
Abstract
We investigated whether a fermented feed supplemented with carrot by-product is associated with body weight and meat quality in pigs, as well as whether it affects the expression of heat shock proteins (HSPs). The chemical composition of the fermented feed was 4% crude protein, 4% crude fat, 2% crude fibre, 0.2% phosphorus and 0.04% calcium. The daily weight gain and finished body weights of pigs fed the treated diet, which supplemented with the fermented diet including carrot by-product, were approximately 175 g and 5 kg greater, respectively, than those that received the basal diet. The transcriptional levels of HSP27 and HSP70 in the longissimus dorsi muscles of pigs fed the treated diet were lower than those that received the basal diets. The correlation coefficients (CORRs) between the expressions of HSP and meat traits including carcass weight, quality grade, crude fat, water holding capacity and drip loss showed strong negative effects. The treated diet reduces the expression levels of HSP27 and HSP70 in pigs, and this is associated with increased daily weight gain, final body weight and higher meat quality as measured by carcass weight, quality grade and water-holding capacity.
Introduction
Carrot is an important root vegetable for many animals including humans. The consumption of carrot-based beverages by humans has been consistently increasing in many countries (Scholten et al. Citation1999; Canibe & Jensen Citation2003). Carrot by-products include valueless carrots sorted during cultivation and remnants from the carrot beverage industry. Carrots contain a variety of micronutrients, such as beta-carotene (vitamin A), vitamin C, vitamin E, chlorophyll, magnesium, iodine and potassium, which play critical roles in overall health and also show antitumor activity (Krinsky Citation1989; Olson Citation1992). Beta-carotene from carrots, which is converted into vitamin A in the body, functions in the protection and improvement of sight, supports fatigue recovery and reduces blood pressure by decreasing cholesterol (Bendich Citation1993). This in turn can help prevent adult diseases such as hypertension and diabetes (Brooks et al. Citation2001).
Fermented feeds for pigs have been gradually changing as a result of advanced fermentation techniques (Scholten & Verdoes Citation1997; Chen & Tang Citation1998). Fermented feeds are prepared using basal medium components supplemented with agro by-products (e.g., carrot) and probiotics including lactic acid bacteria. Applying fermented feed made from food by-products decreases disposal waste, feed-processing costs and the environmental burden of food production (Omenn et al. Citation1996; Schieber et al. Citation2001). Thus, fermented feed processing should be optimised to reduce the amount of waste globally. Functional feed prepared from fermented feed using by-products is an important and rapidly growing component of pig breeding (Chu et al. Citation2011 and Citation2012).
Heat shock proteins (HSPs) are highly conserved proteins that are expressed in response to thermal stress (Ellis & van der Vies Citation1991; Mathew & Morimoto Citation1998). Mammalian HSPs are classified as high- (HSP70 and HSP90) and low-molecular-weight (HSP27) HSPs. High-molecular-weight HSPs are ATP-dependent chaperones and require co-chaperones to modulate their conformation and ATP binding. Conversely, low-molecular-weight HSPs are ATP-independent chaperones (Garrido et al. Citation2001). The overexpression of both HSP27 and HSP70 is associated with the tumorigenic potential of rodent cells (Jaattela Citation1995; Garrido et al. Citation1998). HSPs have a cytoprotective function and improve survival under lethal conditions (e.g., obesity) (Arrigo Citation1998; Benjamin & McMillan Citation1998). Stress can trigger defence mechanisms that are not active under normal conditions to maintain the internal environment (Morimoto Citation1993). HSP responses to stresses have been observed in all living organisms. Sudden changes in gene expression enhance the synthesis of stress proteins such as HSPs (Gupta et al. Citation2010). Almost all forms of stress induce the syntheses of HSPs in cells that are generally adapted to stress (De Maio Citation1999).
In the present study, we examined the impact of a fermented diet supplemented with carrot by-product on final body weight and daily gains in the Berkshire pig breed. We also investigated whether the expression of HSPs was correlated with weight gain and meat quality using RT-PCR.
Materials and methods
Fermentation conditions
Fermented carrot by-product diets mainly contained 67.1% carrot by-products, 32.4% corn grain and 0.3% rice bran, respectively. The moisture of the diets was regulated to approximately 60% by water and each component was simultaneously well mixed and then fermented for 24 h at 37°C after supplementation with 0.1% probiotics. The probiotics per gram contained approximately 4.6 × 107 colony-forming units (cfu) lactic acid bacteria (Lactobacillus plantarium) and 4.4 × 107 thallus-forming units (tfu) yeast (Saccharomyces cerevisae). After the mixture was transferred into anaerobic plastic containers, it was fermented at room temperature (RT) for 14 days. Each composition was assayed according to the methods of previous study (Chu et al. Citation2012).
Animals, rearing and tissue collection
Experimental LYD (Landrace–Yorkshire–Duroc composite populations) pigs (n = 8) were reared at the han-il farm in South Korea. Pigs were fed for seven months in accordance with the Guide for the Care and Use of Laboratory Animals (Gyeongnam National University of Science and Technology Animal Care Committee). The fermented diet including carrot by-product was dried and then converted to powder. The fermented powder was added for pig rearing by ratio of 9% fermented powder and 91% basal diet, named as treated diet. The pigs were fed the treated diet for 45 days from 165 to 210 days post-birth. The animals were allowed free access to unlimited fresh water during the experimental periods. The experimental pigs serviced unlimited feed everyday and the next day residual feed was analysed for feeding efficiency. Initial and finished body weights (IBW and FBW) were measured for average daily gain (ADG). Experimental pigs were slaughtered during the growth stage at approximately 110–120 kg body weight. During transport for slaughter, animals were kept off feed, held for ∼1 h, humanely slaughtered and then the longissimus dorsi muscles were harvested.
RNA extraction
Total RNA was isolated from collected longissimus dorsi using Trizol reagent according to the manufacturer's instruction (Life Technologies, Invitrogen). Briefly, Trizol reagent was added to ground tissue. The mixture was well-suspended with homogeniser, and a aliquot of the mixture was transferred to a 1.5 mL E-tube. The mixture was left alone at RT for 10 min and centrifuged at 12,000 rpm for 10 min, and the solution excluding the cell debris was saved into a new tube. The harvested solution was treated with chloroform, and isopropanol was added to precipitate RNA. The RNA was washed with 70% ethanol and dried at RT. The extracted RNA was dissolved in RNase free water and quantified spectrophotometrically at 260 nm. For each sample, an RNA aliquot was subjected to electrophoresis on a 1.5% agarose gel to verify the extracted RNA.
Semi-quantitative reverse transcription polymerase chain reaction (RT-PCR)
For RT-PCR, first-strand cDNAs of HSP27, HSP70, HSP90 and glyceraldehyde-3-phosphate dehydrogenase (GAPDH) were synthesised using Superscript II Reverse Transcriptase according to the manufacture's protocol (Invitrogen, USA). Briefly, the extracted RNA was added to a reaction mixture consisting of first-strand buffer (Invitrogen, Carlsbad, CA), deoxynucleotide triphosphate (dNTP) (Promega, USA) dissolved in RNase-free water, dithiothreitol (DTT) (Invitrogen, USA), SuperScript Reverse Transcriptase II (Invitrogen, USA), oligo-d(T) 12–18 primer (Invitrogen, USA), RNase Inhibitor (Invitrogen, USA) and RNase free water. The RT step was carried out at 42°C for 1 h, followed by heating at 70°C for 15 min and the addition of RNase H at 37°C for 20 min before storage at 4°C. A set of negative controls was also included, which lacked the reverse transcription reaction. The GAPDH gene was used as an internal control to test the relative amount of target gene from different tissue samples (). The RT-PCR products of the two genes were separated on a 2% tris-acetate-ethylene (TAE) diaminetetraacetic acid agarose gel and visualised by ultraviolet radiation after ethidium bromide staining.
Table 1. Oligonucleotides used in RT-PCR.
Carcass traits, carcass grades and analyses
The Berkshire pigs weighing approximately 110–120 kg were transported to an abattoir near the experimental station. They were slaughtered by stunning with electrical tongs (300 volts for 3 s) after 12 h of feed restriction. The shocked pigs were exsanguinated while being hanged. Carcasses were then placed in a dehairer at 62°C for 5 min and remaining hair was removed by using a knife and flame. Carcasses were eviscerated and split before being placed in a chiller set at 5°C for 12 h.
South Korean carcass grade is explained at the website of Korea Institute for Animal Products Quality Evaluation (KAPE Citation2011). All the pork carcasses in South Korea are graded both in quality and conformation terms. The quality of pork carcasses is graded 1+, 1, 2 and 3 based on the marbling, lean colour and conditions of belly streaks. The conformation terms of pork carcasses is graded A, B, C and D by assessing carcass weight (CW), backfat thickness, balance, muscle, fat condition and so on.
Percentage of dressing was calculated by the ratio of cold CW against live weight. Backfat thickness was measured by using the 10th rib as three-quarters distance along the longissimus dorsi towards the belly. The longissimus dorsi (6th–13th rib) was cut off and kept at 5°C before it was transported to the laboratory for the determination of chemical composition. The concentrations of moisture, crude protein (CP), crude fat (CF) and ash of the longissimus dorsi were determined according to the methods of AOAC (2000) about 24 h after slaughter. The water holding capacity (WHC) was determined as described previously (Kristensen & Purslow Citation2001). The meat lightness (ML) and fat lightness (FL) were recorded after a 30-min blooming at 1°C using a Minolta Chromameter (CR400; Minolta, Japan).
Statistical analyses
Data are presented as means ± SE. Data were analysed using the generalized linear model (GLM) procedure in SAS (SAS Inst. Inc., Cary, NC). Correlations between meat traits and gene expression levels were calculated using the correlation coefficient (CORR) procedure. Results were considered significant when the P value was less than 0.05.
Results and discussion
The treated diet increases body weight and daily gains in fattening pigs
The normal-finishing body weights of both barrow and gilt pigs were used to study the effects of the treated diet. When pigs are fed by the fermented diets including Lactic acid bacteria, short-chain organic acids, including acetic acid, propionic acid and lactic acid, and various minerals, gastrointestinal health in pigs trigger important improvement via decreases of pathogenic bacteria (Boesen et al. Citation2004). describes the ingredients of the basal diet and the treated diet used in the present study. Finished body weight and daily gain were compared between groups fed the different diets for 45 days (). ADG of the treated diet group showed an increased value of 175 g (P < 0.05) when compared with normal diet group. As a result, FBW of the group fed with the treated diet exhibited 5 kg (P < 0.05) greater than that of the normal diet. Anaerobic fermentation enhances the storability, palatability and nutrient value of feedstuffs (Gao et al. Citation2008). Fermented feed can be added to piglet feed due to its high CP and low moisture concentrations (Kim et al. Citation2010). The development of feed ingredients using agriculture by-products may decrease breeding costs in the pig industry. Taken together, our results demonstrate that the treated diet may increase finished body weight by elevating daily weight gain.
Table 2. Ingredients of the basal diet and the fermented diet supplemented with carrot by-product.
Table 3. Body weight (kg) and daily gain (g) of fattening pigs fed the basal and the treated diets.
The treated diet reduces the expression of the HSP27 and HSP70 genes in fattening pigs
To understand the mechanism behind the increased finished body weight caused by the treated diet, genes that play roles in fatigue, blood presssure and stress were considered for analyses. Among them, since stresses expect to play critical roles to pig's physiology such as body weight gain, average daily gain and feeding efficency, first, we analysed the expression of HSP genes. The increased expression of stress-related genes such as HSPs is a ubiquitous response to stress (Santoro Citation2000; Walsh et al. Citation2003). Exposure to stress-causing agents causes an immediate block of every important metabolic process, including DNA replication, transcription and translation, until the cells recover (Biamonti & Caceres Citation2009). HSPs protect cells from injury while promoting recovery to normal growth conditions and ultimately survival (Morimoto & Santoro Citation1998).
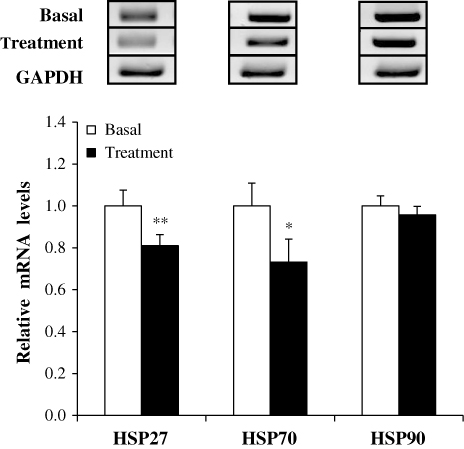
The consumption of the treated diet led to the reduced transcriptional expression of the stress proteins HSP27 (P < 0.01) and HSP70 (P < 0.05) in fattening pigs (). Although HSP90 was also analysed by RT-PCR, the gene did not show any differential expression between basal and the treated groups (). The fermented feed improves intestinal health via removal of pathogenic bacteria owing to lactic acid bacteria, short chain fatty acids, and various mineral components (Niba et al. Citation2009). The improved intestinal health might be trigged to reduced energy losses through efficient maintainence of intracellular environment due to repressed expression amounts of genes related with stresses. Therefore, the results of this study suggest that the fermented feed reduces the expression levels of specific stress-related genes, such as HSP27 and HSP70, resulting in higher body weight via increased daily weight gain.
Expression levels of HSP genes are negatively correlated with meat traits in the longissimus dorsi
To analyse the correlation between economic traits and HSP, the longissimus dorsi muscles from LYD pigs (n = 8) were collected after slaughter and analysed for meat traits. The correlation between meat characteristics and gene expression levels in the longissimus dorsi were measured (). CF, CW, yield grade (YG), quality grade (QG), WHC, and drip loss (DL) showed negative correlations with the expression of the HSP27 and HSP70 genes. ML and FL exhibited positive correlations with the expression HSP27 and HSP70 genes. HSP70 single strand polymorphism (SNP) is differently associated with meat pH, backfat thickness and CW (Jin et al. Citation2006). It is assumed that expression amounts of HSPs directly relate with meat quality traits, because HSP70 SNP carrying different genotypes associates with several meat quality traits via physiologically changed role(s) (Jin et al. Citation2006). Therefore, the results of this study suggest that the downregulations of HSPs associate to increase CW, YG, QG, CF, WHC and DL, but decrease ML and FL in fattening pigs fed the fermented feed with carrot by-product.
Table 4. CORRs between expression levels of HSPs and meat traits in the longissimus dorsi of fattening pigs fed the treated diet.
Conclusion
We predicted that the treated diet supplemented with carrot by-product would increase final body weight by increasing daily weight gain owing to decreased expression of the stress protein HSP27 and HSP70 in fattening pigs. Indeed, the expression levels of HSP27 and HSP70 were negatively correlated with meat traits including CW, QG, WHC and DL. Future studies should focus on identifying micronutrients and dietary factors that regulate the expression of HSPs, which can be incorporated into an improved formula diet.
Funding
This work was supported by grants from the Priority Research Centers Program [2011-0022965] of the Ministry of Education, Science and Technology, Republic of Korea, the support of the Cooperative Research Program for Agriculture Science and Technology Development [Project No. PJ0077672011] of the Rural Development Administration, Republic of Korea, and Gyeongnam National University of Science and Technology Grant.
Additional information
Funding
References
- Arrigo AP. 1998. Small stress proteins: chaperones that act as regulators of intracellular redox state and programmed cell death. Biol Chem. 379:19–26.
- Bendich A. 1993. Biological functions of dietary carotenoids. Ann N Y Acad Sci. 691:61–67. doi:10.1111/j.1749-6632.1993.tb26157.x
- Benjamin IJ, McMillan DR. 1998. Stress (heat shock) proteins: molecular chaperones in cardiovascular biology and disease. Circ Res. 83:117–132. doi:10.1161/01.RES.83.2.117
- Biamonti G, Caceres JF. 2009. Cellular stress and RNA splicing. Trends Biochem Sci. 34:146–153. doi:10.1016/j.tibs.2008.11.004
- Boesen HT, Jensen TK, Schmidt AS, Jensen BB, Jensen SM, Moller K. 2004. The influence of diet on Lawsonia intracellularis colonization in pigs upon experimental challenge. Vet Microbiol. 103:35–45. doi:10.1016/j.vetmic.2004.06.008
- Brooks PH, Beal JD, Niven SJ. 2001. Liquid feeding of pigs: potential for reducing environmental impact and for improving productivity and food safety. Recent Adv Anim Nutr Aust. 13:49–63.
- Canibe N, Jensen BB. 2003. Fermented and nonfermented liquid feed to growing pigs: effect on aspects of gastrointestinal ecology and growth performance. J Anim Sci. 81:2019–2031.
- Chen BH, Tang YC. 1998. Processing and stability of carotenoid powder from carrot pulp waste. J Agric Food Chem. 46:2312–2318. doi:10.1021/jf9800817
- Chu GM, Yang BS, Kim HY, Kim JH, Ha JH, Kim CH, Lee SD, Song YM. 2011. Effects of supplemental fermented agro by-products diet on the growth performances, blood characteristics and carcass traits in fatterning pigs. Asian Australas J Anim Sci. 24:1464–1472. doi:10.5713/ajas.2011.10459
- Chu GM, Yang JM, Kim HY, Kim CH, Song YM. 2012. Effects of fermented mushroom (Flammulina velutipes) by-product diets on growth performance and carcass traits in growing-fattening Berkshire pigs. J Anim Sci. 83:55–62. doi:10.1111/j.1740-0929.2011.00924.x
- De Maio A. 1999. Heat shock proteins: facts, thoughts, and dreams. Shock. 11:1–12. doi:10.1097/00024382-199901000-00001
- Ellis RJ, van der Vies SM. 1991. Molecular chaperones. Annu Rev Biochem. 60:321–347. doi:10.1146/annurev.bi.60.070191.001541
- Gao L, Yang H, Wang X, Huang Z, Ishii M, Igarashi Y, Cui Z. 2008. Rice straw fermentation using lactic acid bacteria. Bioresour Technol. 99:2742–2748. doi:10.1016/j.biortech.2007.07.001
- Garrido C, Fromentin A, Bonnotte B, Favre N, Moutet M, Arrigo AP, Mehlen P, Solary E. 1998. Heat shock protein 27 enhances the tumorigenicity of immunogenic rat colon carcinoma cell clones. Cancer Res. 58:5495–5499.
- Garrido C, Gurbuxani S, Ravagnan L, Kroemer G. 2001. Heat shock proteins: endogenous modulators of apoptotic cell death. Biochem Biophys Res Commun. 286:433–442. doi:10.1006/bbrc.2001.5427
- Gupta SC, Sharma A, Mishra M, Mishra RK, Chowdhuri DK. 2010. Heat shock proteins in toxicology: how close and how far? Life Sci. 86:377–384. doi:10.1016/j.lfs.2009.12.015
- Jaattela M. 1995. Over-expression of hsp70 confers tumorigenicity to mouse fibrosarcoma cells. Int J Cancer. 60:689–693. doi:10.1002/ijc.2910600520
- Jin HJ, Park BY, Park JC, Hwang IH, Lee SS, Yeon SH, Kim CD, Cho CY, Kim YK, Min KS, et al. 2006. The effects of stress related genes on carcass traits and meat quality in pigs. Asian Australas J Anim Sci. 19:280–285.
- KAPE (Korea Institute for Animal Products Quality Evaluation). 2011. Animal products grading service yearbook. 4th ed.
- Kim YI, Seok JS, Kwak WS. 2010. Evaluation of microbially ensiled spent mushroom (Pleurotus osteratus) substrates (bed-type cultivation) as a rough age for ruminants. Korean J Anim Sci Technol. 52:117–124. doi:10.5187/JAST.2010.52.2.117
- Krinsky NI. 1989. Carotenoids and cancer in animal models. J Nutr. 119:123–126.
- Kristensen L, Purslow P. 2001. The effect of ageing on the water-holding capacity of pork: role of cytoskeletal proteins. Meat Sci. 58:17–23. doi:10.1016/S0309-1740(00)00125-X
- Mathew A, Morimoto RI. 1998. Role of the heat-shock response in the life and death of proteins. Ann N Y Acad Sci. 851:99–111. doi:10.1111/j.1749-6632.1998.tb08982.x
- Morimoto RI. 1993. Cells in stress: transcriptional activation of heat shock genes. Science. 259:1409–1410. doi:10.1126/science.8451637
- Morimoto RI, Santoro MG. 1998. Stress-inducible responses and heat shock proteins: new pharmacologic targets for cytoprotection. Nat Biotechnol. 16:833–838. doi:10.1038/nbt0998-833
- Niba AT, Beal JD, Kudi AC, Brooks PH. 2009. Bacterial fermentation in the gastrointestinal tract of non-ruminants: influence of fermented feeds and fermentable carbohydrates. Trop Anim Health Prod. 41:1393–1407. doi:10.1007/s11250-009-9327-6
- Olson RE. 1992. Vitamins and carcinogenesis: an overview. Proceedings of first international congress on vitamins and biofactors in life science. J Nutr Sci Vitaminol Spec. 38:313–316. doi:10.3177/jnsv.38.Special_313
- Omenn GS, Goodman GE, Thornquist MD, Balmes J, Cullen MR, Glass A, Keogh JP, Meyskens FL, Valanis B, Williams JH, et al. 1996. Effects of a combination of b-carotene and vitamin A on lung cancer and cardiovascular disease. N Engl J Med. 334:1150–1155. doi:10.1056/NEJM199605023341802
- Santoro MG. 2000. Heat shock factors and the control of the stress response. Biochem Pharmacol. 59:55–63. doi:10.1016/S0006-2952(99)00299-3
- Scholten RHJ, van der Peet-Schweringa CMC, Verstegen MWA, den Hartoga LA, Schramab JW, Vesseura PC. 1999. Fermented co-products and fermented compound diets for pigs: a review. Anim Feed Sci Technol. 82:1–19. doi:10.1016/S0377-8401(99)00096-6
- Schieber A, Stintzing FC, Carle R. 2001. By-products of plant food processing as a source of functional compounds – recent developments. Trends Food Sci Technol. 12:401–413. doi:10.1016/S0924-2244(02)00012-2
- Scholten RHJ, Verdoes N. 1997. The Dutch benefit of a recycling rule. Pigs. 13:16–17.
- Walsh NP, Alba BM, Bose B, Gross CA, Sauer RT. 2003. OMP peptide signals initiate the envelope-stress response by activating DegS protease via relief of inhibition mediated by its PDZ domain. Cell. 113:61–71. doi:10.1016/S0092-8674(03)00203-4