Abstract
Plastids of Climaconeis (Okedenia) were originally characterized by Mereschkowsky as H-shaped in girdle view with pyrenoids but recent descriptions implicitly or explicitly indicate that the H-shape is seen in valve view. We observed five known taxa–one a variety that we raise to species rank–and two new species: C. silvae, C. riddleae, C. guamensis sp. nov. and C. inflexa (curved species); C. lorenzii (straight species with craticular bars); a straight species without bars that matches Navicula scopulorum var. triundulata = Climaconeis undulata comb. nov.; and a new straight species without bars, C. petersonii sp. nov. In the species with multiple plastids, plastids are plate-like, usually with a pyrenoid in the centre, arranged in pairs along the girdle faces of the cells. In some species with few plastids, including C. riddleae, and possibly C. inflexa, the pyrenoid is also visible in valve view between opposing plastids, such that these species do appear to have plastids that are H-shaped in valve view. We describe the two new species and confirm published SEM observations on other species, including features of the pores that help distinguish C. guamensis from the very similar C. riddleae.
Keywords:
Introduction
Tropical benthic marine diatoms form a significant component of the photosynthetic biodiversity in many habitats on and associated with coral reefs (Lobban & Jordan, Citation2010) but tropical floras have been studied far less than temperate floras. The prospect for new species is evident in the small genus Climaconeis Grunow (Citation1862) emend. E.J. Cox (Citation1982). Six warm-water species have been added during the past decade bringing the total to 13 currently recognized species (Prasad, Citation2003), of which only three are reported as temperate (Reid & Williams, Citation2002). Prasad and co-workers (Prasad et al., Citation2000; Prasad, Citation2003) named four species from Florida (C. koenigii A.K.S.K. Prasad, C. riddleae A.K.S.K. Prasad, C. colemaniae A.K.S.K. Prasad and C. silvae A.K.S.K. Prasad); Reid & Williams (Citation2002) described C. coxii G. Reid & D.M. Williams and C. ghurbensis G. Reid & D.M. Williams from Abu Dhabi. Cox (Citation1982) and Prasad et al. (Citation2000) gave the taxonomic history of the genus and Reid & Williams (Citation2002) sketched the geographic distribution. In the Bahamas (Hein et al., Citation2008), C. koenigii was a dominant species in enumerated samples, C. colemaniae was abundant and C. lorenzii Grunow was common. Navicula scopulorum var. triundulata Meister (Citation1932) was described from Xiamen, China (see Taxonomic accounts below) and C. lorenzii was reported from Palawan, Philippines [Podzorski & Håkansson, Citation1987, as Navicula lorenzii (Grunow) Hustedt]; these are apparently the only Pacific records of Climaconeis to date.
Climaconeis frustules can be curved or straight and some straight species have craticular bars on the valvocopula; most species are readily recognized in living material by the presence of a series of several to many large plastids, which are described as H-shaped with a central pyrenoid (Cox, Citation1979; Prasad et al., Citation2000). However, when we began to observe species in Guam it became clear that some confusion exists about both the shape of the plastid and the location of the pyrenoid and that the generic description of the plastids might need amending.
The history of the H-shaped plastid dates back to Mereschkowsky (Citation1901), whose interest in plastids led him to develop the first endosymbiont hypothesis of plastid origin (Martin & Kowallik, Citation1999). He resurrected Okedenia Eulenstein on the basis of H-shaped plastids with large pyrenoids, including taxa that were then known as Amphora inflexa (Brébisson ex Kützing) H.L. Smith and Navicula scopulorum Brébisson in Kützing. He drew and described plastids that were flat plates in pairs along the girdle faces (). Not all the taxa he illustrated can be clearly assigned to genera (Cox, Citation1982), but his observations of O. scopulorum [=Climaconeis scalaris], O. scopulorum var. fasciculata [=Climaconeis fasciculata] and O. inflexa [=Climaconeis inflexa (Brébisson) Cox] are useful. In the type form of O. scopulorum and in O. inflexa the plastids were H-shaped in girdle view. In the two O. scopulorum taxa the pyrenoids lay entirely within the thickness of the plastid plates but in O. inflexa they protruded inward and met in the middle. Of the six species Cox (Citation1982) included in Climaconeis after her exhaustive study, including Okedenia spp., two have not been observed alive and three have been observed only by Mereschkowsky. Cox (Citation1979) observed C. scalaris (Brébisson) Cox alive, and paid particular attention to its plastids in distinguishing it from Berkeleya scopulorum (Brébisson) Cox (). In her revision of Climaconeis, Cox (Citation1982) followed Mereschkowsky and wrote in her emended description that there are “pairs of H-shaped chloroplasts with conspicuous pyrenoids”. The drawing Cox (Citation1979) presented () looks like Mereschkowsky's drawing of O. scopulorum var. fasciculata (), i.e., a linear cell with quadrate plastids as seen in girdle view rather than his drawing of O. scopulorum (). This variety was raised to species rank (in Navicula) by Hustedt (Citation1961) and Cox (Citation1982) renamed it C. fasciculata (Grunow ex Cleve) Cox; she listed Mereschkowsky's O. scopulorum as synonymous with C. scalaris. In describing live material, Cox (Citation1979) drew faint cytoplasmic connections between some pairs of plastids (), but her drawings do not indicate what she meant by “H-shaped” plastids. Cox & Williams (Citation2006) analysed the plastids in naviculoid diatoms and classified Climaconeis (based on C. scopulorum and C. inflexa) in a group called “two (or more) chloroplasts (each with a central pyrenoid) arranged fore and aft in the cell”. This group was exemplified by Aneumastus spp., in which (their ) they clearly drew the H-shape as seen in valve view and they describe the character state as a 3-dimensional plastid, not as pairs of 2-dimensional plates. Confusion in the number of plastids has crept into recent keys distinguishing C. scalaris and C. fasciculata (see below, under Climaconeis undulata).
Figs 1–4. Drawings of Climaconeis plastids from the literature. . Mereschkowsky's (Citation1901) Okedenia. . O. scopulorum. . O. scopulorum var. fasciculata. . O. inflexa. . Cox's (Citation1979) Navicula scopulorum B. [. reprinted with permission of the British Phycological Society.]
![Figs 1–4. Drawings of Climaconeis plastids from the literature. Figs 1–3. Mereschkowsky's (Citation1901) Okedenia. Fig. 1. O. scopulorum. Fig. 2. O. scopulorum var. fasciculata. Fig. 3. O. inflexa. Fig. 4. Cox's (Citation1979) Navicula scopulorum B. [Fig. 4. reprinted with permission of the British Phycological Society.]](/cms/asset/6049e774-e5ba-46aa-88ec-82a61cbd88e8/tejp_a_490924_o_f0001g.gif)
Among the recently described species, Prasad et al. (Citation2000) were confident that the plastids of C. koenigii were H-shaped and said that those of C. colemaniae were “probably” H-shaped, but they did not mention pyrenoids; nor did Prasad (Citation2003) mention pyrenoids in the two most recent species. In C. riddleae, Prasad (Citation2003) noted “4 (rarely 2)” plastids; he did not characterize these as H-shaped and his suggests either 4 plates along the girdle face or 2 H-shaped; it seems unlikely in this genus that there would be only a single plate on each side of the central cytoplasm. Reid & Williams (1990) were confident that the plastids of C. coxii were H-shaped and wrote that those of C. ghurbensis were “probably” so. As far as we can tell from the published text and illustrations, none of these species was seen alive. The only report from living Climaconeis is John's (Citation1991) account of C. stromatolitis. This species, like the very similar C. colemaniae and the arcuate C. inflexa, has only two plastids, H-shaped in valve view (4 joined plates in Mereschkowsky's terms). Nevertheless, Prasad et al. (Citation2000: 210) revised Cox's English description of the genus and wrote of “H-shaped plastids distributed along the cell, each composed of two girdle-opposed plates connected by a central bridge containing a pyrenoid”. This description, like Cox & Williams’ (Citation2006) more recent characterization of C. scalaris, suggests that the H shape is seen in valve view.
The objectives of this study were to describe the Climaconeis flora from Guam by light and scanning electron microscopy of cells from field collections and cultures, and to determine the character of the plastids and pyrenoids. We find that in the majority of species, plastids are platelike, usually with pyrenoids, and arranged in pairs along the girdle faces of the cells. The previously described taxa are all new records for these islands.
Materials and methods
Study sites and sampling
Samples of diatom communities epiphytic on seaweeds or epipelic on mud were collected by hand at low tide or during SCUBA or snorkelling observations of reefs, chiefly in Guam, with additional samples from Yap (Federated States of Micronesia) and Palau collections ().
Table 1. Collection sites for listed records.
Light microscopy
Fresh samples were examined and photographed in Guam with an Olympus SZ51 dissection microscope and a Nikon Eclipse 600 compound microscope with differential interference contrast and Olympus 7070 consumer cameras. Later a Nikon 80i DIC microscope with DS-Fi1 colour camera head and L2 Controller was used to record images and make measurements. Cells for culture were isolated either in Guam from fresh collections or in Austin from couriered samples.
Cultures
Isolates of Climaconeis riddleae (ECT 3722, isolate 17vi081A), C. guamensis (ECT 3737, isolate 19vi082G) and C. undulata (ECT 3743) were taken from wild collections in Guam at Umatac Bay (benthic turf sample) and Talofofo Bay (epiphytic on Sargassum sp.), respectively. Isolates were grown in Guillard's f/2 medium (Guillard, Citation1975) in a Percival model I-36LL incubation chamber at 27°C. Fluorescent lighting was used in the culture chamber in a 13:11 h light:dark photoperiod.
SEM preparation
Wild and cultured material was centrifuged, the saltwater/media decanted and then rinsed in distilled water several times to remove excess salts. To remove organics, the material was then boiled with an equal volume of nitric acid for 15 minutes and left to sit in the acid overnight. Distilled water was then added and the material allowed to settle; after settling, the water was decanted. This was repeated until the pH of the sample was neutral.
Cleaned material was either allowed to dry on glass coverslips or filtered and dried over Millipore 1.2 µm Isopore membrane filters and fixed to SEM stubs with carbon tape. The samples were coated with 15 nm of iridium using a Cressington 208 Bench Top Sputter Coater. Observations were made on a Zeiss SUPRA 40 VP scanning electron microscope. Additional observations were made using whole mounts of preserved material, dried onto membrane filters, fixed onto stubs with carbon tape, coated with Au/Pd in an Energy Beam Sciences SC7620 Mini Sputter Coater, and observed in an FEI Phenom “tabletop” SEM.
Taxonomic accounts
We discuss the straight species and then the curved species, starting with those for which we have the most information.
STRAIGHT SPECIES
Climaconeis undulata (Meister) Lobban, Ashworth et Theriot, comb. nov.
,
Figs 5–18. Climaconeis plastids, freshly collected living cells, from Guam except as noted. . C. undulata. . Valve views, the cell in 5 showing some plastids possibly joined by pyrenoids (arrows). . Oblique view of small cell. . Girdle view showing pyrenoids. . C. petersonii. . Half of cell in valve view, composite image. . Central portion in girdle view, showing the conspicuous pyrenoids. . C. lorenzii from Palau, photographed after 24 h transport; arrowhead points to craticular bar. . C. silvae valve views. . Half cells showing concentration of plastids in the central area, plastids at an oblique angle in 12. . Details of plastids at several angles in valve views of cells. . C. riddleae (GU44Y-13) and enlargement to show striae. . Cells from GU44Y-13, probably C. riddleae but possibly C. guamensis, displaying pyrenoids in valve and girdle view (py). Scale bar for all = 20 µm.

Figs 19–25. Climaconeis undulata acid-cleaned valves. . Two valves showing radiating stria pattern; field-collected material (GU16V), DIC. . Valve of Navicula scopulorum var. triundulata; slide N11/17 in Hustedt collection, prepared by Hustedt from Meister's material, photo H52078-3a courtesy of AWI (Citation2009) (DIC). . SEM of cultured cells. . Internal apex showing helictoglossa. . External apex. . Internal central area. . External central area. Scale bars: , 10 µm; 22–25 = 2 µm.

BASIONYM: Gomphocaloneis undulata Meister Citation1932, Kieselalgen aus Asien, pp. 41–42, figs 122–123.
SYNONYMS: Navicula scopulorum Bréb. var. triundulata Meister (Hustedt, Citation1934, in Schmidt's Atlas, pl. 394, ); Navicula scopulorum Bréb. f. triundulata Meister (Hustedt, Citation1961, fig. 1186c).
HOLOTYPE: Meister's material appears to be in the Friedrich-Hustedt-Arbeitsplatz, Accession Number 3066, slide N11/17.
TYPE LOCALITY: Amoy Bay (now Xiamen, Fujian Province, China) (Meister Citation1932); “Sumatra, Flußmündung [River delta] des Belawan, 1, Juli 1927, Grundschlamm, leg Voigt, coll Meister” (Hustedt, Citation1934; and as marked on the slides in AWI, Citation2009).
DISTRIBUTION: Southern China, Sumatra?, Guam.
NEW RECORDS: Guam: GU16B, GU16V, GU16Z, GU29E, GU44V-1, GU61–UTA 23 VI 08-1A.
A population of cells initially identified as C. scalaris was first recorded from muddy intertidal rocks near the outfall of the Ylig River (GU16) in Nov. 1988 and was again present when we searched there in 2007 and 2008. The community was dominated by a dense population of the tube-dwelling Nitzschia vidovichii Grunow. This Climaconeis was also found in a similar habitat near low water in Talofofo Bay (GU61) by M.A. and cultured. A small, dense, epiphytic bloom was encountered in a strikingly different coral reef habitat, 10 m deep at GabGab (GU44), not subject to salinity fluctuations or significant sedimentation. A slide of the GabGab material has been deposited at California Academy of Sciences (accession # 627375, slide # 222100) along with type slides of the two new species.
Description
Cells () were straight, 99–117 µm long, 6–7 µm wide at the apex, 7–9 µm wide at the centre (GU16V population), the sides slightly inflated at the apices and centre, and in most cells also halfway between the apex and centre, giving a slightly undulating outline (). Striae 18–20 in 10 µm, weakly radiating at the centre, convergent near the apex, parallel in between, continuing over the valve mantle, except at the apices where there was a radiating set of striae on the valve face separated from a row of punctae around the apical valve mantle (). Pores in the striae oblong, slightly longer in the apical axis. Raphe straight, not deflected at either the central or apical endings and having, in some specimens, an additional slit along one side such that the first row of pores appears displaced (). The helictoglossa characteristic of the genus was evident at the apical raphe endings (). Plastids 9–13 pairs (to 7 in short cells), each comprising a plate with the valve edges turned out slightly (); plastids evenly distributed on each side of nucleus or one more pair on one side. Because of the turned edges, the plastids appeared to have perpendicular lobes when seen in oblique view (). Pyrenoids were clearly seen in healthy cells in girdle view (), similar to Cox's (Citation1979) drawing of C. scalaris and Mereschkowsky's (Citation1901) drawing of C. fasciculata, but were difficult or impossible to see in degraded cells. In some cells () there appeared to be some connection between the plates, but the pyrenoids were not evident in valve view.
Taxonomy
The taxon most similar to our material is Gomphocaloneis undulata Meister (Citation1932), a species he described from China, which was quickly reduced to Navicula scopulorum var. triundulatum Meister by Hustedt (Citation1934). Hustedt (Citation1961: 26) subsequently changed it to the rank of form, citing synonymy with the Atlas listing merely as “als var”. This form has a slightly undulate outline (the character that Hustedt says distinguishes it from the type) and, as shown in Hustedt's (Citation1934, Citation1961) figures, striae that are weakly radiating at the centre, weakly convergent at the apex, and parallel in between. We have examined photos of cells from two slides in the Hustedt collection database (AWI, Citation2009), N11/17 prepared by Hustedt from Meister's material () and ZT7/35 apparently prepared by Meister. Both slides indicate that the material is from Belawan, Sumatra, as given in Hustedt's publications, but while Meister did report diatoms from Belawan samples in his 1932 paper, he gives the origin of the “Gomphocaloneis undulata” as Amoy [Xiamen, China]. The photos of “Navicula scopulorum” from slide W8/28–also some of Meister's material mounted by Hustedt–appear to us to be also the same variety and not the type.
The taxonomic position of Meister's taxon is unclear. While Mereschkowsky considered fasciculata to be a variety of scopulorum, Hustedt considered fasciculata sufficiently different, on the basis of striae being strongly radial at the centre and strongly convergent at the apices, to restore it to species rank; the differences are distinct in his figs 1186–1187. Hustedt saw G. undulata with its more weakly radiating/convergent striae as a form of N. scopulorum rather than of N. fasciculata or a separate species. In contrast, Cox (Citation1982) examined N. fasciculata specimens from the Cleve & Möller exsiccata and British Museum collections and her description and her and show cells with striae parallel except weakly radiating at the apices. She also discussed the taxa illustrated by Mereschkowsky and concluded that there were two similar species: C. scalaris smaller and narrower and with strictly parallel striae, versus the more robust C. fasciculata with striae weakly convergent at the apices. Although the keys in Prasad et al. (Citation2000), Reid & Williams (Citation2002), and Prasad (Citation2003) all indicate different numbers of plastids in these two taxa, with fewer than 10 in C. scalaris, Mereschkowsky's report of 20 plates in C. fasciculata, correctly cited by Cox, is the same as she (Citation1979) showed for C. scalaris (10 only if pairs are counted as one H-shaped plastid). In discussing undulata, Cox (Citation1982) referred to Meister's paper but did not change Hustedt's assignment of it as a form of scopulorum when she split C. scalaris out of N. scopulorum. It therefore remains unclear where Meister's taxon and our material should be placed. The weakly radiating striae in the central area and the undulate outline are not present in either C. scalaris nor C. fasciculata (). However, we observed the unusual additional slit–sometimes not opened up–alongside the raphe in our material, as Cox (Citation1979, , ) described for C. scalaris; we were unable to tell whether it had a row of pores hidden beneath it (cf. Cox Citation1979, ). There are no other recognized species of Climaconeis that are straight and lack both a stauros and craticular bars, except for our new species C. petersonii, which is much longer, with many more plastids (see below). In the absence of molecular data that might support placement of this taxon as a variety under either C. scalaris or C. fasciculata, and in the absence of modern observations of living material of C. fasciculata, we conclude that the best choice for now is to resurrect Meister's name in the new combination Climaconeis undulata.
Figs 26–35. . Climaconeis petersonii acid-cleaned valves, field collected material. . Type specimen, GU55B-4, slide 2. DIC. . Details of central area and apex of type specimen. DIC. . Internal and external central areas, SEM. . External and internal apices, the latter showing the helictoglossa. SEM. . Climaconeis lorenzii acid-cleaned valve from Yap (Y26C). . Frustule showing deflected central raphe endings. . Valvocopula with craticular bars. . Climaconeis silvae, portion of acid-cleaned valve; field-collected material, GU44I-2, DIC. Scale bars: = 10 µm; = 2 µm.

Table 2. Characteristics of selected Climaconeis species. Data from literature and present study.
Climaconeis petersonii Lobban, Ashworth et Theriot, sp. nov.
,
Cellulae rectae, leviter inflatae ad centrum et apices, carens transtris craticulis. Chloroplasti plani cum pyrenoidibus manifestis, circa 80 plerumque binati. Longitudo cellularum 350–380 μm, latitudo 7–8 μm praeter apices et centrum 9–10 μm. Extremitates centralis raphium rectae; striae 20–21 in 10 μm, ubique parallelae, cum poris quadratis vel rectangularibus apicaliter; carens stauro.
HOLOTYPE: Slide GU55B-4-1, deposited in California Academy of Sciences, accession # 627376, slide # 223001.
TYPE LOCALITY: GU55 Agat Bay, “Pete's Reef”, 13 Dec. 2008.
ETYMOLOGY: Species is named for Pete Peterson, who has done much to promote diving and reef conservation on Guam. “Pete's Reef” is a commercial dive site with one of the first mooring buoys installed so that dive boats would not anchor on the reefs.
Cells were straight, 350–380 µm long, 7–8 µm wide, inflated to 9–10 µm wide at centre (in both girdle and valve views) and apex, lacking craticular bars. Valve mantle very shallow (). Central raphe endings straight; 20–21 striae in 10 µm, parallel throughout, no stauros (). About 80 plastids mostly in pairs and most with conspicuous pyrenoids, more or less symmetrically distributed on each side of the central nucleus (). Pores very regular, square to apically rectangular ().
The shape of C. petersonii is most like C. colemaniae and C. lorenzii, but C. colemaniae has a stauros and much finer (35–36) striae; C. lorenzii has a similar number of striae (16) and no stauros, but has craticular bars. Our species is also considerably longer than either of these [except for Prasad et al.'s (Citation2000) very long C. lorenzii]. Climaconeis colemaniae has only 4 plastids. The plates of the plastids in this species sometimes appeared connected, at other times were evidently separate, or even had one plate missing, especially toward the apex.
Climaconeis lorenzii Grunow
,
REFERENCES: Hustedt, Citation1961, fig. 1188 and Podzorski & Håkansson, Citation1987, pl. 31, , both as Navicula lorenzii; Cox Citation1982, and (as Stictodesmis australis Grev.), and figs 4–5 (as Climaconeis frauenfeldii Grun.)
DISTRIBUTION: Widespread in warm oceans: Florida and Caribbean, Western Pacific (see Reid & Williams, Citation2000).
NEW RECORDS: Yap: Y26C, O’Keefe's Island, 25 Sep. 1988; Palau: Ulebsechel PW2009-54, 13 Apr. 2009; Guam: Double Reef, GU6C, 14 Sep. 2008.
To date we have positively recorded this species only from acid cleaned frustules and valvocopulae with craticular bars from a sediment sample from Yap (), but the live cell from Palau () and a valvocopula from Guam (not shown) appear to be this species rather than C. coxii. The key differences between C. lorenzii and C. coxii are in the central raphe endings, which in C. lorenzii are deflected to one side [clearly shown in Hustedt, Citation1961, fig. 1188, Podzorski & Håkansson, Citation1987, pl. 31, (both as Navicula lorenzii) and Reid & Williams, 2000, ] but in C. coxii are straight, and in the higher number of striae (20 in 10 µm C. lorenzii compared with 16 in C. coxii) (). These features cannot be seen when only the valvovopula and craticular bars are observed, as in our Guam sample and in Hein et al. (Citation2008) pl. 26, . Cells of C. coxii are shorter and narrower than C. lorenzii and our cells are relatively small, but C. lorenzii has been reported from New Caledonia (Greville's original description as Stictodesmis australis, see Cox, Citation1982), the Philippines (Podzorski & Håkansson, Citation1987) and Yap (this paper), whereas C. coxii is known so far only from Abu Dhabi (Reid & Williams, 2000). We therefore feel confident in assigning our Palau cell to C. lorenzii. In both species there is probably one pair of plastids between each two craticular bars (Cox, Citation1982; Prasad et al., Citation2000) and, although the craticular bars in our live cell were mostly obscured by the plastids, it appears from our figure that this suggestion is correct and that the nucleus and the plastids on each side of it share the larger space between the two central bars. In our cells there was one more bar and pair of plastids in one half than in the other; the asymmetry can also be seen in illustrations in Podzorski & Håkansson (Citation1987) and Cox (Citation1982).
CURVED SPECIES
Climaconeis silvae A.K.S.K. Prasad
, 35–38
REFERENCE: Prasad, Citation2003, figures 1–25.
DISTRIBUTION: Florida, Western Pacific islands.
NEW RECORDS: Guam: GU44I-1, GU44I-2, GU44AB-7. Palau: Mekeald reef, PW2009-8.
Cells 190–210 µm long, central area almost straight and slightly wider than the apices (10 vs.7 µm), distal portions of cell curved in the same direction. Striae 28–30 in 10 µm, parallel throughout. Raphe markedly biarcuate, each half curving across the valve face from the convex (dorsal) side at the apex to the concave (ventral) side at the central ends. Plastids 12–18 pairs of lozenge-shaped plates, more or less symmetrical on each side of the central nucleus, generally restricted to the central straight portion, leaving the curved ends of the cell clear (). There was a dimple in the girdle face of the plastid but in the rare girdle views we were unable to observe pyrenoids. We could detect no pyrenoids between the plates, but there were sometimes indistinct cytoplasmic connections between the paired plates. The shape of the plastids often appeared complex, with the valvar edges of the plastid giving an impression of four perpendicular lobes, but in those cases the plastid pair was in oblique view, even though the cell itself was in valve view (). Areolae in our specimens were irregularly elongate (), as shown by Prasad (Citation2003).
Figs 36–38. Climaconeis silvae SEM of cells cultured from Guam collection. . Internal and external apices and girdle bands. . Internal central area. . External central area. Scale bars = 2 µm.

The shape of C. silvae is generally similar to all the other curved species except the more lunate C. inflexa–i.e. C. ghurbensis, C. koenigii and C. riddleae, but these all have distinctive combinations of size, numbers of plastids and stria density (). Climaconeis riddleae has only 2 H-shaped plastids, C. ghurbensis and C. koenigii have 40 or more (counted as pairs or H-shaped), in contrast to the 8–18 reported for C. silvae.
Climaconeis riddleae A.K.S.K. Prasad
, 18(?),
REFERENCES: Prasad, Citation2003, figs 26–41.
Figs 39–43. Climaconeis riddleae from Guam. . Acid-cleaned valve from GU44Y-13. . SEMs from isolate ECT 3722. . External and internal views of central area. . Internal and external views of apex. Scale bars: = 10 µm; = 2 µm.

DISTRIBUTION: Florida, Guam.
RECORDS: Guam: GU44Y-13; GU60, isolate ECT 3722.
Several valves were observed in a live sample and in acid-cleaned material that also included C. guamensis (see below) and both had been isolated into culture. They are almost the same size but the striae of C. riddleae were clearly visible under oil immersion, and can even be seen in live cells (), whereas those of C. guamensis could not. Climaconeis riddleae had only 27 striae in 10 µm, conforming to Prasad's species from Florida Bay and was smaller than his C. silvae. In addition, the curving raphes of C. riddleae are almost central, whereas those of C. silvae and C. guamensis arc toward the convex side of the cell; both species also have stria counts >30 in 10 µm. The plastids on each side of the central area appear H-shaped in valve view () but it is not clear if the plastids are separate plates joined by compound pyrenoids or three-dimensional plastids as in Cox & Williams’ (Citation2006) Aneumastus type. In girdle view, apparently, the pyrenoids are visible and the plastids are also somewhat H-shaped (), but we must qualify that it is hard to get girdle views of these curved cells, and in girdle view there is no way to distinguish C. riddleae from C. guamensis. Cultured cells of C. riddleae did not show pyrenoids.
Climaconeis riddleae and C. guamensis are readily distinguished in the SEM by the pore shape. Climaconeis riddleae has square pores arranged in apical and transapical rows; some pores are slightly elongated in the apical axis. There is an irregularly thickened raphe sternum () in C. riddleae as shown by Prasad (Citation2003).
Climaconeis guamensis Lobban, Ashworth et Theriot, sp. nov.
Figs 44–52
Cellulae arcuatae, leviter capitatae, cum 2 chloroplastis H-formis. Longitudo cellularum 90–120 μm, latitudo 3–4 μm. Raphe biarcuata accedens versus marginem convexum, moderate deflexa versus latus ventrale ad extrema centralia. Striae 33–34 in 10 μm, ubique parallelae cum poris plus minusve rectangularibus et elongatis transapicaliter.
HOLOTYPE: GU44Y-13, slide 1; deposited in California Academy of Sciences, accession # 627373, slide # 222098.
SYNTYPE: GU59, Pago Bay near Marine Laboratory, 17 June 2008, slide made from cultured isolate ECT 3737, deposited in California Academy of Sciences, accession # 627374, slide # 222099.
TYPE LOCALITY: GabGab reef, Apra Harbor, Guam, 10 May 2009.
ETYMOLOGY: This species is named for the island where it was discovered.
ADDITIONAL RECORDS: Guam: GU44AB-1, GU54A-F.
Cells arcuate, weakly capitate, 90–120 µm long, 3–4 µm wide (). Raphe biarcuate, arcing toward the convex (dorsal) side with central endings deflected toward the ventral side at the central endings (). Striae 33–34 in 10 µm, parallel throughout, barely visible in LM. Plastids similar to those of C. riddleae. In a live sample with both species (GU44Y-13) we were unable to distinguish the two unless striae were visible (C. riddleae). Confirmation of plastid form came from a sample () in which the material on the slide used for live photographs was preserved and later dried onto a filter and examined with SEM. Pores rounded rectangular, irregularly elongated transapically (, ).
Figs 44–49. Climaconeis guamensis acid-cleaned valves. . Holotype specimen, GU44Y-13, slide 1. DIC. . Specimen from isotype slide, isolate ECT 3737. DIC. . SEM of cultured cells, isolate ECT 3737. . Internal and external views of apex. . External and internal views of central area. Scale bars: = 10 µm; = 2 µm.

Figs 50–52. Climaconeis guamensis from GU54A-F. . Live cell. . SEM from whole mount of same material. Scale bars: = 50 µm; = 5 µm.
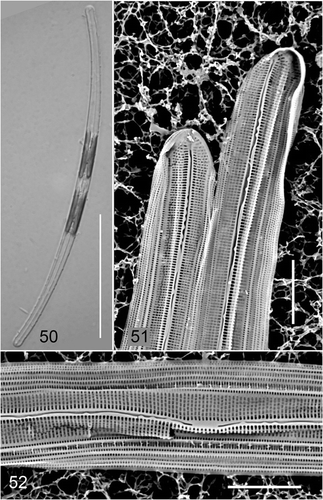
The species described here is similar in size and shape to the two smaller curved Climaconeis with two/four plastids, i.e. C. inflexa and C. riddleae, but has denser striae than these (). The differences between C. guamensis and C. riddleae are (1) the number of striae, (2) the shape of the areolae, which in the former are transapically elongated (as in C. silvae), whereas those of C. riddleae are square or slightly elongated apically (as in C. petersonii); (3) C. guamensis is weakly capitate, although this is not always evident. The overall cell shape, the curvature of the raphe, stria density and areola shapes are all similar in C. guamensis and C. silvae, but the latter, like C. ghurbensis, has numerous plastids. The size ranges of C. guamensis and C. silvae do not overlap.
Climaconeis inflexa (Brébisson) Cox
REFERENCES: Cox Citation1982, , ; Hendey Citation1964, pl. 37, ; Mereschkowsky Citation1901 (all as Okedenia inflexa).
RECORDS: Guam: GU7.
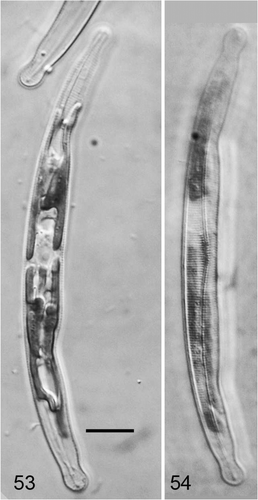
This species is known from Guam so far only from photographic records made in 2005. However the following characteristics are evident from the Guam material that strongly suggest this is C. inflexa. Cells arcuate-lunate, the convex side uniformly curved, the concave side of the valve straight in the middle section, resulting in a wider central area than the apices; raphe biarcuate. Apices distinctly capitate. Striae 18 in 10 µm, parallel at the centre. Plastids appear H-shaped in valve view; as with C. guamensis, it is not clear if the plastids are separate plates joined by compound pyrenoids or one plastid.
The cells described here are within the ranges of C. inflexa as described by Mereschkowsky (Citation1901) and Cox (Citation1982). Cell shape, plastid shape and stria density distinguish it from C. riddleae and C. guamensis ().
Figs 53–54. Climaconeis inflexa from Guam. . Cell with plastids in focus. . Cell with striae in focus. Scale bar = 10 µm.
Discussion
It is evident from our observations that the plastid characters of Climaconeis vary enough that they cannot be generically characterized as H-shaped, although this interpretation does fit some with only two plastids (C. riddleae, C. guamensis, C. inflexa, C. stromatolites and perhaps C. colemaniae). Nevertheless, the species with multiple plastids are readily recognized in live material. Although Mereschkowsky (Citation1901) showed “pairs of H-shaped chloroplasts” (as stated by Cox in her emended description), the H shape, in girdle view, has not been seen in recent studies, nor in the present study with one possible exception. In most cells the two plates are close together and there may be some more or less distinct material between them, as illustrated also in Cox's (Citation1979) drawing of C. scalaris. We conclude that Climaconeis plastids are usually 2-dimensional plates; the distinction–and the variation–is significant if plastids are categorized, since plastids that are H-shaped in valve view are 3-dimensional (Cox & Williams, Citation2006). Pyrenoids can be detected with acidified Azocarmine-G (Schmid, Citation2001), but it may take transmission electron microscopy to settle whether the material between the plates is a single pyrenoid within one plastid or a compound structure between two [cf. plastids in the phaeophyte Asteronema breviarticulatum (J. Agardh) Ouriques & Bouzon (Ouriques & Bouzon, Citation2000)]. In this study, our objectives were to reexamine whether the character of a “prominent” pyrenoid was consistent, and clarify its location, rather than exploring the potential of new pyrenoid characters to support taxonomy. The pyrenoid, when visible, is usually in the middle of each plate rather than in the cytoplasm between them, and we must revise the emended description given by Prasad et al. (Citation2000). The revised wording also takes into the account the occurrence of Climaconeis as epiphytes (Reid & Williams, 2000 and this study) and in fouling circumstances (Reid & Williams, 2000); they are clearly not restricted to epipelic habitats. We propose the following opening lines for the generic description of Climaconeis (Grun.) emend. E.J. Cox:
Cells marine; epipelic, epiphytic, or fouling; plastids 4 to numerous plates, usually paired along the girdle faces, often with pyrenoids in the plates, or 2 three-dimensional plastids, H-shaped in valve view, with pyrenoid between. Plastids distributed more or less symmetrically on either side of the central nucleus, throughout the cell or restricted to the middle portion.
The genus Climaconeis is similar to Berkeleya and hard to define, especially on the basis of valve characters, because of the diverse characters (especially straight versus curved valves and the presence or absence of craticular bars). Although Cox (Citation1979) was able to use plastids to distinguish C. scalaris from B. scopulorum, some Climaconeis have H-shaped plastids like Berkeleya, but so far at least two such plastids, whereas Berkeleya species have only one. The “tuning-fork” appearance of the raphe in Berkeleya species that have distant central raphe endings is not seen in B. scopulorum (Cox, Citation1979). Berkeleya species usually occur in mucilage tubes, whereas Climaconeis are solitary.
As we discover more species of Climaconeis, some very similar to others, it is becoming increasingly important to use fine structure characters. One such character that seems useful is the shape of the areolae. There are at least three groups: species with very regular square–rectangular areolae, e.g. C. ghurbensis (Reid & Williams, Citation2002, ), C. riddleae and C. petersonii; those with irregularly elongated areolae, e.g. C. silvae and C. guamensis; and those with circular pores, e.g. C. colemaniae and C. koenigii (Prasad et al., Citation2000), and C. scopulorioides Hustedt (Cox, Citation1982: figs 23–26). Other species have intermediate or distinctive pores, and do not fit into these groups, e.g. C. inflexa (Cox, Citation1982: ), C. coxii (Reid & Williams, Citation2002, figs 25–29), C. stromatolitis (John, Citation1991) and C. undulata.
In our experience, several Climaconeis species were notable for occurring in dense but very localized patches, a few centimetres in diameter. Most were collected because they formed a visible brown coating on other algae, such as dense clumps of Halimeda opuntia (on reef flats in Guam and Palau) and filamentous turf. Hein et al.'s (Citation2008) records of high per cent frequencies of Climaconeis in enumerated samples–maximum 100% for C. koenigii–suggests that they also collected such patches, and Prasad (Citation2003) mentioned large numbers of C. silvae in some collections.
This is clearly an interesting genus in terms of biodiversity, particularly in warm waters, with ten species added since Cox's (Citation1982) taxonomic revision, and we expect more new species and many new records of known species in future tropical collections. Their plastids make most Climaconeis species distinctive in live material.
Acknowledgements
The microscopes used at U. Guam were funded by NIGMS RISE award GM063682 and US Dept. Education MSEIP award P120A040092. Funding for the Theriot laboratory was through NSF EF-0629410.
CL thanks COMNAVMAR for access to the study site on Guam Naval Facility, María Schefter for field support, and Friedel Hinz at Alfred-Wegener-Institut für Polar-und Meeresforschung for help with the Friedrich-Hustedt-Arbeitsplatz database.
References
- AWI (2009). Friedrich-Hustedt-Arbeitsplatz database. http://www.awi.de/en/research/research_divisions/biosciences/biological_oceanography/diatom_centre/. Accessed 21 Sep. 2009.
- Cox , EJ . 1979 . Studies on the diatom genus Navicula Bory. Navicula scopulorum Bréb. and a further comment on the genus Berkeleya Grev . Br. Phycol. J. , 14 : 161 – 174 .
- Cox , EJ . 1982 . Taxonomic studies on the diatom genus Navicula Bory. IV. Climaconeis Grun., a genus including Okedenia inflexa (Bréb.) Eulenst. ex de Toni and members of Navicula Sect. Johnsonieae sensu Hustedt . Br. Phycol. J. , 17 : 147 – 168 .
- Cox , EJ and Williams , DM . 2006 . Systematics of naviculoid diatoms (Bacillariophyta): a preliminary analysis of protoplast and frustule characters for family and order level classification . Systematics Biodiv. , 4 : 385 – 399 .
- Grunow , A . 1862 . Die österreichischen Diatomeen nebst Anschluss einiger neuen Arten von andern Lokalitäten und einer kritischen Übersicht der bisher bekannten Gattungen und Arten. Erste Folge. Epithemieae, Meridioneae, Diatomeae, Entopyleae, Surirelleae, Amphipleureae . Verhandlungen der kaiserlich-königlichen zoologish-botanischen Gesellschaft in Wien , 12 : 315 – 472 .
- Guillard , RRL . 1975 . “ Culture of phytoplankton for feeding marine invertebrates ” . In Culture of Marine Invertebrate Animals , Edited by: Smith , WL and Chanley , MH . 26 – 60 . New York : Plenum Press .
- Hein , MK , Winsborough , BM and Sullivan , MJ . 2008 . Bacillariophyta (Diatoms) of the Bahamas , Ruggell : Gantner Verlag .
- Hendey , NI . 1964 . An Introductory Account of the Smaller Algae off British Coastal Waters. Part V. Bacillariophyceae (Diatoms) , London : Her Majesty's Stationery Office .
- Hustedt, F. (1934). Vorläufige Erläuterungen zu Dr. A. Schmidt's Atlas der Diatomaceenkunde. Tafel 394. J. Kramer Reprint.
- Hustedt, F. (1961). Die Kieselalgen Deutschlands, Österreichs und der Schweiz. Rabenhorst's Kryptogamenflora Band 7, Teil 3. Lief 1. Koeltz, Koenigstein.
- John , DM . 1991 . Climaconeis stromatolitis, a new species of diatom from Shark Bay, Western Australia . Diatom Res. , 6 : 49 – 54 .
- Lobban, C.S. & Jordan, R.W. (2010). Diatoms on coral reefs and in tropical marine lakes. In The Diatoms: Applications for the Environmental and Earth Sciences. 2nd ed. (Smol, J.P., Stoermer E.F., editors). Cambridge University Press, Cambridge. (in press).
- Martin , W and Kowallik , KV . 1999 . Annotated English translation of Mereschkowsky's 1905 paper ‘Über Natur und Ursprung der Chromatophoren im Pflanzenreiche’ . Eur. J. Phycol. , 34 : 287 – 295 .
- Meister , F . 1932 . Kieselalgen aus Asien , Berlin : Gebrüder Borntraeger .
- Mereschkowsky, C. (1901). On Okedenia Eul. Ann. Mag. Nat. Hist., Ser. 7, 8: 415–423 and plate 7.
- Ouriques , LC and Bouzon , ZL . 2000 . Stellate chloroplast organization in Asteronema breviarticulatum comb. nov. (Ectocarpales, Phaeophyta) . Phycologia , 39 : 267 – 271 .
- Podzorski , AC and Håkansson , H . 1987 . Freshwater and marine diatoms from Palawan (a Philippine island) . Bibliotheca Diatomologica , 13 : 1 – 245 .
- Prasad , AKSK . 2003 . Fine structure and taxonomy of two species of the marine diatom genus Climaconeis (Berkeleyaceae, Bacillariophyta): C. silvae sp. nov. and C. riddleae sp. nov. from the Caribbean Sea and Florida Bay, U.S.A . Hidrobiológica , 13 : 9 – 22 .
- Prasad , AKSK , Riddle , KA and Nienow , JA . 2000 . Marine diatom genus Climaconeis (Berkeleyaceae, Bacillariophyta): two new species, Climaconeis koenigii and C. colemaniae, from Florida Bay, U.S.A . Phycologia , 39 : 199 – 211 .
- Reid , G and Williams , DM . 2002 . The marine diatom genus Climaconeis (Berkeleyaceae, Bacillariophyta): two new species from Abu Dhabi, United Arab Emirates . Diatom Res. , 17 : 309 – 318 .
- Schmid , A-MM . 2001 . “ Value of pyrenoids in the systematics of the diatoms: their morphology and ultrastructure ” . In 16th International Diatom Symposium , Edited by: Economou-Amilli , A . 1 – 31 . Greece : University of Athens .