
ABSTRACT
The objective of this study was to compare the diet selection and productive performance of beef cattle (Asturiana de los Valles breed) and crossbred horses grazing on heather-gorse shrublands with different available surface of improved pasture (IP): 80% (IP80) or 25% (IP25). Twelve and 10 lactating dams per species with their offspring were managed in IP80 (19 ha) and IP25 (22 ha), respectively, during two grazing seasons (spring-autumn). Visual recordings at IP25 indicated that horses grazed for longer than cattle. Both species grazed preferentially on IP and increased the utilization of shrublands as season advanced and available sward height in the IP area decreased. Diet composition estimates using faecal markers showed greater dietary percentages of white clover in cows than in mares during spring-summer. During autumn cows selected greater percentages of heather and lower of gorse than mares. Mares achieved more favourable body weight (BW) changes than cows during the whole grazing season, although differences were mainly confined to IP80. Calves showed higher BW gains than foals during the whole grazing season, with scarce differences between vegetation mixtures. Mares compete with cattle for the use of quality pastures, negatively affecting BW recoveries of cows when both species graze together.
1. Introduction
Large herbivores have replaced small ruminants in less-favoured areas of Cantabrian Mountains in the NW of the Iberian Peninsula, dominated by heathlands, despite the more efficient use of this poor quality vegetation by sheep and goats (lower intake per metabolic weight, higher digestibility of selected diets, and higher performance because of their lower body size and absolute nutrient requirements) (Celaya et al. Citation2008; Osoro et al. Citation2017). In these plant communities, as in other shrublands, the limiting factor for livestock performance is not usually the amount of available phytomass, but its poor nutritive quality (Celaya et al. Citation2011). Thus, maintaining profitable grazing systems in these less-favoured areas requires livestock access to certain areas with improved pasture (IP) to better meet the nutritional needs of animals. The presence of grassland patches within heathland areas together with mixed species grazing could promote a more efficient use of vegetation according to the diet selection of different herbivore species (Wright and Connolly Citation1995), in addition to higher levels of biodiversity (Rosa García et al. Citation2010; Fraser et al. Citation2014).
In general, cattle and horses graze preferentially on similar habitats, composed of herbaceous communities such as grasslands (Putman et al. Citation1987; Gordon Citation1989; Menard et al. Citation2002), showing a high degree of overlap in their diets (Hansen et al. Citation1977; Lamoot et al. Citation2005; Ferreira et al. Citation2013), which implies a great potential for competition for the same feed resource. Despite this high overlap, the differences in behaviour and digestive physiology between cattle and horses (Demment and Van Soest Citation1985; Duncan et al. Citation1990) could indicate certain complementarity in the use of heterogeneous vegetation under mixed grazing (Bullock and Armstrong Citation2000; Loucougaray et al. Citation2004). As some differences are observed, mainly in relation to the use of shrub communities (Pratt et al. Citation1986; Gordon Citation1989; Celaya et al. Citation2011), it could be possible to achieve sustainable grazing systems with cattle and horses in grassland-heathland mosaics. However, there is little information comparing the foraging behaviour and performance of these large herbivores in those mixed communities to assess the possible advantages (complementarity) or disadvantages (competition) of this management strategy.
The aim of this study was to compare the diet selection and productive performance of lactating beef cows and mares grazing with their offspring during spring-autumn season at different vegetation mixtures regarding the dominant plant community, i.e. heather-gorse shrubland or IP.
2. Materials and methods
2.1. Study site
The experiment was conducted between 2010 and 2013 at the Carbayal Research Station, Sierra de San Isidro, Illano, western Asturias (NW Spain, 43° 19′ N; 6° 54′ W), located at 900–1000 m above sea level. The climate is humid temperate but with heavier rains and colder temperatures than in lowlands because of the elevation and exposure. Mean annual precipitation recorded at the site during the study years was 1649 mm, mostly occurring from October to June. Mean annual temperature was 9.3°C, with a minimum monthly mean of 3.4°C in February and a maximum monthly mean of 15.8°C in August ().
Table 1. Monthly mean temperatures and rainfalls at the Carbayal Research Station (Asturias, Spain) in the four study years and during the experimental grazing season (GS: from April-May to October-November).
The characteristic plant community in the area is a heather-gorse shrubland dominated by heather species (Erica cinerea L., Erica umbellata Loefl. ex L. and Calluna vulgaris (L.) Hull, among others) and western gorse (Ulex gallii Planch.), a thorny and woody legume. The most abundant herbaceous species are grasses such as Pseudarrhenatherum longifolium Thore (Rouy) and Agrostis curtisii Kerguélen, covering the gaps remaining among the shrubs.
2.2. Experimental design and animal management
The experimental design consisted of two herbivore species (cattle and horses) grazing together on two plots with different available surface of IP: 80% (IP80) or 25% (IP25). The rest of the area was heather-gorse shrubland. Pasture surface was mechanically cleared using a brush cutter, and improved by ploughing, dressing and sowing perennial ryegrass (Lolium perenne L.) and white clover (Trifolium repens L.). Beef cows (Asturiana de los Valles breed, 512 ± 11.6 kg body weight – BW) and crossbred horses (mostly Cabalo Galego de Monte, 320 ± 7.4 kg BW) were managed during two grazing seasons in each vegetation mixture. In IP80 (19 ha), during two consecutive years (2010 and 2011), 12 cows and 12 mares with their offspring were managed from early May to late November at stocking rates of 1.3 adult animals/ha. In IP25 (22 ha), during two consecutive years (2012 and 2013), 10 cows and 10 mares with their offspring were managed from late April to mid-October or mid-November at stocking rates of 0.9 adult animals/ha. In 2013 cattle were taken off the plot 20 days before horses because of symptoms of undernourishment.
All adult animals had grazed heather-gorse vegetation with IP during the preceding years and were familiar with the plant species present in the experimental plots. Foals and calves were born in winter-early spring and reared with their mothers until the experimental grazing season began when they were aged 1–4 months. All animals were dosed against gastrointestinal nematodes and other parasites 2 weeks before turn out. Mares were dosed with ivermectin (Equimel©, Virbac group, Esplugues de Llobregat, Spain) by oral administration, using a dosing rate of 0.2 mg/kg BW. Cows were orally dosed with netobimin (Hapasil©, Schering-Plough, Segré, France), using a dosing rate of 0.13 mL/kg BW. Fresh water was available at both plots. No mineral supplements or additional feeds were given during the experiment. All procedures were carried out in accordance with European Union Directive 2010/63/EU on the protection of animals used for scientific purposes.
2.3. Vegetation measurements
Samples of the main plant components, i.e. heather, gorse, herbaceous plants (mostly grasses) growing in the heathland area, and IP herbage, were collected by hand-plucking (one sample per component and plot) three times each year (i.e. June, August and October) for chemical analysis to estimate nutritive quality. Samples were analysed following the procedures of the Association of Official Analytical Chemists (AOAC Citation2006) for ash (no. 942.05) and nitrogen (N, no. 990.03). Crude protein (CP) was calculated as N × 6.25. Neutral detergent fibre (NDF) was analysed according to the procedures described by Mertens (Citation2002). Acid detergent fibre (ADF, no.973.18) was analysed following the methods of AOAC handbook (2006), and expressed inclusive of residual ash. Lignin (sa) was determined by solubilizing cellulose with sulphuric acid (Robertson and Van Soest Citation1981). All contents are given as concentrations on dry matter (DM) basis.
In IP80 botanical composition was sampled in June 2011 by recording 250 vertical hits at 50 random sites in the shrubland area, and 500 vertical hits at 100 random sites in the IP area with a point-quadrat (Grant Citation1981) to obtain cover percentages. In IP25 botanical composition was sampled both years in June with the same apparatus by recording 1000 hits at 200 random sites in the shrubland areas, and 500 hits at 100 random sites in the IP area. Green pasture availability was assessed fortnightly during the grazing season by measuring the sward surface height at 100 random sites on the IP area using a calibrated swardstick (Barthram Citation1986). Sward heights from each plot and year were averaged across time for spring, summer, autumn, and the whole grazing season.
2.4. Diet selection and grazing time
The composition of the diet selected by the animals was estimated at different times using n-alkanes and long-chain alcohols (LCOH) as faecal markers (Dove and Mayes Citation2006). Samplings were performed in mid-June, late August, early October and early November 2011 in IP80; and in mid-June, late August and lat|e October 2012, and late July and early October 2013 in IP25. Several logistic problems prevented us from carrying out samplings reliably in 2010. On each occasion, one faecal grab sample from individual cows and mares was collected. At the same time, samples of the following potential dietary components were collected by hand-plucking: white clover, IP grasses, heather, gorse and heath-grasses. All samples were stored at −20°C and then freeze-dried and milled through a 1-mm pore-size mesh before analytical procedures. n-Alkanes (from C21 to C36) and LCOH (from C20-OH to C30-OH) from plant and faecal samples were extracted using the procedures described by Dove and Mayes (Citation2006), and their concentrations determined by gas chromatography, using a Perkin Helmer Clarus 580, equipped with flame ionization detector (FID) and an auto-sampler. Identification was performed by comparison of retention times of the samples components with the previously injected mixture of n-alkanes and of LCOH. The response factors for the individual n-alkanes and LCOH were calculated from peak areas and the known concentrations. The n-alkane concentrations were quantified relative to known amounts of the internal standards C22 (n-docosane) and C34 (n-tetratriacontane), whereas C27-OH (1-heptacosanol) was used as the internal standard for the LCOH quantification. All internal standards were added to the samples at the beginning of the extraction process.
The proportions of the plant components in the diet were estimated for each animal using an iterative least squares procedure that minimizes the discrepancies between the observed concentrations of each n-alkane (from C25 to C33) and LCOH (even-chain ones from C20-OH to C30-OH) in the faeces, and the estimated proportions of plant components in the diet, according to Salt et al. (Citation1994), using the Solver routine in Microsoft Excel. The other n-alkanes and LCOH were not used in the calculations due to their low concentrations in all dietary components. Before calculations, cows’ marker faecal concentrations were corrected for incomplete faecal recovery, according to data obtained in previous validation studies with cattle fed similar diets (López López et al. Citation2015). Mares’ marker faecal concentrations were not adjusted as previous studies (Ferreira et al. Citation2007; López López et al. Citation2015) indicated that the faecal recovery of these markers is unaffected by their carbon-chain length in this animal species.
At IP25, the time spent grazing by each animal species on each vegetation type (heathland and IP) was determined by recording the grazing activity of the animals every 15 min from dawn to dusk during two consecutive days on two occasions in 2012 (late June and early September) and once in 2013 (mid-August). At IP80, simultaneous visual recording of all the animals in the plot was impracticable because of the configuration of the terrain.
2.5. Animal performance
All animals were periodically weighed during the grazing season. Daily BW changes were calculated for spring, summer, autumn, and for the whole grazing season. Given the natural differences in BW existing between the cattle and horse breeds studied, daily BW changes were also calculated as percentages in relation to initial BW (i.e. in BW%/day).
2.6. Statistical analysis
Data on plant chemical composition (CP, NDF, ADF and lignin concentrations) were subjected to a mixed model ANOVA for repeated measures to test the fixed effects of plant component (C: heather, gorse, heath-grasses and IP herbage), vegetation mixture (V: IP80 and IP25), season (S: spring, summer and autumn as repeated measures), and the full interactions. Mean IP sward heights during each season and the whole grazing season were analysed with 1-way ANOVA for V effect. Sward height data disaggregated by seasons were analysed to test the effects of V, S (three repeated measures) and the interaction V × S. n-Alkane and LCOH concentrations in plant components were analysed with 2-way ANOVA to examine the differences between the diet components, including the effect of season and their interaction.
Diet composition data (percentages of white clover, IP grasses, heather, gorse and heath-grasses) from individual animals were angular-transformed and analysed within each year with a mixed model ANOVA, including the effects of livestock species (Sp), S (four, three and two repeated measures for 2011, 2012 and 2013, respectively), and the interaction Sp × S. Back-transformed least-squares means and standard errors (SEM) are presented for dietary percentages. For individual data on daily BW changes (dams or offspring) during each season and the whole grazing season, the model included the fixed effects of Sp, V, the interaction Sp × V, and the random effect of year nested within V. Differences between means were examined by Tukey's test. All analyses were performed using Statistica 8.0 software (StatSoft, Inc., Tulsa, OK).
3. Results
3.1. Available vegetation
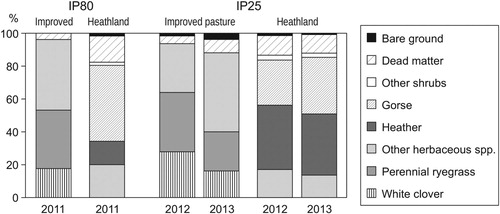
At IP80, IP in June 2011 was dominated by common bent (Agrostis capillaris L., a widespread grass in acid grasslands) and perennial ryegrass (38 and 36%, respectively), while white clover accounted for 18% cover. Heathland vegetation was dominated by gorse (37%), with heather and herbaceous species covering 21 and 20%, respectively (). At IP25, a greater percentage of white clover was found in the IP in 2012 (28%), but both clover and ryegrass cover diminished in the next year, whereas native species (mostly common bent) increased its percentage. Heathland was on average composed of 38% heather, 31% gorse and 15% herbaceous plants, with the rest being dead matter and bare ground ().
Figure 1. Botanical composition of heather-gorse shrublands and improved ryegrass-clover pastures in the experimental plots grazed by cattle and horses (IP80: 80% improved pasture; IP25: 25% improved pasture).
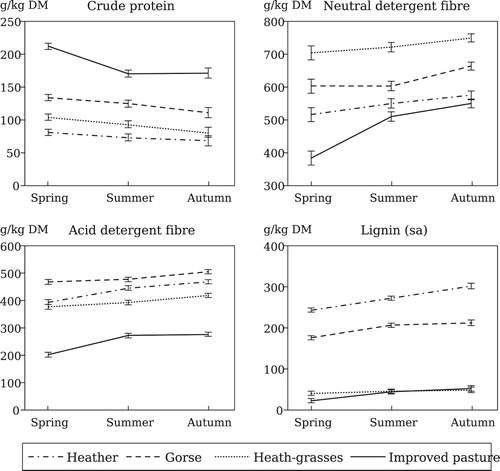
Chemical composition differed (P < 0.001) between the analysed plant components (). The IP herbage showed the highest CP content in all seasons, averaging 185 g/kg DM, followed by gorse and heath-grasses (123 and 92 g/kg DM, respectively), while heather showed the lowest CP contents (74 g/kg DM). Cell wall components represented the highest fraction in all plant components, with NDF ranging from a minimum of 384 g/kg DM (IP in spring) to a maximum of 750 g/kg DM (heath-grasses in autumn), while lignin (sa) showed values between 202 (IP in spring) and 506 g/kg DM (gorse in autumn). Large differences between herbaceous and woody species were observed in lignin content, which accounted for 175–300 g/kg DM in heather and gorse, and 21–51 g/kg DM in herbaceous species. Plot effect was not significant for any chemical variable. There were differences (P < 0.001) among seasons, with mean CP contents decreasing from spring to summer-autumn, whereas mean fibre fraction contents increased from spring to summer and to autumn. The seasonal variations were somewhat different between plant components (interactions C × S: P < 0.01 for lignin; P < 0.05 for CP, NDF and ADF; ).
Figure 2. Chemical composition of the green shoots and leaves of the main plant components of heather-gorse shrublands and improved pasture.
Mean sward height of the IP across the whole grazing season was higher in IP80 than in IP25 (7.40 vs. 6.43 cm; SEM = 0.093; P < 0.05). Sward height decreased in both plots as season advanced from a mean of 10.62 cm in spring to 6.45 cm in summer and 3.96 cm in autumn (SEM = 0.711; P < 0.01). Although V × S interaction was not significant, lower mean sward heights were observed in IP25 than in IP80 in summer (5.07 vs. 7.84 cm; SEM = 0.177; P < 0.01) and autumn (3.05 vs. 4.87 cm; SEM = 0.561; P = 0.148).
3.2. Foraging behaviour
Dietary plant components showed different concentrations of almost all n-alkanes and LCOH (). Heather was characterized by higher concentrations of most n-alkanes and shorter chain LCOH, whereas herbaceous species had higher total LCOH concentration than heather and gorse (P < 0.001). Concentrations of most markers did not vary greatly among seasons.
Table 2. n-Alkane and long-chain alcohol (LCOH) contents (mg/kg DM) of the main plant components (C) of heather-gorse shrublands (heather, gorse and grasses) and improved pasture (IP) at different seasons of the year (Spr, spring; Sum, summer; Aut, autumn).
The estimated animal diets at IP80 showed greater mean percentages of IP grasses (65.8 vs. 57.5%; SEM = 3.46; P < 0.05) and lower of gorse (1.1 vs. 7.4%; SEM = 1.30; P < 0.001) in cows than in mares. In general, IP components decreased (P < 0.001) while those from heathlands increased (P < 0.01) from spring-summer to autumn in the diets of both herbivore species, although Sp × S interactions were observed for all components (). In June and August, both species fed almost exclusively on IP, with cows showing greater percentages of white clover (23.6 vs. 11.3%; SEM = 2.49; P < 0.001) and lower of IP grasses (74.7 vs. 84.7%; SEM = 2.67; P < 0.001) than mares. However, these differences were somewhat reversed during autumn, with greater clover percentages estimated in mares’ diets in October (15.5 vs. 4.7%, SEM = 2.83; P < 0.01) and greater (P < 0.01) IP grass percentages in cows’ diets in October (67.1 vs. 37.5%; SEM = 6.99) and November (45.8 vs. 15.6%; SEM = 7.58). From October to November, dietary percentages of heath-grasses increased more in cows, whereas those of shrubs increased more in mares, resulting in especially greater percentages of gorse in mares’ diets late in the season (32.0 vs. 0.1%; SEM = 6.76; P < 0.001), in addition to greater heather percentages (7.4 vs. 1.1%; SEM = 1.28; P < 0.001) and lower heath-grass percentages (29.9 vs. 49.6%; SEM = 7.36; P < 0.05) compared to cows’ diets ().
Figure 3. Diet selection of cows (C) and mares (M) grazing on partially improved heathlands (IP80: 80% improved pasture; IP25: 25% improved pasture) across the grazing season.
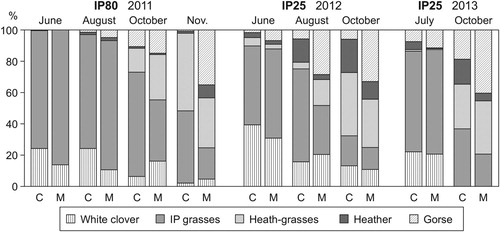
At IP25, in both years greater mean dietary percentages of IP grasses (42.5 vs. 29.1%; SEM = 5.38; P < 0.05 in 2012; 49.7 vs. 43.7%; SEM = 2.96; P = 0.083 in 2013) and heather (11.7 vs. 4.9%; SEM = 0.89; P < 0.001 in 2012; 9.2 vs. 1.8%; SEM = 1.10; P < 0.001 in 2013), and lower of gorse (3.5 vs. 18.8%; SEM = 2.20; P < 0.001 in 2012; 11.4 vs. 23.3%; SEM = 2.73; P < 0.01 in 2013) were estimated in cows than in mares (). As in IP80, heathland components increased as season advanced, with important Sp × S interactions being found, especially in 2012. Clover percentage in June was greater in cows than in mares (39.2 vs. 30.7%; SEM = 2.40; P < 0.01) and it decreased to similar values of 16 and 12% in August and October, respectively. For IP grasses, greater percentages were estimated in cows than in mares’ diets in August (59.4 vs. 26.5%, SEM = 9.07; P < 0.01). As season advanced, greater heather percentages and lower gorse percentages were found in cows than in mares in both years. Between-species differences in heath-grasses were subtle and variable among sampling seasons ().
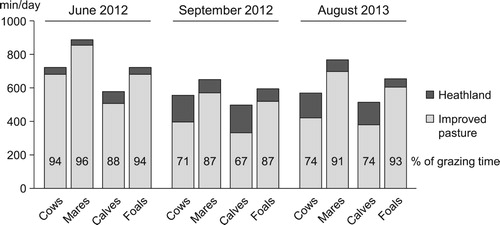
Regarding the visually recorded animals’ behaviour at IP25, mares were found to graze for longer than cows in the three observation periods (770 vs. 618 min/day). Mean daily grazing time was longest in June (806 min/day) and shortest in September (604 min/day). In June, both mares and cows spent around 95% of the daily grazing time on IP. In August and September, mares continued to graze mostly on IP (89%), whereas cows grazed on shrublands to a greater extent (28%; ). The grazing pattern of the offspring was similar to that of the dams, with foals grazing for longer (657 vs. 530 min/day) and proportionally less on shrublands (9 vs. 24% on average) than calves.
Figure 4. Time spent grazing by cows, mares, calves and foals on a partially (25%) improved heathland at different times of the year.
3.3. Animal performance
Animals’ daily BW changes and as percentages in relation to initial BW (BW%) across the whole grazing season differed between grazer species, with cows in general showing worse BW changes than mares (148 vs. 270 g/day; SEM = 28.7; P < 0.01; 0.03 vs. 0.09%/day; SEM = 0.007; P < 0.001). The main effect of available vegetation in the plot was not significant, although a Sp × V interaction was found (P < 0.01), as the more favourable BW changes in mares than cows were observed in IP80 but not in IP25 (). Daily BW and BW% changes were also affected by year within each vegetation mixture (P < 0.05). Differences in BW changes between animal species increased as season advanced. In spring, mares and cows showed a similar performance in both plots (mean gain of 1156 g/day). In summer, mares had better BW changes than cows (138 vs. 37 g/day; SEM = 32.2; P < 0.05). In autumn, both species showed negative BW changes, with mares presenting lower losses than cows (−322 vs. −820 g/day; SEM = 44.9; P < 0.001). Daily BW% changes showed a better performance of mares than cows in all seasons (P < 0.001 in spring and autumn; P < 0.01 in summer; ). Available vegetation influenced animal performance in spring, with greater BW (P < 0.05) and BW% (P = 0.051) gains in IP80 than in IP25, and in autumn, with lower BW% losses in IP80 than in IP25 (P < 0.05). In addition, an interaction Sp × V (P < 0.05) in spring was found for BW% change, indicating that the greater gains achieved by mares occurred especially in IP80 ().
Table 3. Daily body weight (BW) changes and as percentage of initial BW (BW%) of lactating cows and mares grazing in plots with different vegetation availability (IP80: 80% improved ryegrass-clover pasture, 20% heather-gorse shrubland; IP25: 25% improved ryegrass-clover pasture, 75% heather-gorse shrubland).
Calves and foals had positive BW gains throughout the whole grazing season, without significant differences between vegetation mixtures, although daily BW changes were affected by year within each plot (P < 0.001; ). In spring, daily BW gains were higher for foals than for calves (1124 vs. 989 g/day; SEM = 35.7; P < 0.01), whereas in summer and autumn, calves showed higher profits (914 vs. 707 g/day in summer; SEM = 22.6; P < 0.001; 435 vs. 278 g/day in autumn; SEM = 43.6; P < 0.05), which resulted in overall higher gains compared with foals (831 vs. 697 g/day; SEM = 22.6; P < 0.001). Examining BW% changes, although the differences between species across each season showed similar trends to changes in g/day (higher gains of foals than calves in spring, the reverse in summer and autumn), such differences were cancelled out during the whole grazing season (). There was a trend for higher gains in IP80 compared with IP25 (1.33 vs. 1.21%/day; SEM = 0.079; P = 0.053).
Table 4. Daily body weight (BW) changes and as percentage of initial BW (BW%) of calves and foals grazing in plots with different vegetation availability (IP80: 80% improved ryegrass-clover pasture, 20% heather-gorse shrubland; IP25: 25% improved ryegrass-clover pasture, 75% heather-gorse shrubland).
4. Discussion
4.1. Available vegetation and animals’ foraging behaviour
Data on the chemical composition showed the lower nutritive quality of heathland vegetation, particularly of heather, compared with the improved ryegrass-clover pasture, as reported in previous studies (Celaya et al. Citation2008; Ferreira et al. Citation2013), and pointed out the convenience of establishing certain IP areas available to grazing animals in order to supplement the diet offered by shrub vegetation and better meet their nutritional requirements (Celaya et al. Citation2011; Osoro et al. Citation2017).
Grazing times recorded at IP25 indicated that mares grazed for longer than cows, as earlier observed in different vegetation conditions (Arnold Citation1984; Menard et al. Citation2002; Lamoot et al. Citation2005; Ferreira et al. Citation2013). It is well known that horses, being hindgut fermenters, compensate the less efficient fibre digestion compared to ruminants by increasing daily intake at the cost of spending more time grazing (Demment and Van Soest Citation1985; Duncan et al. Citation1990). At the beginning of the grazing season, in spring, both herbivore species fed almost exclusively on IP areas, as grasslands are much preferred compared to shrubby communities (Pratt et al. Citation1986; Gordon Citation1989; Ferreira et al. Citation2013). Once sward surface height decreased over summer and autumn below 8 cm, considered near the optimal for cattle performance on ryegrass-clover pastures (Wright and Whyte Citation1989), cows became limited because of reduced bite depth and DM intake (Hodgson Citation1985), and thus reduced their grazing time on IP, as previously observed in a similar situation (Celaya et al. Citation2008), and made a greater use of heathlands than mares. Other studies have shown a higher use of scrub by cattle compared with horses (Gordon Citation1989; Lamoot et al. Citation2005), which benefit from their greater capacity to graze on short swards thanks to their dental anatomy (Menard et al. Citation2002; Hongo and Akimoto Citation2003). It is expected that cattle are forced to abandon short grasslands to feed on other communities before than horses (Menard et al. Citation2002; Cornelissen and Vulink Citation2015).
On the other hand, apart from their shorter grazing time (around 15% less), offspring showed a grazing behaviour very similar to that of their dams. Although there are very few reports on the grazing behaviour of nursing calves and foals, a high correlation between dams and their offspring is to be expected given the precociousness and follow-up learning behaviour of both calves and foals (Bailey and Lawson Citation1981; Carson and Wood-Gush Citation1983).
The greater proportional grazing time on heathlands in cows than in mares was not reflected in greater percentages of woody plants consumed by cows. This could be partly due to faeces samplings and visual recordings being not coincident in time, in addition to different diets selected by cows and mares within heathlands. Although differences in the percentages of heath-grasses were inconsistent, mares in general consumed higher gorse percentages than cows in both IP25 and IP80. Inversely, except in November 2011 at IP80, cows ate proportionally more heather than mares. Horses have been observed to have a good capacity to ingest the spiny shoots of gorse species (Putman et al. Citation1987; Ferreira et al. Citation2013; López López et al. Citation2017). However, they are more reluctant to browse on heather than cattle (Putman et al. Citation1987; Celaya et al. Citation2011). It has been hypothesized that horses are more constrained by plant secondary compounds than ruminants (Duncan et al. Citation1990; Menard et al. Citation2002), so the tannins present in heather could be restricting its use by horses. Contrary to these chemical defences against herbivory, physical defences such as thorns seem to be less effective in horses than in cattle. The greater CP concentrations in gorse relative to heather makes this legume more palatable to horses, as equines do not recycle urea back to the digestive tract as efficiently as ruminants to support microbial protein synthesis (Santos et al. Citation2011).
Regarding the diet selection within IP, cows in general had greater dietary percentages of white clover than mares when herbage availability was high enough, i.e. in June and August 2011 in IP80 (mean sward heights of 7.5–8.6 cm) and in June 2012 in IP25 (mean sward height of 9.6 cm). Nevertheless, clover percentages in mares’ diets were similar to or even greater than in cows’ diets in other seasons with shorter sward heights. It is commonly accepted that horses prefer grasses over clover, but reliable evidence is very limited (Archer Citation1973; Hunt et al. Citation1989). In fact, Archer (Citation1973) observed that a grass-clover mixture was the most palatable forage for horses among 26 species (including grasses, legumes and herbs) or mixtures tested, and, though white clover was less palatable than most perennial ryegrass varieties, it was much more preferred than common bent (the co-dominant grass species in our IP). Our results suggest that horses eat white clover quite willingly without apparent rejection despite its potential toxicity by cyanogenic glycosides (McGorum et al. Citation2012).
4.2. Animal performance
Mares in general showed better performances than cows, despite the lower fibre digestive efficiency of horses compared with ruminants (Demment and Van Soest Citation1985; Cymbaluk Citation1990; Osoro et al. Citation2017). Horses are able to maintain high levels of intake thanks to a fast throughput rate of digesta, and are well adapted to utilize fibrous graminoids (Duncan et al. Citation1990). Particularly, mares showed much lower BW losses compared with cattle during autumn, when available IP sward height was very short, which seems to be related to horses’ ability to prehend foliage from short swards (Menard et al. Citation2002) and to make a better use of heath-grasses and gorse plants available in heathlands (Putman et al. Citation1987; Celaya et al. Citation2011).
Except in spring, calves achieved greater BW gains than foals during the whole grazing season, in agreement with other studies performed on heather-gorse shrublands with or without IP areas (Celaya et al. Citation2011; Osoro et al. Citation2017). The better performance of calves could be related to the ability of lactating cows to mobilize body reserves to maintain milk production (Osoro Citation1989). Although forage intake progressively substitutes calf reliance on milk intake after peak lactation (Le Du and Baker Citation1979), milk intake is still important to meet the energy needs of growing calves until weaning (Bailey and Lawson Citation1981). By contrast, lactating mares have a lower ability to mobilize body reserves and are unable to maintain milk production under conditions of nutritional deficiency (Deichsel and Aurich Citation2005). Therefore, the energy consumption by foals is reduced, adversely affecting their BW gains (Doreau and Boulot Citation1989).
Regarding the effects of available vegetation (grassland/heathland ratio) on animal performance, direct comparisons between plots should be cautiously taken as animals were managed at each vegetation mixture in different years. Although such comparisons could be affected by climatic differences among years, meteorological data indicate greater differences in mean temperatures and rainfalls between the two grazing seasons studied within each plot than between the pairs of years at each plot (i.e. 2010–2011 vs. 2012-2013; see ). The great inter-annual variability observed in animal performance restricted the significance of the effects of available vegetation on BW changes, with scarce superiority found for IP80 at particular seasons. Those differences would be related to the greater IP area in IP80 compared with IP25, resulting in lower grazing pressure and higher sward heights maintained in the former, particularly at the end of the grazing season, and thus mitigating BW losses in both cattle and horses (Aiken et al. Citation1989; Wright and Whyte Citation1989). It should be noted that grazing seasons were 17 days shorter on average in IP25 than in IP80 since the animals had to be housed before to be fed, thus reducing the differences between plots. The shorter grazing season in IP25 caused by the smaller IP area and a faster herbage depletion rate entails higher feeding costs during winter housing.
5. Conclusion
In view of the poor nutritive quality of heathland vegetation and the performance obtained by cattle and horses in these conditions, we can conclude the need for the livestock of good quality pasture surfaces for long grazing seasons (7–8 months). Both cows and mares spend most of their time grazing on IP, which means that with larger pasture surface available, for similar stocking rates, sward height is less reduced and BW losses in autumn are lower, which will increase their overall production capacity. In spite of the complementary use of heathland vegetation, mares compete strongly with cattle for the use of high quality pastures, negatively affecting BW recoveries of cows when both species graze together. As an alternative management, mares could remain with their offspring in heathland areas with available gorse and grasses until autumn, and then be moved to the improved areas previously grazed by cattle, to consume vegetation not used by ruminants, favouring a clean regrowth of quality pasture in the next spring.
Disclosure of interest
The authors report no conflict of interest.
Acknowledgments
The authors thank the staff of the Carbayal Research Station for their help in animal management and field work. This work was supported by the Spanish National Institute for Agricultural and Food Research and Technology (INIA) under Project RTA2010-00136-00-00 and European Regional Development Fund. Carlos López López was recipient of a predoctoral fellowship from INIA during the study.
Disclosure Statement
No potential conflict of interest was reported by the authors.
ORCID
Rafael Celaya http://orcid.org/0000-0002-7452-8233
Luis Miguel M. Ferreira http://orcid.org/0000-0002-3341-861X
Additional information
Funding
References
- Aiken GE , Potter GD , Conrad BE , Evans JW. 1989. Growth performance of yearling horses grazing bermudagrass pastures at different grazing pressures. J Anim Sci. 67(10):2692–2697. doi: 10.2527/jas1989.67102692x
- Archer M. 1973. The species preferences of grazing horses. J Br Grassld Soc. 28:123–128. doi: 10.1111/j.1365-2494.1973.tb00732.x
- Arnold GW. 1984. Comparison of the time budgets and circadian patterns of maintenance activities in sheep, cattle and horses grouped together. Appl Anim Behav Sci. 13:19–30. doi: 10.1016/0168-1591(84)90048-0
- Association of Official Analytical Chemists . 2006. Official methods of analysis, 18th ed. Gaithersburg, MD : AOAC.
- Bailey CB , Lawson JE. 1981. Estimated water and forage intakes in nursing range calves. Can J Anim Sci. 61:415–421. doi: 10.4141/cjas81-049
- Barthram GT. 1986. Experimental techniques: the HFRO swardstick. In: The Hill Farming Research Organisation biennial report 1984–85. Midlothian, UK : HFRO; p. 29–30.
- Bullock DJ , Armstrong HM. 2000. Grazing for environmental benefits. In: Rook AJ , Penning PD , editor. Grazing management. The principles and practice of grazing, for profit and environmental gain, within temperate grassland systems. Reading, UK : British Grassland Society; p. 191–200.
- Carson K , Wood-Gush DGM. 1983. Equine behaviour: I. A review of the literature on social and dam—foal behaviour. Appl Anim Ethol. 10:165–178. doi: 10.1016/0304-3762(83)90138-4
- Celaya R , Benavides R , García U , Ferreira LMM , Ferre I , Martínez A , Ortega-Mora LM , Osoro K. 2008. Grazing behaviour and performance of lactating suckler cows, ewes and goats on partially improved heathlands. Animal. 2:1818–1831. doi: 10.1017/S1751731108003224
- Celaya R , Ferreira LMM , García U , Rosa García R , Osoro K. 2011. Diet selection and performance of cattle and horses grazing in heathlands. Animal. 5:1467–1473. doi: 10.1017/S1751731111000449
- Cornelissen P , Vulink JT. 2015. Density-dependent diet selection and body condition of cattle and horses in heterogeneous landscapes. Appl Anim Behav Sci. 163:28–38. doi: 10.1016/j.applanim.2014.12.008
- Cymbaluk NF. 1990. Comparison of forage digestion by cattle and horses. Can J Anim Sci. 70:601–610. doi: 10.4141/cjas90-072
- Deichsel K , Aurich J. 2005. Lactation and lactational effects on metabolism and reproduction in the horse mare. Livest Prod Sci. 98:25–30. doi: 10.1016/j.livprodsci.2005.10.003
- Demment MW , Van Soest PJ. 1985. A nutritional explanation for body size patterns of ruminant and non-ruminant herbivores. Am Nat. 125:641–672. doi: 10.1086/284369
- Doreau M , Boulot S. 1989. Recent knowledge of mare milk production: a review. Livest Prod Sci. 22:213–235. doi: 10.1016/0301-6226(89)90057-2
- Dove H , Mayes RW. 2006. Protocol for the analysis of n-alkanes and other plant-wax compounds and for their use as markers for quantifying the nutrient supply of large mammalian herbivores. Nat Protoc. 1:1680–1697. doi: 10.1038/nprot.2006.225
- Duncan P , Foose TJ , Gordon IJ , Gakahu CG , Lloyd M. 1990. Comparative nutrient extraction from forages by grazing bovids and equids: a test of the nutritional model of equid/bovid competition and coexistence. Oecologia. 84:411–418. doi: 10.1007/BF00329768
- Ferreira LMM , Celaya R , Benavides R , Jáuregui BM , García U , Santos AS , Rosa García R , Rodrigues MAM , Osoro K. 2013. Foraging behaviour of domestic herbivore species grazing on heathlands associated with improved pasture areas. Livest Sci. 155:373–383. doi: 10.1016/j.livsci.2013.05.007
- Ferreira LMM , Garcia U , Rodrigues MAM , Celaya R , Dias-da-Silva A , Osoro K. 2007. The application of the n-alkane technique for estimating the composition of diets consumed by equines and cattle feeding on upland vegetation communities. Anim Feed Sci Technol. 138:47–60. doi: 10.1016/j.anifeedsci.2006.11.007
- Fraser MD , Moorby JM , Vale JE , Evans DM. 2014. Mixed grazing systems benefit both upland biodiversity and livestock production. PLoS ONE. 9(2):e89054. doi: 10.1371/journal.pone.0089054
- Gordon IJ. 1989. Vegetation community selection by ungulates on the isle of Rhum. II. vegetation community selection. J Appl Ecol. 26:53–64. doi: 10.2307/2403650
- Grant SA. 1981. Sward components. In: Hodgson J , Baker RD , Davies A , Laidlaw AS , Leaver JD , editor. Sward measurement handbook. Hurley, UK : British Grassland Society; p. 71–92.
- Hansen RM , Clark RC , Lawhorn W. 1977. Foods of wild horses, deer, and cattle in the Douglas Mountain area. Colorado. J Range Manage. 30:116–119. doi: 10.2307/3897750
- Hodgson J. 1985. The control of herbage intake in the grazing ruminant. Proc Nutr Soc. 44:339–346. doi: 10.1079/PNS19850054
- Hongo A , Akimoto M. 2003. The role of incisors in selective grazing by cattle and horses. J Agric Sci. 140:469–477. doi: 10.1017/S0021859603003083
- Hunt WF , Hay RJM , Clark D. 1989. Pasture species preferences by horses in New Zealand. In: XVI International Grassland Congress. Nice, France: Association Française pour la Production Fourragère; p. 797–798.
- Lamoot I , Meert C , Hoffmann M. 2005. Habitat use of ponies and cattle foraging together in a coastal dune area. Biol Conserv. 122:523–536. doi: 10.1016/j.biocon.2004.09.009
- Le Du YLP , Baker RD. 1979. Milk-fed calves: 5. The effect of a change in milk intake upon the herbage intake and performance of grazing calves. J Agric Sci. 92:443–447. doi: 10.1017/S0021859600062985
- López López C , Celaya R , Santos AS , Rodrigues MAM , Osoro K , Ferreira LMM. 2015. Application of long-chain alcohols as faecal markers to estimate diet composition of horses and cattle fed with herbaceous and woody species. Animal. 9:1786–1794. doi: 10.1017/S1751731115001196
- López López C , Ferreira LMM , García U , Moreno-Gonzalo J , Rodrigues MAM , Osoro K , Ferre I , Celaya R. 2017. Diet selection and performance of horses grazing on different heathland types. Animal. 11:1708–1717. doi: 10.1017/S1751731117000465
- Loucougaray G , Bonis A , Bouzilé JB. 2004. Effects of grazing by horses and/or cattle on the diversity of coastal grasslands in western France. Biol Conserv. 116:59–71. doi: 10.1016/S0006-3207(03)00177-0
- McGorum BC , Pirie RS , Fry SC. 2012. Quantification of cyanogenic glycosides in white clover (Trifolium repens L.) from horse pastures in relation to equine grass sickness. Grass Forage Sci. 67:274–279. doi: 10.1111/j.1365-2494.2011.00844.x
- Menard C , Duncan P , Fleurance G , Georges JY , Lila M. 2002. Comparative foraging and nutrition of horses and cattle in European wetlands. J Appl Ecol. 39:120–133. doi: 10.1046/j.1365-2664.2002.00693.x
- Mertens DR. 2002. Gravimetric determination of amylase treated neutral detergent fiber in feeds with refluxing in beaker or crucibles: collaborative study. J AOAC Int. 85:1217–1240.
- Osoro K. 1989. Manejo de las reservas corporales y utilización del pasto en los sistemas de producción de carne con vacas madres establecidos en zonas húmedas [management of body reserves and pasture utilization in suckler beef production systems established in humid areas]. Invest Agr Prod San Anim. 4:207–240. Spanish.
- Osoro K , Ferreira LMM , García U , Martínez A , Celaya R. 2017. Forage intake, digestibility and performance of cattle, horses, sheep and goats grazing together on an improved heathland. Anim Prod Sci. 57:102–109. doi: 10.1071/AN15153
- Pratt RM , Putman RJ , Ekins JR , Edwards PJ. 1986. Use of habitat by free-ranging cattle and ponies in the New Forest, southern England. J Appl Ecol. 23:539–557. doi: 10.2307/2404035
- Putman RJ , Pratt RM , Ekins JR , Edwards PJ. 1987. Food and feeding behaviour of cattle and ponies in the New Forest, Hampshire. J Appl Ecol. 24:369–380. doi: 10.2307/2403881
- Robertson JB , Van Soest PJ. 1981. The detergent system of analysis. In: James WPT , Theander O , editor. The analysis of dietary fiber in food. New York, NY : Marcel Dekker; p. 123–158.
- Rosa García R , Ocharan FJ , García U , Osoro K , Celaya R. 2010. Arthropod fauna on grassland-heathland associations under different grazing managements with domestic ruminants. C R Biol. 333:226–234. doi: 10.1016/j.crvi.2009.12.008
- Salt CA , Mayes RW , Colgrove PM , Lamb CS. 1994. The effects of season and diet composition on the radiocaesium intake by sheep grazing on heather moorland. J Appl Ecol. 31:125–136. doi: 10.2307/2404605
- Santos AS , Rodrigues MAM , Bessa RJB , Ferreira LM , Martin-Rosset W. 2011. Understanding the equine cecum-colon ecosystem: current knowledge and future perspectives. Animal. 5:48–56. doi: 10.1017/S1751731110001588
- Wright IA , Connolly J. 1995. Improved utilization of heterogeneous pastures by mixed species. In: Journet M , Grenet E , Farce MH , Thériez M , Demarquilly C , editor. Recent developments in the nutrition of herbivores. Paris : INRA Editions; p. 425–436.
- Wright IA , Whyte TK. 1989. Effects of sward height on the performance of continuously stocked spring-calving beef cows and their calves. Grass Forage Sci. 44:259–266. doi: 10.1111/j.1365-2494.1989.tb02163.x