
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Early and late season kudzu leaves were collected to analyse dry matter (DM), crude protein (CP), acid detergent fibre (ADF), and neutral detergent fibre (NDF) in situ rumen disappearance to assess kudzu quality. Four studies were conducted during different growing periods [2 repeated early season (ES); 2 repeated late season (LS)] to determine age variability effects. Kudzu was incubated in the rumen between 0.25 and 72 h to determine total rumen degradation (%) and the rate of disappearance (% h−1). Data were analysed as a randomized complete block design (RCBD) with repeated measures. Regression analysis was used to determine the degradation rate. Overall season effects (across incubation times) for dry matter disappearance (DMD) were significantly different (P < .0001). There were also significant season by incubation time interactions for DMD. Rumen degradation rate was determined to be 2.41% h−1 across both seasons, with no significant seasonal difference (P = .3396). Results from this study are indicative that ES kudzu is more degradable than LS. Although there are significant seasonal effects on kudzu DMD during certain incubation times, DMD overall for ES and LS is still indicative of high rumen degradability.
KEYWORDS:
Introduction
Kudzu (Pueraria montana var. lobata), an invasive semi-woody perennial vine weed species native to eastern Asia, was introduced to the United States in the late 1800s as a land restoration crop for soil erosion control. However, due to its perenniality and pervasiveness, kudzu was later classified by the USDA as an invasive weed species in the 1970s (Everest et al. Citation1999). Historically recognized as an exceptional forage legume in the tropical Caribbean island region (Vicente-Chandler et al. Citation1953), kudzu was well-known for its adaptability and summer persistency. However, as a common trait for vine-type plants, the easy-to-propagate feature and pronounced vigour of kudzu also make it one of the most invasive plant species that could easily encroach into existing pasture ecosystems, preventing other predominate forage species from receiving adequate solar radiation and soil nutrients to survive. Most importantly, kudzu has indicated exceptional tolerance to soil pH change (ranging from 4.0 to 8.0; Loustalot and Telford Citation1948) and remarkable cold tolerance (Coiner et al. Citation2018), making it even more advantageous in the climate-transitioning Southeastern U.S., where ample rainfall increased soil acidity rapidly and rising incidence of late-spring frost constantly jeopardized forage production in this region. Animal performance data on the use of kudzu as a major feed source is extremely limiting. The reasons are multifold, including anti-quality concerns such as bloat caused by highly soluble protein content, and effects of certain isoflavones identified in kudzu with complex medicinal properties are largely uncertain for livestock (Reppert et al. Citation2008). Furthermore, grazers such as beef cattle typically ignore vine plants unless the availability of other predominant forages is highly limited. However, browsers (e.g. goats and deer) are herbivores that are naturally adapted to feeding on leaf and twig growth of shrubs, vines, and trees. Therefore, intensive browsing by goats could serve as a viable option to biologically control kudzu, allowing other forages to thrive, meanwhile, providing high-quality feed for supporting desirable animal performance (Hart Citation2001; Bohnert and Stephenson Citation2016).
Forage nutritive value of kudzu (17.5% CP, 48.1% NDF, and 38.2% ADF) is comparable to other high-quality legumes such as alfalfa (18.7% CP, 46.0% NDF, and 36.9% ADF) at similar growth stages (Corley et al. Citation1997; NRC Citation2012). However, there are few historic examples of kudzu being used as a supplement in livestock and poultry diets. Polk and Gieger (Citation1945) demonstrated that an inclusion rate of 9% kudzu meal could be supplemented in place of alfalfa meal in chick diets. Emery and Norman (Citation1947) observed that Guernsey cows found tropical kudzu (Pueraria phaseoloides) palatable, and was grazed and browsed successfully by oxen, dairy cows, and goats. However, research-based data on the degradability of kudzu in ruminants is limited. Corley et al. (Citation1997) performed a rumen in situ study on kudzu leaf, stem, and tuber over a 24-h incubation time and found that kudzu canopy biomass provides excellent nutritive value, a slower digestion rate, but a greater digestible fraction. However, only one-time sampling was conducted in that study, thus, no information was available on effects caused by growth stage differences. This repeated rumen in situ study aimed at investigating total rumen degradability of dry matter (DM), crude protein (CP), acid detergent fibre (ADF), and neutral detergent fibre (NDF) of kudzu above-ground biomass (detached leaves from petioles) harvested during ES and LS in middle Tennessee. The general reduction in forage nutritive value and voluntary intake with progressing senescence is well observed. However, quantifying the extent of contributory effects caused by the growing season of such an invasive forage species in the middle Tennessee region could provide important scientific data to both forage and livestock producers on how to better control, utilize, and manage kudzu in their production systems.
Materials and methods
Kudzu used in this research was collected from nine naturally growing sites across seven counties in the middle Tennessee area (Bedford, Cannon, Coffee, Dekalb, Lawrence, Maury, and Rutherford). Early season kudzu leaf samples were collected in June and LS in July of 2018, respectively. During each sampling, a 30.5-cm2 PVC square quadrat was randomly tossed into clusters of kudzu. Above-ground biomass within the quadrat was collected by clipping an estimated 15 cm depth of kudzu vine at a 2-cm cutting height. After collection, all forage biomass were stored in paper bags and dried at 60°C for 48 h until reaching constant dry weight. Once dried, kudzu leaves were ground and passed through a 2-mm screen, then compiled into composite samples across all counties for each growing season (ES or LS). Forage nutritive value was determined using near-infrared reflectance spectroscopy (NIR) analysis (Dairy One, Ithaca, NY) and results were summarized in .
Table 1. Near-infrared reflectance spectroscopy (NIR) chemical composition of early (ES) and late season (LS) kudzu leaves incubated in experimental fistulated bovine during in situ rumen degradation study periods (DM basis).
A modified in situ degradation protocol was followed according to Vanzant et al. (Citation1998). For each composite ground sample, 24 5-g subsamples were weighed and individually placed in an Ankom® nylon forage bag [(10 × 20 cm; 50 ± 10 micron porosity)]. Each sample bag was prepared in triplicate. Each nylon bag was heat sealed three times before ruminal fluid incubation. A ruminally fistulated animal (Bos taurus steer) was used for each in situ study. The steer was provided ad libitum mixed grass hay (), soybean meal (2.27 kg d−1; 46.50% CP, 4.00% CF, 0.50% crude fat), and free-choice vitamin and mineral premix () 7 d prior to initiation of sample incubation and throughout entirety of in situ incubation periods. All animal handling procedures were approved by the MTSU Institutional Animal Care and Use Committee (IACUC) and conformed to accepted practices (FASS Citation1999). This in situ procedure was repeated two times for both ES and LS forage samples. Twenty-one nylon bags were suspended in the rumen at time intervals of 0.25, 1, 12, 24, 36, 48, and 72 h. Zero hour bags were processed after each study’s completion. Samples were subsequently placed in a large mesh bag (28 × 74 cm) beginning with the 72 h incubation time. After the 0.25 h incubation time was completed, all samples were removed and submerged in water (26°C) to halt fermentation activity and were then hand-rinsed, dried, and analysed for DMD and disappearance of CP, ADF, and NDF. A modified Vanzant et al. (Citation1998) hand-rinsing procedure was used for residue processing. Residue samples were hand-wash agitated in 39°C water for 1 min and spun dry for 2 min. This protocol was repeated 5 times for each sample. After rinsing, residue samples were dried at 60°C for 48 h and were then air-equilibrated for 36 h. Samples were weighed for dry matter disappearance (DMD) determination and were then removed from nylon bags and analysed for CP, ADF, and NDF (Dairy One, Ithaca, NY) using AOAC and Van Soest procedures (Goering and Van Soest Citation1970; AOAC Citation2016). The disappearance proportions at all measuring intervals were later used to determine average CP, ADF, and NDF disappearance rate.
Table 2. Near-infrared reflectance spectroscopy (NIR) chemical composition of early (ES) and late season (LS) mixed grass hay fed to experimental fistulated bovine during in situ rumen degradation study periods (DM basis).
Table 3. Composition of free-choice vitamin and mineral supplement provided to experimental fistulated bovine during in situ rumen degradation study periods.
Sampling season was considered as main treatment effect and repeated sampling time was considered as blocking effect to account for temporal correlation. Ruminal incubation time interval was considered as repeated measurement effect. Thus, the whole dataset was analysed as a randomized complete block design with repeated measures using the PROC mixed procedure of SAS (SAS Institute Citation2017) to determine treatment (season), incubation time, and treatment (season) by incubation time interactions. Regression analysis was conducted to determine in situ rumen degradation rate (% h−1) using data obtained between 0 and 24 h incubation periods. Data were considered significant at P < .05 level. Additionally, a standard in situ digestion experiment model was constructed according to an established model for kinetics of digestion originally proposed by Mertens and Loften (Citation1980), described in the following equation:(1)
(1) where R represents the residue (% DM), D
0 and U represent the digestible and indigestible fractions (% DM), respectively, kd
is the rate of digestion (% DM h−1), L is the lag time (h) that marks the time interval from the beginning of incubation time to active disappearance phase, and t is incubation time period (h). Nonlinear, least-square based parameter estimation routine using MATLAB Programming Language was implemented (The Math Works Inc. 2017, Natick, MA, U.S.A.).
Results and discussion
Forage nutritive value analysis of ES and LS kudzu biomass indicates that it is an excellent forage legume species for ruminant animals without considering potential anti-quality factors (). The obtained nutritive index values in this research far exceed those from other common legume hay, which typically offer 18% or more CP and 64% or more total digestible nutrients (TDN) (Rivera and Parish Citation2010). In this study, the ES and LS kudzu indicated a CP concentration of 30.5% and 26.7% and TDN of 76% and 59%, respectively (). However, Corley et al. (Citation1997) found kudzu biomass with an average CP concentration of 17.5%. This difference could be caused by the fact that this research focused on initial and mid-to-late growing season of kudzu leaves, whereas Corley et al. (Citation1997) mainly sampled those well-senesced tissues at the end of the growing season. Glass and Al-Hamdani (Citation2016), however, reported similar nutritive values of kudzu leaves to our findings (24.46% CP, 44.00% NDF, 28.67% ADF, and 55.99% TDN).
Overall, ES and LS kudzu incubated in the rumen from 0 to 72 h demonstrated seasonal effects for DM, NDF, ADF, and CP at various incubation intervals. Both incubation time and its two-way interaction with sampling season were significant (P < .001) for all indices (DMD, ADF, NDF, and CP). However, these two-way interactive effects were mainly caused by magnitude differences in response (–). Early season samples generally provided greater degradation rates at an early incubation period compared with LS, but those advantages diminished over time. Slope analysis based on the first 24 h incubation time indicated significant seasonal effects in CP (P < .01), ADF (P < .05), and NDF (P < .01) degradation but not in DMD (P = .25).
Figure 1. Total in situ rumen dry matter disappearance (DMD) of early season (ES) and late season (LS) kudzu. *Indicates significant seasonal effect at a given incubation time (P < .05).
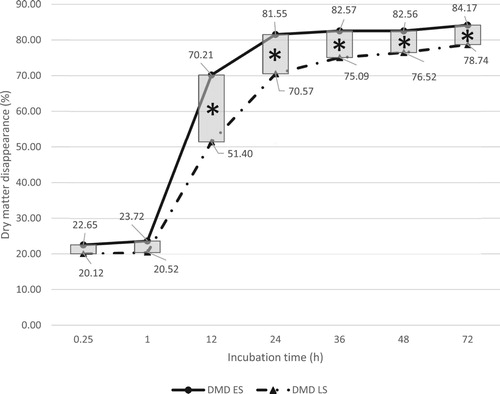
Figure 2. Total in situ rumen neutral detergent fibre (NDF) disappearance of early season (ES) and late season (LS) kudzu. *Indicates significant seasonal effect at a given incubation time (P < .05).
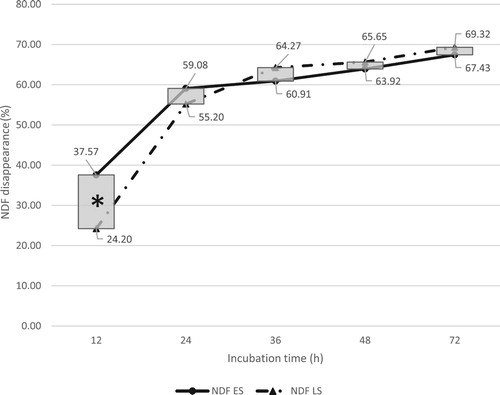
Figure 3. Total in situ rumen acid detergent fibre (ADF) disappearance of early season (ES) and late season (LS) kudzu. *Indicates significant seasonal effect at a given incubation time (P < .05).
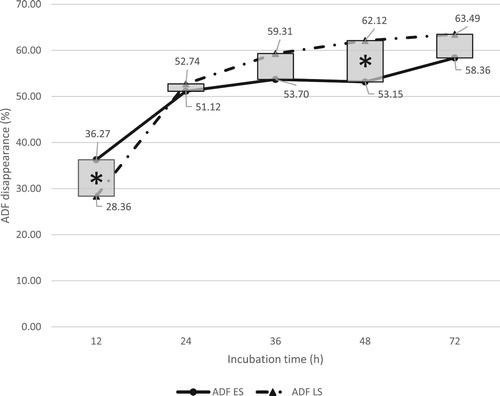
Figure 4. Total in situ rumen crude protein (CP) disappearance of early season (ES) and late season (LS) kudzu. *Indicates significant seasonal effect at a given incubation time (P < .05).
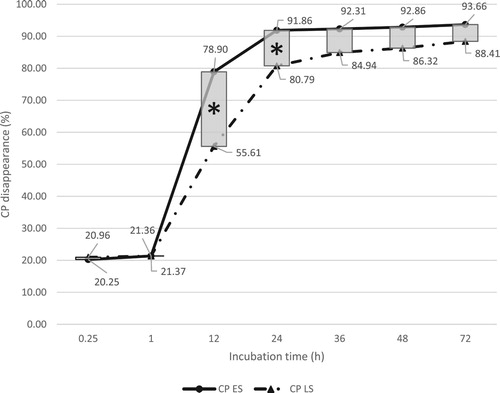
As indicated in , total rumen DM degradability of ES reaches a relatively stable phase starting from 24 h, and LS from 36 h of incubation. Slope of linear regression line of DMD within the first 1–24 h period indicated an average DM rumen degradation rate of 2.41% h−1 across both harvesting seasons, with no significant differences between the two slopes. In situ total DMD was significantly different between ES and LS (P < .0001) and among all incubation times (P < .0001). Significant seasonal effects on DMD were found at 12, 24, 36, 48, and 72 h (P < .001) of incubation (). In situ DMD for ES and LS samples were indicated in . Both ES and LS kudzu samples were highly rumen degradable, but there were seasonal differences in degradability. Average rumen degradability of ES kudzu was 9.75% higher than LS between 12 and 72 h. With ES kudzu being younger, containing less cell wall content (e.g. NDF), and insoluble lignin, it had an overall higher degradability than LS (). Less NDF in ES kudzu also indicated that it contained a higher concentration of neutral detergent solubles (containing primarily sugars, starches, and proteins) than LS, resulting in a higher degradability. Corley et al. (Citation1997) found that total in situ DMD for leaf and stem (1:1, wt:wt) was 78% at 24 h. In the present study, across both ES and LS; DMD for kudzu at 24 h of incubation was analysed to be 78.8%, which is comparable to the results reported by Corley et al. (Citation1997).
Table 4. Kudzu age by in situ incubation time interaction effects for DM, NDF, ADF, and CP disappearance.
shows ES and LS kudzu in situ rumen disappearance of NDF. Overall, across all incubation times, there was a significant difference between ES (41.27%) and LS (39.81%) kudzu NDF disappearance (P < .05), with ES being more degradable than LS kudzu. In situ disappearance of NDF for ES and LS kudzu at 12 h was 37.57% and 24.20%, respectively. A significant seasonal effect at 12 h (P < .001) was observed (; ). This could be indicative of ES kudzu having an increased early rumen degradability. After 12 h of incubation, there were no seasonal effects. This is likely due to low lignification of both ES and LS kudzu, thereby affecting NDF disappearance (). Additionally, the linear regression-based slope analysis indicated that ES and LS NDF degradation rates were different from each other and averaged at 2.59 and 2.27% h−1, respectively; which agreed with the ANOVA-based analysis results.
Early and late season kudzu in situ rumen disappearance of ADF is shown in . Overall, no significant differences were found between ES (36.09%) and LS (38.39%) kudzu ADF disappearance rates (P = .06). However, there were significant seasonal effects at 12 and 48 h (P < .05) (). In situ disappearance of ADF for ES and LS kudzu at 12 h was 36.72% and 28.36% and at 48 h 53.15% and 62.12%, respectively. In this research, kudzu age had minimal effects on ADF degradability. Likewise, the slope analysis also indicated a slight but significant difference between ES (2.30% h−1) and LS (2.23% h−1) ADF degradation rates.
Minimal seasonal effects were observed between ES and LS kudzu CP disappearance, as indicated in . Overall, across all incubation times, there was no significant difference between ES (70.17%) and LS (62.63%) kudzu CP disappearance (P = .09). However, there were significant seasonal effects at 12 and 24 h (P < .05) (). Early and late season kudzu in situ rumen degradation of CP at 12 h was 78.90 and 55.61% and at 24 h 91.86 and 80.79%, respectively. Although not significant, there was a tendency toward higher CP disappearance (P = .09) in the ES growth of kudzu. This was likely due to ES kudzu containing 28.9% available protein, whereas LS contained 24.9% (). Slope analysis also indicated that ES CP degradation rate (3.39% h−1) was significantly greater than LS values (2.60% h−1). Corley et al. (Citation1997) found kudzu leaves contained 17.5% CP. In the current study, ES and LS kudzu leaves had 30.5% and 26.7% CP, respectively. These differences in composition are likely due to kudzu variety or soil nitrogen levels. Corley et al. (Citation1997) collected kudzu between June and October, whereas in this research kudzu samples were collected between June and July. As indicated by Corley et al. (Citation1997), the leaf part of kudzu contains the greatest amount of CP (compared to stems and roots) and will meet the requirements of most ruminant animals (NRC Citation2012).
Finally, traditional nonlinear model-fitting techniques were used to evaluate the seasonal effects on DM, ADF, and CP disappearance as influenced by incubation time (). Overall, results from this conventional model-fitting approach agreed well with our ANOVA-based analysis. For DMD, ES digestible DM was greater than the LS. As expected, ES indigestible DM was lower than the LS, but the overall rate of digestion of DM were similar. The longer lag time associated with LS samples were caused by an increased time for hydrating more fibrous cell wall materials and prolonged time required for microbial attachment for the degradation on more senesced forage tissues. Similar results were also found in CP. Again, ES tissues indicated greater CP concentration, less lag time, and lower indigestible fraction compared with LS. No differences were detected in model parameters of NDF except for the indigestible DM. Again, older forage tissues tend to have more indigestible fibre such as lignin. Estimated parameter values of kudzu were similar to average values obtained from various forage species as reported by Mertens and Loften (Citation1980), with a slightly lower rate of digestion on fibre content. Additionally, estimated D 0 and kd values for DM were greater than Tifton 85 bermudagrass as reported by Smith et al. (Citation2017). Meanwhile, both D 0 and U values of NDF or ADF were smaller than bermudagrass and lag time was also much shorter. These findings indicate that kudzu, as a high-quality legume, should provide a much greater digestible biomass/fibre content with higher rates of digestion caused by a lower concentration of indigestible fibre than a warm-season grass.
Table 5. Estimated nonlinear model parameter values of early (ES) and late season (LS) kudzu DM, NDF, ADF, and CP disappearance from a traditional fitting technique as influenced by in situ rumen fluid incubation time.
Conclusion
Early and late season kudzu nutritive value and rumen degradability indicated that it is a nutrient-rich, highly degradable forage species for ruminant animals. Significant age variability effects influenced kudzu DMD, but the rate of kudzu rumen degradability was similar across seasons. Early season kudzu is more rumen degradable than LS, indicating ES is the most beneficial growing season for animal production. Using ruminant animals to control kudzu can reap the benefits of both invasive plant species control and providing a high-quality feedstuff to produce ruminant animals. Browsing early growth kudzu would yield a higher degradability in the rumen, potentially leading to greater animal performance. Likewise, defoliating kudzu during the early growing season should cause greater stress to forage growth (due to the plant being young and having limited carbohydrate stores). Further research is needed to assess primary total nonstructural carbohydrate storage organs and the dynamics of nutrient allocation of kudzu during kudzu’s growing season. This information could assist producers in managing kudzu population to use it as a companion plant for enhancing soil health and effectively controlling its growth and production through livestock browsing. Although, well known for its medicinal properties for treating several human diseases (Reppert et al. Citation2008), the major anti-quality/nutritional factors inherent to kudzu that could compromise animal performance are still largely unknown; thus, should be an important future research direction. Overall, this research indicates that kudzu is a highly degradable and nutritious forage legume for ruminants, and, when available and accessible, should be considered as a supplement to a ruminant’s diet.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- AOAC . 2016. Official methods of analysis. 20th ed. Arlington (VA ): AOAC International.
- Bohnert DW , Stephenson MB. 2016. Supplementation and sustainable grazing systems. J Anim Sci. 94:15–25. doi:10.2527/jas.2016-0520.
- Coiner HA , Hayhoe K , Ziska LH , Van Dorn J , Sage RF. 2018. Tolerance of subzero winter cold in kudzu (Pueraria montana var. lobata). Oecologia. 187:839–849. doi: 10.1007/s00442-018-4157-8
- Corley RN , Woldeghebriel A , Murphy MR. 1997. Evaluation of the nutritive value of kudzu (Pueraria lobata) as a feed for ruminants. Anim Feed Sci Tech. 68:183–188. doi:10.1016/S0377-8401(97)00038-2.
- Emery AT , Norman CF. 1947. Tropical kudzu in Puerto Rico. Pasturing. Federal experiment station in Puerto Rico. Circular. 27:23–24.
- Everest JW , Miller JH , Ball DM , Patterson M. 1999. Kudzu in Alabama: history, uses, and control. Alabama Cooperative Extension System. ANR-65.
- Federation of Animal Science Societies . 1999. Guide for the care and use of agricultural animal in agricultural research and teaching. General guidelines for animal husbandry. p. 7–19.
- Glass D , Al-Hamdani S. 2016. Kudzu forage quality evaluation as an animal feed source. Amer J Plant Sci. 7:702–707. doi:10.4236/ajps.2016.74063.
- Goering HK , Van Soest PJ. 1970. Forage fiber analyses. ARS/USDA handbook no. 379. Superintendent of Documents, US Government Printing Office, Washington, DC. P15.
- Hart SP. 2001. Recent perspectives in using goats for vegetation management in the USA. J Dairy Sci. 84:E170–E176. doi:10.3168/jds.S0022-0302(01)70212-3.
- Loustalot AJ , Telford EA. 1948. Physiological experiment with tropical kudzu. J Amer Soc Agron. 40(6):503–511. doi: 10.2134/agronj1948.00021962004000060003x
- Mertens DR , Loften JR. 1980. The effect of starch on forage fiber digestion kinetics in vitro. J Dairy Sci. 63:1437–1446. doi:10.3168/jds.S0022-0302(80)83101-8.
- NRC . 2012. Nutrient requirements of beef cattle. 11th rev. ed. Washington (DC ): Natl. Acad. Press.
- Polk HD , Gieger M. 1945. Kudzu in the ration of growing chicks. Miss Agr Expt Sta Bull. 414:1–14.
- Reppert A , Yousef GG , Rogers RB , Lila MA. 2008. Isolation of radiolabeled isoflavones from kudzu (Pueraria lobata) root cultures. J Agric Food Chem. 56:7860–7865. doi: 10.1021/jf801413z
- Rivera D , Parish J. 2010. Interpreting forage and feed analysis reports. Mississippi State University Extension Service. Publication 2620.
- SAS Institute . 2017. SAS/STAT® user’s guide version 14.3. Cary (NC ): SAS Institute.
- Smith WB , Foster JL , McCuistion KC , Tedeschi LO , Rouquette FM Jr . 2017. In situ degradation pattern of ‘Tifton 85’ bermudagrass with dried distillers’ grains supplementation. Crop Sci. 57:1773–1783. doi: 10.2135/cropsci2016.12.0981
- Vanzant ES , Cochran RC , Titgemeyer EC. 1998. Standardization of in situ techniques for ruminant feedstuff evaluation. J Anim Sci. 76:2717–2729. doi:10.2527/1998.76102717x.
- Vicente-Chandler J , Caro-Costas R , Figarella J. 1953. The effect of two heights of cutting and three fertility levels of the yield, protein content and species composition of a tropical kudzu and molassesgrass pasture. Agron J. 45:397–400. doi: 10.2134/agronj1953.00021962004500090001x