
Abstract
Two new marine diatom species from Argentinean coastal waters, Pleurosigma hinzianum Sterrenburg, Sunesen & Sar, sp. nov. and P. frenguellianum Sunesen, Sterrenburg & Sar, sp. nov., are described. The characters permitting their identification are specified, based on comparison with type material of the morphologically similar species P. amara Stidolph and P. elongatum W. Smith in the light (LM) and scanning electron microscope (SEM). New information on the type material of P. elongatum is presented and its taxonomic concept is emended. The main criteria for separation of the species discussed here are: valve outline, path of the raphe-sternum, raphe angle and angle of intersection of the oblique striae in LM; and external central and terminal raphe fissures, internal details of the central area, external and internal apical structure, and morphology of the hymen-occluded internal pores in SEM. The occurrence of these (and probably other) species in the genus Pleurosigma in net samples is adventitious and not indicative of a true planktonic mode of life.
Introduction
The genus Pleurosigma W. Smith has a complex taxonomic history (Sar et al., Citation2012) and now includes more than 306 accepted species (Reid, Citation2012). It is characterized by more or less flattened valves that are sigmoid to straight in outline, a raphe-sternum that is sigmoid and more or less eccentric or straight and central, three stria systems (one transverse and two oblique) and loculate areolae with apically elongated foramina externally and hymen-occluded pores internally.
Diagnostic features for separating species of Pleurosigma with the LM were summarized and discussed by Sterrenburg (Citation1991a) and include valve shape, raphe-sternum path, raphe angle, intersection angle of the oblique striae, apical structure and stria density. Additional criteria for species characterization with SEM proposed by Cardinal et al. (Citation1989) and Sterrenburg (Citation1991b) are: morphology of the central bars, central area and external central raphe fissures, and the organization of the areolae around the central nodule. Stidolph (Citation1992), Reid (Citation2002) and Sar et al. (Citation2012) included additional ultrastructural features that were shown to be stable with a small intraspecific but a marked interspecific variation: morphology of the internal occluded pores of the areolae and the external terminal raphe fissures and fine external and internal apical details.
As part of a study of diatom populations from coastal waters of the Provinces of Buenos Aires and Río Negro, Argentina, numerous species and one variety of the genus Pleurosigma collected in several net samples were described (Sar et al., Citation2013; Sunesen et al., Citation2013). Here we describe two new taxonomic entities which at first sight are similar in general appearance to Pleurosigma amara Stidolph and P. elongatum W. Smith respectively.
Pleurosigma amara was described by Stidolph (Citation1992) from a salt marsh near Napier, New Zealand. Pleurosigma elongatum was described by Smith (1852: 6, pl. 1, ) from marine and brackish material from Poole Bay, UK, without designation of a holotype. The species was characterized by having ‘valves linear-lanceolate, flexure slight, extremities acute, median line central …’. Dimensions given (converted to metric values) were: length 330 µm, width 25 µm and stria separation of 0.56 µm (c. 18 striae in 10 µm). Additional morphometric data were given in Smith (1853: 64). Subsequently, Hendey (Citation1964) examined the slide BM23651 from the W. Smith collection and designated this as the type slide of P. elongatum. Stidolph (Citation1992) morphologically separated P. elongatum from P. amara and Reid (Citation2002) showed a few ultrastructural details of P. elongatum, based on SEM study of the type material.
The aims of this study are to describe two new taxa – which are rather similar in LM but clearly different in SEM – and to compare them with two morphologically similar species, P. amara and P. elongatum. The taxonomic concept of P. elongatum is emended, based on examination of its type material.
Materials and methods
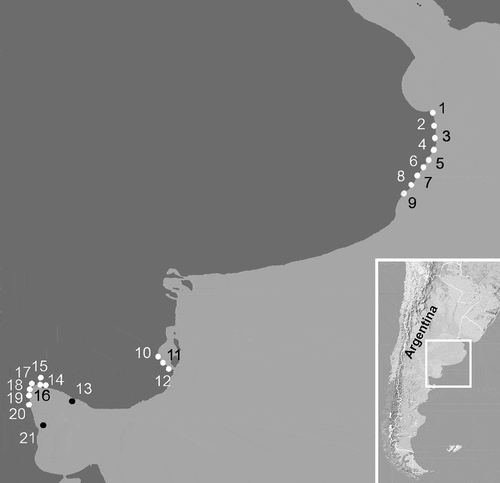
The material from Argentina was collected in three areas (), as follows. (1) At several stations along the northern coast of Buenos Aires Province, San Clemente del Tuyú, Santa Teresita, Mar del Tuyú, La Lucila del Mar, Mar de Ajó, Nueva Atlantis, Pinamar, Villa Gesell and Mar Azul, from November 1994 to September 2000 and from March 2008 to March 2013. (2) At several stations along the southern coast of Buenos Aires Province, Los Pocitos, Ría del Jabalí and Bahía San Blas, from May 2008 to March 2013. (3) At several stations in the northern area of San Matías Gulf (Río Negro Province): Punta Orengo, Las Garzas, Banco Reparo, San Antonio Oeste, Los Álamos, Las Grutas, El Sótano and El Fuerte, from April 1998 to May 2000 and from March 2006 to April 2007.
Fig. 1. Map of the marine littoral of Buenos Aires Province and North of Río Negro Province, showing sampling stations and location of the area in Argentina. 1: San Clemente del Tuyú, 2: Santa Teresita, 3: Mar del Tuyú, 4: La Lucila del Mar; 5: Mar de Ajó, 6: Nueva Atlantis, 7: Pinamar, 8: Villa Gesell, 9: Mar Azul, 10: Los Pocitos, 11: Ría del Jabalí, 12: Bahía San Blas, 13: Punta Orengo, 14: Las Garzas, 15: San Antonio Oeste, 16: Banco Reparo, 17: Los Álamos, 18: Las Grutas, 19: Piedras Coloradas, 20: El Sótano, 21: El Fuerte.
Qualitative samples were taken from the surface layer of the water column (between 0 and 5 m) with 30 μm net hauls and fixed with 4% formalin. In the laboratory, the preserved samples were rinsed with distilled water to remove salt and preservatives, and then the organic matter was oxidized according to Hasle & Fryxell (Citation1970) and Prygiel & Coste (Citation2000). The cleaned material was mounted for light and scanning electron microscopy after Ferrario et al. (Citation1995). Permanent mounts were made with Naphrax (Brunel Microscopes, Chippenham, UK).
The materials have been deposited in the Herbarium of the División Ficología ‘Dr. Sebastián A. Guarrera’, Facultad de Ciencias Naturales y Museo, Universidad Nacional de La Plata under the numbers LPC 4250–4495, LPC 4550–4643, LPC 11001–11353, LPC 11601–11908, LPC 13648–13685.
Observations were made with Leica DM 2500 (phase contrast and differential interference contrast; Leica Microsystems, Wetzlar, Germany), Zeiss Axiovert 40 CFL (phase contrast and Nomarski interference contrast; Zeiss Microimaging, Göttingen, Germany) and Nikon Microphot-FX microscopes. The photomicrographs were made with Leica DM 2500 and Zeiss Axiovert 40 CFL microscopes and Jeol JSMT 100 and Jeol JSM 6360 LV (JEOL, Tokyo, Japan) scanning electron microscopes.
The type slide of Pleurosigma elongatum, BM 23651, ‘Poole Bay, June 1849’, from the Natural History Museum, London, was examined with LM and photomicrographs were made with apochromatic objectives on Kodak TP 2415.
The type material of P. elongatum, from ‘Poole Bay, June 1849’, kept in the Van Heurck Collection, National Botanic Garden of Belgium, Brussels, was cleaned by conventional methods and photomicrographs were made with the Jeol JSM 6360LV. The photomicrographs are mostly reproduced at the same scale, to facilitate comparisons.
Terminology follows Ross et al. (Citation1979), Round et al. (Citation1990), Sterrenburg (Citation1991a, Citation1991b), Stidolph (Citation1992), Sterrenburg et al. (Citation2003) and Reid (Citation2012). The procedure for measurement of the intersection angle of the oblique striae and the raphe angle was described in Sterrenburg (Citation1991a); the term ‘calcar’ – a spur-like detail at the apex seen in LM and represented in SEM by an external fissure in the valve surface and/or particular internal details – was introduced by Sterrenburg in Stidolph (Citation1992).
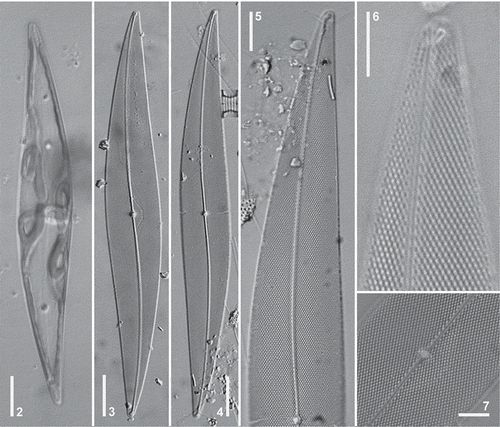
Figs 2–7. Pleurosigma hinzianum. Holotype (Figs 4–5); Paratype (Figs 3, 6–7). LM. , Cell showing ribbon-shaped chloroplast. , General appearance of valve. , Detail of valve apex. , Note terminal area unilaterally dilated. , Note terminal helictoglossa. , Detail of the central area of the valve. Note central nodule showing internal raphe ends. Scale bars: Figs 2–4 = 20 µm, Fig. 5 = 10 µm, Figs 6–7 = 5 µm.
Figs 8–17. Pleurosigma hinzianum. Type material from sample used to prepare the holotype slide. SEM. External views. , General appearance of the valve. , Valve apex showing straight terminal fissure straight, bent towards the concave side of the valve and the adjacent buttonhole-shaped opening (arrowhead). , Central part of the valve. Note apically elongated slit-like foramina. , Detail of the central area showing raphe fissures. Scale bars: Figs 8–9 = 20 µm, Fig. 14 = 5 µm, Figs 10–13, 15–17 = 2 µm.

Figs 18–24. Pleurosigma hinzianum. Type material from sample used to prepare the holotype slide. SEM. Internal views. , Valve showing eccentric raphe-sternum. , Valve apex showing polar endings finishing in loop-shaped helictoglossa. Note a buttonhole-shaped opening adjacent to the helictoglossa (arrowhead). , Central part of the valve showing central area short and raised central area with central bars surrounding the oval raphe nodule. Note pairs of C-shaped pores (arrowheads). , Detail of the central area. Note central coaxial raphe endings coaxial and pairs of C-shaped pores (arrowhead). , Elliptical, hymen-occluded pores crossed by a bar. Detail of paired C-shaped pores (arrowheads). Scale bars: Figs 18–19 = 20 µm, Fig. 22 = 5 µm, Figs 20–21, 23 = 2 µm.
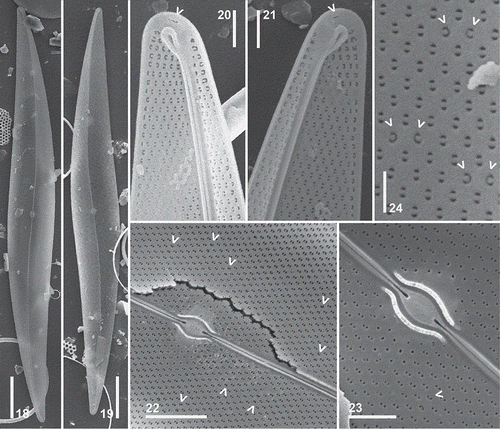
Table 1. Morphometric data obtained from specimens of Pleurosigma hinzianum and P. frenguellianum and from type material of P. amara and P. elongatum. Abbreviations: nd, no data; *, measured or observed from pictures in the literature.
Results
Pleurosigma hinzianum Sterrenburg, Sunesen & Sar, sp. nov. (–, )
Description in LM: Cell with one or two ribbon-shaped chloroplasts () sometimes convoluted at the valve centre and lying appressed to the girdle towards the poles. Valve lanceolate, slightly sigmoid, with acute apices (–), 181–206 µm long and 20–28 µm wide. Ratio length/width 7.0–8.7. Raphe-sternum eccentric, with single sigmoid curvature, closer to the convex sides towards the apices (–). Raphe angle +7° to +8° (–). Central area circular, small, with a thick barely bilaterally dilated central raphe nodule (–, ). Central internal raphe endings coaxial (). Terminal areas small, unilaterally dilated, funnel-shaped, in apical position (–), with helictoglossa twisted towards the concave side of the valve (). Striation dominated by the oblique striae, 19–22 in 10 µm, intersecting at an angle between 64° and 69° in the central part of the valve, and crossed by transverse striae, 22–24 in 10 µm (–).
Description in SEM: External view: Valve vaulted towards the apices, angled at the raphe (–). Raphe bordered by a narrow, inconspicuous, hyaline area (–). Central external raphe fissures deeply projecting into the central area, variable in undulation, either overlapping for some distance (–) or not overlapping (), and curving in opposite directions. Terminal raphe fissures straight, obliquely bent towards the concave side of the valve (–). Apices with a short, buttonhole-shaped opening, near the terminal part of the raphe fissure, on the concave side of the valve (–, arrowheads). Areolae arranged in decussate striae, opening by apically elongated slit-like foramina (–). Foramina around the central area shorter than in the remainder of the valve (, ).
Internal view: Raphe-sternum narrow, thickened (–). Central area short and raised, with slender central bars of approximately equal length, surrounding the oval raphe nodule (–). Central raphe endings coaxial, dilated in the proximal part (–). Polar raphe endings terminated in a well-developed, looped helictoglossae, turned towards the concave side of the valve (–). Apices with a buttonhole-shaped opening located near the helictoglossa (–, arrowheads). Areolae opening by elliptical, hymen-occluded pores crossed by siliceous bars except at the periphery of the central area (–). Pores paired, C-shaped, appearing scattered over the valve surface (–, arrowheads). Areolae bordering the raphe-sternum and the non-perforated valve margin not completely aligned (–).
This new species is named after Friedel Hinz of the Alfred Wegener Institute for Polar and Marine Research in Bremerhaven, Germany in recognition of her long-term assistance to the diatom research community.
Holotype: Slide LPC 4262 (1) here designated, labelled ‘holotipo de Pleurosigma hinzianum, Nueva Atlantis, 27/03/1995’ (deposited in the Herbarium of the División Ficología ‘Dr. Sebastián A. Guarrera’).
Isotype: Slide LPC 4262 (2) here designated, labelled ‘isotipo de Pleurosigma hinzianum, Nueva Atlantis, 27/03/1995’ (deposited in the Herbarium of the División Ficología ‘Dr. Sebastián A. Guarrera’).
Paratype: Slide LPC 4263 (2) here designated, labelled ‘paratipo de Pleurosigma hinzianum, Pinamar, 27/03/1995’ (deposited in the Herbarium of the División Ficología ‘Dr. Sebastián A. Guarrera’).
Type locality: Nueva Atlantis, 36°45′81′′S-56°40′04′′W, Buenos Aires Province, Argentina.
Distribution in the study area: Pleurosigma hinzianum occurred in samples collected at Nueva Atlantis 27 March 1995, Pinamar 27 March 1995, Villa Gesell 21 March 2011 and Santa Teresita 8 March 2013, in the Province of Buenos Aires. Pleurosigma hinzianum only appeared in samples collected in the late summer and early autumn.
Remarks: The combination of characters observed in the type material of the marine species Pleurosigma hinzianum can be summarized as follows: (1) valve lanceolate, slightly sigmoid with acute apices, (2) raphe-sternum eccentric, (3) raphe angle +7° to +8°, (4) angle of intersection of the oblique striae 64° to 69°, (5) terminal external raphe fissures straight, bent towards the concave side of the valve, (6) buttonhole-shaped opening adjacent to the raphe fissure, obliquely oriented towards the concave side of the valve, (7) apices with a buttonhole-shaped opening adjacent to the looped helictoglossa, (8) hymen-occluded pores crossed by a siliceous bar.
Pleurosigma frenguellianum Sunesen, Sterrenburg & Sar, sp. nov. (Figs 25–47, ).
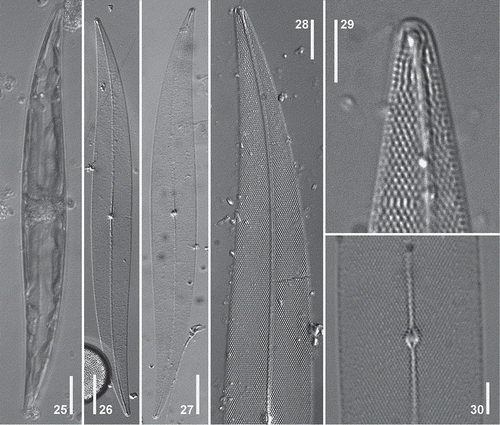
Figs 25–30. Pleurosigma frenguellianum. Holotype (, –). Material from Bahía San Blas 07/05/11 () and from Piedras Coloradas 05/26/06 (, ). LM. , Cell showing ribbon-shaped chloroplast. , General appearance of the valve. , Detail of valve showing striation pattern. , Note terminal unilaterally dilated area and change of striation pattern at valve apex. , Detail of the central area of the valve. Note central nodule showing internal raphe ends. Scale bars: – = 20 µm, Fig. 28 = 10 µm, Figs 29–30 = 5 µm.
Figs 31–39. Pleurosigma frenguellianum. Type material from sample used to prepare the holotype slide (, , ) and the paratype slide (, –, ). Material from Ría del Jabalí 09/29/08 () and from Nueva Atlantis 03/27/95 (). SEM. External views. , General appearance of the valve. , Valve apex showing hook-shaped terminal fissure, bent towards the concave side of the valve and two adjacent buttonhole-shaped openings (arrowheads). , Central part of the valve. Note apically elongated slit-like foramina. , Detail of the central area showing raphe fissures. Note that ends are more or less parallel, overlapping, spoon-shaped dilated. Scale bars: Figs 31–32 = 20 µm, Fig. 36 = 5 µm, Figs 33–35, 37–39 = 2 µm.

Figs 40–47. Pleurosigma frenguellianum sp. nov. Type material from sample used to prepare the holotype slide () and the paratype slide (, ). Material from Nueva Atlantis 03/27/95 (–, , ). SEM. Internal views. , Valve showing almost central raphe-sternum, sigmoid towards the apices. , Valve apex showing change of striation pattern and polar endings in helictoglossa. Note two isolated pores adjacent to the helictoglossa (arrowheads). , Central part of the valve showing short and raised central area with central bars surrounding the oval raphe nodule. Note pairs of C-shaped pores (arrowheads). , Detail of the central area. Note coaxial central raphe endings. , Elliptical, hymen-occluded pores crossed by a bar. Detail of paired C-shaped pores (arrowheads). Scale bars: Figs 40–41 = 20 µm, Fig. 45 = 5 µm, Figs 42–44, 46 = 2 µm, Fig. 47 = 1 µm.
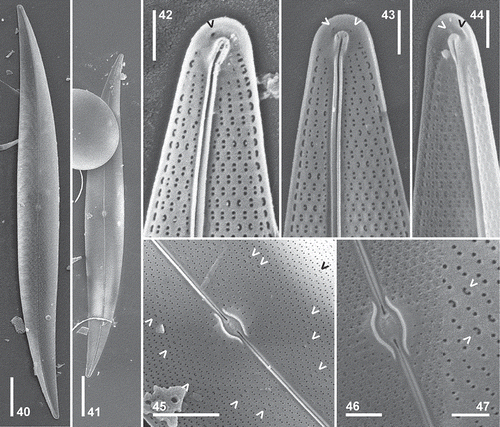
Description in LM: Cell with one or two ribbon-shaped chloroplasts (Fig. 25) sometimes convoluted at the valve centre and lying appressed to the girdle towards the poles. Valve linear-lanceolate, slightly sigmoid, with acute apices (–), 161–289 µm long and 20–29 µm wide. Ratio length/width 8.2–11.2. Raphe-sternum almost central, sigmoid towards the apices approaching the convex side of the valve. Raphe angle +6° to +7° (–). Central area oval, small, with a thick barely bilaterally dilated central raphe nodule (–, ). Central internal raphe endings dilated, coaxial (). Terminal areas small, unilaterally dilated, funnel-shaped, in apical position (–), with helictoglossa slightly twisted towards the concave side of the valve (). Striation pattern dominated by the oblique striae, 18–20 in 10 µm, intersecting at an angle between 53° and 58° in the central part of the valve, and crossed by transverse striae, 20–22 in 10 µm (, ). At the valve apex the stria orientation changing from the 3-system (sensu Pleurosigma) to the 2-system (sensu Gyrosigma) (–) as in P. angulatum.
Description in SEM: External view: Valves slightly vaulted near the apices (–). Raphe bordered by a narrow, inconspicuous, hyaline area (). Central external raphe fissures deeply projecting into the central area, undulating in the same direction, with the ends more or less parallel, overlapping, and spoon-shaped dilated (–). Terminal raphe fissures hook-shaped, obliquely bent towards the concave side of the valve. Apices with two short, diagonally arranged buttonhole-shaped openings near the terminal part of the raphe fissure (–, arrowheads). Areolae arranged in decussate striae, opening by apically elongated slit-like foramina (–). Areolae bordering the non-perforated valve margin perfectly aligned (–, ).
Internal view: Raphe-sternum narrow, thickened (–). Central area short and raised, with slender central bars of similar length, surrounding the oval raphe nodule (–). Central raphe endings coaxial, dilated in the proximal part (–). Polar endings terminated in a well-developed helictoglossa with parallel lips, slightly turned towards the concave side of the valve (–). Apices with two isolated pores, one on each side of the helictoglossa (–, arrowheads). Areolae opening by elliptical, hymen-occluded pores crossed by siliceous bars (–, –) except at the periphery of the central area (–). Pores paired, C-shaped, appearing scattered over the valve surface (, , arrowheads). Areolae bordering the non-perforated valve margin completely aligned and C-shaped (–).
This new species is named after Dr Joaquín Frenguelli of the Museo de La Plata, Argentina, who started the study of diatoms in Argentina in the early 20th century.
Holotype: Slide LPC 11220 (1) here designated, labelled ‘holotipo de Pleurosigma frenguellianum, Villa Gesell, 02/07/2010’ (deposited in the Herbarium of the División Ficología ‘Dr. Sebastián A. Guarrera’).
Isotype: Slide LPC 11220 (2) here designated, labelled ‘isotipo de Pleurosigma frenguellianum, Villa Gesell, 02/07/2010’ (deposited in the Herbarium of the División Ficología ‘Dr. Sebastián A. Guarrera’).
Paratype: Slide LPC 11221 (1) here designated, labelled ‘paratipo de Pleurosigma frenguellianum, Mar Azul, 02/07/2010’ (deposited in the Herbarium of the División Ficología ‘Dr. Sebastián A. Guarrera’).
Type locality: Villa Gesell, 37°16′50,70′′S–56°58′53′′W, Buenos Aires Province, Argentina.
Distribution in the study area: Pleurosigma frenguellianum occurred in samples collected in Santa Teresita 27 March 1995; Nueva Atlantis 27 March 1998; Villa Gesell 2 February 2010, 22 March 2011; Mar Azul 26 March 2010, 2 July 2010, 8 March 2013; Ría del Jabalí 29 September 2008 and Bahía San Blas 29 September 2010, 5 July 2011, 11 October 2011, 18 March 2013 in the Buenos Aires Province and in Piedras Coloradas 26 March 2006, Gulf of San Matías, Río Negro Province. Pleurosigma frenguellianum shows a wider distribution in space and time than P. hinzianum appearing between late summer and spring.
Remarks: The combination of characters observed in the type material of the marine species Pleurosigma frenguellianum can be summarized as follows: (1) valve linear-lanceolate, slightly sigmoid, with acute apices, (2) raphe-sternum almost central, sigmoid towards the apices, (3) raphe angle +6° to +7°, (4) angle of intersection of the oblique striae 53° to 58°, (5) stria pattern changing in orientation from 3-system to 2-system near the valve apex, (6) ends of the central external raphe fissures more or less parallel, overlapping, spoon-shaped dilated, (7) terminal external raphe fissures hook-shaped, (8) two diagonally arranged buttonhole-shaped openings adjacent to the raphe fissure, (9) two isolated pores adjacent to the helictoglossa, which has parallel lips, (10) areolae bordering the non-perforated valve margin perfectly aligned, internally C-shaped, (11) hymen-occluded pores crossed by a siliceous bar.
Analysis of type material of Pleurosigma elongatum W. Smith (–, )
Figs 48–55. Pleurosigma elongatum Smith. Type material from ‘Poole Bay, June 1849’, kept in Van Heurck Collection (Figs 48–55). –, LM. Figs 51–55, SEM. , General appearance of the valve. , Detail of the central part of the valve. , Detail of valve apex showing calcar (arrowhead). , Detail of the central area of the valve in external view showing W-shaped raphe fissures. , Detail of valve apex in internal view showing helictoglossa bordered by a series of rosette-shaped clustered pores and one apical rosette (arrowhead). , Detail of the elliptical hymen-occluded internal pores, some of which are bisected by a bar. , Detail of the central area of the valve in internal view. Note circular hymen-occluded pores not bisected by a bar. Scale bars: Fig. 48 = 20 µm, – = 10 µm, – = 2 µm.

The type material was examined in LM and SEM to obtain more data on the morphology, such as the internal morphology of the areolae and the ultrastructural equivalent of the ‘calcar’ seen in LM. Specimens of the type material pictured by Reid (2002, , ) show the areolar pores adjacent to the raphe-sternum crossed by bars whilst elsewhere they do not show bars. In Sterrenburg et al. (Citation2005), it was shown that such bars develop in the last stage of valve formation, but not necessarily in all pores.
A typical specimen in the type material, on slide BM 23651, is shown in –. The valve is slender lanceolate, moderately sigmoid, gradually tapering and vaulted towards the ends, with somewhat acute apices, 200 µm long, 20 µm wide. The raphe-sternum is almost straight for most of its length (–), becoming moderately sigmoid and eccentric near the apices (). The terminal areas are small, unilaterally dilated, funnel-shaped, in apical position (). A calcar is clearly visible in LM as a spur-like shadow at the apex (, arrowhead). Raphe angle +5°. The central area is small, oval and symmetric (). The transverse striae number 20–22 in 10 µm, the oblique 18–20 in 10 µm. The angle of intersection of the oblique striae is 63°–65°.
SEM study of the type material in the Van Heurck Collection yielded three valves, an incomplete one in external view and two in internal view. The central external raphe fissures are twice deflected to the same side, becoming W-shaped, ending at the same level in the central area (). The loculate areolae are arranged in decussate striae, opening by apically elongated slit-like foramina, shorter around the central area than elsewhere (). The raphe-sternum is internally thickened (–). The central area is small and raised, with central bars of approximately equal length surrounding the oval raphe nodule () and central internal raphe endings coaxial, dilated as small triangles (). The terminal internal raphe ends in helictoglossae turned towards the concave side of the valve. On the concave side, they are bordered by a series of rosette-shaped clustered pores (–, arrowheads), which form the SEM equivalent of the ‘calcar’ seen in LM. There is a cluster of 2–4 small pores adjacent to the helictoglossa (–, arrows). The areolae open internally by hymen-occluded pores, roundish only in a small area around the central nodule () and elliptical in the rest of the valve (–). The hymen-occluded pores in the central part are not crossed by a bar, whilst a row of pores adjacent to the raphe-sternum does show such narrow silica bars and the rest of the valve shows pores with, as well as others without, bars (–).
Records from Argentina: Pleurosigma elongatum was recorded from Buenos Aires coastal waters by Müller Melchers (Citation1959) and from continental waters of Argentina by Solari & Claps Citation(1984) (Arroyo San Clemente, Buenos Aires Province), Romero (Citation1993) (Río Colorado, Province of La Pampa), Vouilloud & Leonardi (Citation2001) (in artificial channels, Buenos Aires Province) and Rodríguez et al. (Citation2006) (epixylic in a section of the lower river Luján and a tributary, Buenos Aires Province), without morphological analysis or illustrations. Maidana (Citation1994) determined material from the Quaternary sediments of the Salinas del Bebedero as P. elongatum, but the dimensions given and the valve outline of the specimens drawn do not match those of P. elongatum or of the new species described here. Thus we are not able to confirm the presence of P. elongatum in Argentina.
Remarks: The combination of characters observed in the type material of the marine species Pleurosigma elongatum can be summarized as follows: (1) valve linear-lanceolate, slightly sigmoid, with acute apices, (2) calcar present, (3) raphe-sternum almost central, sigmoid towards the apices, (4) raphe angle +4° to +5°, (5) angle of intersection of the oblique striae 63°–68° (including data of Reid, Citation2002), (6) central external raphe fissures W-shaped, (7) helictoglossae turned towards the concave side of the valve and laterally bordered by a series of rosette-shaped clustered pores, (8) hymen-occluded pores of the areolae may be bisected by a bar.
Discussion
In Sterrenburg et al. (Citation2003) it was pointed out that Pleurosigma species have a fully functional raphe, allowing active movement across and through substrata, which would serve no useful purpose for a planktonic diatom. The presence of Pleurosigma species and other raphe-bearing diatoms in the plankton can be attributed to three causes. Firstly, in shallow turbulent waters such as the localities examined here, benthic diatoms are easily dislodged from the substratum and suspended in the water column. Secondly, long-established epiphyton readily detaches from macrophytes to enter the water column. The samples studied here also contain raphe-bearing diatoms of the typically epiphytic genus Cocconeis. Thirdly, even in stagnant pools sediments rich in benthic diatoms may become suspended and dispersed as a result of photosynthetic oxygen production. A planktonic occurrence does not necessarily equate to a planktonic mode of life; such an incidental presence has been characterized as tychoplanktonic.
In the LM, Pleurosigma hinzianum Sterrenburg, Sunesen & Sar sp. nov. (–) resembles Pleurosigma frenguellianum Sunesen, Sterrenburg & Sar sp. nov. (–) in general appearance and dimensions (see ). Both species show subtle differences in the valve outline (more lanceolate in P. hinzianum and more linear in P. frenguellianum), and in the raphe-sternum path (more eccentric in the middle part of the valve in P. hinzianum and almost central in P. frenguellianum). The principal LM features separating P. hinzianum and P. frenguellianum are the angle of the oblique striae in the central part of the valve (64°–69° in the former and 53°–58° in the latter) and the orientation of the striae at the valve apex, in 3-systems in P. hinzianum and in 2-systems in P. frenguellianum.
In the SEM, these species show several important differences in the external central raphe fissures and in the external and internal apical structure. The overlapping part of the central external raphe fissures is not dilated in P. hinzianum (–) whereas it is dilated and spoon-shaped in P. frenguellianum (–). The apices in external view differ in the morphology of the terminal raphe fissures: straight, oblique, with an adjacent buttonhole-shaped opening in P. hinzianum (–) and hook-shaped, with two adjacent diagonally arranged buttonhole-shaped openings in P. frenguellianum (–). In internal view, the helictoglossa is accompanied by one buttonhole-shaped opening in P. hinzianum (–, arrowheads) and by two isolated pores in P. frenguellianum (–, arrowheads).
Pleurosigma hinzianum (–) resembles P. amara Stidolph (see – in Stidolph Citation1992) in general appearance, but shows subtle differences in the dimensions of the valves, stria density, angle of intersection of the oblique striae and raphe angle. Pleurosigma frenguellianum (–) resembles P. amara (Stidolph Citation1992, –) in the path of the raphe-sternum and in the raphe angle but differs in valve outline, in the angle of intersection of the oblique striae () and in the orientation of the striae at the valve apex. The ranges of valve dimensions and stria density are not useful for separating P. hinzianum and P. frenguellianum from P. amara. More useful LM features are: the path of the raphe-sternum, which is sigmoid in P. hinzianum (–) but almost central in the middle of the valve becoming sigmoid only towards the apices in P. amara (Stidolph Citation1992, ). The calcar is present in P. amara (Stidolph Citation1992, –) but absent in both P. hinzianum (–) and P. frenguellianum (–). Although some characters separate P. hinzianum and P. frenguellianum from P. amara in the LM, the main characters for differentiation are ultrastructural, principally related to details of the external and internal apical structure. The terminal external raphe fissures are straight, obliquely bent towards the concave side of the valve in P. hinzianum (–), hook-shaped and obliquely bent towards the concave side in P. frenguellianum (–) and broadly curved over the valve apex in P. amara (Stidolph Citation1992, figs 64–65). Additionally, there is a small buttonhole-shaped opening adjacent to the raphe fissure towards the concave side of the valve in P. hinzianum, two diagonally arranged buttonhole-shaped openings adjacent to the raphe fissure in P. frenguellianum and two terminal long and slender grooves parallel to the valve margin in P. amara.
The internal apical view shows modified slit-like areolae in the proximity of the helictoglossa in P. amara (see figs 68–69 in Stidolph Citation1992) while the areolae adjacent to the helictoglossa are similar to those at the valve margin in P. hinzianum () and P. frenguellianum (). Another useful feature for separating P. frenguellianum and P. amara is the morphology of the end of the central external raphe fissures: generally parallel, constantly overlapping and dilated in the former () and markedly variable without overlapping or dilatation in the latter (see fig. 76 A–G in Stidolph, Citation1992).
Another species that looks similar to the new species described here is P. elongatum. The type material of P. elongatum has been previously analysed and some ultrastructural information was added by Reid (2002, –). Our specimens of P. elongatum () perfectly match those shown by Smith (1852, pl. 1, ; see also Smith, Citation1853, pl. 20, fig. 199) and by Reid (2002, –, ) in valve outline, path of the raphe-sternum, angle of intersection of the oblique striae and raphe angle (see ). The transverse striae are slightly denser than the oblique in our specimens and in those shown and described in Reid (2002, , 26) ().
Ultrastructural details of the internal central area () match those shown in Reid (2002, ) as regards outline, extension and morphology of the internal raphe ends, central bars around the nodule and hymen-occluded pores. The central external raphe fissures, which have not been previously shown in the type material, are W-shaped in the specimen retrieved () and resemble those pictured by Stidolph (1992, fig. 79) for the Sterrenburg Collection sample from Robbenoordbos, North Holland, material. However, as is evident from the various shapes seen in P. hinzianum, the external central raphe fissure shape is not always a constant character, as pointed out in Sterrenburg Citation(1990). The apex of the specimen pictured by Reid (2002, ) in internal view is apparently eroded, with an atypical areola pattern that might be interpreted as an early stage of the valve morphogenesis (Sterrenburg et al. Citation2005). Specimens analysed in the present study are not eroded and show apical details that could be considered consistent with those described by Reid (2002, ), the helictoglossae are laterally bordered by a series of rosette-shaped clustered pores including a terminal one (, arrowheads). The type specimens examined and the specimens from Robbenoordbos examined by Stidolph (1992, figs 80–82) showed the same aspect of internal occluded pores: circular, single, without bars, in the middle part of the valve and elliptical, bisected by a bar in the vicinity of the raphe-sternum. Additionally, the material of the present study () and that of Stidolph (1992, fig. 81) show bisected pores alternating with elliptical single pores over the valve. It is unlikely that all valves are in the early stages of morphogenesis, thus it is probable that the internal pores are more morphologically variable than claimed for this genus (see Round et al., Citation1990).
The LM comparison between Pleurosigma frenguellianum () and P. elongatum () shows some similarities in valve outline, path of the raphe-sternum and stria density. Nevertheless, both species can be separated by the angle of intersection of the oblique striae (53°–58° in the former and 63°–68° in the latter), the raphe angle (+6°/+7° in the former and +4°/+5° in the latter) and the pattern of striation at the apex. Pleurosigma hinzianum () differs from P. elongatum in valve outline, path of the raphe-sternum and raphe angle but it is similar in the angle of intersection of the oblique striae (). A calcar is present in P. elongatum () and absent in both P. hinzianum () and P. frenguellianum (). The comparison of these species in SEM reveals differences in details of the internal apical structure: there is a series of rosette-shaped clustered pores laterally to the helictoglossa with a cluster at the apex in P. elongatum (), whereas the areolae adjacent to the helictoglossa are similar to those at the valve margin, with one isolated apical pore in P. hinzianum () and with two isolated apical pores in P. frenguellianum (). In addition, in the new species, all the internal hymen-occluded pores except for a few surrounding the central area are crossed by a bar, unlike the situation in P. elongatum.
The main LM features of the species discussed are the valve outline, the path of the raphe-sternum, the raphe angle, the angle of intersection of the oblique striae and the striation pattern at the apex. In SEM, the external central and terminal raphe fissures, internal details of the central area, external and internal apical details and morphology of the hymen-occluded pores are important characters. The results of the present study, like those of Stidolph (Citation1992) and Sar et al. (2012, 2013) for instance, demonstrate that detailed electron microscopic studies are necessary for the description of new species in the genus Pleurosigma and re-examination of the type material of similar species is also necessary to avoid potential taxonomic confusion.
Acknowledgements
The electron photomicrographs of the type material of Pleurosigma elongatum in the Van Heurck Collection at the National Botanic Garden of Belgium, Brussels, were made available to us by Prof. Bart Van de Vijver and Mrs Myriam de Haan whose assistance is gratefully acknowledged.
This work was supported by Universidad Nacional de La Plata [grant number 11/N640] and by Consejo Nacional de Investigaciones Científicas y Técnicas [grant number PIP0067].
References
- Cardinal, A., Poulin, M. & Bérard-Therriault, L. (1989). New criteria for species characterization in the genera Donkinia, Gyrosigma and Pleurosigma (Naviculaceae, Bacillariophyceae). Phycologia, 28: 15−27.
- Ferrario, M.E., Sar, E.A. & Sala, S.E. (1995). Metodología básica para el estudio del fitoplancton con especial referencia a las diatomeas. In Manual de Métodos Ficológicos (Alveal, K., Ferrario, M.E., Oliveira, E.C. & Sar, E., editors), 1–23. Universidad de Concepción, Concepción.
- Hasle, G.R. & Fryxell, G.A. (1970). Diatoms: cleaning and mounting for light and electron microscopy. Transactions of the American Microscopical Society, 89: 469–474.
- Hendey, N.I. (1964). An Introductory Account of the Smaller Algae of British Coastal Waters. V. Bacillariophyceae (Diatoms). Her Majesty’s Stationery Office, London. 317 pp.
- Maidana, N.I. (1994). Fossil diatoms from Salinas del Bebedero (San Luis, Argentina). Diatom Research, 9: 99–119.
- Müller Melchers, F.C. (1959). Plankton diatoms of the Southern Atlantic Argentine and Uruguay coasts. Comunicaciones Botánicas del Museo de Historia Natural deMontevideo, 3: 1−53.
- Prygiel, J. & Coste, M. (2000). Guide Méthodologique pour la mise en ouvre de l´Indice Biologique Diatomées. Agences de l’Eau, Ministère de l’Aménagement du Territoire et de l’Environnement, Direction de l’Eau & CEMAGREF, France. 134 pp.
- Reid, G. (2002). Four new species of Pleurosigma (Bacillariophyta) from Alexandria, Egypt. Botanical Journal of the Linnean Society, 140: 77−92.
- Reid, G. (2012). A revision of the family Pleurosigmataceae (Bacillariophyta). Diatom Monographs 14. A.R.G. Gantner, Ruggell, Liechtenstein.
- Rodríguez, P.L., Pizarro, H., Dos Santos Afonso, M., Maidana, N. & Bonaventura, S.M. (2006). Epixylic algae from a polluted lowland river of Buenos Aires Province (Argentina). Cryptogamie, Algologie, 27: 63−83.
- Romero, O.E. (1993). Diatomeas (Bacillariophyceae) de la Provincia de La Pampa (Argentina). I. Darwiniana, 32: 303−313.
- Ross, R., Cox, E.J., Karayeva, N.I., Mann, D.G., Paddock, T.B.B., Simonsen, R. & Sims, P.A. (1979). An amended terminology for the siliceous components of the diatom cell. In Proceedings of the Fourth Symposium on Recent and Fossil Marine Diatoms (Simonsen, R., editor). Nova Hedwigia, Beiheft, 54: 513–533.
- Round, F.E., Crawford, R. & Mann, D.G. (1990). The Diatoms. Morphology and Biology of the Genera. Cambridge University Press, Cambridge. 747 pp.
- Sar, E.A., Hinz, F., Sterrenburg, F.A.S., Lavigne, A.S., Lofeudo, S. & Sunesen, I. (2012). Species of Pleurosigma (Pleurosigmataceae) with lanceolate or slightly sigmoid valve outline. Analysis of type materials. Diatom Research, 27: 237–253.
- Sar, E.A., Sterrenburg, F.A.S., Lavigne, A.S. & Sunesen, I. (2013). Diatomeas de ambientes marinos costeros de Argentina. Especies del género Pleurosigma (Pleurosigmataceae). Boletín de la Sociedad Argentina de Botánica, 48: 17–51.
- Smith, W. (1852). Notes on the Diatomaceae, with descriptions of British species included in the genus Pleurosigma. Annals and Magazine of Natural History, 9, 2: 1–12.
- Smith, W. (1853). A Synopsis of the British Diatomaceae; with Remarks on their Structure, Functions and Distribution; and Instructions for Collecting and Preserving Specimens. Vol. 1. 89 pp., pls. 1–32. John Van Voorst, London.
- Solari, L.C. & Claps, M.C. (1984). El fitoplancton del arroyo San Clemente (Bahía Samborombón). Revista del Museo de La Plata, 13, Bot. 76: 149−164.
- Sterrenburg, F.A.S. (1990). Studies on the genera Gyrosigma and Pleurosigma (Bacillariophyceae). A new phenomenon: co-existence of dissimilar raphe structures in populations of several species. In Ouvrage dedié à la Mémoire du Professeur Henry Germain (1903–1989) (Ricard, M., editor), 235–242. Koeltz, Koenigstein.
- Sterrenburg, F.A.S. (1991a). Studies on the genera Gyrosigma and Pleurosigma (Bacillariophyceae). Light-microscopical criteria for taxonomy. Diatom Research, 6: 367–389.
- Sterrenburg, F.A.S. (1991b). Studies on the Genera Gyrosigma and Pleurosigma (Bacillariophyceae). The typus generis of Pleurosigma, some presumed varieties and imitative species. Botanica Marina, 34: 561–573.
- Sterrenburg, F.A.S., Meave Del Castillo, M.E. & Tiffany, M.A. (2003). Studies on the genera Gyrosigma and Pleurosigma (Bacillariophyceae): Pleurosigma species in the plankton from the Pacific coast of Mexico, with the description of P. gracilitatis sp. nov. Cryptogamie, Algologie, 24: 291–306.
- Sterrenburg, F.A.S., Tiffany, M.A. & Meave Del Castillo, M.E. (2005). Valve morphogenesis in the diatom genus Pleurosigma W. Smith (Bacillariophyceae) – Nature’s alternative sandwich. Journal of Nanoscience and Nanotechnology, 5: 140–145.
- Stidolph, S.R. (1992). Observations and remarks on the morphology and taxonomy of the diatom genera Gyrosigma Hassal and Pleurosigma W. Smith. III. Gyrosigma sterrenburgii sp. nov. and Pleurosigma amara sp. nov. Diatom Research, 7: 345–366.
- Sunesen, I., Sterrenburg, F.A.S. & Sar, E.A. (2013). Pleurosigma guarreranum, sp. nov. (Pleurosigmataceae, Bacillariophyta), from the Gulf of San Matías, Argentina and comparison with the allied species Pleurosigma exemptum and P. obesum. European Journal of Phycology, 48: 451–460.
- Vouilloud, A.A. & Leonardi, P.I. (2001). Bacillariophyceae de los canales de Corfo (Río Colorado, Buenos Aires, Argentina). Boletín de la Sociedad Argentina de Botánica, 36: 229–241.