
ABSTRACT
Data from eight experiments were jointly analysed to characterize the ingestive behavior, displacement patterns, use of feeding stations and the dynamics of meals of beef heifers in different phenological stages of winter forages. The experiments included 360 heifers, with initial age of eight months and average body weight of 145.0 ± 17 kg. The grazing method was put-and-take stocking with variable number of animals. Grazing time, duration of meal and interval between meals were similar in the morning shift, regardless of the phenological stage. At the vegetative and pre-reproductive stages, in the afternoon, meals were longer. The average daily gain was greater at pre-reproductive being associated with the duration of the meal in the afternoon shift. Heifers change their ingestive behavior, bite rate, duration of meals, interval between meals, patterns of forage selection and number of steps per minute according to phenological stages and evaluation shifts in order to maintain a similar bite mass.
Introduction
Canopy structure is a central characteristic that determines both: the forage growth dynamics and the ingestive behavior of grazing animals. This structure is influenced by the management, plant phenological stage and the interaction between them. The temporal variation in the quality and quantity of forage available is physiological and herbivorous animals develop grazing strategies along the grazing cycle, as a tool for compensation of these changes. Thus, it is expected that the ingestive behavior reflects the variations occurred in the amount and quality of ingested forage (Stobbs Citation1975).
Studies on annual winter grasses showed the influence of the plant growth stage on forage intake rate (Pedroso et al. Citation2004), animal performance (Pilau et al. Citation2005), ingestive behavior and displacement patterns of beef heifers (Glienke et al. Citation2010). In general, the nutritional value of forage plants decreases along its phenological stages through reduced leaf: stem ratio and increased cell wall lignification (Van Soest Citation1994).
The knowledge of ingestive behavior strategies of herbivorous animals can help to establish a pasture management that allows optimal use of forage and provide efficiency to the system. Such strategies determine the nutrient intake of these animals, as well as the location and level of the impact caused by such animals on the vegetation (Prache and Peyraud Citation2001). This intake depends on factors that determine the beginning and interruption of successive meals as well as their temporal arrangements (Gregorini et al. Citation2008). There is little progress in the understanding of the factors mediating these relationships and more information is required in order to understand them (Chilibroste et al. Citation2015).
In the scientific context, a single experimental work may not confirm and generalize its results and conclusions. Thus, there is a repetition of the same experiment at several different locations with the goal of checking the repeatability of results. In this way, the grouping of data for the same set of information allows broader conclusions at a lower cost (Lovatto et al. Citation2007).
Assuming that changes in the pasture growth stages modify the canopy structure and, consequently, herbivorous animals change their ingestive behavior, this study analysed and characterized the ingestive behavior, displacement patterns, use of feeding stations and the dynamics of meals of beef heifers grazing on winter forage species at three phenological stages. Also, the sward structure, chemical composition of the forage from the grazing simulation and the average daily gain of heifers were related with these variables.
Material and methods
Data analysed came from eight experiments conducted between 2003 and 2013 at the Federal University of Santa Maria (UFSM). The ingestive behavior of beef heifers, from eight to twelve months of age, on cool season pasture, at three phenological stages of the grass (vegetative, pre-reproductive and reproductive) was evaluated and comprised the database analysed.
The experiments were conducted in the Central Depression of Rio Grande do Sul, Brazil. The climate in the region is humid subtropical, according to the Köppen classification. The soil is classified as Paleudalf (EMBRAPA Citation2006). Mean values for the chemical characteristics of the soil were: pH-H2O: 5.0; pH SMP: 5.8; clay: 19.2 m V−1; P: 13.4 mg L−1; K: 92 mg L−1; OM: 2.7 m V−1; Al3+: 0.2 cmolc L−1; Ca2+: 4.6 cmolc L−1; Mg2+: 2.2 cmolc L−1; base saturation: 56.6%; Al saturation: 3%.
The experiments included 360 beef heifers with initial age of eight months and average body weight (BW) of 145.0 ± 17 kg. The pastures consisted of black oat (Avena strigosa Schreb.), ryegrass (Lolium multiflorum Lam.) and red clover (Trifolium pratense). Fertilization in the experimental areas followed the recommendations of the Chemicals and Soil Fertility Committee RS/SC (2016). In coverage, 77 kg of nitrogen ha−1 was applied as urea.
The grazing method was put-and-take stocking to maintain 1500–2000 kg dry matter (DM) ha−1 of forage mass (FM). In all experiments, two or three repetitions of the area were used, with three test animals in each repetition and a variable number of animals to maintain the forage mass. The phenological stages of grass were divided into vegetative (FM with 0% inflorescence, 45.4% leaf blade and 23.4% stem); pre-reproductive (FM composed of 4.1% inflorescence, 32.9% leaf blade and 30.7% stem) and reproductive (FM composed of 14.1% inflorescence 17.0% leaf blade and 31.8% stem).
Forage and pasture variables comprising the database were: forage mass (kg DM ha−1), canopy height (cm), leaf blade mass (kg DM ha−1), stem mass (kg DM ha−1), dead material mass (kg DM ha−1), leaf: stem ratio and leaf: dead material ratio. Variables related to forage as grazed were: crude protein (Euclides et al. Citation1992), in situ dry matter digestibility was determined by incubation for 48 h in the rumen of a fistulated bovine (Demarquilly et al. Citation1969) and neutral detergent fiber (Senger et al. Citation2008), all in percentage of DM. The variable related to the performance of heifers was the average daily gain (kg).
Evaluations related to ingestive behavior, displacement patterns and use of feeding stations were: time of grazing, rumination and other activities, in minutes and/or hours; bite rate (bites minutes−1), number of feeding stations per minute, time spent at each feeding station (second), the number of steps taken per minute, the number of steps taken between each feeding station and bites per feeding station. A feeding station was defined as the half-cylinder-shaped area directly in front of and on each side of the animal when its front feet are stationary (Laca et al. Citation1992), and a footstep was defined as each movement of the forelegs. Ingestive behavior was measured through visual observation in continuous periods of 18 h. The activities of grazing, rumination and other activities were recorded every 10 min (Jamieson and Hodgson Citation1979). From these data, the duration and number of meals and the time interval between meals were calculated. A meal was considered a long sequence of grazing with, at least, two successive 10 min in the grazing activity. The minimum interval between meals was considered to be 20 min. The variables of ingestive behavior, displacement patterns, use of feeding stations and dynamics of meals were calculated for each grazing shift, with the morning shift being the interval from 7:00 to 12:59h; the afternoon shift being the interval from 13:00 to 18:59h; the night shift was considered as the interval from 19:00 to 00:59h.
The variables measured in the experiments were stratified according to the phenological stages of grass (vegetative, pre-reproductive and reproductive) and evaluation shifts (morning, afternoon, night). A graphical analysis of residuals tested linearity deviation. Data were analysed using the software SAS 8.2, subjected to analysis of variance and F-test at 5% probability using the Mixed procedure and, when detected differences, the means were compared using the lsmeans test. We used a mixed model with fixed effects (stage of grass, evaluation shifts and their interactions) and random effects (year of study, the residual and the nested paddocks for each phenological stages of grass). We performed a structure selection test using the Bayesian information criterion (BIC) to determine the model that best represented the data. The variables were subjected to Pearson correlation test. We used a stepwise method for multiple regression analysis to identify the most influential independent variables. From the equations obtained, we selected the one with the lowest p value, residual variance and number of independent variables, and the highest coefficient of determination.
Results
At the vegetative stage, forage mass (FM) was made up of 45.4% leaves, 23.4% stems and 17.9% dead material (). Forage mass was 668.6 kg of dry matter (DM) ha−1 (43.9%) lower when compared to the reproductive stage. At the vegetative, we observed 317.5 kg DM ha−1 (85.1%) more leaf blades in the FM, 1.9 times less mass of stems and mass of dead material was 2.9 times lower. The leaf: stem ratio and leaf: dead material ratio were 6 and 5.2 times higher, respectively, when compared with the reproductive stage. The leaf blades bulk density was 2.2 times higher and the dead material bulk density was 2.0 times lower at this stage than at the reproductive stage. At the pre-reproductive stage, these variables presented intermediate values to other growth stages.
Table 1. Structural and qualitative parameters of cold season forage species under beef heifers grazing in different phenological stages.
Total bulk density (157.27 kg cm−1) and stem bulk density (43.61 kg cm−1), at the vegetative stage, were 1.3 and 1.5 times lower, respectively, compared to the pre-reproductive and reproductive stages, which were not significantly different from each other. At this stage, forage as grazed exhibited crude protein content 9.6% higher and neutral detergent fiber (NDF) 14.0% lower when compared to the reproductive stage (). At the pre-reproductive stage, the values of crude protein and NDF were intermediate to the other phenological stages.
Canopy height (11.7 cm), stocking rate (1019.9 kg BW ha−1) and in situ digestibility (82.0% DM) were similar between vegetative and pre-reproductive stages. In these stages, forage digestibility was 9.5% higher while canopy height and the stocking rate were 3.6 cm (30.5%) and 558.7 kg BW ha−1 (54.8%), respectively, lower than in the reproductive stage. Daily weight gain of heifers was similar (0.939 kg) between vegetative and reproductive stages. At the pre-reproductive stage, this gain was 0.187 kg day−1 (19.9%) higher than that registered in other growth stages.
Crude protein content of forage as grazed, at the growth stages evaluated, was explained by leaf:stem ratio (LSR), according to the equation: Ŷ = 11.93 + 8.62LSR; P < 0.0001; r 2 = 0.65. The neutral detergent fiber content of forage as grazed was correlated with the amount of dead material (r = 0.56; P < 0.0001), which was lower at the vegetative stage (). The DM in situ digestibility was correlated with the mass of leaves (r = 0.47; P < 0.0001), which was lower in at the reproductive stage of the pasture ().
At the pre-reproductive stage, FM consisted of 32.9% leaves, 30.7% of stems and 26.7% dead material. At the reproductive stage, FM consisted of 17.0% leaves, 31.8% stems and 36.7% dead material ().
Bite rate was similar between vegetative and pre-reproductive stages (51.1 bites min−1). This variable was 4.8 bites min−1 (10.3%) higher in these stages compared to the reproductive stage. The bite rate was negatively associated with forage mass (r = −0.64; P < 0.0001).
The changes in canopy structure () in the phenological stages of the grass caused no significant change in daily grazing time (348.6 min; P = 0.1025) and the bite mass (0.37 g; P = 0.5429). At the vegetative stage, rumination time was 40.19 min (21.4%) shorter and the time spent in other activities was 38.4 min (11.4%) longer compared to the pre-reproductive and reproductive stages. At the pre-reproductive and reproductive stages, the daily time spent in rumination (228.4 min) and in other activities (346.6 min; ) was similar.
Table 2. Variables of intake behaviour of heifers in cool season pasture grazing.
There was interaction between phenological stages × evaluation shifts for the variables time spent in grazing (P < 0.0001), ruminating (P = 0.0038) and in other activities (P < 0.0001), duration of meal (P = 0.0004) and the interval between meals (P < 0.0001; ). In the morning shift, grazing time (142.3 min), duration of meal (56.6 min) and the interval between meals (104.3 min) were similar, regardless of the phenological stage. The duration of meal in the morning shift was positively correlated with rumination time in the night shift (r = 0.27; P = 0.0197). The rumination time was 25.4 min (38.5%) shorter in the vegetative stage compared to the reproductive stage. At the pre-reproductive stage, the rumination time was intermediate to other stages. The rumination time in the morning shift, at the vegetative stage, was negatively associated with DM in situ digestibility (DMD; r = −0.50; P < 0.0001). The time spent in other activities was 24.9 min (18.9%) longer in the vegetative stage compared to other stages. The time in other activities (131.9 min) was similar between the pre-reproductive and reproductive stages.
Figure 1. Variables of intake behaviour and meals dynamics of heifers in cool season pasture grazing during the phenological stages (vegetative, pre-reproductive and reproductive) and evaluation shifts (morning, afternoon and night).
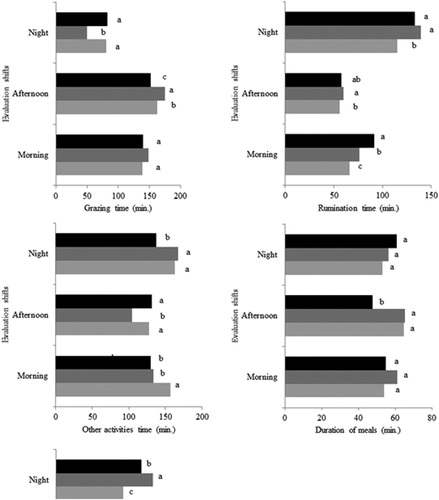
In the afternoon, grazing time was intermediate at the vegetative stage and this variable was 23.2 min (15.3%) longer at pre-reproductive than at reproductive. At the pre-reproductive stage, grazing time was explained by the dead material mass (DMM), according to the equation: Ŷ = 135.08 + 0.007DMM; P = 0.0014; r 2 = 0.32. The increase of 1 kg DM ha−1 dead material is expected to cause an increase of 0.007 min in the grazing time of heifers. The rumination time was 4.2 min (7.6%) shorter at the vegetative stage than at pre-reproductive. The rumination time at the reproductive stage was similar to other evaluated stages. The time spent in other activities, in this shift, was similar (129.2 min) at the vegetative and reproductive stages. This variable was 24.6 min (23.5%) longer in these stages when compared to pre-reproductive. The duration of meal was similar (64.9 min) between vegetative and pre-reproductive stages, being 17.2 min (36.0%) longer in those stages compared to the reproductive stage. The duration of meal, in this shift, at the reproductive stage was associated with the mass of leaf blades (r = 0.57; P = 0016) and with DM in situ digestibility (r = 0.48; P < 0.0001). The interval between meals, regardless of the phenological stage, was similar in this shift (88.4 min).
In the night shift, grazing time was similar between vegetative and reproductive stages (81.6 min). The grazing time was 32.1 min (67.0%) longer in these stages compared to the pre-reproductive stage. The rumination time was 21.4 min (18.6%) shorter at the vegetative stage when compared to the other stages. The rumination time was similar (136.5 min) between the pre-reproductive and reproductive stages. The time spent in other activities was similar (165.3 min) between the vegetative and pre-reproductive stages, and this variable was 27.5 min (19.9%) longer in these stages compared to the reproductive stage. The duration of meal was similar (56.8 min), independent from the phenological stage. The interval between meals was 76.0 min (43.7%) shorter at the vegetative stage when compared to pre-reproductive. At reproductive, the interval between meals was intermediate to other stages. Regardless of the phenological stage, the interval between meals was used in other activities in the morning (r = 0.38; P < 0.0001) and afternoon (r = 0.25; P = 0.0021). The longest interval between meals in the night shift was related to the long rumination time (r = 0.20; P = 0.0170) and time spent in other activities (r = 0.24; P = 0.0044) in this shift, at the pre-reproductive stage.
There was no interaction between phenological stages × evaluation shifts for the number of meals (P = 0.3031). The number of meals was similar (2.3), regardless of the phenological stage. In the morning shift, the number of meals was intermediate as compared to the other shifts. In the afternoon, the number of meals was 1.1 (72.6%) higher than in the night shift.
There was interaction between growth stages × evaluation shifts for time spent at each feeding station (P = 0.0407), number of feeding stations per minute (P = 0.0819) and number of steps per minute (P = 0.0302; ). In the morning shift, time spent at each feeding station was similar (10.2 s), regardless of the phenological stage. The number of feeding stations per minute was 0.6 feeding station (8.9%) lower in the vegetative stage when compared to the reproductive stage. At pre-reproductive, the number of feeding stations visited per minute was similar to other phenological stages. Displacement per minute at the vegetative stage was similar to other phenological stages. The number of steps per minute was 2.0 steps (18.5%) higher at reproductive as compared to the pre-reproductive stage.
Figure 2. Use of feeding station and p displacement pattern of heifers in cool season pasture grazing during the phenogical stages (vegetative, pre-reproductive and reproductive).
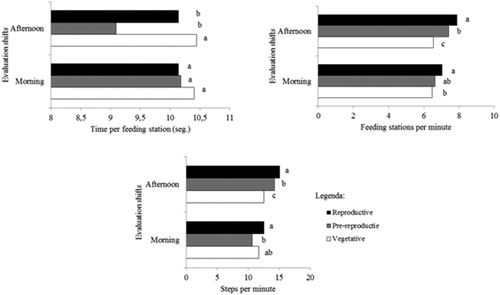
In the afternoon, time spent at each feeding station was 0.8 s (8.6%) longer in the vegetative stage compared to other stages. In this shift, the time at each station was similar (9.6 s) in the stages of pre-reproductive and reproductive. The time in each feeding station, in the vegetative stage, was negatively associated with the forage mass (r = −0.30; P = 0.0119). The number of feeding stations per minute was 1.5 (20.7%) and the number of steps per minute was 2.6 (20.5%) lower in the vegetative when compared to the reproductive stage. These variables showed intermediate values at the pre-reproductive stage. The number of feeding stations per minute was positively correlated with dead material mass (r = 0.44; P = 0.0046).
There was no interaction between phenological stages × evaluation shifts for the number of steps between feeding stations (P = 0.4621). The number of steps between stations (1.9) was similar, regardless of the phenological stage. The number of steps between stations was 0.2 step (11.8%) lower (P < 0.0001) in the morning when compared to the afternoon. The number of bites per feeding station was 1.4 bites (23.6%) higher in the vegetative stage when compared to the reproductive stage. The number of bites in each feeding station was intermediate at the pre-reproductive stage. This variable, in the vegetative stage, was explained by the stem mass (SM; r = 0.32) and dead material mass (r = 0.10), according to equation: Ŷ = 9.658–0.004SM – 0.0016DMM; P = 0.0296; r 2 = 0.42. At pre-reproductive, the number of bites per feeding station was explained by DM in situ digestibility (DMD), according to the equation: Ŷ = 20.12–0.17DMD; P < 0.0001; r 2 = 0.75. At the reproductive stage, this variable was explained by leaf:stem ratio (LSR; r = 0.13) and dead material mass (DMM; r = 0.22), according to equation: Ŷ = 5.841 + 1.0606LSR – 0.002DMM; P = 0.0036; r 2 = 0.35. At pre-reproductive, the average daily gain was positively correlated with duration of meal in the afternoon (r = 0.37; P = 0.0255). The average daily gain of heifers at the reproductive stage was negatively correlated with the dead material mass (r = −0.41; P = 0.0004).
Discussion
In the experiments, considering the forage mass (FM) greater than 1500 kg DM ha−1 (Hodgson Citation1990) and canopy height above 9 cm (Prache and Peyraud Citation2001), there was no limitation to forage intake by heifers. The change in stocking rate, according to the phenological stages evaluated, resulted from the management adopted to maintain the forage mass between 1500 and 2000 kg DM ha−1. This FM allowed to keep, in all growth stages, the mass of leaf blade above 350 kg ha−1, which does not limit the forage intake (Silva et al. Citation2005). It is expected a reduced participation of this component in the canopy with advancing of phenological stages (Drescher et al. Citation2006).
The change in the chemical composition of forage grazed, in the phenological stages investigated, results from decreasing the amount of cell content, rich in nitrogen (N), whose degradation is more rapid, and increased cell wall proportion (Prache and Peyraud Citation2001). The leaf blades are the structure of plants with greater concentration of N (Van Soest Citation1994). In this way, there is a reduction in crude protein content in forage as grazed (). The positive association between neutral fiber detergent content and dead material mass, and between in situ DM digestibility and leaf blade mass may explain the higher neutral detergent fiber content and the decrease in forage digestibility at reproductive stage.
Moreover, the bite rate is within the range of 30–70 bites minute−1 observed in herbivorous animals (Carvalho et al. Citation2001). At the reproductive stage, increasing forage mass is mainly associated with the largest number of stems. In this condition, the animals need more time to find and perform the bite. Stems require longer intervals between bites for chewing and handling of the harvested forage, which leads to longer intervals between bites (Baggio et al. Citation2009), resulting in decreased bite rate at this stage. The activities of harvesting the bite it and chewing are the grazing activities whose energy cost is higher (Gregorini Citation2012).
Heifers spent 37.8% time in grazing, regardless of the phenological stage, confirming that most herbivorous animals spend about 1/3 daily time in grazing (Carvalho et al. Citation2001). The grazing time is below the range of time cited by Krysl and Hess (Citation1993), from 359 to 771 min., in a wide range of ambient temperatures, supplementation schemes, grazing management and forage types.
The high content of soluble carbohydrates and low content of structural carbohydrates at the vegetative stage (Baumont et al. Citation2000) determine the shortest daily time in rumination. Thus, animals harvest forage with lower content of neutral detergent fiber and rumination time is shorter (). The time in other activities is related to the excluding character of activities of the feeding behavior, and these activities are performed in finite time (time scale of days; Carvalho et al. Citation2001).
A similar duration of meal, in the morning shift, independent from the phenological stage of the pasture, can be explained by the emptying of the gastrointestinal tract, as this variable is positively associated with rumination time on the night shift. This emptying causes hunger signals, since the animals spend only 20% of night time in grazing. In response to hunger, animals reduce their selectivity (De Paula et al. Citation2009). According to Van Soest (Citation1994), rumination time is influenced by the cell wall content of the forage, which is lower in the vegetative stage, reducing its retention time in the rumen (Decruyenaere et al. Citation2009).
In agreement with Prache and Peyraud (Citation2001), the presence of dead material in the canopy acts as a barrier to the bite depth. Thus, in the afternoon, at pre-reproductive, heifers increase the time spent in grazing in response to the modified structure of the pasture (), whereas the bite rate is similar to that observed in the vegetative stage (). Animals analyse the profitability of grazing and reduce the time when this is not energetically advantageous (Prache and Peyraud Citation2001) and this may have occurred at the reproductive stage, during this shift, depending on the existing canopy structure ().
In the vegetative and pre-reproductive stages, in the afternoon, meals were longer and it was expected a more efficient use of forage, characterized by greater nutritional value in this shift (Gregorini Citation2012). Hence, the reduction in cellular content and, consequently, in digestibility () and the increase in the amount of cell wall can lead to a more rapid filling of the rumen (Baumont et al. Citation2000). Thus, the duration of meal, in this shift, is reduced at the reproductive stage.
Higher leaf: stem ratio () in the vegetative stage is related to the time spent in the feeding station, which is determined by the amount of leaf blades (Prache and Roguet Citation1996). Probably, the longer time in each feeding station at this stage, in the afternoon, is because the herbivorous animals, by recognizing the site as rich in potential bites (Charnov Citation1976), remains in the feeding station until forage depletion and the perception or expectation to find other stations motivate it to change the feeding station (Baumont et al. Citation2000).
According to Ogura et al. (Citation2004), the number of feeding stations is related to the time spent at each feeding station. The increase of dead material mass at reproductive provided an increased number of feeding stations in the afternoon. In these situations, the animals needed to increase the total number of daily feeding stations in order to maintain the rate of forage consumption (Baggio et al. Citation2009).
Furthermore, the sum of the percentages relative to the grazing time in the morning (38%) and afternoon (43%) equates to 81% of the total daily grazing time, regardless of the phenological stage. Thereby, the night grazing contributes little to the daily forage intake and this behavior is related to the strategy to avoid predators (Gregorini Citation2012).
The change in the canopy structure, in the evaluated stages (), was not enough to alter the number of meals, which is an indicator of the pastoral environment (Carvalho et al. Citation2008). The beginning of the next meal, in animals kept under continuous grazing, is triggered by the emptying of the gastrointestinal tract (Gregorini Citation2012). The better quality of forage in the afternoon (Gregorini Citation2012) probably led heifers to empty faster their gastrointestinal tract and thereby provide the accomplishment of a greater number of meals in this shift.
The shorter grazing in the night shift, at the pre-reproductive stage, corresponds to the longer diurnal grazing time, because the ingestive behavior reflects the variations in consumption and quality of forage (Stobbs Citation1975). Thus, the night shift, at this stage, is mainly used for rumination and/or other activities. In the night shift of the vegetative and reproductive stages, the longer time in grazing is probably because heifers have not maximized their rumen filling during the day (Gregorini et al. Citation2008). In the first evening hours, when predation risk increases, the animal grazes to seek a stable relationship of nutrients (Gregorini Citation2012). This may explain the similarity in the duration of meal in this shift, regardless of the phenological stage of the grass.
The interval between meals is used for activities like rumination, rest, water intake, social interactions, among other activities. With the increased amount of dead material and stem in forage mass and decreased leaf: stem ratio, through the vegetative to the reproductive stage (), the access to leaf blade is impaired (Baggio et al. Citation2009) and may have resulted in longer time interval between one bite and another. This prevents diet selection within a single feeding station. Thus, heifers decrease the number of bites in each feeding station () and the chemical composition of forage as grazed is changed in the phenological stages ().
According to Prache et al. (Citation1998), bite mass is independent from the phenological stage of the grass and is associated with the mass of leaf blades in the canopy. The value of leaf blade mass (373.1 kg DM ha−1, ), at the reproductive stage, did not limit the bite mass. The displacement of herbivorous animals between each feeding station is associated with the bite mass in the last feeding station (Carvalho and Moraes Citation2005), which may explain the similar displacement between the feeding stations in the phonological stages evaluated.
Herbivorous animals seek to be efficient in their displacements (Carvalho and Moraes Citation2005), and the highest quality of forage in the afternoon (Gregorini Citation2012) provides more time for heifers to choose and travel to a new feeding station. In accordance with Roguet et al. (Citation1998), the greater displacement between feeding stations corresponds to greater selectivity of forage by the animals.
Additionally, lower weight gain at the vegetative stage may be due to the low DM content of the forage associated with low intake of heifers (Pilau et al. Citation2005). The afternoon shift corresponds to the higher intake of DM (Gregorini et al. Citation2008), therefore, heifers increase the duration of meal in this shift, which may have resulted in higher daily weight gain of heifers at the pre-reproductive stage. Changes occur throughout the phenological cycles and the canopy structure may deteriorate in a way that the animal performance decreases at the end of the season, which may explain the association between average daily gain and dead material mass at the reproductive stage.
The advance in phenological stages entails changes in canopy structure and chemical composition of forage as grazed. Heifers modify the ingestive behavior, duration of meal and interval between meals, forage selection patterns and number of steps per minute depending on the phenological stage and evaluation shifts, and use them as strategies to keep a similar bite mass. The reduction in bite rate is related to the presence of stems in the canopy. In the afternoon shift, the longer time grazing at the pre-reproductive stage is determined by the dead material mass, and the shorter duration of meal at the reproductive stage is associated with the leaf blade mass and in situ dry matter digestibility.
Changes in canopy structure caused no change in the number of meals and displacement between the feeding stations. The average daily gain at the pre-reproductive stage is associated with longer duration of meal in the afternoon.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Baggio C , Carvalho PCF , Silva JLS , Anghinoni I , Lopes MLT , Thurow JM. 2009. Padrões de deslocamento e captura de forragem por novilhos em pastagem de azevém-anual e aveia-preta manejada sob diferentes alturas em sistema de integração lavoura-pecuária. R Bras Zootec. 38:215–222. doi: 10.1590/S1516-35982009000200001
- Baumont R , Prache S , Meuret M , Morand-Fehr P. 2000. How forage characteristics influence behaviour and intake in small ruminants: a review. Livest Prod Sci. 64:15–28. doi: 10.1016/S0301-6226(00)00172-X
- Carvalho PCF , Da Silva SC , Nabinger C , Moraes AD , Genro TC. 2008. Managing natural grasslands in a changing world: grazing ecology insights to accomplish re-oriented management expectations. XXI International Grassland Congress and VIII International Rangeland Congress Hohhot, China. p. 415–421.
- Carvalho PCF , Moraes A. 2005. Comportamento ingestivo de ruminantes: bases para o manejo sustentável do pasto. Maringá : Manejo Sustentável em Pastagem; p. 1–20.
- Carvalho PCF , Ribeiro Filho HMN , Poli CHEC , Morais A , Delagarde R , 2001. Importância da estrutura da pastagem na ingestão e seleção de dietas pelo animal em pastejo. 39ª Reunião Anual da Sociedade Brasileira de Zootecnia, Piracicaba (Brazil), pp. 853–871.
- Charnov EL. 1976. Optimal foraging, the marginal value theorem. Theor Popul Biol. 9:129–136. doi: 10.1016/0040-5809(76)90040-X
- Chilibroste P , Gibb MJ , Soca P , Mattiauda DA. 2015. Behavioural adaptation of grazing dairy cows to changes in feeding management: do they follow a predictable pattern? Anim Prod Sci. 55:328–338. doi: 10.1071/AN14484
- Decruyenaere V , Buldgen A , Stilmant D. 2009. Factors affecting intake by grazing ruminants and related quantification methods: a review. Biotechnol Agron Soc Envir. 13:559–573.
- Demarquilly C , Chenost M , Aubry J , Chevalier D , Chenost MJ. 1969. Etude de la digestion des fourrages dans le rumen par la methods des sachets de nylon; liaisons avec la valeur alimentaire. Annales de Zootechaie. 18:419–430. doi: 10.1051/animres:19690405
- De Paula EFE , Stupak EC , Zanatta CP , Poncheki JK , Leal PC , Monteiro ALG. 2009. Comportamento ingestivo de ovinos em pastagens: Uma revisão. R Tróp Ci Agr Biol. 4:42–51.
- Drescher M , Heitkönig IMA , Raats JG , Prins HT. 2006. The role of grass stems as structural foraging deterrents and their effects on the foraging behaviour of cattle. Appl Anim Behav Sci. 101:10–26. doi: 10.1016/j.applanim.2006.01.011
- EMPRESA BRASILEIRA DE PESQUISA AGROPECUÁRIA – EMBRAPA , editor. 2006. Sistema Brasileiro de Classificação de Solos. Brasília: Embrapa Produção de Informação/Rio de Janeiro, Embrapa Solos. 412 pp.
- Euclides VPB , Macedo MCM , Oliveira MP. 1992. Avaliação de diferentes métodos de amostragem para estimar o valor nutritivo de forragens sob pastejo. Rev Bras Zootecn. 21:691–701.
- Glienke CL , Rocha MG , Roso D , Pötter L , Costa VG , Machado JM. 2010. Ingestive behavior and displacement patterns of beef heifers on Italian ryegrass pasture. R Bras Zootec. 39:247–254. doi: 10.1590/S1516-35982010000200003
- Gregorini P. 2012. Diurnal grazing pattern: its physiological basis and strategic management. Animal Prod Sci. 52(7):416–430. doi: 10.1071/AN11250
- Gregorini P , Gunter SA , Beck PA , Soder KJ , Tamminga S. 2008. Review: The interaction of diurnal grazing pattern, ruminal metabolism, nutrient supply, and management in cattle. Prof Anim Sci. 24:308–318. doi: 10.15232/S1080-7446(15)30861-5
- Hodgson J. 1990. Grazing management: science into practice. London : Longman Scientific & Techinical; 203 pp.
- Jamieson WS , Hodgson J. 1979. The effect of daily herbage allowance and sward characteristics upon the ingestive behaviour and herbage intake of calves under strip-grazing management. Grass Forage Sci. 34:261–271. doi: 10.1111/j.1365-2494.1979.tb01478.x
- Krysl LJ , Hess BW. 1993. Influence of supplementation on behavior of grazing cattle. J Anim Sci. 71:2546–2555. doi: 10.2527/1993.7192546x
- Laca EA , Ungar ED , Seligman NG , Ramey MR , Demment MW. 1992. An integrated methodology for studying short term grazing behaviour of cattle. Grass Forage Sci. 47:81–90. doi: 10.1111/j.1365-2494.1992.tb02250.x
- Lovatto PA , Lehnen CR , Andretta I , Carvalho AD , Hauschild L. 2007. Meta-análise em pesquisas científicas - enfoque em metodologias. R Bras Zootec. 36:285–294. doi: 10.1590/S1516-35982007001000026
- Ogura SI , Sekino R , Hirata M. 2004. Foraging behavior of cattle in a bahiagrass (Paspalum notatum Flugge) pasture: investigation at a feeding station scale. Jap Jou Grass Sci. 50(2):147–153.
- Pedroso CES , Medeiros RB , Silva MA , Jornada JBJ , Saibro JC , Teixeira JRF. 2004. Comportamento de ovinos em gestação e lactação sob pastejo em diferentes estádios fenológicos de azevém anual. R Bras Zootec. 33:1340–1344. doi: 10.1590/S1516-35982004000500028
- Pilau A , Rocha MG , Restle J , Silva JHS , Freitas FK , Macari S. 2005. Desenvolvimento de novilhas de corte recebendo ou não suplementação energética em pastagem com diferentes disponibilidades de forragem. R Bras Zootec. 34:1483–1492. doi: 10.1590/S1516-35982005000500008
- Prache S , Peyraud J. 2001. Foraging behaviour and intake in temperate cultivated grasslands. In: JA Gomide, WRS Mattos, SC Da Silva, editor. 19th International Grassland Congress, São Pedro (Brazil); p. 309–319.
- Prache S , Roguet C. 1996. Influence de la structure du couvert sur le comportement d´ingestion. Clermont-Ferrand : Institut National de la Recherche Agronomique; pp. 22–24.
- Prache S , Roguet C , Petit M. 1998. How degree of selectivity modifies foraging behaviour of dry ewes on reproductive compared to vegetative sward structure. Appl Anim Behav Sci. 57:91–108. doi: 10.1016/S0168-1591(97)00110-X
- Roguet C , Prache S , Petit M. 1998. Feeding station behaviour of ewes in response to forage availability and sward phenological stage. Appl Anim Behav Sci. 56:187–201. doi: 10.1016/S0168-1591(97)00104-4
- Senger CD , Kozlosky GV , Sanches LMB , Mesquita FR , Alves TP , Castagnino DS. 2008. Evaluation of autoclave procedures for fibre analysis in forage and concentrate feedstuffs. Anim Feed Sci Technol. 146:169–174. doi: 10.1016/j.anifeedsci.2007.12.008
- Silva ACF , Quadros FLF , Trevisan NB , Martins CEN , Bandinelli DG. 2005. Alternativa de manejo de pastagem hibernal: níveis de biomassa de lâmina foliar verde. R Bras Zootec. 34:472–478. doi: 10.1590/S1516-35982005000200014
- Stobbs TH. 1975. The effect of plant structure on the intake of tropical pasture. III. Influence of fertilizer nitrogen on the size of bite harvested by Jersey cows grazing Setaria anceps cv. Kazungula swards. Aust J Agric Res. 26:997–1007. doi: 10.1071/AR9750997
- Van Soest PJ , ed. 1994. Nutritional ecology of the ruminant. Ithaca (NY ): Cornell University Press; 476 pp.