Abstract
This study was carried out to evaluate the dietary valine requirement by measuring means of the plasma free valine and ammonia concentrations in rainbow trout, Oncorhynchus mykiss, after dorsal aorta cannulation. A total of 35 fish averaging 498±7.2 g (initial body weight, mean±SD) were randomly distributed into seven groups with five fish in each group. After 48 h of feed deprivation, each group was fed one of seven L-amino acid based diets containing graded levels of valine (0.45, 0.95, 1.20, 1.45, 1.70, 1.95 or 2.45% of diet, dry matter basis) by intubation at 1% body weight. Blood samples were taken at 0, 5 and 24 h after intubation. Post-prandial plasma free valine concentrations (PPval, 5 h after intubation) and post-absorptive free valine concentrations (PAval, 24 h after intubation) of fish fed diets containing 1.45% or more valine were significantly higher than those of fish fed diets containing 1.20% or less valine (P<0.05). Post-prandial plasma ammonia concentrations (PPA, 5 h after intubation) remained low or constant from fish fed diets containing 0.45–1.45% valine, but increased linearly from fish fed diets containing 1.45–2.45% valine. Post-absorptive plasma ammonia concentrations in the present study were not significantly different among the groups. Broken-line model analyses on PPval, PAval and PPA indicated that the dietary valine requirements of rainbow trout was between 1.41 (3.85) and 1.50% (4.10) of diet (% of dietary protein on a dry matter basis).
1. Introduction
Valine is a branched-chain amino acid along with isoleucine and leucine, which are not synthesised in animals; so it must be ingested, usually as valine or valine containing proteins. Valine plays very important roles in certain biochemical reactions and in the growth of monogastric and preruminant terrestrial animals. It is involved in protein synthesis, synthesis of the amine neurotransmitters serotonin and the catecholamines dopamine and norepinephrine, which are derived from the aromatic amino acids tryptophan, phenylalanine, and tyrosine, the production of energy and the compartmentalisation of glutamate (Fernstrom Citation2005). Valine deficiency resulted in poor feed conversion and growth recession in some fish species such as catla, Catla catla (Ravi and Devaraj Citation1991), milkfish, Chanos chanos (Borlongan and Coloso Citation1993) and Indian major carp, Labeo rohita (Murthy and Varghese Citation1997). So, it is important to determine the valine requirements of cultured fish species for normal growth and feed utilisation. Dietary valine requirements have been estimated for commonly cultured species of fish, and wide variations of 2.5–4.0% of dietary protein have been reported (Wilson Citation2002).
The above requirements were determined by feeding trials. Feeding trials are expensive and last for several weeks or months before appreciable responses are observed. Determination of amino acid requirements by measuring plasma amino acid and ammonia concentrations on the other hand lasts for just a week, with insignificant labour and material costs. This method has been employed in several species and results are similar to those obtained by feeding trials (Cowey Citation1995; Ok Citation2002). Hence, it is advisable that this procedure be confirmed in important species so that they could be employed in determination of yet undetermined amino acid or other nutrient requirements in those species.
Plasma free amino acid (PFAA) concentrations were used to investigate amino acid metabolism and to evaluate the quality of dietary protein in rainbow trout, Oncorhynchus mykiss (Nose Citation1972; Walton and Wilson Citation1986; Schuhmacher et al. Citation1997), carp, Cyprinus carpio (Plakas et al. Citation1980; Dabrowski Citation1982) and Atlantic salmon, Salmo salar (Espe et al. Citation1993; Sunde et al. Citation2003). Evaluation of PFAA concentrations has led to the discovery of genetic defects of amino acid metabolism as a result of primary renal or liver disease and the effects of amino acid deficiencies, imbalances and toxicities on amino acid metabolism (Park et al. Citation2005). The relationships between the concentration of PFAA and intake of dietary amino acids have been investigated by several researchers (Plakas et al. Citation1980; Murai et al. Citation1987; Bai et al. Citation2003). PFAA of fish fed graded levels of the amino acid tested has been used in several experiments in an attempt to confirm requirement values assessed by growth. Although the effects of dietary protein or amino acid mixtures on PFAA concentrations in rainbow trout, O. mykiss (Murai et al. Citation1987; Schuhmacher et al. Citation1997; Vermeirssen et al. Citation1997), and sea bass, Dicentrarchus labrax (Thebault et al. Citation1985), have been reported, the complete dose response relationships for amino acids have not been investigated. In our previous studies, requirements for dietary essential amino acids such as arginine, methionine, lysine and threonine were obtained in rainbow trout, O. mykiss, by using surgically modified methods in our laboratory (Ok et al. Citation2001; Ok Citation2002; Park et al. Citation2005).
As a part of our studies on essential amino acid requirements of rainbow trout, the purpose of the present study was to estimate the dietary valine requirement based on the plasma free valine and ammonia concentrations in dorsal aorta cannulated rainbow trout, O. mykiss.
2. Materials and methods
2.1. Experimental fish
Rainbow trout, O. mykiss, averaging 493±7.2 g (initial body weight, mean±SD) were obtained from Ewhajung trout farm in Sangju, Korea. Seven net cages (1.3×1.3×1.3 m) were placed in a flow-through concrete raceway with a water flow rate of 60 L min−1. Fish were then fed a commercial rainbow trout diet (Woo-sung Feed Co. Ltd., Daejon, Korea) for 72 h until the fish recovered from the dorsal aorta cannulation operation. Supplemental aeration was also provided to maintain the dissolved oxygen at 6.7±0.5 mg L−1, and water temperature was maintained at 15±0.3°C.
2.2. Preparation of experimental diets
A basal diet containing 36.6% crude protein (29.6% crystalline amino acids mixture, 5% casein and 2% gelatin) was formulated by the modification of Kim (1997). Seven experimental diets were formulated to contain graded levels of valine (0.45, 0.95, 1.20, 1.45, 1.70, 1.95 or 2.45% of diet, on a dry matter basis) based on the basal diet. Equal amounts of aspartic acid and glutamic acid by weight were substituted for the graded amounts of valine in the basal diet to maintain the seven experimental diets isonitrogenous. Formulation and amino acid composition of the experimental diet are shown in and , respectively. The ingredient mixtures without oil were stored at –80°C until used and the diets were prepared by adding fish oil (10% of diet) and water (diet plus 0.4 parts of distilled water per diet) before intubation.
Table 1. Composition of the experimental diets (% of dry matter basis).a
Table 2. Amino acid composition of the experimental diet (% of dry matter basis).
Table 3. Plasma post-prandial (5 h after intubation) and post-absorptive (24 h after intubation) valine and ammonia concentrations (nmol mL−1) of rainbow trout fed graded levels of dietary valine.*
2.3. Dorsal aorta cannulation and intubation (force-feeding)
Rainbow trout were anaesthetised with 200 mg L−1 3-aminobenzoic acid ethyl ester methanesulfonate (MS-222; Sigma Chemical Company, St. Louis, MO, USA) for 3–5 min, placed on a V-shape table, and their gills were irrigated continuously with 16°C water containing 100 mg L−1 of MS-222 during the operation. A 50-cm-long cannula (Clay Adams PE 50 tubing, Parsippany, NJ, USA) with a bubble of about 5–6 cm on one end was washed with the heparinized Cortland saline solution; a 13-gauge needle was used to pierce a hole on the right nostrum of fish (ventral side up) as an exit for the cannula. A 19-gauge needle was used to bore a small hole in the roof of the mouth at the mid-line behind the third gill arc at a 30° angle, and a piano wire was inserted into the PE 50 tubing as a guide. The proper insertion was verified by the observation of a slow blood flow after the wire was withdrawn from the cannula. A 3 mL syringe with a 23-gauge needle was used to remove air and blood clot, and the cannula was flushed with the heparinized saline solution. The cannula was sutured behind the bubble on the roof of the mouth, led out from the right nostrum, plugged with a colour head pin, and sutured at the dorsal fin (Ok et al. Citation2001; Bai et al. Citation2003; Park et al. Citation2005). Thirty five dorsal aorta cannulated rainbow trout were randomly distributed into seven groups of five fish per group and fed one of the seven experimental diets per group. After 24 h feed deprivation, the fish were anaesthetized with 200 mg L−1 of MS-222 and fed the experimental diet by the stomach intubation method using a 3 ml syringe.
2.4. Sample collection and analysis
Five fish per group were anaesthetised with 200 mg L−1 MS-222 and blood was sampled using a 3 ml syringe from each fish at 0, 5 and 24 h after intubation of the experimental diets. Plasma samples were prepared by centrifugation at 3000 ×g for 10 min at room temperature. For deproteinization, plasma samples were mixed with a 10% 5-sulphosalicylic acid solution in the ratio of 4:1 (v/v), cooled on ice for 30 min and re-centrifuged. The protein-free supernatant was dissolved in pH 2.2 lithium citrate sample dilution buffer in the ratio of 1:1 (v/v), and the samples were stored at –80°C until analysis. PFAAs were quantified using an S433 amino acid analyzer (Sykam, Gilching, Germany) using the ninhydrin method. Plasma ammonia concentrations were analysed using the Berthelot reaction (Sigma).
2.5. Statistical analysis
Data were subjected to one way analysis of variance test using Statistix 3.1 (Analytical Software, St Paul, MN, USA). When a significant treatment effect was observed, a Least Significant Difference test was used to compare means. Treatment effects were considered significant at P<0.05. The breakpoints for post-prandial plasma free valine concentrations (PPval), post-absorptive free valine concentrations (PAval) and post-prandial plasma ammonia concentrations (PPA) were estimated by using the broken line model of Robbins et al. (1979).
3. Results
3.1. Plasma free valine concentrations
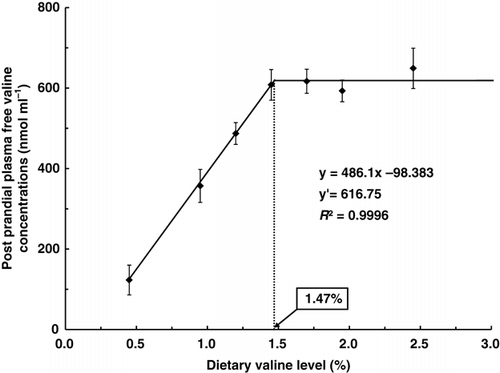
Results of PPval (5 h after intubation) and PAval (24 h after intubation) in dorsal aorta cannulated rainbow trout force-fed with diets containing seven graded levels of valine are summarised in . PAval of fish fed diets containing 1.45% or more valine were significantly higher than those of fish fed diets containing 1.20% or less valine (P<0.05). There were no significant differences in PPval among fish fed diets containing 1.45% or more valine (P>0.05). Broken-line model analyses on the basis of PPval as shown in indicated that the dietary valine requirement of trout could be 1.47% (4.02) of diet (% dietary protein on a dry matter basis).
Figure 1. Broken line analysis of post-prandial plasma valine concentrations (nmol mL−1) in rainbow trout fed graded levels of dietary valine. Values of the X-axis are the valine levels in the experimental diets. Values are means±SD of five replicates. Y=486.1x −98.383, Y′=616.75, R 2=0.9996.
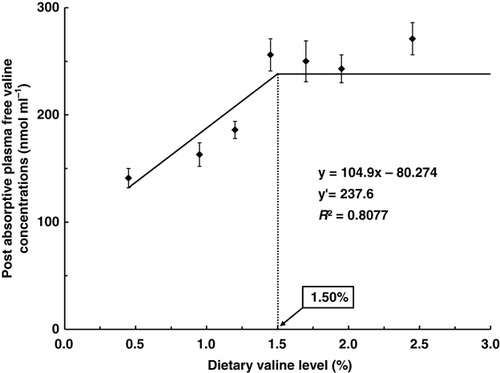
Post-absorptive plasma free valine concentrations of fish fed diets containing 1.45% or more valine were significantly higher than those of fish fed diets containing 1.20% or less valine (P<0.05). Post-absorptive plasma free valine concentration of fish fed the diet containing 2.45% valine was significantly higher than that of fish fed the diet containing 1.95% valine (P<0.05), but these were not significantly different among fish fed diets containing 1.45 and 1.70% valine (P>0.05). Broken-line model analyses on the basis of PAval in indicated that the dietary valine requirement of trout could be 1.50% (4.10) of diet (% dietary protein on a dry matter basis).
Figure 2. Broken line analysis of post-absorptive plasma valine concentrations (nmol mL−1) in rainbow trout fed graded levels of dietary valine. Values of the X-axis are the valine levels in the experimental diets. Values are means±SD of five replicates. Y=104.9x −80.274, Y′=237.6, R 2=0.8077.
3.2. Plasma ammonia concentrations
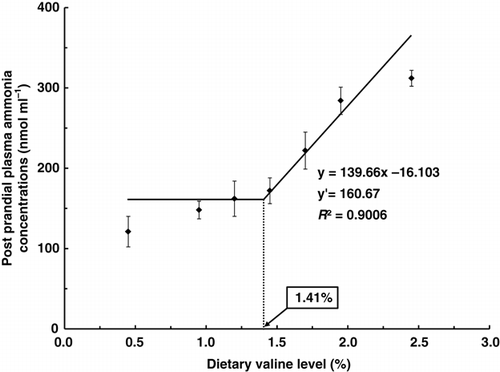
Post-prandial plasma ammonia concentrations (PPA, 5 h after intubation) and post-absorptive plasma ammonia concentrations (PAA, 24 h after intubation) in rainbow trout force-fed graded levels of dietary valine are shown in . Post-prandial plasma ammonia concentration of fish fed the diet containing 2.45% valine was significantly higher than those of fish fed diets containing 1.95% or less valine (P<0.05). Post-prandial plasma ammonia concentrations linearly increased with dietary valine levels from 1.45 to 2.45%. Post-absorptive plasma ammonia concentrations in the present study were not significantly different among the groups (P>0.05). Broken-line model analyses on the basis of PPA () indicated that the dietary valine requirement of rainbow trout could be 1.41% (3.85) of diet (% dietary protein on a dry matter basis).
Figure 3. Broken line analysis of post-prandial plasma ammonia concentrations (nmol mL−1) in rainbow trout fed graded levels of dietary valine. Values of the X-axis are the valine levels in the experimental diets. Values are means±SD of five replicates. Y=139.66x −16.103, Y′=160.67, R 2=0.9006.
4. Discussion
In the present study, the dietary valine requirement for the rainbow trout based on plasma free valine concentrations (PPval and PAval) was between 1.47 and 1.50% of the dry diet. This value corresponds to 4.02–4.10% of the dietary protein on a dry weight basis. In previous studies, the dietary valine requirements of rainbow trout have been reported to be 1.30% of diet by Ogino (Citation1980) and 1.55–1.58% of diet by Rodehutscord et al. (Citation1997) based on dose response or protein accretion. Results of the present study were similar to those reported for Rohu, L. rohita, 1.50% of diet (Murthy and Varghese Citation1997). Somewhat lower requirement values have been reported for other species including 0.71% of diet in channel catfish, Ictalurus punctatus (Wilson et al. Citation1980); 0.78% in lake trout, Salvelinus namaycush (Hughes et al. Citation1983); 0.78% in Nile tilapia, Oreochromis niloticus (Santiago and Lovell Citation1988); 1.20% in chum salmon, Oncorhynchus keta (National Research Council Citation1993); 1.30% in Chinook salmon, Oncorhynchus tshawytscha (Chance et al Citation1964); 1.42% in milkfish, C. chanos (Borlongan and Coloso Citation1993) and 1.42% in Catla, C. catla (Ravi and Devaraj Citation1991), while higher levels of valine in diet have been reported for mrigal, Cirrhinus mrigala, 1.52% (Ahmed and Khan Citation2006).
These wide variations observed in the valine requirements among fish species may be due to the differences in dietary protein sources, the reference protein for which amino acid pattern is being imitated, diet formulation, size and age of fish, genetic differences, feeding practices and rearing conditions (Rodehutscord et al. Citation1997). Zhou et al. (Citation2007) reported that the dietary lysine requirements of fish species that have high protein requirements, such as marine fish and freshwater carnivorous fish, are higher than those of the species that have low protein requirements including omnivores and herbivorous fish. However, the difference in the dietary valine requirement values listed above indicates that there is no apparent distinction in food habits or environmental needs which may help to give a clear classification of dietary valine requirements for various fish species.
The effects of dietary valine intake on PPval and PAval were dependent on the relative adequacy of the dietary valine supply, increasing up to the dietary valine requirement levels with no further increase above the requirement values. The pattern of post-prandial or absorptive plasma free amino acid concentrations of fish fed graded levels of the amino acid tested has been used in several studies in an attempt to confirm requirement values assessed by growth (Wilson et al. Citation1980; Thebault et al. Citation1985; Tibaldi and Tulli Citation1999). The plasma limiting amino acid concentration is expected to be low in fish given sub-requirement levels of the test amino acid, however when adequate levels are fed the plasma concentration tends to increase linearly as the dietary concentrations rise (Cowey Citation1995). As indicated above, a similar response was observed in the present study in that a breakpoint was observed in the plasma concentration value at 1.07% (PPval) or 0.97% (PAval) dietary valine.
In the present study, the dietary valine requirement for rainbow trout based on post-prandial plasma free ammonia concentration (PPA, 5 h after intubation) could be 1.41% of the dry diet. This value corresponds to 3.85% of the dietary protein on a dry weight basis. PPA was significantly affected by the dietary valine levels. However, PAA (24 h after intubation) was not affected by the dietary valine level. PPA remained low and virtually constant, but increased linearly when dietary valine level was higher than the requirement level. Similar observations were reported that PPA was affected in response to dietary arginine, lysine, methionine and threonine levels in rainbow trout (Ok Citation2002). This observation could be attributed to an increase in net protein synthesis with the increase in dietary amino acids. The ammonia excretion is related to the intake of protein (Koshio et al. 1993) and dietary amino acids balance (Kaushik et al. 1988). Considering protein and amino acid metabolism, results of the present study indicated that the dietary valine requirement in rainbow trout can be estimated based on PPA. Furthermore, the results showed that estimated valine requirement value based on PPA is lower than that based on PPval and PAval. This relationship between plasma free valine concentrations and plasma ammonia concentrations was not observed in this study. Future studies would look into the cause of the discrepancy in these values.
In conclusion, broken-line model analyses on the basis of PPval, PAval and PPA concentrations indicated that the dietary valine requirements of rainbow trout could be between 1.41% (3.85) and 1.50% (4.10) of diet (% of dietary protein basis).
Acknowledgements
This research was supported by the funds of the Feeds and Foods Nutrition Research Center, Korea Sea Grant Program and Brain Korea 21 Program at Pukyoung National University, Busan, Korea, Ewhajung trout farm in Sangju, Korea.
References
- Ahmed , I and Khan , MA . 2006 . Dietary branched-chain amino acid valine, isoleucine and leucine requirements of fingerling Indian major carp, Cirrhinus mrigala (Hamilton) . British Journal of Nutrition , 96 : 450 – 460 .
- Bai SC , Ok IH , Park GJ , Kim KW , Choi SM . 2003 . Development of modeling system for assessing essential amino acid requirements using surgically modified rainbow trout . Journal of Aquaculture 16 1 : 1 – 7 . in Korean, with English abstract .
- Borlongan , IG and Coloso , RM . 1993 . Requirements of juvenile milkfish, Chanos chanos Forsskal for essential amino acids . Journal of Nutrition , 123 : 125 – 132 .
- Chance , RE , Mertz , ET and Halver , JE . 1964 . Nutrition of salmonid fishes. XII. Isoleucine, leucine, valine and phenylalanine requirements of chinook salmon and interrelations between isoleucine and leucine for growth . Journal of Nutrition , 83 : 177 – 185 .
- Cowey , CB . 1995 . Protein and amino acid requirements: a critique of methods . Journal of Applied Ichthyology , 11 : 199 – 204 .
- Dabrowski , K . 1982 . Postprandial distribution of free amino acids between plasma and erythrocytes of common carp, Cyprinus carpio . Comparative Biochemistry and Physiology , 72A : 753 – 763 .
- Espe , M , Lied , E and Torrissen , KR . 1993 . Changes in plasma and muscle free amino acids in Atlantic salmon, Salmo salar during absorption of diets containing different amounts of hydrolysed cod muscle protein . Comparative Biochemistry and Physiology , 105A : 555 – 562 .
- Fernstrom , JD . 2005 . 4th amino acid assessment workshop: branched chain amino acids and brain function . Journal of Nutrition , 135 : 1539 – 1546 .
- Hughes , SG , Rumsey , GL and Nesheim , MC . 1983 . Dietary requirements for essential branched-chain amino acids by lake trout . Transactions of the American Fisheries Society , 112 : 812 – 817 .
- Jun-Young , B , Dae-Jung , K , Kwang-Yeol , Y , Sun-Gyu , K , Jeong-Yeol , L and Sungchul , CB . 2010 . Effects of Dietary Arachidonic Acid (20:4n-6) Levels on Growth Performance and Fatty Acid Composition of Juvenile Eel, Anguilla japonica . Asian-Aust. J. Anim. Sci. , 23 ( 4 ) : 508 – 514 .
- Kaushik , SJ , Fauconneau , B , Terrier , L and Gras , J . 1988 . Arginine requirement and status assessed by different biochemical indices in rainbow trout (Salmo gairdneri R.) . Aquaculture , 70 : 75 – 95 .
- Kim , KI . 1997 . Re-evaluation of protein and amino acid requirements of rainbow trout (Oncorhynchus mykiss) . Aquaculture , 151 : 3 – 7 .
- Koshio , S , Teshima , S , Kanazawa , A and Watase , T . 1993 . The effect of dietary protein content on growth, digestion efficiency and nitrogen excretion of juvenile kuruma prawns, Penaeus japonicus . Aquaculture , 113 : 101 – 114 .
- Murai , T , Ogata , H , Hirasawa , Y , Akiyama , T and Nose , T . 1987 . Portal absorption and hepatic uptake of amino acids in rainbow trout force fed complete diets containing casein or crystalline amino acids . Nippon Suisan Gakkaishi , 53 ( 10 ) : 1847 – 1859 .
- Murthy , HS and Varghese , TJ . 1997 . Quantitative dietary requirements of the Indian major carp, Labeo rohita for the essential amino acid valine . Indian Journal of Animal Sciences , 67 : 1028 – 1030 .
- National Research Council . 1993 . Nutrient requirement of fish . Washington , DC : National Academy Press .
- Nose , T . 1972 . Changes in patterns of free plasma amino acids in rainbow trout after feeding . Bulletin of Freshwater Fisheries Research Laboratory , 22 : 137 – 144 .
- Ogino , C . 1980 . Requirements of carp and rainbow trout for essential amino acids . Bulletin of the Japanese Society for the Science of Fish , 46 : 171 – 174 .
- Ok IH . 2002 . Determination of essential amino acid requirements by using plasma free amino acid concentrations in rainbow trout, Oncorhynchus mykiss [PhD dissertation] . Pukyong National University
- Ok , IH , Bai , SC , Park , GJ , Choi , SM and Kim , KW . 2001 . The patterns of plasma free amino acids after force-feeding in rainbow trout, Oncorhynchus mykiss with and without dorsal aorta cannulation . Aquaculture Research , 32 ( s1 ) : 70 – 75 .
- Park , GJ , Bai , SC , Ok , IH , Han , KM , Hung , SSO , Rogers , QR and Min , TS . 2005 . Post prandial plasma free arginine concentrations increase in rainbow trout fed arginine deficient diets . Asian-Australasian Journal of Animal Sciences , 18 : 396 – 402 .
- Plakas , SM , Katayama , T , Tanaka , Y and Deshimaru , O . 1980 . Changes in the levels of circulating plasma free amino acids of carp, Cyprinus carpio after feeding a protein and an amino acid diet of similar composition . Aquaculture , 21 : 307 – 322 .
- Ravi , J and Devaraj , KV . 1991 . Quantitative essential amino acid requirements for growth of catla, Catla catla (Hamilton) . Aquaculture , 96 : 281 – 291 .
- Robbins , KR , Norton , HW and Baker , DH . 1979 . Estimation of nutrient requirements from growth data . J. Nutr. , 109 : 1710 – 1714 .
- Robinson , EH , Wilson , RP and Poe , WE . 1981 . Arginine requirement and apparent absence of a lysine-arginine antagonist in fingerling channel catfish . J. Nutr. , 111 : 46 – 52 .
- Rodehutscord , M , Becker , A , Pack , M and Pfeffer , E . 1997 . Response of rainbow trout, Oncorhynchu mykiss to supplements of individual essential amino acids in a semi purified diet, including an estimate of the maintenance requirement of essential amino acids . Journal of Nutrition , 126 : 1166 – 1175 .
- Santiago , CB and Lovell , RT . 1988 . Amino acid requirements for growth of Nile tilapia . Journal of Nutrition , 118 : 1540 – 1546 .
- Schuhmacher , A , Wax , C and Gropp , JM . 1997 . Plasma amino acids in rainbow trout, Oncorhynchus mykiss fed intact protein or a crystalline amino acid . Aquaculture , 151 : 15 – 28 .
- Sunde , J , Kiessling , A , Higgs , D , Opstvedt , J , Venturini , G and Torrissen , KR . 2003 . Evaluation of feed protein quality by measuring plasma free amino acids in Atlantic salmon, Salmo salar after dorsal aorta cannulation . Aquaculture Nutrition , 9 : 351 – 360 .
- Thebault , H , Alliota , E and Pastoureauda , A . 1985 . Quantitative methionine requirement of juvenile seabass, Dicentrarchus labrax . Aquaculture , 50 : 75 – 87 .
- Tibaldi , E and Tulli , F . 1999 . Dietary threonine requirement of juvenile European seabass, Dicentrarchus labrax . Aquaculture , 175 : 155 – 166 .
- Vermeirssen , ELM , Scott , AP and Liley , NR . 1997 . Female rainbow trout urine contains a pheromone which causes a rapid rise in plasma 17α, 20β-dihydroxy-4-pregnen-3-one levels and milt amounts in males . Journal of Fish Biology , 50 : 107 – 119 .
- Walton , MJ and Wilson , R . 1986 . Postprandial changes in plasma and liver free amino acids of rainbow trout fed complete diets containing casein . Aquaculture , 51 : 105 – 115 .
- Wilson , RP . 2002 . “ Amino acids and proteins ” . In Fish nutrition , 3rd edn , Edited by: Halver , JE and Hardy , RW . 143 – 179 . San Diego , CA : Academic Press .
- Wilson , RP , Poe , WE and Robinson , EH . 1980 . Leucine, isoleucine, valine and histidine requirements of fingerling channel catfish . Journal of Nutrition , 110 : 627 – 633 .
- Zhou , QC , Wu , ZH , Chi , SY and Yang , QH . 2007 . Dietary lysine requirement of juvenile cobia, Rachycentron canadum . Aquaculture , 273 : 634 – 640 .