Abstract
Analysis of biogeographic affinities is a key tool to establish and improve the resolution of hierarchical biogeographic systems. We describe patterns of species richness of the marine macroalgal flora across Lusitanian Macaronesia (Azores, Madeira, the Salvage Islands and the Canary Islands), and test (i) whether such differences are related to differences in proximity to the nearest continental shore and size among islands. We also explore biogeographic affinities in the composition of macroalgal assemblages (= presence/absence of each taxon in multivariate datasets) to determine (ii) whether each archipelago is a biogeographic unit within this ecoregion and (iii) whether patterns in assemblage composition are related to proximity (i.e. distances) among islands. Presence/absence matrices were created to test and visualize multivariate affinities among archipelagos. A total of 872 taxa were compiled. Species richness peaked at the Canary Islands and decreased towards the Azores; the pattern matched a progressive increase in distance from the nearest continental shores, matching the classical island biogeography theory. Intra-archipelago differences in species richness were largely related to variations in island size. Biogeographic similarities among archipelagos were hierarchically structured. Madeira and the Salvage Islands constituted one biogeographic unit. Floras from the Azores, Madeira and the Salvage Islands were barely separable from each other, but were different from those at the Canary Islands. Such biogeographic similarities among islands were negatively correlated with the geographical separation (i.e. distances) among them. Proximity to nearby continental shores, in conjunction with large- and meso-scale oceanographic patterns, seems to interact to create patterns in richness and composition of algal assemblages across Lusitanian Macaronesia.
Introduction
Understanding broad-scale patterns in the composition and diversity of communities is important for the theoretical development of macroecology–the interface between ecology and biogeography–and for the implementation of conservation and management policies (Brown, Citation1995).
Biogeographic units are defined using a hierarchical system based on taxonomic configurations, and analyses of biogeographic affinities are a key tool to establish such a system (Spalding et al., Citation2007). Normally, significant levels of internal biogeographic heterogeneity exist within any particular biogeographic level. A more detailed analysis is thus necessary to improve the resolution of our biogeographic systems. Different factors influence the evolutionary history of biota at any given biogeographic level. Dispersion of species to insular systems is strongly affected by two key factors: (i) proximity (i.e. distance) to donor areas, normally the nearest continental shore, but sometimes other islands, and (ii) the size of the islands, which determine the amount of available habitat. These ideas are the core of the ‘Island Biogeography Theory’ (hereafter IBT, MacArthur & Wilson, Citation1967), which specifically predicts that proximity to adjacent continental shores influences colonization rates, while the island size affects extinction rates. Moreover, patterns of ocean circulation (e.g. currents and other oceanographic elements such as upwellings), particularly for marine organisms, can also influence the composition of the biota (Whittaker & Fernández-Palacios, Citation2007). Importantly, dispersal abilities of each species ultimately influence the pool of organisms reaching any oceanic island. For example, species with considerable dispersal capabilities, and thus large range sizes (Lester et al., Citation2007), are typically found on oceanic islands far away from continental masses (Whittaker & Fernández-Palacios, Citation2007), and this pattern has been highlighted for marine algae (van den Hoek, Citation1987).
The oceanic archipelagos located in the central-east (northern hemisphere) Atlantic Ocean are grouped under the term Macaronesia. Five archipelagos are included (from north to south, ): Azores, Madeira, Salvage Islands, Canary Islands, and Cape Verde. These archipelagos share similar geomorphological traits; they are all volcanic in origin, and although there is no general consensus, most are thought to be the result of several geologic hotspots (Carracedo, Citation2001). They have a unique biogeography: all native flora and fauna reached the archipelagos via long-distance dispersal from adjacent continental areas (Whittaker & Fernández-Palacios, Citation2007) or, in some circumstances, adjacent archipelagos (Domingues et al., Citation2008). Marine biotas of the different archipelagos are interconnected via oceanic currents; surface currents generally (but not always) move from the Azores towards Madeira and the Canary Islands (Morton et al., Citation1998). The Azores, Madeira, the Salvage and Canary Islands belong to the same ecoregion within the so-called Lusitanian province (Spalding et al., Citation2007). The archipelago of Cape Verde is under the influence of a more tropical climate (e.g. coral reefs are found in contrast to the other archipelagos) and is included within the West African Transition province (Spalding et al., Citation2007).
Fig. 1. Map of Lusitanian Macaronesia showing the location of each archipelago in the eastern Atlantic Ocean. Cape Verde not shown.
The macaronesian archipelagos, and their islands, have differing degrees of isolation from the nearest continental shores. Such differences have been advocated to explain differences in the composition and diversity of the marine biota (Hawkins et al., Citation2000). Previous phytogeographic studies have highlighted a high degree of similarity in the composition of the marine flora at each archipelago and the nearest (donor) continental coasts. For example, the marine flora of the Azores presents elements in common with the northern Atlantic, the western Mediterranean, and the eastern American coasts (Prud’homme van Reine, Citation1988), while the Canary Islands show stronger affinities with the western Mediterranean and the tropical western Atlantic (van den Hoek, Citation1987). However, quantitative analyses exploring the affinities in the composition of the marine flora among archipelagos are rather sparse. In this study, we described patterns in species richness of the macroalgal flora across Lusitanian Macaronesia and tested (i) whether such differences are related to differences in proximity (= distance) to the nearest continent and/or size among islands. We also analytically explored biogeographic affinities in the composition of algal assemblages (= presence/absence of each taxa in multivariate datasets) to determine; (ii) whether each archipelago is a biogeographic unit within this ecoregion and (iii) whether patterns in assemblage composition are related to proximity among islands.
Materials and methods
Presence/absence matrices for red, green and brown algae, and the entire algal dataset as well, were created (Appendix 1, see supplemental material available online at www.informaworld.com/mpp/uploads/tuya___haroun_appendix_1.xls) by searching in literature for all available phytogeographic records across Lusitanian Macaronesia. These matrices take into account published checklists and subsequent additions, for the different islands that constitute each archipelago (Appendix 2, available online at www.informaworld.com/mpp/uploads/tuya___haroun_appendix_2.doc).
Patterns in species richness
We calculated the total number of taxa of red, green, and brown algae in each archipelago, and the number of shared and unshared taxa among the archipelagos. Multiple regression models tested whether proximity (= distance) to the nearest continent (linear distance in km) and island size (the coastal perimeter of each island was used as a proxy) influenced species richness across the study region. To visualize patterns in species richness separately for each of these two variables, simple regression models tested the relationship between species richness at each island and proximity to the nearest continent, and the relationship between species richness and the coastal perimeter of each island separately for each archipelago; so we were able to explore the effects of within-archipelago differences in island sizes on species richness patterns. An exponential model was selected for the former, and a linear for the latter.
Patterns in assemblage composition
Canonical Analysis of Principal coordinates (CAP, Anderson & Willis, Citation2003) was used as a constrained ordination procedure to test and visualize differences in assemblage composition among archipelagos. CAP is an appropriate technique to analyse biogeographic affinities in assemblage composition (Edgar et al., Citation2004). Essentially, CAP finds axes in multivariate space to maximize the separation among groups (archipelagos in this case). First, we conducted a Principal Coordinates Analysis (PCO); we then carried out the CAP based on that subset of the PCO axes for which additional PCO axes did not add explanatory power. Analyses were based on Jaccard dissimilarities, which are ideal for incidence data (presence/absence data, Clarke & Warwick, Citation2001). The ‘trace statistic’ was used to test (via 999 permutations of the data) differences in assemblage composition among archipelagos. The CAP routine also calculated miscalculation errors for regional groupings using the ‘Leave-one-out Allocation success’ (LoA): each island was removed from the dataset, the CAP analysis re-run using the remaining observations, and then the removed datapoint classified to the nearest group centroid in the canonical space. Comparison of known with allocated groups provided miscalculation errors (Anderson & Willis, Citation2003). Pairwise comparisons in assemblage composition between each pair of archipelagos were carried out with 1-way ANOSIM, using 999 permutations, via calculation of the R statistic. This statistic fluctuates between −1 and 1 (R equals 0 if the null hypothesis is true, while values significantly different from 0 indicate some degree of discrimination). Interpretations of pairwise R-statistic values followed Clarke (Citation1993).
Distance-based redundancy analysis (db-RDA, Legendre & Anderson, Citation1999) was used to visualize whether variation in distance to the nearest continental shore and island perimeter explained variation in assemblage composition across the study region. Multivariate multiple regression, using the DISTLM routine via 999 permutations of the data (Anderson, Citation2001), tested the significance of these relationships by fitting a linear model based on Jaccard dissimilarities. We additionally determined whether geographical affinities in the composition of algal assemblages across the study region were related to the geographical separation by means of a correlation analysis between the pairwise dissimilarity matrix between each pair of islands and a pairwise matrix containing the linear distances (in km) between each pair of islands. Rho values were calculated as Spearman rank correlations, and p-values using 999 permutations, through the RELATE routine.
All multivariate procedures were carried out by means of the PRIMER 6.0 package (Clarke & Warwick, Citation2001).
Results
Patterns in species richness
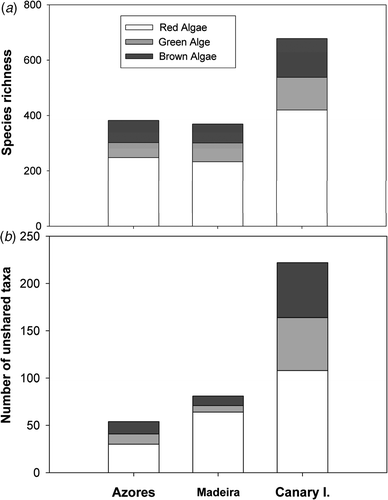
A total of 872 taxa were compiled across the study region (552 red algae, 143 green algae, and 177 brown algae, Appendix 1, see supplemental material). Overall, the number of taxa of each algal group peaked at the Canary Islands (678), and decreased towards the north of the study region (), though the Azores (382) hosted a slightly larger number of taxa relative to Madeira (369). Only 28 taxa (3.21%) were found in all archipelagos, whereas 366 taxa (41.97%) were exclusively restricted to one archipelago. The Canary Islands also hosted the greatest number of taxa exclusive of any other archipelago (a total of 222 species), which represents around one-third of its algal flora (). This proportion was smaller for Madeira (∼22%), and particularly low for the Azores (∼14%) ().
Fig. 2. (a) Algal richness and (b) number of taxa restricted to one archipelago across the Lusitanian Macaronesia. Data from Madeira and the Salvage Islands were pooled.
Species richness was significantly affected by both ‘distance’ (to the nearest continental shore) and (island) ‘perimeter’, but ‘distance’ explained a larger amount of variation (). Species richness decreased exponentially with increasing distance from adjacent continental shores (, p < 0.0001 in all cases). At the archipelago level, species richness linearly increased with increasing insular coastal perimeter (, p < 0.05 in all cases), although the Azores showed lower R 2 values in all cases (except green algae).
Fig. 3. Relationship between species richness at each island and distance to the nearest continental shore for (a) the overall algal dataset, (b) red algae, (c), green algae, and (d) brown algae. p-values provide goodness-of-fit tests to examine the significance of the decay in species richness with increasing distance from the continent. Data from Madeira and the Salvage Islands were pooled (see results). ×: Azores, ▴: Madeira and Salvage Islands, ▪: Canary Islands.
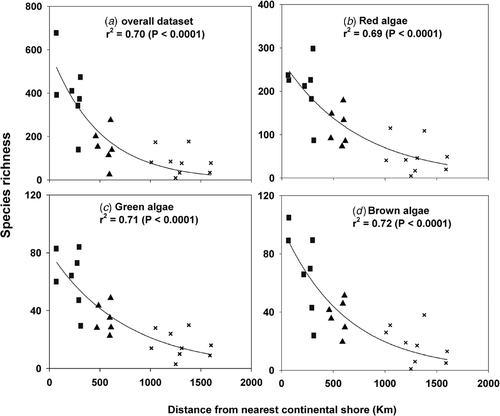
Fig. 4. Relationship between species richness at each island and its corresponding coastal perimeter for (a) the overall algal dataset, (b) red algae, (c), green algae, and (d) brown algae, separately for each archipelago. p-values provide goodness-of-fit tests to examine the significance of the increase in species richness with increasing island perimeter. Data from Madeira and the Salvage Islands were pooled (see results). ×: Azores, ▴: Madeira and Salvage Islands, ▪: Canary Islands.
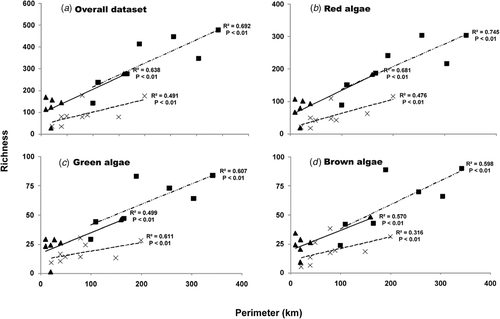
Table 1. Results of multiple regressions testing of the effect of ‘distance’ (to the nearest continental shore) and (island) ‘perimeter’ on species richness across the study region.
Patterns in assemblage composition
The CAP analyses () efficiently separated floras from the different archipelagos along the first canonical axis (δ1 2 was, in all cases, significant, p < 0.01). Biogeographic similarities, however, were hierarchically structured. First, Madeira and the Salvage Islands constituted the same biogeographic unit (, one-way ANOSIM: R < 0.0, p > 0.8 in all cases); their algal assemblages clustered together in the ordination space regardless of the algal group (), and the LoA indicated an important misclassification for the Salvage Islands; they were repeatedly misclassified as belonging to Madeira. Second, floras from the Azores, Madeira and the Salvage Islands were barely separable (, one-way ANOSIM: R < 0.4 in all cases), but were different from those found on the Canary Islands (, one-way ANOSIM: R > 0.5 in all cases). In addition, none of the Canary Islands were misclassified as belonging to the other three archipelagos for any of the algal groups.
Fig. 5. Constrained canonical ordination plots (CAP) of assemblage composition for (a) the overall algal dataset, (b) red algae, (c), green algae, and (d) brown algae. ×: Azores, ▾: Madeira, ▴: Salvage Islands, ▪: Canary Islands. Analyses were based on 8, 7, 7 and 6 PCO axes, respectively. ‘Leave-one-out Allocation’ success is indicated (=LoA, i.e. the percentage of points correctly allocated into each group). δ2: square canonical correlation with CAP-I.
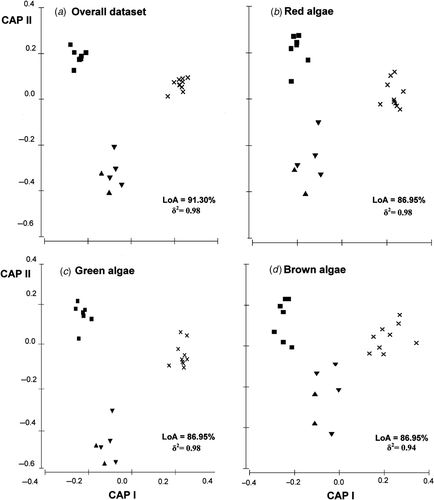
Table 2. One-way ANOSIM testing differences in assemblage composition between each pair of archipelagos.
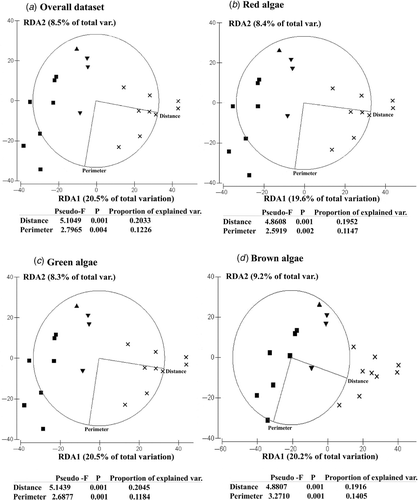
The first two axes from the db-RDA explained, in all cases, ∼30% of the total variability in algal composition. In the biplots of the first two db-RDA axes (), ‘distance’ (to the nearest continental shore) was positively correlated with the first axis (RDA-1). ‘Distance’ was the best predictor of variation in assemblage composition across the study region: all multivariate multiple regressions indicated that ‘distance’ explained ∼20% total variability (inserted Tables in ). The second axis of all db-RDAs was negatively correlated with the (island) ‘perimeter’, contributing significantly to an additional 11–14% of total variability (inserted tables in ). These results indicated that, although variation in coastal perimeter explained a considerable proportion of the variation in assemblage composition, ‘distance’ to the nearest continental shore was the main factor explaining patterns of variation in floral composition across Lusitanian Macaronesia. In fact, the similarities in assemblage composition between each pair of islands were negatively correlated with the geographical separation between them (overall dataset: Rho = 0.585, red algae: Rho = 0.575, green algae: Rho = 0.572, brown algae: Rho = 0.524; p ≤ 0.01 in all cases).
Fig. 6. Distance-based Redundancy Analysis (db-RDA) biplots of first and second axes relating ‘distance’ (to the nearest continental shore) and (island) ‘perimeter’ to algal composition across the study region. ×: Azores, ▾: Madeira, ▴: Salvage Islands, ▪: Canary Islands. Results of multivariate multiple regressions testing the relationship between ‘distance’ and ‘perimeter’ and the composition algal assemblages are indicated. Analyses were based on Jaccard similarities. To retain variables with explanatory power, the AIC procedure was chosen as model selection criterion (Anderson, Citation2001). Abbreviations: Var.: variation.
Discussion
This study has documented that inter-archipelago variability in species richness and algal composition is explained by differences in proximity to the nearest continental shores, while intra-archipelago differences were considerably affected by the size (i.e. the perimeter) of each island.
As predicted by the IBT (MacArthur & Wilson, Citation1967), the richness of algal species at oceanic islands decreased with increasing distance from the nearest continental shore and so the Canary Islands were floristically richer than the Azores and Madeira. In turn, quasi-permanent filaments advected westward away from the north-eastern African coasts are common (Davenport et al., Citation2002), thus allowing a considerable degree of connectivity between the Canary Islands and the adjacent continental coasts, likely sustaining high immigration rates to this archipelago (Whittaker & Fernández-Palacios, Citation2007). On the above basis, we would anticipate that one would observe greater algal diversity at Madeira than the Azores; the absence of such a pattern is likely to be due to the larger coastal area of the Azores (i.e. the coastal perimeter of the Azores is more than three times larger than that of Madeira), and so extinction rates have probably been higher around Madeira. Our study showed that most algae on the Azores (∼86%) may also be found on any of the other macaronesian archipelagos. This suggests that most algae on the Azores have large range sizes and so probably have large dispersion patterns (van den Hoek, Citation1987).
Several oceanographic processes can also be superimposed on differences in proximity to continental shores among archipelagos to create and maintain patterns in richness and composition of algae across Lusitanian Macaronesia. Firstly, sea water temperature is typically colder in the Salvage Islands (16–24°C), Madeira (16–24°C) and Azores (12–23°C) than in the Canary Islands (17–25°C) and so the presence of species with tropical affinities in the former archipelagos can be limited compared with the latter (e.g. fishes, Brito et al., Citation2007). It has been reported that tropical biota have typically narrower geographical ranges than temperate biota–Rappoport's rule–(Stevens, Citation1989). For example, hotspots in algal diversity are typically characterized by the existence of numerous algae with restricted geographical ranges (Santelices & Marquet, Citation1998). Secondly, the Canarian Archipelago lies in the transition between the northwest African upwelling and the oligotrophic open ocean (the so-called Northwest African Coastal Transition Zone). Considerable spatial variation in sea surface temperature occurs across an east–west gradient perpendicular to the African coast, with an average difference of 2°C between the eastern and western islands (Davenport et al., Citation2002). As a result of this meso-scale variability, marine assemblages at widely separated islands of this archipelago (100 s of km) are subjected to different oceanographic conditions, that produce differences in the composition (Haroun et al., Citation2003) and structure (Tuya & Haroun Citation2006) of macroalgae between the eastern and western islands. The eastern islands predominantly host taxa with warm-temperate affinities, while the western islands have a flora with more tropical affinities (Haroun et al., Citation2003). Such oceanographic gradients, on the other hand, are absent around the other macaronesian archipelagos, preventing the substantial environmental heterogeneity normally associated with higher richness and diversity (Brown, Citation1995; Whittaker & Fernández-Palacios, Citation2007).
It is also important to realize that the amount of research effort across the four archipelagos has not been equal. The Canaries and the Azores host research groups that have studied the phycology of both archipelagos for >25 years ago. For example, the number of published articles containing the keyword ‘algae’ () is largest for the Canary Islands (76), followed by the Azores (26) and then Madeira (11). This has resulted in the largest number of taxonomic (including revisions and additions) studies (; Appendix 2, see supplemental material: www.informaworld.com/mpp/uploads/tuya___haroun_appendix_2.doc) for the Canary Islands (29), followed by the Azores (18), Madeira (8) and to date only four publications on the algal flora of the Salvage Islands. For this reason, we can not rule out that algal species richness could be underestimated in Madeira and the remote Salvage Islands. However, Madeira and the Salvage Islands have considerably lower total coastal perimeters compared to the Azores and particularly when compared to the Canary Islands. If one considers the ratio between the amount of research effort (e.g. the number of works as a proxy, ) and the total coastal perimeter of each archipelago, the values tend to be comparable (i.e. same order of magnitude), minimizing differences in research effort among archipelagos as an important confounding factor of our interpretations.
Table 3. Number of published articles containing the keywords ‘algae’ and the name of each archipelago (the search was performed thorough the ISI Web of Science) and number of taxonomic works (including revisions and additions) carried out at each archipelago (see online supplemental data: Appendix 2). The ratio between the number of articles, and taxonomic works, per archipelago, and the total archipelago coastal perimeter is also indicated.
On the basis of congruent divisions for algal assemblages, Lusitanian Macaronesia can be separated into two major biogeographical areas: (i) the Canary Islands and (ii) Azores, Madeira and the Salvage Islands. The latter can be further subdivided into two areas: (iia) Azores and (iib) Madeira and the Salvage Islands. Importantly, this outcome was consistent across the different algal groups (red, green and brown algae), which reinforces this criterion as a potential valid model for Lusitanian Macaronesia. In general, these biogeographic affinities among the studied archipelagos were inversely related to their geographical separation. However, the oceanographic peculiarities of the study region were also important in shaping such similarities. The Salvage Islands are a group of islets approximately midway between Madeira and the Canary Islands. One might anticipate that the algal floras would be a mixture of both the Madeira and the Canaries' flora. However, the algal composition on the Salvage Islands was very similar to that observed on Madeira. The main oceanographic feature of the study region is the Canary current, which flows towards the south-west. Thus, it is likely that the Salvage Islands are the main recipient, through passive dispersal, of those floral elements present on Madeira. This connectivity has resulted in a high degree of similarity between marine floras on Madeira and the Salvage Islands. It is appreciated that further data from the Salvage Islands is required unambiguously to clarify the appropriateness of this hypothesis. The large distances between the continental shores and the Azores, Madeira and the Salvage Islands (>400 km), in addition to their similar oceanographic contexts, could help to explain why algal floras of these three archipelagos are similar.
In summary, proximity to adjacent continental shores, in conjunction with large and meso-scale oceanographic patterns, seem to interact primarily to create patterns in richness and composition of algal assemblages across Lusitanian Macaronesia. Moreover, differences in island sizes affect intra-archipelago variations in both species richness and composition of algae.
Supplementary Data
Download Zip (49.8 KB)Acknowledgements
FT was supported by a postdoctoral research fellowship within the framework ‘Plan de Formación y Perfeccionamiento’ of the Spanish Ministry of Education and Sciences. We thank T. Wernberg for providing positive comments on a draft of the manuscript and two anonymous reviewers for their help in improving this paper.
Related Research Data
References
- Anderson , MJ . 2001 . Permutation tests for univariate and multivariate analysis of variance and regression . Can. J. Fish. Aquat. Sci. , 58 : 626 – 639 .
- Anderson , MJ and Willis , TJ . 2003 . Canonical analysis of principal coordinates: a useful method of constrained ordination for ecology . Ecology , 84 : 511 – 525 .
- Brito , A , Falcón , JM and Herrera , R . 2007 . Características zoogeográficas de la ictiofauna litoral de las islas de Cabo Verde y comparación con los archipiélagos macaronésicos . Rev. Acad. Canar. Cienc. , 18 : 93 – 109 .
- Brown , JH . 1995 . Macroecology , Chicago (IL), , USA : University of Chicago Press .
- Carracedo , JC . 2001 . “ Volcanismo reciente y riesgo volcánico ” . In Naturaleza de las Islas Canarias. Ecología y conservación , Edited by: Fernández-Palacios , JM and Martín , J . 65 – 76 . Santa Cruz de Tenerife, , Canary Islands : Turquesa .
- Clarke , KR . 1993 . Non-parametric multivariate analyses of changes in community structure . Aust. J. Ecol. , 18 : 117 – 143 .
- Clarke , KR and Warwick , RM . 2001 . Changes in Marine Communities: an Approach to Statistical Analysis and Interpretation , Plymouth, , UK : PRIMER-E .
- Davenport , R , Never , S , Helmke , P , Perez-Moreno , J and Llinás , O . 2002 . Primary productivity in the northern Canary Islands region as inferred from Seawifs imagery . Deep Sea Res. II , 49 : 3481 – 3496 .
- Domingues , VS , Alexandrou , M , Almada , VC , Robertson , DR , Brito , A , Santos , RS and Bernardi , G . 2008 . Tropical fishes in a temperate sea: evolution of the wrasse Thalassoma pavo and the parrotfish Sparisoma cretense in the Mediterranean and the adjacent Macaronesian and Cape Verde Archipelagos . Mar. Biol. , 154 : 1432 – 1793 .
- Edgar , GJ , Banks , S , Fariña , JM , Calvopiña , M and Martínez , C . 2004 . Regional biogeography of shallow reef fish and macro-invertebrate communities in the Galapagos Archipelago . J. Biogeogr. , 31 : 1107 – 1124 .
- Haroun , RJ , Gil-Rodríguez , MC and Wildpret De La Torre , W . 2003 . Plantas Marinas de las Islas Canarias , Edited by: Canseco . Spain : Talavera de La Reina .
- Hawkins , SJ , Corte-Real , HBSM , Pannacciulli , FG and Weber , LC . 2000 . Thoughts on the ecology and evolution of the intertidal biota of the Azores and other Atlantic islands . Hydrobiologia , 440 : 3 – 17 .
- Legendre , P and Anderson , MJ . 1999 . Distance-based redundancy analysis: testing multispecies responses in multifactorial ecological experiments . Ecol. Monogr. , 69 : 1 – 24 .
- Lester , SE , Ruttenberg , BI , Gaines , SD and Kinlan , BP . 2007 . The relationship between dispersal ability and geographic range size . Ecology Letters , 10 : 745 – 758 .
- MacArthur , RH and Wilson , EO . 1967 . The Theory of Island Biogeography , Princeton (NJ), , USA : Princeton University Press .
- Morton , B , Britton , JC , De Frias , AM and Martins , H . 1998 . Coastal Ecology of the Azores , Ponta Delgada, , Acores, Portugal : Sociedade Afonso Chaves .
- Prud’homme Van Reine , WF . 1988 . Phytogeography of seaweeds of the Azores . Biogeography of Marine Benthic Algae , 42 : 165 – 185 .
- Santelices , B and Marquet , P . 1998 . Seaweeds, latitudinal diversity patterns, and Rappoport's rule . Divers. Distrib. , 4 : 71 – 75 .
- Spalding , MD , Fox , HE , Allen , GR , Davidson , N , Ferdana , ZA , Finlayson , M , Halpern , BS , Jorge , MA , Lombana , A , Lourie , SA , Martin , KD , McManus , E , Molnar , J , Recchia , CA and Robertson , J . 2007 . Marine ecoregions of the world: a bioregionalization of coastal and shelf areas . Bioscience , 57 : 573 – 583 .
- Stevens , GC . 1989 . The latitudinal gradients in geographical ranges: how so many species co-exist in the tropics? . Am. Natur. , 133 : 240 – 256 .
- Tuya , F and Haroun , RJ . 2006 . Spatial patterns and response to wave exposure of photophilic algal assemblages across the Canarian Archipelago: a multiscaled approach . Mar. Ecol. Prog. Ser. , 311 : 15 – 28 .
- Van Den Hoek , C . 1987 . The possible significance of long-range dispersal for the biogeography of seaweeds . Helgol. Mar. Res. , 41 : 261 – 272 .
- Whittaker , RJ and Fernández-Palacios , JM . 2007 . Island Biogeography. Ecology, Evolution and Conservation , London, , UK : Oxford University Press .