Abstract
Complex bristle types formed by species in the genus Mallomonas include those with helmet or lance-shaped apices. The ornamentation on each side of the helmet has been thought to be equivalent or symmetrical, whereas on a lance-shaped bristle an expanded portion folds over one side of the shaft to form an asymmetrical structure. We describe, for the first time, helmet bristles with a distinctly asymmetrical design, also formed by the folding of a siliceous membrane over one side of the helmet. We postulate that the asymmetrical helmet represents a structure that combines the formation of a symmetrical helmet and a lance-shaped design on the same bristle. Further, we report structurally similar asymmetrical helmet bristles, lance-shaped bristles and scales that are unambiguously assigned to Mallomonas asmundiae in Middle Eocene sediments from a maar lake in northern Canada, supporting the hypothesis that scale and bristle morphology in the Synurophyceae has undergone extensive prolonged evolutionary stasis. Given differences in scale morphology and the presence of asymmetrical helmet bristles, we transfer the North American endemic Mallomonas acaroides var. muskokana to the rank of species. Further, we formally describe Mallomonas dispar and M. lancea, fossil species with asymmetrical helmet bristles and lance-shaped bristles, respectively. The taxonomic and biogeographic significance of asymmetrical and lance-bearing bristles is discussed.
Introduction
Mallomonas Perty, the largest genus within the class Synurophyceae, consists of motile unicellular organisms with one or two plastids, two apically inserted flagella and a siliceous cell covering composed of scales and bristles. Scales are flat, plate-like structures that are precisely arranged around the cell membrane and have species-specific structures (Siver, Citation1991 a). Bristles, found on all but a few species, are elongate structures consisting of a shaft and an expanded proximal end termed the foot (Siver, Citation1991 a). The foot is tucked under the distal end of a scale and the shaft radiates out from the cell. Although bristles also have species-specific characteristics, historically they have been utilized less than scales for determining taxa at the species level. There are more than 172 known species and subspecific taxa of Mallomonas (Kristiansen, Citation2002) and the genus is at least 40 Ma years old, with representatives having been uncovered from Middle Eocene lake sediments (Siver & Wolfe, Citation2005 a, Citation b ).
The shaft of the bristle is slender, most often curved, circular or ribbed in cross section, and sometimes serrated (Siver, Citation1991 a). The shaft may be solid or hollow, the latter type formed by a folding or rolling of the silica sheet such that a slit occurs along the axis where the margins converge. Many species possess bristles with a single serration, most often along the convex margin or ridge, however some have two or three rows of teeth. The teeth are usually pointed, sometimes recurved, and in a few cases are bifurcate or trifurcate.
The distal tip of the bristle also varies greatly between species and can be of taxonomic significance. In many species the tip is drawn out into a sharp point, however, other designs include forked, hooked, plume, paddle, lance and helmet apices (Siver, Citation1991 a; Kristiansen, Citation2002). The lance and helmet forms, two of the more complex morphologies, are of particular interest in our current work. In the case of the lance bristle, one of the margins near the apex becomes expanded, folded and attached to another ridge, forming a triangular-shaped tip referred to as the lance. Because the fold is unidirectional, it results in an asymmetrical form. Helmet tips are formed when the ventral margin of the shaft becomes expanded and moulded into a C-shaped design. Because the expanded portion of the shaft is not folded, the design of the helmet is identical on each side producing a symmetrical structure.
Mallomonas acaroides var. muskokana Nicholls and M. asmundiae (Wujek & Van der Veer) Nicholls produce helmet and lance-tipped bristles, respectively (Kristiansen, Citation2002). Mallomonas acaroides var. muskokana, originally described from the Muskoka District of Ontario, was distinguished from what was believed to be its closest relative, Mallomonas acaroides Perty emend. Iwanoff, on the basis of slight differences in scale and bristle structure, and because it has domeless scales (Nicholls, Citation1987 a). We have discovered that the design of M. acaroides var. muskokana helmet bristles is remarkably different and unique among all known living species of Mallomonas, and further that this bristle design dates to at least the Eocene. We have also uncovered scales and lance bristles from Eocene sediments that are remarkably similar to, if not identical with, those of M. asmundiae.
The current study is based on modern collections taken from lakes and ponds distributed from Connecticut to Newfoundland, which form part of a large-scale biotic survey and inventory of scaled chrysophytes and diatoms (Siver et al., Citation2005; Siver & Lott, Citation2006), and samples from an ancient North American Eocene maar lake situated near the Arctic Circle and known as Giraffe Pipe (Siver & Wolfe, Citation2005 a; Wolfe et al., Citation2006). The Early and Middle Eocene, including the time period when the Giraffe Pipe maar lake existed, represented a warm period in the Earth's history (Zachos et al., Citation2001; Pagani et al., Citation2005), referred to as the Cenozoic hot house, when subtropical environments were well established at high latitudes across North America (Greenwood & Basinger, Citation1994; Zachos et al., Citation2001; Pagani et al., Citation2005). During this time period, latitudinal temperature gradients were greatly reduced, the planetary cryosphere was vastly diminished or absent, and large expanses of the Arctic Ocean were considerably less saline (Pagani et al., Citation2005; Brinkhuis et al., Citation2006).
The objectives of this paper are to: (i) describe for the first time asymmetrical helmet bristles, discuss their potential evolutionary connection to lance-shaped bristles and report that both bristle types are at least as old as the Middle Eocene; (ii) report the discovery of siliceous remains from Eocene sediments of a taxon with affinities to the modern species, Mallomonas asmundiae; (iii) transfer M. acaroides var. muskokana to the species level and; (iv) formally describe Mallomonas dispar sp. nov. and M. lancea sp. nov., fossil North American species with asymmetrical helmet and lance bristles, respectively. The taxonomic significance of asymmetrical and lance bristles, the biogeography of species possessing these bristle types and further evidence for evolutionary stasis within scaled chrysophytes will also be discussed.
Materials and methods
Samples examined as part of this study can be divided into three groups. First, surface sediments and plankton tows from 31, 26 and 31 water-bodies situated along coastal Maine, Nova Scotia and Newfoundland were sampled during the summers of 2002, 2005 and 2006, respectively. These lakes form part of a much larger study that includes over 300 water-bodies situated along the east coast of North America (Siver et al., Citation2005; Lott & Siver, Citation2005; Siver & Lott, Citation2006). The second set consists of sediment and plankton samples from 56 lakes in the State of Connecticut (Canavan & Siver, Citation1994). The third set consists of sediment core samples from a Middle Eocene maar lake, referred to as Giraffe Pipe, which existed between 40 and 48 Ma (Siver & Wolfe, Citation2005 a, Citation b ). The Giraffe Pipe locality (64°44′N; 109°45′W) is a kimberlite diatreme that was infilled by a sequence of lacustrine and paludal sediments, and subsequently covered by Neogene glacial deposits (Siver & Wolfe, Citation2005 a, Citation b ; Wolfe et al., Citation2006). The core contains 113.1m of stratified organic sediment of Middle Eocene age, including 44.8 m of peaty material underlain by 68.3m of lacustrine facies. Fossil remains from eight lacustrine facies, corresponding to lake-water depths ranging from 0.55 m to 41.09 m were used in the current study ().
Table 1. Samples from the Giraffe Pipe core used in this study and containing Mallomonas asmundiae, M. dispar and M. lancea.
For all modern localities, plankton samples were taken with a 10 µm mesh net from the centre of each water-body. Approximately 1–2 ml of plankton sample was air dried onto heavy duty aluminium foil on the day of collection. Surface sediments from these lakes were oxidized with a sulphuric acid/potassium dichromate mixture, cleaned with distilled water and an aliquot dried onto heavy duty aluminium foil. Small chips (∼200 mg) of organic mudstone from each Giraffe Pipe sample were oxidized with 30% H2O2, centrifuged, rinsed and also dried onto aluminium foil. The aluminum foil samples were attached onto aluminium stubs with Apiezon® wax and coated with a gold and palladium mixture for one minute with a Polaron model E5100 sputter coater (Siver, Citation1987). For each fossil sample, small chips of mudstone were also mounted directly onto aluminium SEM stubs using carbon tape, coated with the same gold and palladium mixture and grounded with silver paint. All samples were observed with a Leo (Zeiss) 982 SEM.
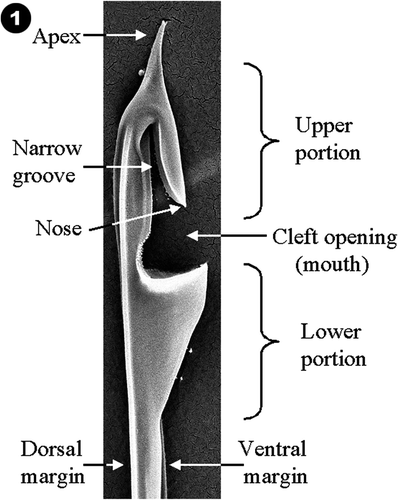
The following terminology will be used to describe the structure of a helmet bristle (). The expanded distal end of the bristle is known as the helmet. The helmet consists of upper and lower portions separated by a cleft-like opening referred to as the mouth. The side of the bristle containing the cleft-shaped opening is the ventral margin, and the side of the shaft that lacks the opening is the dorsal margin. If the bristle lies such that the cleft opening faces to the right with the apex pointed upwards, the side of the helmet that is visible is defined as the right side and the side that is not visible (lying on the surface) as the left side. The tip of the margin lining the upper part of the cleft is referred to as the nose.
Fig. 1. Labelled components of a helmet bristle illustrating the ventral surface, dorsal surface, cleft opening, lower portion, upper portion, mouth, nose, narrow groove and apex.
Results
Bristles
Asymmetrical helmet bristles in modern and extinct species of Mallomonas
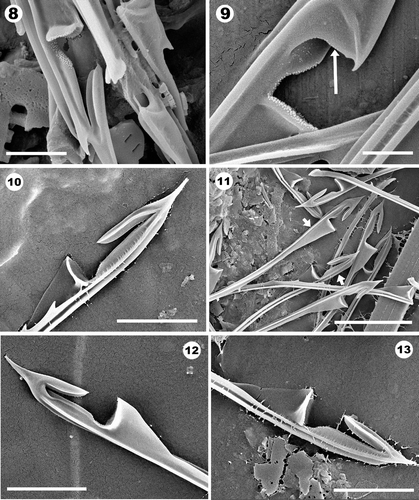
Helmet bristles on most modern species of Mallomonas, including M. acaroides (, ) and M. crassisquama (Asmund) Fott (, ), are symmetrical with respect to the structure on each side of the helmet. For these taxa, the C-shaped opening and the narrow grooves that run parallel with the shaft are identical on each side of the helmet. Bristles that displayed a distinctively different structure on each side of the helmet, referred to herein as asymmetrical helmet bristles, were consistently found on cells of the modern taxon M. acaroides var. muskokana (Figs 2–9) and on an undescribed, probably extinct, species of Mallomonas from the Giraffe Pipe core (Figs 25–30).
Figs 2–7. Helmet bristles from modern populations of Mallomonas muskokana. . Left-hand side of the helmet denoting the open nature of the cleft and groove. The rib on the left side (upper arrow in ) terminates as a subapical tooth (lower arrow). . Right-hand side of the helmet showing the closed nature of the narrow groove caused by folding of an expanded portion of the ridge. The sequence of three arrows in mark the ridge that folds (left arrow), the point where it folds (upper right arrow), and its termination at the nose (lower right arrow). Note that the dorsal-most ridge terminates in a second subapical tooth and the bristle apex ends in a tiny bifurcation. . Ventral view of a helmet bristle showing the flattened nature of the floor of the mouth and overall asymmetry of the helmet. Arrows denote the folded flap on the left side (lower arrow) and one of the subapical teeth (upper arrow). . Close-up of the cleft region lined with numerous tiny papillae. Scale bars are 2 µm () and 200 nm ().

Helmet bristles of M. acaroides var. muskokana are ribbed, serrated, range in length from 10 to 39 µm, and possess a helmet on the distal end of the shaft (Figs 2–9). There are two subapical teeth along the dorsal margin just above the main portion of the helmet, and the bristle terminates in a pointed bifurcate apex (Figs 2–5). The helmet forms along the ventral surface between two ribs. The bottom portion of the helmet that aligns the mouth is expanded, flattened and lined with minute papillae (Figs 6–9). On each side, the cleft-shaped opening narrows and forms a groove that bends distally and runs along the upper portion of the helmet (). The bristle is distinctly three-ribbed, with one rib running along each side of the shaft bordering the helmet, and a third along the dorsal margin. The dorsal rib continues just past the helmet and terminates as one of the two subapical teeth; this rib is never serrated. The left and right ribs, which line the sides of the helmet, are serrated and the morphology of the serration can differ on each rib. The rib on the left side can have teeth that are bifurcate or trifurcate, while the serration on the right rib is usually composed of simple teeth. The rib on the left side runs along the side of the helmet then bends slightly towards the dorsal surface and terminates as the second subapical tooth (arrows, ; top arrow, ). The rib on the right side expands, folds over the upper portion of the helmet covering the groove and terminates as a small point at the tip of the nose (arrows on , ). The groove on the left side of the cleft is not covered and remains open (). It is the folded portion enclosing the groove on the right side of the helmet that results in the asymmetrical design. The same type of asymmetrical helmet bristle found on M. acaroides var. muskokana has also been discovered on a fossil species from Giraffe Pipe sediments formally described below.
Figs 8–13. Helmet bristles from modern populations of Mallomonas muskokana (), M. crassisquama (, ) and M. acaroides (). . View of the left side of a bristle (on the left) and a ventral view (specimen on the right) denoting the asymmetrical nature of the helmet. . Close-up of the right side of a helmet showing the folded flap (arrow). . Left side of a helmet bristle. . Group of bristles depicting the open nature of both the right side (upper arrow) and left side (lower arrow) of the helmet. . Both sides of the helmet from a symmetrical bristle type. Scale bars: 500 nm (); 1 µm (); 2 µm (); 5 µm ().
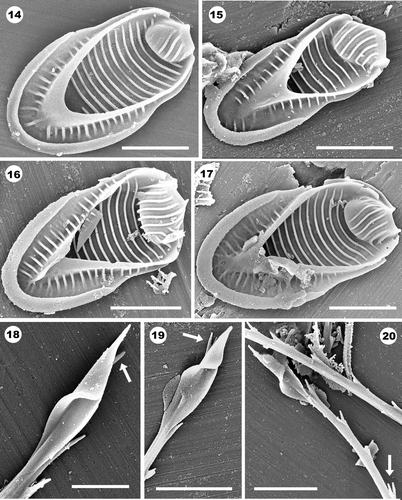
Evidence of taxa with lance-shaped bristles, including Mallomonas asmundiae, in Giraffe Pipe sediments
Numerous scales and bristles that closely match those of the extant species M. asmundiae were common in seven sections of the Giraffe Pipe core (). The morphology of the scales was virtually identical in all aspects with those of M. asmundiae, such that it was not possible to distinguish the fossil specimens from the extant congeners. As is true for modern specimens, the fossil scales possessed large and well developed domes, lateral incurvings, V-ribs that are continuous with the anterior submarginal ribs, parallel sets of ribs on the dome, shield and posterior flange, and ranged in size from 5.2–6.7 × 2.9–3.8 µm, within that given by Nicholls (Citation1987 b) for M. asmundiae (). Similarly, most characteristics of the bristles found in the Giraffe Pipe sediments matched those of modern populations of M. asmundiae. The bristles ranged in length from 15 to 31 µm, were strongly ribbed with a double (sometimes triple) row of teeth along the convex margin and terminated in either a sharp tooth or a lance-shaped apex (). In fact, the asymmetrical design of the lance-shaped tip on the fossil forms appears virtually identical to modern specimens. The only difference between the Giraffe Pipe bristles and those normally reported for M. asmundiae in modern collections was that the teeth aligning the shafts on the fossil specimens were more pronounced and longer than ones reported today (arrow, ). Bristles with similar lance-shaped apices as on M. asmundiae were additionally observed on another fossil species from Giraffe Pipe also described below.
Figs 14–20. Scale (Figs 14–17) and bristle (Figs 18–20) remains of fossil specimens of Mallomonas asmundiae from Giraffe Pipe. Note the large dome, broad V-rib, lateral incurvings and sets of parallel and evenly-spaced ribs on the shield, dome and posterior flange. The arms of the V-rib are continuous with the anterior submarginal ribs. Most of the ribs on the posterior flanges do not reach the margins. Scales depicted in represent the most common morphotypes, while the one in has a larger length-to-width ratio and more widely spaced shield ribs. . Lance-shaped bristle tips. One ridge expands and folds over onto another membrane forming the characteristic triangular-shaped apex. Arrows denote elongated subapical tooth. . Two bristles denoting the serrated shaft (upper bristle) and a lance-shaped apex. Arrow denotes the large double-toothed nature of the teeth. Scale bars: 1 µm (); 2 µm (Figs 14–17, 19–20).
Taxonomy
Transfer of Mallomonas acaroides var. muskokana to the species level
Mallomonas acaroides var. muskokana was originally described by Nicholls (Citation1987 a) based on populations from ten dilute, softwater lakes in Ontario, Canada. Nicholls (Citation1987 a) separated var. muskokana from the type on the basis of slight differences in the bristles, ribbing on the shield of the scales, ornamentation on the domes, and the fact that cells of var. muskokana had posterior domeless scales. He further noted that the closely related taxon, M. crassisquama, was distinguished from M. acaroides by the presence of posterior spined scales (Nicholls, Citation1987 a). Although the original description of M. acaroides var. muskokana was based solely on TEM, Siver (Citation1989; Citation1991a , Citation b ) later presented SEM micrographs of the siliceous cell covering along with a description of the stomatocyst. Even though the asymmetrical nature of the helmet bristles was overlooked in all of these works, this feature is clearly visible on micrographs in the original description (Nicholls, Citation1987 a, ) as well as ones illustrated subsequently in Siver (Citation1991 a, fig 3.44B) and Kristiansen (Citation2002, fig. 64d).
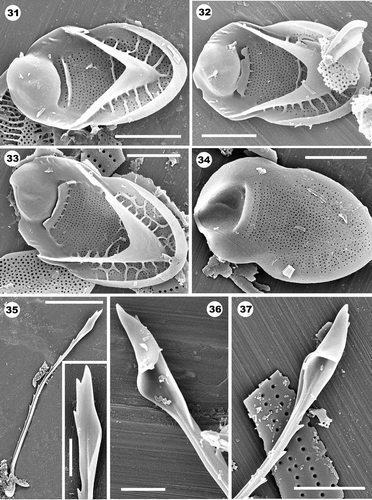
Figs 21–24. Domed and domeless scales of the fossil taxon Mallomonas dispar. . Note the large oval dome, shallow V-rib and single prominent transverse shield rib. Other than the transverse rib, the scale lacks additional secondary structures. . Distal end and anterior flanges of domeless scales lack base plate pores. Scale bars: 1 µm (); 2 µm ().

Figs 25–30. Helmet bristles of the fossil taxon Mallomonas dispar. . Group of bristles presumably from the same cell displaying the ribbed nature of the shaft. . Right-hand side of a helmet apex with a folded membrane covering the narrow groove. The arrows denote a subapical tooth along the left side of the structure. . Left-hand side of a helmet with an open, uncovered groove. . Right-hand side of a helmet apex. Note the expanded lower portion of the helmet with small transverse ribs (arrow on 30). Scale bars: 1 µm (); 2 µm ().

After further examination of scales and bristles, five primary features serve to separate M. acaroides var. muskokana from M. acaroides var. acaroides and support transfer to the rank of species. First, while all of the scales on M. acaroides var. acaroides cells possess domes, M. acaroides var. muskokana has both domed and domeless scales (Nicholls, Citation1987 a; Siver, Citation1989, Citation1991a ). Second, the domes on M. acaroides var. muskokana scales are usually ornamented with a series of ribs and papillae, whereas those of the type are not (Kristiansen, Citation2002). Third, the anterior submarginal ribs are continuous with the arms of the V-rib on M. acaroides var. muskokana (Siver, Citation1991 a). On M. acaroides var. acaroides scales the arms of the V-rib extend to the margin of the scale and are not continuous with the anterior submarginal ribs. Fourth, the shields of M. acaroides var. muskokana scales rarely have secondary structure, whereas those of M. acaroides var. acaroides are traversed by a series of ribs that often fuse to form a reticulated meshwork. Fifth, as noted in this work, the helmet bristles of M. acaroides var. muskokana are asymmetrical and fundamentally different in design to the symmetrical ones on M. acaroides var. acaroides.
The differences between M. acaroides var. muskokana and M. acaroides var. acaroides are sufficient to warrant recognition at the species level and the following combination is proposed:
Mallomonas muskokana (Nicholls) Siver nov. comb.
BASIONYM: Mallomonas acaroides var. muskokana Nicholls, Citation1987, Can. J. Bot., 65: 1779–1784, (holotype)–20.
Description of new species from Giraffe Pipe
Mallomonas dispar Siver, Lott & Wolfe nov. sp. (Figs 21–30)
LATIN DIAGNOSIS: Cellulae squamis cupuliferis et squamis cupula carentibus obtectae et setis apice galeiformi instructae. Lamina basalis squamae perforata, taenia reflexa tenui, et costa v-formi. Pori laminae basalis in scuto seriebus transversalibus, sed in limbo posteriore paulo fortuite dispositi. Cupula magna, rotunda ad subelliptica, costis paucis parallelibus. Squamae cupuliferae costa transversali manifesta in scuto prope basim cupulae. In squamis cupuliferis rami costae v-formis in limbos anteriores exeunt. In squamis sine cupula pori laminae basalis in parte distale squamae et in limbis anterioribus carentes. Setae manifeste costatae, apice galeiformi asymmetrico, ad apicem distalem acuminatae. Canalis apicis galeiformis tenuis et axi setae parallela. Canalis lateris sinistri aperta, lateris dextri velata. In latere dextro, una ad tres costae curtae e jugo ad partem inferiorem extensam apicis galeiformis.
TYPE: Here designated as (domed body scale).
TYPE LOCALITY: Sample marked “13-3-90” from Giraffe Pipe core. This section is 76.82 m down in the core and corresponds to a water-body with a maximum depth of approximately 7.24 m.
DIMENSIONS OF SCALES AND BRISTLES: domed scales 3.0–3.9 × 2.4–2.6 µm; domeless scales 2.9–3.3 × 2–2.2 µm; bristles 10–16 µm (n = 20). The epithet reflects the unequal nature of the helmet bristles.
Both domed and domeless scales and asymmetrical helmet bristles comprise the cell covering of M. dispar (Figs 21–30). Scales consist of a perforated base plate, shallow upturned proximal rim and a V-rib. The base plate pores are arranged in transverse rows on the shield, but are more randomly spaced on the posterior flange. The dome is large, circular to slightly elliptical and often with a few parallel aligned ribs (). All domed scales possess a single, prominent, transverse rib on the shield located directly behind the base of the dome (). Smaller ribs may connect the large transverse rib to the dome (). Other secondary structures are lacking. On domed scales that presumably surround the flagellar end of the cell, the arms of the V-rib fuse with relatively short anterior submarginal ribs that, in turn, fuse with the sides of the dome. On other domed scales the arms of the V-rib terminate on the anterior flange. Smaller domed scales can have slight lateral incurvings, but these are generally lacking on larger scales. Domeless scales lack base plate pores on the distal portion of the scale and along the anterior flanges ().
Bristles are strongly ribbed, possess an asymmetrical helmet and terminate with an elongated and pointed distal apex (). The opening or groove of the helmet is very narrow and is aligned almost parallel with the axis of the bristle (). The groove on the left side of the helmet remains open, but the groove on the right side is enclosed forming an asymmetrical design (). On the right side of the bristle, one to three short ribs extend from the ridge onto the lower expanded portion of the helmet (arrow, ). The rib aligning the left side of the helmet forms a subapical tooth (arrows, ). Abundant remains of M. dispar were found at the 11-3-90 and 13-3-90 sections of the Giraffe Pipe core.
Mallomonas lancea Siver, Lott & Wolfe nov. sp. ()
Figs 31–37. Scale () and bristle (Figs 35–37) remains of the fossil species, Mallomonas lancea from Giraffe Pipe sediments. . Scales with large, well-developed domes, prominent V-ribs, winged anterior flanges with parallel ribs, a single large transverse shield rib and widely-spaced ribs on the posterior flange. Base plate pores on the shield are small and in transverse rows. . Undersurface of a scale depicting a large dome cavity. . Ribbed bristle with a ventral serration and lance apex. Insert denotes a close up of the left side of the lance. . Close-ups of lance apices showing the triangular-shaped membrane folded over onto a ventral membrane. If the membrane along the ventral surface were to develop a cleft-like opening, it would begin to resemble an asymmetrical helmet bristle. See text for details. Scale bars: 1 µm (); 2 µm (); 5 µm ().
LATIN DIAGNOSIS: Squamae magnae, robustae, lateribus conspicue incurvatis, cupulis, costis v-formibus, et limbis anterioribus valde evolutis. Pori minuti in seriebus transversalibus in scuto, sed in limbo posteriore radialiter dispositi. Cupula magna, rotunda ad subelliptica, et levis. Costa una, manifesta, in scuto post basim cupulae disposita. Costae additionales interdum e costa v-formi in scutum radiantes and saepe costa secunda minor prope angulum costae v-formis dispositae. Limbus posticus sive serierum costarum rararum radiantum sive reticulo aperto instructus. Costa v-formis magna, ramis ad marginem squamae extendibus. Limbus anterior ala curta extendit, serie costarum parallelarum ornatus Setae manifeste costatae, serie dentium prope marginem ventralem et magnis, conspicuis apicelibus instructae. Dentes unidentati sive bifurcati.
TYPE: Here designated as .
TYPE LOCALITY: Sample marked “13-2-120” from Giraffe Pipe core. This section is 78.14 m down in the core and represents a waterbody with a maximum depth of approximately 8.56 m.
DIMENSIONS OF SCALES AND BRISTLES: scales 5.4–7.3 × 3.1–4.2 µm; bristles 15–20 µm (n = 20). The epithet reflects the lance-shaped nature of the bristle apices.
Scales are large and robust with prominent lateral incurvings, domes, V-ribs and well developed anterior flanges (). The shield is perforated with small pores arranged in transverse rows, whereas on the posterior flange the pores are more or less radially aligned. The dome is large, circular to slightly elliptical, and smooth (). A single, prominent, transverse rib is positioned on the shield directly behind the base of the dome. This rib often bends forward and comes to rest on the dome (). The posterior margin of the transverse rib often has a series of smaller ribs connecting it to the shield. Additional ribs may radiate from the V-rib onto the shield and often a second smaller transverse rib is observed near the base of the V-rib. The posterior flange consists of a series of widely spaced and radially aligned ribs (). On some scales the posterior flange ribs form a reticulated design (). The hood of the V-rib is well formed and the arms of the V-rib extend to the margin of the scale at the base of the anterior flange. The anterior flange extends as a short wing and is marked with a series of short parallel ribs ().
Bristles are strongly ribbed with a single row of teeth along the ventral margin and large, prominent lance-shaped apices (). The teeth can be single toothed or bifurcate (). At the apex, the ventral margin and the ridge along the right side of the bristle become expanded, folded and connect with another expanded portion along the ventral margin to form the lance (). With SEM, the folded membrane is not evident when viewed from the left-hand side (, insert).
Discussion
As illustrated, helmet bristles of Mallomonas species can clearly differ in regards to symmetry. On some species, such as M. crassisquama, M. acaroides and M. galeiformis Nicholls, the cleft-shaped openings on each side of the helmet are identical and neither side becomes occluded with a folded portion of the shaft. However, on M. muskokana one of the ribs expands along the distal part of the shaft and folds over the right side of the helmet covering the majority of the cleft. The rib on the left side of the shaft does not expand and fold, but rather continues past the helmet and terminates as a prominent subapical tooth along the dorsal side of the bristle. As a result, the cleft opening on the left side of the helmet remains uncovered, resulting in a distinctly asymmetrical design. What is more intriguing is the fact that the same asymmetrical pattern observed on M. muskokana bristles is also present on the Eocene fossil taxon, M. dispar.
In our opinion, the process where the rib expands and subsequently folds over the right side of the helmet on bristles of M. muskokana and M. dispar is similar, if not identical, to what occurs in the formation of lance-shaped bristles found on M. intermedia Kisselev emend. Péterfi & Momeu, M. corcontica (Kalina) Péterfi & Momeu, M. sphagnicola Nicholls, M. asmundiae (Kristiansen, Citation2002) and M. lancea (this paper). In each of these species, one of the ribs of the bristle expands and folds over an adjacent rib of the shaft forming a lance-like apex (Nicholls, Citation1987 b; Kristiansen, Citation2002). In the case of M. asmundiae and M. lancea, the membrane along the right ridge folds over and connects to another expanded portion positioned along the ventral margin. If the expanded membrane along the ventral surface were to develop a cleft it would resemble a helmet and essentially combine characteristics of lance and helmet bristles in a manner similar to that is observed in M. muskokana and M. dispar. Thus, instead of folding over an adjacent rib of the shaft, as occurs on a lance bristle, the flared rib on a M. muskokana bristle folds over and connects to the helmet. In this sense, the helmet bristles of M. muskokana possess characteristics of species within the Series Mallomonas (e.g. a helmet) as well as ones (e.g. the expanded and folded rib) found within the Series Intermediae Momeu & Péterfi and the Series Corconticae Asmund & Kristiansen (later emended by Nicholls, Citation1987 b). Based on the design of the bristle, M. muskokana may represent a taxon intermediate between species within the Series Mallomonas and the Series Intermediae and Corconticae. With further work, including analyses based on a full spectrum of morphological and molecular characters, it would not be surprising to find M. muskokana more closely aligned with taxa in the Series Intermediae and Corconticae, especially since these organisms are also similar in having domed and domeless scales and V-ribs that are continuous with the anterior submarginal ribs.
The symmetry of helmet bristles appears to be a very stable and consistent feature within populations of M. muskokana, M. acaroides, M. crassisquama and M. galeiformis. Siver (Citation1991 a) reported on populations of all four taxa based on seasonal collections made in 65 Connecticut lakes. Upon re-examination of the Connecticut material, as well as 46 populations from the 88 lakes in Maine, Nova Scotia and Newfoundland, all specimens of M. muskokana had asymmetrical helmet bristles, while the other three species possessed only symmetrical forms. Published micrographs of M. crassisquama confirm that this taxon has symmetrical helmet bristles with opened clefts on each side of helmet (e.g. Takahashi, Citation1978, figs 77, 78; Siver & Skogstad, Citation1988, ; Siver, Citation1991 a, fig 3.46E; Kristiansen, Citation2002, fig. 67d). Likewise, previously published micrographs of helmet bristles for M. acaroides (e.g. Nicholls, Citation1987 a, ) and M. galeiformis (Nicholls, Citation1988, ; Nicholls, Citation1990, ; Siver, Citation1988, figs 7, 8; Siver, Citation1991 a, figs 3.37E, F; Kristiansen, Citation2002, fig. 60d) are of the symmetrical type. We conclude that the symmetry of the bristle is a stable character, useful in distinguishing between Mallomonas species.
As we have already pointed out, there are sufficient differences in the scale and bristle morphologies between M. muskokana and M. acaroides to warrant separate species designations. Recognition of M. muskokana at the species level is further supported by the fact that M. acaroides shares key characteristics with M. crassisquama, but not with M. muskokana. First, the secondary design of M. crassisquama scales, especially on thinly ornamented specimens, can be similar to that of M. acaroides such that isolated scales cannot provide a definitive species identification (Kristiansen, Citation2002). Second, the arms of the V-rib on both M. crassisquama and M. acaroides scales extend to the scale margin and are not continuous with the anterior submarginal ribs (Siver et al., Citation1990). In addition, the helmet bristles of M. acaroides and M. crassisquama have a similar design and both lack the asymmetry found in M. muskokana. Lastly, the fact that cells of M. muskokana bear domed and domeless scales, like those of M. crassisquama, while cells of M. acaroides have only domed scales (Siver, Citation1991 a; Kristiansen, Citation2002) provides further evidence that M. muskokana needs to be recognized at the species level, and not as a subspecific taxon within M. acaroides.
Asmund & Kristiansen (Citation1986) placed M. asmundiae in the Series Striata under the Sectio Striatae. In that same publication, species with lance-shaped bristles were placed either in the Series Intermediae or the Series Corconticae, both under the large Sectio Mallomonas (Asmund & Kristiansen, Citation1986). Shortly thereafter, Nicholls (Citation1987 b) described M. sphagniphila Nicholls and reported finding M. asmundiae cells with lance-shaped bristles. Nicholls (Citation1987 b) further noted that the presence of lance-shaped bristles on M. asmundiae negated its inclusion in the Series Striata. Based largely on the presence of lance-shaped bristles, he emended the Series Corconticae and recommended inclusion of three species, M. asmundiae, M. corcontica and M. sphagniphila. Although Kristiansen (Citation2002) recognized the changes to the Series Corconticae made by Nicholls (Citation1987 b), he maintained M. asmundiae within the Series Striata. Our findings of M. asmundiae cells with lance-shaped bristles in Giraffe Pipe sediments clearly confirm the observations made by Nicholls (Citation1987 b). Thus, if the presence of lance-shaped bristles is given the taxonomic significance suggested by Nicholls (Citation1987 b) when he emended the Series Corconticae, M. asmundiae should be transferred into this series.
Regardless of the Series designation, isolated scales of M. asmundiae are similar and sometimes difficult to separate from those of M. sphagniphila, M. corcontica, M. cratis Harris & Bradley and M. pseudocratis Dürrschmidt. Accurate species identifications are paramount for understanding the current and historical biogeographies of these taxa and for determining endemic status (Kristiansen & Lind, Citation2004). Scales of M. asmundiae are similar to those of M. sphagniphila and M. corcontica, and all three taxa can have serrated bristles with either toothed or lance-shaped apices. Scales of M. asmundiae differ from those of M. sphagniphila and M. corcontica in possessing a larger number of parallel-aligned ribs on the dome that are orientated with, or at a slight angle to, the longitudinal axis of the scale. In addition, on M. sphagniphila and M. corcontica scales the arms of the V-rib extend to the margin and are not continuous with the anterior submarginal ribs as they are on M. asmundiae. The posterior flange ribs on M. asmundiae scales are less developed than those of M. sphagniphila and often do not extend to the posterior margin. Scales of M. corcontica also differ in regards to the design of the anterior flange and they contain a series of short ribs on the shield at the base of the dome that run parallel with the longitudinal axis of the scale. Compared with M. asmundiae, scales of M. pseudocratis lack distinct lateral incurvings and have roughly half the number of ribs (ca. 16 vs 30) on the posterior flange (Kristiansen, Citation2002). Scales of M. cratis have distinctly U-shaped dome ribs, ribs on the anterior flanges and lack well developed anterior submarginal ribs (Siver, Citation1991 a). Based on a comparison of all features, the fossil scales from the Giraffe Pipe core definitively match those of M. asmundiae and support the hypothesis of Siver and Wolfe (Citation2005 a) that the designs of the siliceous cell covering on multiple Mallomonas species have undergone prolonged evolutionary stasis.
Although the suite of morphological characteristics representing M. dispar is unique, this taxon is similar to the modern taxon M. galeiformis. Mallomonas galeiformis has both domed and domeless scales of similar size and shape (Nicholls, Citation1988; Siver, Citation1988). Most scales of M. galeiformis have a series of well developed transverse shield ribs, however, it is not uncommon to find scales with only a single prominent rib situated near the base of the dome, which are very similar to those of M. dispar (e.g. Siver, Citation1991 a, figs 3.36, 3.37). Both species also have helmet bristles with deep, narrow and similarly shaped grooves, however, those of M. dispar differ in being asymmetrical. In addition, cells of M. galeiformis generally have posterior scales with short spines, a scale type not uncovered to date in the Giraffe Pipe remains containing M. dispar. Despite differences, these two species are undoubtedly closely related and it is possible that M. dispar, or a similar species, was the precursor for the North American endemic, M. galeiformis.
Scales of M. dispar also share some features with the European endemic, M. intermedia. Both taxa have dome scales that bear a single prominent transverse shield rib near the dome and domeless scales with a flat smooth distal area in place of the dome. Domed scales of M. intermedia are larger with more developed anterior flanges and this taxon has lance-shaped bristles instead of ones with asymmetrical helmets. However, if as we suggest, the folded portion on asymmetrical helmet bristles is homologous with that on lance bristles, then it is highly likely that M. dispar and M. intermedia share a common ancestor.
In the descriptions of M. dispar and M. lancea we have linked specific bristle types with each scale morphology. We are confident of these assignments based on three lines of evidence. First, for each species, we examined intact sediment flakes and found the bristles in question next to the appropriate scale type within the mudstone matrix, presumably representing the remains of whole cells. Second, each scale/bristle combination was observed in at least one sample that lacked other, potentially confounding, scale types. Third, appropriate ratios of domed scales to bristles were observed for each taxon in each sample in which they were encountered.
Modern taxonomic works on Mallomonas (e.g. Takahashi, Citation1978; Wee, Citation1982; Siver, Citation1991 a; Citation2003; Kristiansen, Citation2002) are based largely on the morphological design of the siliceous scales and less on the structure of the bristles. However, as observed in this study, the fine detail of the bristles on M. muskokana and M. dispar relative to other species in the genus indicates that closer examination and comparison of bristle structure between species is warranted. In addition to providing another valuable character to distinguish subgeneric taxa, bristle morphology may help elucidate evolutionary shifts and biogeographic patterns within the genus. For example, it is intriguing that half the taxa examined or reviewed in this work with either helmet or lance bristles are reported to have restricted biogeographic ranges and, in fact, are considered to be endemic to specific regions of the world (Kristiansen & Lind, Citation2004). The helmet-bearing organisms, M. muskokana and M. galeiformis, are both endemic to North America and are distributed primarily along the eastern portion of the continent (Nicholls, Citation1987 a; Citation1988; Siver, Citation1988, Citation1991a ; Kristiansen, Citation2002). The lance bristle-bearing species M. sphagnicola is also endemic to North America (Kristiansen, Citation2002; Kristiansen & Lind, Citation2004), while M. intermedia is widespread, but endemic to Europe (Kristiansen, Citation2002; Kristiansen & Lind, Citation2004). Mallomonas corcontica, another lance-bearing species, has been reported only from Europe and North America, mostly from temperate and northern latitudes. On the other hand, M. crassisquama and M. acaroides, both with symmetrical helmet bristles, as well as M. asmundiae with lance bristles, are commonly reported and widely distributed. Thus, the formation of asymmetrical helmet or lance bristles, in conjunction with other morphological and molecular characters, may be a valuable means of tracking historical biogeographic patterns within the genus and for determining potentially endemic species within the genus.
In this regard, it is of considerable interest that both asymmetrical helmet bristles and lance-shaped bristles, among the most complex types of asymmetrical bristles known within the genus Mallomonas, have been uncovered in Middle Eocene Giraffe Pipe sediments. This finding supports the idea that these structures are ancient, at least 40 Ma old, and that, as has been found for scales (Siver & Wolfe, Citation2005 a, Citation b ), the structures of at least some bristle types have also undergone little change since the Eocene. These data further indicate that, during the warm Eocene, additional Mallomonas species possessing asymmetrical helmet or lance-shaped bristles existed, and that a taxon possibly related to the modern endemic species M. galeiformis was present in northern North America water-bodies.
Acknowledgments
This research was supported by the National Science Foundation (USA; NSF-DEB-0343355 and NSF-DEB-0716606) and the Natural Sciences and Engineering Research Council (Canada). We thank BHP Billiton Diamonds Inc. (Kelowna, Canada) and the Geological Survey of Canada (Calgary) for providing access to the Giraffe Pipe material and Jørgen Kristiansen for providing the Latin diagnoses. Jim Romanow and Marie Cantino (University of Connecticut) kindly aided with SEM facilities.
Related Research Data
References
- Asmund , B and Kristiansen , J . 1986 . The genus Mallomonas (Chrysophyceae). A taxonomic survey based on the ultrastructure of silica scales and bristles . Opera Botanica , 85 : 1 – 128 .
- Brinkhuis, H., Schouten, S., Collinson, M.E., Sluijs, A., Sinninghe Damsté, J.S., Dickens, G.R., Huber, M., Cronin, T.M., Onodera, J., Takahashi, K., Bujak, J.P., Stein, R., van der Burgh, J., Eldrett, J.S., Harding, I.C., Lotter, A.F., Sangiorgi, F., van Konijnenburg-van Cittert, H., de Leeuw, J.W., Matthiessen, J., Backman, J., Moran, K. & The Expedition 302 Scientists. (2006). Episodic Fresh surface waters in the Eocene Arctic Ocean. Nature, 441: 606–609
- Canavan , RW and Siver , PA . 1994 . Chemical and physical properties of Connecticut lakes . Lake and Reservoir Management , 10 : 173 – 186 .
- Greenwood , DR and Basinger , JF . 1994 . The paleoecology of high latitude Eocene swamp forests from Axel Heiberg Island, Canadian High Arctic . Rev. Paleobot. Palynol. , 81 : 83 – 97 .
- Kristiansen , J . 2002 . The genus Mallomonas (Synurophyceae): A taxonomic survey based on the ultrastructure of silica scales and bristles . Opera Bot. , 139 : 1 – 218 .
- Kristiansen , J and Lind , JF . 2004 . Endemicity in silica-scaled chrysophytes . Nova Hedwig. Beih. , 128 : 65 – 83 .
- Lott , AM and Siver , PA . 2005 . An inventory of scaled chrysophytes from the Atlantic Coastal Plain of North Carolina, USA . Proceedings of the Sixth International Chrysophyte Symposium , : 211 – 230 .
- Nicholls , KH . 1987a . The distinction between Mallomonas acaroides v. acaroides and Mallomonas acaroides v. muskokana var. nov. (Chrysophyceae) . Can. J. Bot. , 65 : 1779 – 1784 .
- Nicholls , KH . 1987b . Form variation in Mallomonas asmundiae and a description of Mallomonas sphagniphila sp. nov. (series Corconticae, Mallomonadaceae) . Can. J. Bot. , 65 : 627 – 634 .
- Nicholls , KH . 1988 . Descriptions of three new species of Mallomonas (Chrysophyceae). M. hexagonis sp. nov., M. liturata sp. nov., and M. galeiformis sp. nov . Br. Phycol. J. , 23 : 159 – 166 .
- Nicholls , KH . 1990 . The taxonomy of problematic Mallomonas species (Synurophyceae). II. M. costata and M. galeiformis . Nord. J. Bot. , 10 : 243 – 247 .
- Pagani , M , Zachos , JC , Freeman , KH , Tipple , B and Bohaty , S . 2005 . Marked decline in atmospheric carbon dioxide concentrations during the Paleogene . Science , 309 : 600 – 603 .
- Siver , PA . 1987 . The distribution of Synura species (Chrysophyceae) in Connecticut, U.S.A. including the description of a new form . Nordic J. Bot. , 7 : 107 – 116 .
- Siver , PA . 1988 . Morphology and ecology of Mallomonas galeiformis, a potentially useful paleolimnological indicator . Trans. Am. Microsc. Soc. , 107 : 152 – 161 .
- Siver , PA . 1989 . The separation of Mallomonas acaroides v. acaroides and v. muskokana (Synurophyceae) along a pH gradient . Nova Hedwig. Beih. , 95 : 111 – 117 .
- Siver , PA . 1991a . The Biology of Mallomonas: Morphology, Taxonomy and Ecology , Dordrecht, , The Netherlands : Kluwer Academic Publishers .
- Siver , PA . 1991b . The stomatocyst of Mallomonas acaroides v. muskokana (Chrysophyceae) . J. Paleolimnol. , 5 : 11 – 17 .
- Siver , PA . 2003 . “ The Synurophyceae ” . In Freshwater Algae of North America , Edited by: Wehr , J and Sheath , B . 523 – 558 . New York (NY), , USA : Academic Press .
- Siver , PA and Lott , AM . 2006 . Further observations on the scaled Chrysophyceae and Synurophycean flora of the Ocala National Forest, Florida, U.S.A . Nordic J. Bot. , 24 : 211 – 233 .
- Siver , PA and Skogstad , A . 1988 . The morphology and ecology of Mallomonas crassisquama . Nordic J. Bot. , 8 : 99 – 107 .
- Siver , PA and Wolfe , AP . 2005a . Eocene scaled chrysophytes with pronounced modern affinities . Int. J. Plant Sci. , 166 : 533 – 536 .
- Siver , PA and Wolfe , AP . 2005b . Scaled Chrysophytes in Middle Eocene Lake Sediments From Northwestern Canada, Including Descriptions of Six New Species . Proceedings of the Sixth International Chrysophyte Symposium , : 295 – 308 .
- Siver , PA , Hamer , J and Kling , H . 1990 . Separation of Mallomonas duerrschmidtiae sp. nov. from M. crassisquama and M. pseudocoronata: Implications for paleolimnological research . J. Phycol. , 26 : 728 – 740 .
- Siver , PA , Hamilton , PB , Stachura-Suchoples , K and Kociolek , JP . 2005 . Diatoms of North America: The Freshwater Flora of Cape Cod . Iconographia Diatomologica , 14 : 1 – 463 .
- Takahashi , E . 1978 . Electron microscopical studies of the Synuraceae (Chrysophyceae) in Japan, taxonomy and ecology , Tokyo, , Japan : Tokai University Press .
- Wee , J . 1982 . Studies on the Synuraceae (Chrysophyceae) of Iowa . Bibl. Phycol. , 62 : 1 – 183 .
- Wolfe , AP , Edlund , MB , Sweet , AR and Creighton , S . 2006 . A first account of organelle preservation in Eocene nonmarine diatoms: observations and paleobiological implications . Palaios , 21 : 298 – 304 .
- Zachos , JC , Pagani , M , Sloan , L , Thomas , E and Billups , K . 2001 . Trends, rhythms, and aberrations in global climate 65 Ma to present . Science , 292 : 686 – 693 .