following the protocol described by Wolfersberger (Citation1993). Binding ability of anti-idiotypic phage antibodies of Cry2Aa toxin with Plutella xylostella BBMV was determined by ELISA. First, 96-well plates were coated with 10 μg/mL BBMV. Then, after blocking with 2% MPBS, 106 phages (precipitated by PEG/NaCl and resolved in PBS) for each clones for each clones (n = 3) were added to the well. The bound phage antibodies were detected using HRP-conjugated anti-M13 monoclonal antibodies (1:5000 diluted by PBS). The subsequent steps were performed as a competitive phage ELISA. PBS and an irrelevant phage clone which recognize bovine serum albumin (BSA) from Tomlinson (I + J) library were used as negative controls.
Results and discussion
Constructions of chain-shuffled library
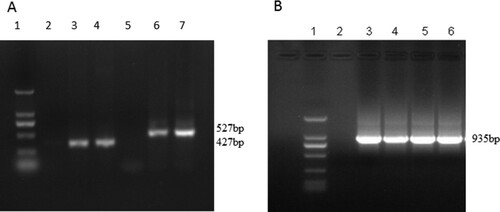
The amplified VL and VH genes of B10, F2 and Tomlinson I library were analysed by agarose gel electrophoresis ((A)). The calculated sizes of VH and VL products were approximately 527 and 427 bp, respectively. The base length of SOE-PCR products was approximately 935 bp ((B)). The sizes of constructed heavy- and light-chain-shuffled libraries were determined to be 1.4 × 106 cfu and 1.2 × 106 cfu.
Figure 1. Analysis of amplified VH, VL and SOE-PCR products by agarose gel electrophoresis. (A) 1:DNA ladder, from top to bottom, 2000,1000,750, 500, 250, 100 bp; 3–4:VL (Tomlinson I); 6:VH (B10); 7: VH (F2). (B) 1: DNA ladder, from top to bottom, 2000, 1000, 750, 500, 250, 100 bp 3: VL (Tomlinson I)-VH (B10); 4: VL (Tomlinson I)-VH (F2); 5: VL (B10)-VH (Tomlinson I); 6: VL (F2)-VH (Tomlinson).
Isolation of mutants from chain-shuffled libraries
Before panning, the whole affinity of heavy- and light-chain-shuffled libraries was determined by polyclonal phage ELISA. The absorbance of light-chain-shuffled library was 4.2-fold higher than heavy-chain-shuffled library. It is thus demonstrated that in the present study, the heavy chains played a more important role than light chains for the binding ability.
After one round of panning, the output clone numbers for the heavy- and light-chain-shuffled libraries were 59 and 182, respectively. These clones were screened by monoclonal phage ELISA, in which 8 and 93 positive clones were isolated from the heavy- and light-chain-shuffled library. The positive rates for the two libraries were 13.6% and 51.1%. The high-positive rate may benefit the more functional antibodies from the chain-shuffled libraries than the libraries constructed by random mutagenesis.
After DNA sequencing, we finally obtained one heavy-chain-shuffled clones (2H1 originated from B10) and seven light-chain-shuffled clones (MTU10, MUT19, MUT21 originated from B10; 2F10, 3B1, 3F1 and 3H11 originated from F2) (). By comparing the sequence of mutants with their parental clones, we found that all the mutants have the same length of CDR loops as the parental clones, and seven clones (MUT10, MUT19, MUT21, 2F10, 3F1, 3H11 and 3B1) have one to five different amino acids with their parental clones, which probably arose by a somatic mutation from the same initial B-cell clones. The heavy-chain-shuffled clone (2H1) has 11 different amino acids in CDR regions, which may have derived from a different germline family.
Table 2. Protein sequences of parental clones and mutants derived from chain-shuffled librariesa.
Affinity determination
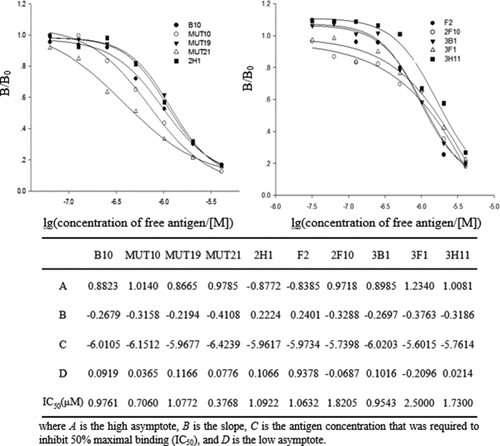
Affinity is an essential determinant of antibody function. The affinity of mutants could be rapidly ranked by a competitive phage ELISA even when the concentrations of scFvs are unknown (Clackson & Lowman, Citation2004). The ratios of the antigen concentration that required to inhibit 50% maximal binding (IC50) of parental clones to mutants obtained from competitive phage ELISA were a close approximation to the ratio of affinities; it permitted the ranking of the binding activities of mutants.
IC50 and parameters of mutants originated from B10 and F2 are summarized in . The IC50 of the best mutants (MUT21 from B10 and 3B1 from F2) were 0.3768 and 0.9543 μM. The ratios of IC50(B10)/IC50 (MUT21) and IC50(F2)/IC50(3B1) were 2.59 and 1.11, MUT21 showed a moderately improved affinity by competitive phage ELISA. The apparent affinity is defined as the reciprocal of the concentration required to IC50 in a competitive ELISA. Hence, the apparent affinity for the best clone (MUT21) was 2.65 × 106 M−1.
Figure 2. Determination of the apparent affinity of mutants and parental clones by competitive phage ELISA (n = 3). B/B0 stands for the ratio ELISA mean absorbance values of antibody-binding response in the presence of free antigen to the absence of antigen. Various concentrations of free antigen inhibited the binding of phage antibodies with the coating antigen, the signal was detected by using HRP-conjugated anti-M13 monoclonal antibodies. The calibration curves were constructed by using a four-parameter equation by Sigmaplot 12.0.
In this study, we do not isolate highly improved clones from chain-shuffled libraries, which may be caused by the low diversity of the VH, VL genes repertories from the same non-immunized library. The total diversity of Tomlinson I library is 1.47 × 108. If the diversity of light chain and heavy chain was equal, the estimated diversity of light chain or heavy should have been around 1.21 × 104 members. This indicated that the diversity of the heavy- and light-chain-shuffled libraries was approximately 2.42 × 104 (two parental clones). This may be not sufficient to isolate highly improved clones.
Binding analysis of anti-idiotypic antibodies to BBMV of Plutella xylostella
Plutella xylostella is a target insect of Cry2Aa toxin. If these anti-idiotypic scFvs mimicked the toxin-receptor binding surface of Cry2Aa toxin, it could be used to map the toxin-binding site instead of Cry2Aa toxin. In , the binding ability of these anti-idiotypic antibodies to BBMV of Plutella xylostella were determined. Generally, F2 and its mutants (2F10, 3B11, 3F1, 3H11) showed weaker binding ability than B10 and its mutants. MUT21 (a mutant of B10) was the strongest binder to BBMV of Plutella xylostella larvae by phage ELISA. The discrepant signal of phage antibodies may be due to the fact that they mimicked the toxin epitopes, which have different distance or affinity to the binding site. The specific characters and application of these anti-idiotypic scFvs are under investigation by our group and will be discussed in the future publications.
Figure 3. Determination of binding ability of anti-idiotypic phage antibodies with BBMV of Plutella xylostella larvae by ELISA (n = 3). For each clones, 106 phages/well were added to ELISA plate coated with BBMV and detected by HRP-conjugated anti-M13 monoclonal antibodies. Blank is the mean signal of wells which had been added PBS instead of phage antibodies. BSA is an irrelevant phage antibody from Tomlinson (I + J) library which recognizes BSA.
Conclusion
The present work indicates that the chain shuffling could rapidly generate panels of antibodies with similar binding abilities but of distinct idiotypic compositions. Moreover, mutants with moderately improved affinities may also be obtained by the chain shuffling, even with the same naïve library where the antibody was originally isolated. The serials of mutants can be used to map the binding epitopes of the receptor in target insects instead of the Cry2Aa toxin or to be used as a cost-efficient surrogate molecule instead of the toxin in ELISA kit for toxin detection.
Disclosure statement
No potential conflict of interest was reported by the authors.
Additional information
Funding
Related Research Data
References
- Brockmann, E. C. (2010). Evolution of bioaffinity reagents by phage display (PhD Thesis). University of Turku, Turku.
- Chen, D. F., Hu, B. S., Qian, G. L., Zhang, Q., Gu, C. Y., Fan, J. Q., & Liu, F. Q. (2008). A novel probe for harpin receptor in nonhost plants: Monoclonal anti-idiotypic antibodies as internal images of harpin(Xoo) active sites. Physiological and Molecular Plant Pathology, 73, 138–146. doi: https://doi.org/10.1016/j.pmpp.2009.05.001
- Cheung, N. K., Guo, H. F., Modak, S., & Cheung, I. Y. (2003). Anti-idiotypic antibody facilitates scFv chimeric immune receptor gene transduction and clonal expansion of human lymphocytes for tumor therapy. Hybridoma and Hybridomics, 22, 209–218. doi: https://doi.org/10.1089/153685903322328938
- Clackson, T., & Lowman, H. B. (2004). Phage display: A practical approach. Oxford: Oxford University Press.
- de Wildt, R. M. T., Mundy, C. R., Gorick, B. D., & Tomlinson, I. M. (2000). Antibody arrays for high-throughput screening of antibody-antigen interactions. Nature Biotechnology, 18, 989–994. doi: https://doi.org/10.1038/79494
- Fitzgerald, J., Leonard, P., Darcy, E., Danaher, M., & O’Kennedy, R. (2011). Light-chain shuffling from an antigen-biased phage pool allows 185-fold improvement of an anti-halofuginone single-chain variable fragment. Analytical Biochemistry, 410, 27–33. doi: https://doi.org/10.1016/j.ab.2010.11.009
- Goletz, S., Christensen, P. A., Kristensen, P., Blohm, D., Tomlinson, I., Winter, G., & Karsten, U. (2002). Selection of large diversities of antiidiotypic antibody fragments by phage display. Journal of Molecular Biology, 315, 1087–1097. doi: https://doi.org/10.1006/jmbi.2001.5314
- Huang, T., & Campadelli-Fiume, G. (1996). Anti-idiotypic antibodies mimicking glycoprotein D of herpes simplex virus identify a cellular protein required for virus spread from cell to cell and virus-induced polykaryocytosis. Proceedings of the National Academy of Sciences of the United States of America, 93, 1836–1840. doi: https://doi.org/10.1073/pnas.93.5.1836
- Irving, R. A., Kortt, A. A., & Hudson, P. J. (1996). Affinity maturation of recombinant antibodies using E-coli mutator cells. Immunotechnology, 2, 127–143. doi: https://doi.org/10.1016/1380-2933(96)00044-9
- Kang, A. S., Jones, T. M., & Burton, D. R. (1991). Antibody redesign by chain shuffling from random combinatorial immunoglobulin libraries. Proceedings of the National Academy of Sciences of the United States of America, 88, 11120–11123. doi: https://doi.org/10.1073/pnas.88.24.11120
- Kiyohara, T., Totsuka, A., Yoneyama, T., Ishii, K., Ito, T., & Wakita, T. (2009). Characterization of anti-idiotypic antibodies mimicking antibody- and receptor-binding sites on hepatitis A virus. Archives of Virology, 154, 1263–1269. doi: https://doi.org/10.1007/s00705-009-0433-6
- Kramer, K. (2002). Evolutionary affinity and selectivity optimization of a pesticide-selective antibody utilizing a hapten-selective immunoglobulin repertoire. Environmental Science and Technology, 36, 4892–4898. doi: https://doi.org/10.1021/es010209s
- Lan, H. N., Hong, P., Li, R. N., Suo, L., Shan, A. S., Li, S., & Zheng, X. (2017). Internal image anti-idiotypic antibody: A new strategy for the development a new category of prolactin receptor (PRLR) antagonist. Molecular Immunology, 87, 86–93. doi: https://doi.org/10.1016/j.molimm.2017.04.006
- Lan, H., Zheng, X., Khan, M. A., & Li, S. (2015). Anti-idiotypic antibody: A new strategy for the development of a growth hormone receptor antagonist. The International Jounal Biochemistry and Cell Biology, 68, 101–108. doi: https://doi.org/10.1016/j.biocel.2015.09.004
- Liu, Y., Lin, M. M., Zhang, X. F., Hu, X. D., Lin, J. R., Hao, J., … Liu, X. J. (2017). Development of competitive ELISA for the detection of bovine serum albumin using single-chain variable fragments. Analytical Biochemistry, 525, 89–91. doi: https://doi.org/10.1016/j.ab.2017.03.003
- Liu, Y., Zhang, C.Z., Yu, X.Y., Zhang, Z. Y., Zhang, X., Liu, R. R., Liu X.J., Gong, Z. M. (2007). Development and evaluation of immunoassay for zeranol in bovine urine. Jounal of Zhejiang University-Science B, 8, 900-905. doi: https://doi.org/10.1631/jzus.2007.B0900
- Lu, D., Shen, J. Q., Vil, M. D., Zhang, H. F., Jimenez, X., Bohlen, P., … Zhu, Z. P. (2003). Tailoring in vitro selection for a picomolar affinity human antibody directed against vascular endothelial growth factor receptor 2 for enhanced neutralizing activity. Journal of Biological Chemistry, 278, 43496–43507. doi: https://doi.org/10.1074/jbc.M307742200
- Marks, J. D., Griffiths, A. D., Malmqvist, M., et al. (1992). By-passing immunization: Building high affinity human antibodies by chain shuffling. Nature Biotechnology, 10, 779–783. doi: https://doi.org/10.1038/nbt0792-779
- Ness, J. E., Kim, S., Gottman, A., Pak, R., Krebber, A., Borchert, T. V., … Minshull, J. (2002). Synthetic shuffling expands functional protein diversity by allowing amino acids to recombine independently. Nature Biotechnology, 20, 1251–1255. doi: https://doi.org/10.1038/nbt754
- Osbourn, J. K., Field, A., Wilton, J., Derbyshire, E., Earnshaw, J. C., Jones, P. T., … McCafferty, J. (1996). Generation of a panel of related human scFv antibodies with high affinities for human CEA. Immunotechnology, 2, 181–196. doi: https://doi.org/10.1016/S1380-2933(96)00046-2
- Ossysek, K., Uchanski, T., Kulesza, M., Bzowska, M., Klaus, T., Wos, K., … Bereta, J. (2015). A new expression vector facilitating production and functional analysis of scFv antibody fragments selected from tomlinson I plus J phagemid libraries. Immunology Letters, 167, 95–102. doi: https://doi.org/10.1016/j.imlet.2015.07.005
- Rojas, G., Talavera, A., Munoz, Y., Rengifo, E., Krengel, U., Angstrom, J., Gavilondo, J., Moreno, E. (2004). Light-chain shuffling results in successful phage display selection of functional prokaryotic – expressed antibody fragments to N-glycolyl GM3 ganglioside. Journal of Immunological Methods, 293, 71-83. doi: https://doi.org/10.1016/j.jim.2004.07.002
- Shi, L., Wheeler, J. C., Sweet, R. W., Lu, J., Luo, J. Q., Tornetta, M., … Tsui, P. (2010). De novo selection of high-affinity antibodies from synthetic fab libraries displayed on phage as pIX fusion proteins. Journal of Molecular Biology, 397, 385–396. doi: https://doi.org/10.1016/j.jmb.2010.01.034
- Tu, Z., Xu, Y., He, Q. H., Fu, J. H., Liu, X., & Tao, Y. (2012). Isolation and characterisation of deoxynivalenol affinity binders from a phage display library based on single-domain camelid heavy chain antibodies (VHHs). Food and Agricultural Immunology, 23, 123–131. doi: https://doi.org/10.1080/09540105.2011.606560
- Wolfersberger, M. G. (1993). Preparation and partial characterization of amino-acid transporting brush border membrane-vesicles from the larval midgut of the gypsy-moth (lymantria-dispar). Archives of Insect Biochemistry and Physiology, 24, 139–147. doi: https://doi.org/10.1002/arch.940240304
- Wu, A. H. (2015). Expression and activity analysis of engineering antibodies against Bt Cry2Aa toxin (MS thesis). Nanjing Agricultural University, Nanjing.