
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Oregano essential oil (OEO), one kind of Labiata has been known for a long time as a popular remedy. The essential oil of oregano has anti-bacterial, anti-oxidant, anti-fungal and nematicidal properties mainly due to the basic constituents. Once the shell’s microbial barriers have been breached, Gram-negative bacteria are more capable of withstanding the antimicrobials present in the albumen. Eggs were divided into, control group (GI), LPS group (GII) and LPS plus OEO 0.5% group (GIII), as natural disinfectants. Statistical data were analysed, histopathological, RT-PCR and in situ hybridization studies were performed to explain the OEO modulatory role. Lipopolysaccharide (LPS) group represented the highest (P < 0.05) increase of embryonic mortality percentage, greatest significance (P < 0.01) egg weight loss, a decrease of hatchability and high teratogenic effect compared with those for other groups in all types of experiments. Chicks produced from egg subjected to oregano 0.5% and LPS significantly (P < 0.05) recorded large body weight gain, enhancement of hatchability and produces no teratogenic effects in the developing chicken. OEO modulates the effect of LPS in histopathological, RT-PCR and in situ hybridization studies throughout modulation of the Daam2 and Tbx2 gene expression towards the normal one. Using Oregano oil as a natural disinfectant in this study modulate significantly the effect of LPS oxidative effect on the chick embryo even after hatching, so, we conclude that OEO is an effective anti-oxidant that recommended to be used in hatchery for good quality chicks, chick growth and finally improvement of laying production performance.
1. Introduction
Natural active compounds extracted from plants are more acceptable and less harmful than synthetic chemical compounds and represent principle agents to control diseases. The secondary metabolites of the natural medicinal plants have shown to be used to control infectious organisms and to control the problems associated with synthetic chemicals, plant essential oils, having antimicrobial effect (Mousa et al. Citation2016; Sevim et al. Citation2020). In the present work, we attempt to detect the application of natural essential oils to control microbial activity on eggshell and to determine its effects on different parameters as hatchability of fertile egg, embryonic growth and development of chicks after hatch.
Once the shell’s microbial barriers are breached, Gram-negative bacteria are more capable of withstanding the antimicrobials present in the albumen. Salmonella Enteritidis and Pseudomonas fluorescens were both able to survive at different rates in various parts of the egg. While Salmonella Enteritidis survived best on the exterior surface of the shell, Pseudomonas fluorescens was better able to transverse the shell membranes and infect the contents of the egg (Jones et al. Citation2004).
Oregano, one kind of Labiata known for a long time as a popular remedy. The essential oil of oregano has anti-bacterial, anti-oxidant, anti-fungal, cytotoxic and nematicidal (Meneses et al. Citation2009; Hang et al. Citation2016; Leyva et al. Citation2017; Vidács et al. Citation2017; Baylan et al. Citation2018) properties which had been recorded by many groups of researchers as a result of several contents of essential oils such as carvacrol, terpenes, thymole, … etc. Natural products in plant were recommended as modulatory components for human being’s health. Where the secondary metabolites of the naturally occurring active plant compound are found to control and verse the oxidative and destructive effect of the synthetic chemical compounds, so understanding the role of natural plant component is very important chemicals (Vidács et al. Citation2017).
The environmental factors that are most critical to the normal development of the embryo are those that occur before and during incubation and hatching processes. These factors include using sanitizers and disinfectants (Meijerhof Citation2000). Egg infection can be risk factors for bone integrity leading to bone weakness (Rath et al. Citation2000).
Embryonic morphogenesis usually considered to result from cell movements and changes in cell behaviour resulting in embryonic morphogenesis, integrated with environmental factors include using sanitizers and disinfectants that are most critical to the normal development of the embryo before and during incubation and hatching processes. Embryos localized zones of high cell proliferation in the epiblast layer are thought to be important in determining several phases of early development.
Lipopolysaccharide (LPS) is an endotoxin found in the cell wall of gram-negative bacteria. These endotoxins are released by bacterial cell wall destruction. LPS acts as a powerful inflammatory-inducing agent. LPS consists of three main specific parts O-antigen, central core and lipid A (Alessandra and Paola Citation2014). LPS structure differs from one species to other and this structure is used to differentiate between different species of Bactria. It causes direct injury to many organs of the body. It causes elevation of lipid peroxidation as an index of oxidative stress (Giriwono et al. Citation2011). So LPS are used as an excellent model for inflammation studies (Tonelli et al. Citation2008). It is a potent activator of innate immune response (Miyake Citation2006). Most of the inflammation responses are due to LPS mediated throughout the Toll-like receptor (TLR) 4 that leads to differential expression of many pro-inflammatory genes such as IL-1β and TNF … etc. (Wang et al. Citation2011). LPS induce pro-inflammatory cytokines in different cell lines such as RAW264.7 (Cheng et al. Citation2018). When this cell line is treated with oregano oils it showed anti-inflammatory effect by decreasing nitrogen oxide (NO) (Marrelli et al. Citation2018). Treating with different essential oils causes an improvement in growth performance and reduction of cholesterol in the blood (Altop et al. Citation2018). LPS effect reduced significantly in broilers when oregano oil was supplemented in diet (Cetin et al. Citation2016). Oregano oil has an immunomodulatory role against LPS-induced cell and tissue damage. Oregano oil has many effective compounds indicating their different biological properties. In our study, we aim to spotlight the protective role of oregano oil against the destructive effect of LPS on chick embryo before and after hatching throughout the investigation of egg weight change, chick body weight, mortality rate, histological studies of chick embryo as a whole, expression of Daam2 and Tbx2.
2. Materials and methods
2.1. Animals
The present experiment studies were conducted at the Biological Sciences Department, Faculty of Science, King Faisal University (KFU). Fresh eggs were collected from KFU veterinary farm. Faecal contaminated eggshells and eggshells with visible cracks discarded.
Females with males from chicken strain were housed in floor pens during the laying stage (32–40 wks) of age. Layer diets contain 16.5% crude protein and 2750 ME kcal/kg diet. A total of 360 hatched eggs from the chicken strain was used in this experiment. Eggs were divided into three groups, representing the LPS injected group, disinfectants used Oregano 0.5%, as disinfectants from natural sources for combination with LPS group and control group (). Each group for studying the hatching parameter contains 40 hatching eggs and replicated three times for each treatment. The active ingredient of Origanum vulgare L (Oregano) β-pinene (0.5%) – thymol (0.86%), terpinene and p-cymene – Carvacrol (57.01%) has an effect on killing microorganism, anti-oxidant, anti-bacterial, anti-fungal. The basis of drug dose selection in our study was our own previous study (Zeweil et al. Citation2015). The chicks were fed ad libitum. All houses were provided with a number of nests sufficient to allow one nest for every seven birds. Fresh eggs were collected manually four times daily from the nests. Faecal contaminated eggshells and eggshells with visible cracks were discarded. Eggs were numbered consecutively and weighed before setting in an incubator, which operated at 37.5Co and 55% relative humidity. All eggs were weighed individually again during incubation on the 5th, 10th, 15th, 18th days and at first signs of pipping in order to obtain egg weight loss percentages for each incubation.
Table 1. Experimental design.
Chicks for either males or females were reared together, and sex was determined at the end of the 6 weeks growing period. Hatching failed eggs were broken and examined under a microscope every day. Embryonic mortality rate was recorded as a percentage of fertile eggs’ total number in each group. Data collected in three periods as infertile, early dead embryos (1–7) day, mid dead embryos (7–14) day, late dead embryos (14-pipping), embryos that died after external pipping and for hatched chicks.
For histological preparation and studying, embryos were rinsed in saline water and fixed in Bouin,s fluid for 24 h and processed for routine Haematoxylin and Eosin as described in the method of Gabe (Citation1976).
2.2. Statistical analyses
The statistical analysis was conducted using SAS program (SAS, Institute Citation1998) software, the following model was used:
where Yij = observation record, M = the overall mean, Li = is the effect of disinfection, i = 1–14 and eij = is the random error.
2.3. RT-PCR study
In this study, we used pre-hatching (18 days incubation) and post-hatching (2 weeks old) chickens. Eggs before hatching and chicks after hatching were maintained at the college of Science facilities in accordance with the guidelines for animal experiments at King Faisal University and the protocol approved by the experimental animal committee. Chicken (pre-and post-hatching) were grouped into three main groups: control group receive only phosphate buffer saline (Pbs) and alcohol, stimulated group injected with lipopolysaccharides (LPS) 10 µg/ml (rough strain) from Salmonella typhimurium SL1181 (RE mutant) (Sigma, U.S.A.), and treated group injected with LPS and oregano oil 0.5%. Tissues and organs subjected to our experiment (brain, liver, kidney, spleen, thymus, heart, lung and bursa of Fabricius) were isolated from samples. Total RNA extracted from the designated tissue using isogene (life science) from which mRNA purified. One μg of mRNA was then used for cDNA synthesis for each sample by oligo-dt12-28 primer (GIBCO-Invitrogen, USA). The transcripted cDNA was purified using phenol–chloroform extraction method. This cDNA was stored at −20°C until a further investigation had been done. Daam2 and Tbx2 genes expression was investigated in both pre-hatched and post-hatched ones. Chicken cDNA samples used to analyse target genes expression analysis were prepared from the selected tissues by RT-PCR using the primers 5′-TAGGGGCTGTCTGCTTGG-3′, 5′ -ATAGGGATAGGCATCTGT-3′ and 5′ -GCCGGGGCGGCCGAGGCC-3′, 5′ -CCGAGATGTTGTTGGTGA-3′ as forward and reverse primers for Daam2 and Tbx2, respectively, and chicken tissues cDNA as a template.
2.4. In situ hybridization
In this section, chicken tissues (spleen and bursa of Fabricius) were subjected to frozen section preparations. In situ hybridization was performed using digoxygenin-labeled (Roche Applied Science) probes. Sense and antisense probes were prepared from chicken cDNAs obtained from prepared samples according to the manufacturer manual with the following modifications. Linearizing the transcript (both sense and antisense) with T3 and T7/sp6 RNA polymerase The +1 G of the RNA polymerase promoter sequence in the DNA template is the first base incorporated into the transcription product. To make sense RNA, the 5' end of the coding strand must be adjacent to or just downstream of, the +1 G of the promoter. For antisense RNA to be transcribed the 5' end of the noncoding strand must be adjacent to the +1 G. If the insert is in a vector, the vector should be linearized downstream from the promoter and the inserted sequence to be transcribed . The hybridization protocol was carried out according to Olivier and Walter (Citation1998) with a few modifications as following. Slides were immersed in 4% paraformaldehyde in 0.1% DEPC-treated PBS for 30 min for fixation. Followed by 0.1% active DEPC-PBS 15 min × 2 times for RNase inactivation, slides then dipped in DEPC-treated 5× SSC (0.75 M NaCl, 0.075 M Na-citrate) for 15 min. Followed by 58°C for 2 h in 50% formamide/5× SSC buffer, 40 μg/ml salmon sperm DNA as a prehybriidization step, hybridization was done for 28 h at 58°C, with 500 ng/ml probe of DIG-labelled chicken Daam1 and Tbx2 fragment, in 50% formamide/5× SSC buffer, 40 μg/ml salmon sperm DNA. The incubation was carried out in 50% formamide /5× SSC buffer saturated chamber. Washed by 2× SSC buffer at room temperature for 1 h in 2× SSC at 65°C., 1 h in 0.1 SSC and finally for 10 min in buffer 1 (Tris 100 mM, NaCl 150 mM, pH 7.5) as an equilibrium medium. Then lifted overnight with anti-DIG antibody, Pod-coupled, diluted 1:300 in buffer 2 (buffer 1 with 0.5% Boehringer Blocking reagent) at 4°C. Washed two times for 20 min with buffer 1, then 5 min in buffer 3 (Tris 100 mM, NaCl 150 mM, Mgcl2 5o mM, pH 9.5), stained with substrate kit for peroxidase (vector® NovaRED, Funakoshi, Japan) for 30 min, under tap water for 10 min and finally stained with Methylene green for 10 min, dried and mounted.
3. Results
3.1. Embryonic weight
Results of average absolute and relative embryonic weights at different incubation ages as affected by different disinfectants and LPS infection are summarized in (). It appears from () that eggs subjected to disinfectants from natural sources had the heaviest (P < 0.05) absolute embryonic weight daily from 2 to 17 days compared to those infected to LPS and control one. Absolute embryonic weights produced from eggs treated with OEO recorded heaviest (P < 0.05) than those of LPS treated group except for18 day incubation control eggs. LPS treated group recorded the lowest absolute embryonic weight among all groups. The best absolute embryonic weight is reported in the OEO-treated group as a whole during 2–18 d of incubation (). Relative embryonic weight results showed a parallel track of recording where OEO showed a significant (P < 0.05) modulation in chick relative embryonic weight when compared with LPS injected group. The most significant (P < 0.05) results were obtained on days 16, 17 and 18. It is supposed that the increase of embryonic development and consequently in embryonic weight as illustrated in –6 resulting from using disinfectant from natural sources compared to those LPS and control is due to that of these disinfectants did not adversely affect the cuticle and shell properties. This explanation highlights the main causes, which may influence the growth and weights of embryo at different incubation days with natural disinfectant. Moreover, it has been demonstrated that microbial infection is an important cause of embryonic death and the principal microbes are those environmental or faecal bacteria and fungi. Results of total embryonic mortality for LPS group were the worst one compared to other natural and control treatments ().
Table 2. Effect of hatching egg disinfectants from natural sources and LPS on daily absolute embryonic weights (X ± SE) Group I (control) Group II (LPS injected) Group III (LPS + OEO).
Table 3. Effect of hatching egg disinfectants from natural sources and LPS on daily relative embryonic weights (X ± SE). Group I (control) Group II (LPS injected) Group III (LPS + OEO).
Table 4. Effect of hatching egg disinfection on absolute and relative embryonic weight at different ages during incubation (X ± SE). Group I (control) Group II (LPS injected) Group III (LPS + OEO).
Table 5. Effect of hatching egg disinfection on absolute and relative embryonic weight at different ages during incubation (X ± SE). Group I (control) Group II (LPS injected) Group III (LPS + OEO).
Table 6. Effect of hatching egg disinfection on absolute and relative embryonic weight at different ages during incubation (X ± SE). Group I (control) Group II (LPS injected) Group III (LPS + OEO).
Table 7. Effect of hatching egg disinfection on absolute and relative embryonic weight at differentages during incubation (X ± SE). Group I (control) Group II (LPS injected) Group III (LPS + OEO).
Table 8. Effect of hatching egg disinfection on embryonic mortality during different stages of incubation (X ± SE). Group I (control) Group II (LPS injected) Group III (LPS + OEO).
The results of the present investigation, which reveal the increase of embryonic mortality and decrease of hatchability percentage in infected egg groups and the reverse in oregano and LPS, egg groups, which demonstrate the increase of hatchability with the decrease embryonic.
3.2. Hatchability
Hatchability percentage is a major estimator from the studied criterion for the evaluation of egg disinfectant and LPS infection. As it appears from , egg hatchability percentages in LPS group was lessened compared to those for other treated groups including control groups and other disinfected group. These findings support the results of the lowest hatchability percentage for the group of Oregano 0.5% treatment () with the greatest egg weight loss during the setting phase and whole incubation period.
Table 9. Effect of hatching disinfection on macroscopic fertility and hatchability percentages (X ± SE). Group I (control) Group II (LPS injected) Group III (LPS + OEO).
3.3. Effect of hatching egg disinfection on post-hatch growth
3.3.1. Body weight
Effects of hatching egg disinfection from different sources compared with control on body weight of post-hatch growing chicks are presented in . It could be noticed from this table that chicks from eggs subjected to disinfectants from the natural source were significantly (P < 0.05) heavier than those hatched from other infected eggs group throughout the different ages of the growing period.
Table 10. Effect of hatching egg disinfection on body weight of post-hatch growing chicks (X ± SE): Group I (control) Group II (LPS injected) Group III (LPS + OEO).
3.4. Histopathological studies
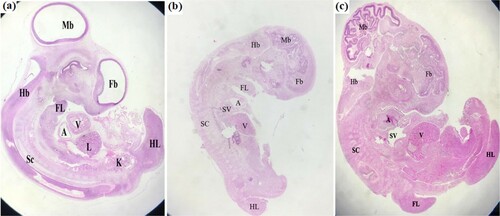
Control cases with alcohol revealed that the embryo was more developed, high-weighed and the morphology showed progress in development than that of 4-day where the tail buds upwardly curved, eye was developed and the mesencephalon (2optic lobes) was more lengthened and the leg buds slightly larger than wing buds. At the level of cervical flexure, we recorded the approximate right angle between the axis of medulla and the axis of the posterior trunk. Furthermore, the trunk flexure recorded a shifting to the lumbar region. So the rotation extends the chick embryo to the posterior part. The tail bud turned to the right at about an angle of 90° to the axis of the posterior trunk. Also, the heart was more developed ((a)). While the embryos infected with LPS showed low-weighed embryo, underdeveloped, the morphology was delayed lesser than 4-day where the tailbud was less curved, eye was malformed and mesencephalon (2 optic lobes) was more degenerated. The dorsal contour of the trunk was a straight line or slightly curved and the anterior and posterior contours were parallel at their bases sometimes extended to the tip of the tail. Malformed brain and heart ((b)). On the other hand, all natural disinfectant oregano treated cases restored to their normal morphology where the wing and leg buds distinctly longer than wide, the dorsal contour of the trunk was bent and eye pigmentation distinct., eye and mesencephalon (2 optic lobes)were developed, long limb buds were lengthened, otherwise show little change in shape ((c)).
Figure 1. Photomicrographs showing sagittal sections of chick embryo after 3 days of incubation (a) Control chick embryo showing normal morphology brain chambers fore, mid and hind brain FB, MB, HB, limb buds fore and hind FL HL heart chambers SV, A V tissue of liver L and kidney K and spinal cord SC (b) Embryo injected with LPS showing underdeveloped and malformed morphology (c) Embryo injected with LPS and dipping with Oregano oil showing normal morphology appearances.
Histologically, in the control case, the liver of group I showed normal structure ((a)) with hepatic parenchyma and hepatic cords and also focal areas of sinusoids aggregated nucleated WBCs. Photomicrographs of the liver of Group II injected with LPS showed an adverse degree of histopathological modifications in the liver as compared to the control groups. These modifications including breaking of hepatic cords, degeneration of hepatocytes some hepatocytes were characterized by cytoplasmic vacuolation, losing, shrinkage and swelling. Varying degrees of necrosis and apoptosis of hepatocytes were seen in addition to the formation of rosettes of regenerating hepatocytes (Rh) ((b)).
Figure 2. Transverse sections of liver (a–c) and heart (d–f) of 3-days chick embryos: (a) Normal structure of liver of control cases with normal hepatocyte cords Hc and sinusoids S (b) Liver of group 2 showing adverse degree of histopathological modifications: inflammatory infiltrations and rosette of hepatocytes (black arrows) (c) Liver of group 3 showing normal architecture in appearance with hepatocyte cord in line (black arrows)and single hepatocytes (white arrows) (d) Heart of group 1 control case with normal atrial chambers left and right (RA, LA)and thick walled ventricle (V) (e) Heart of group 2 injected with LPS showing malformed atrium an(A) and small and thin walled ventricle.
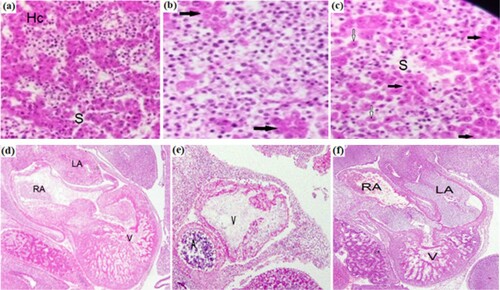
It was obvious that lymphoid cell infiltration was diffused in hepatic parenchyma. The inflammatory infiltrates may breach the interface between the portal tract and hepatic parenchyma. The inflammatory cells are mostly lymphocytes sometimes forming lymphoid follicles, plasma cell containing gamma globulins mostly igG type and macrophages containing iron granules may be present within the infiltrate.
In Group III [combination of LPS and Oregano 0.5%]. The hepatocyte parenchyma with more or less normal architecture ((c)) Linear rearrangement of hepatic cords, no obvious of necrotic and apoptotic hepatocytes in this liver section. There was tiny lymphoid infiltration. This may perhaps indicate the protective effect of herbal extract over LPS-induced pathological changes in hepatic tissue. It also showed normal hepatic parenchyma without focal changes in hepatocytes, mild degree of pathological alterations of degenerative nature in hepatocytes that signify the protective effect of herbal drug under study ((c)).
The heart was well developed in the control chick embryo ((d)). It had been differentiated into atria and ventricles with a complete interventricular septum. The ventricular walls showed a thick myocardial layer. When these heart structures were in the infected cases with LPS, it was found to be affected adversely, as thinning and occasional loss of myocardium and ventricular septal defect and hypomorphic atria ((e)). Treatment with oregano oil showed normal heart structure with normal development ((f)).
3.5. RT-PCR and in situ hybridization studies
shows the RT- PCR amplification of Daam2 and Tbx2 genes on the 18th day of incubation. RT-PCR amplification showed that the expression of Daam2 and Tbx2 is significantly increased in LPS-injected group if compared with the control one in all organs studied ((a,c)). Treatment with oregano oil showed a significant modulation of the LPS effects and the expression of this gene showed almost no change than that of control samples in most of organs under investigation ((a,c)). These results were confirmed by insitu hybridization study ((b,d)) of spleen and Bursa of Fabricus for both genes (Daam2 and Tbx2). Signal strength is very high in LPS injected group ((bii, bv, dii and dv)) if compared by the control one ((bi, biv, di and div)) in both organs. The oregano oil treated group showed a moderate signal ((biii, bvi, diii and dvi)) when compared by LPS treated group and near that of control.
Figure 3. Modulatory effect of OEO on the expression of both Daam2 and Tbx2 gens. (a) RT-PCR expression profile of Daam2 gene in different organs at 18th day incubated chick. (b) Expression rate status in spleen (i group I (control), ii Group II (LPS) and iii Group III (OEO)) and bursa (iv group I, v Group II and vi Group III). (C) RT-PCR expression profile of Tbx2 gene in different organs at 15th day post-hatcing chick. (d) Expression rate status in spleen (i group I, ii Group II and iii Group III) and bursa (iv group I, v Group II and vi Group III). Arrows showed the expressed mRNA in both tissue in different treatments.
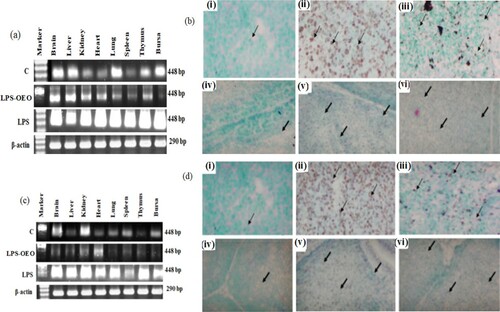
showed a protective effect of oregano oil against LPS of two weeks old chick spleen and bursa of Fabricus. Both investigated genes showed a high expression rate in LPS injected group in all organs under investigation ( (a,c)). We investigated Spleen and bursa for both genes by in situ hybridization. Results showed that LPS injection increase significantly the mRNA expression of both investigated genes in spleen and bursa ((b,d)). The oregano oil-treated group showed decreased significantly the expression rate ((a–d)) of both genes.
Figure 4. Modulatory effect of OEO on the expression of both Daam2 and Tbx2 gens. (a) RT-PCR expression profile of Daam2 gene in different organs at 15th day post-hatcing chick. (b) Expression rate status in spleen (i group I (control), ii Group II (LPS) and iii Group III (OEO)) and bursa iv group I, v Group II and vi Group III). (C) RT-PCR expression profile of Tbx2 gene in different organs at 15th day post-hatcing chick. (d) Expression rate status in spleen (i group I, ii Group II and iii Group III) and bursa iv group I, v Group II and vi Group III). Arrows showed the expressed mRNA in both tissue in different treatments.

4. Discussion
It is supposed that the increase of embryonic development and consequently in embryonic weight as illustrated in resulting from using disinfectant from natural sources compared to those LPS and control is due to that of these disinfectants did not adversely affect the cuticle and shell properties. These results are in accordance with those reported by Copur et al. (Citation2010) reported that oregano essential oil had no detrimental impact on the development embryo (Kusstatscher et al. Citation2017; Baylan et al. Citation2018). Otherwise, Sevim et al. (Citation2020), also reported the same result by using rosemary essential oil as a disinfectant.
It could indicate that LPS may have a toxic effect on embryo viability. Microbes on eggshells of newly laid eggs can multiply rapidly when exposed to appropriate ambient conditions, can penetrate the eggshell through pores, and can lead to a dramatic reduction in hatching success. This may lower the risk of infection and reduced hatching success because the likelihood of trans-shell infection is positively related to microbe densities on eggshells (Cook et al. Citation2005).
Results of the present investigation reveal an increase of embryonic mortality and decrease of hatchability percentage in injected egg group and the reverse in oregano and LPS egg groups which demonstrate the increase of hatchability with the decrease embryonic mortality are in accordance with those reported by Adaszyńska-Skwirzyńska and Szczerbińska (Citation2017) and Zhang et al. (Citation2019), who reported that LPS acts on proteins and also on nucleic acid. It is feasible that formaldehyde gas diffused into the egg at an early stage of embryonic development will alkylate the nitrogen atoms of purine and pyrimidine bases in DNA thus inhibiting their functions. This in turn, blocks embryonic development at an early stage.
In , we noticed that chicks from eggs subjected to disinfectants from the natural source were significantly (P < 0.05) heavier than those hatched from other infected eggs group throughout the different ages of the growing period. This result in accordance with Roofchaee et al. (Citation2011) Also, LPS group exhibited the lowest significant (P < 0.05) chick body weights compared to other experimented treated groups at 0, 4 and 8 weeks of growing ages. Moreover, egg disinfection with LPS control groups did not significantly affect the subsequent chick body weight at 12 weeks of age. The hatchability improvement due to the use of experimented disinfectants is the main goal of this experiment. Many factors affecting hatchability success are associated with the functional properties of the eggshell (Melo et al. Citation2019).
There is little literature on the effect of LPS on the development of chick embryos and its mechanism of action as a teratogenic agent. LPS showed a strong oxidative effect on the chick embryos. On the other hand, it was found that in the treatment with Oregano oil in combination with LPS, there were no specific malformations associated with Oregano oil treatment in the set of the experiment which were not seen in the controls. Defects in the neural and heart development, malformed axial skeleton, morphological defects and histological damage were reported during chick embryo development. that are inconsistent with studies of (Bokariya and Umarji Citation2012; Baylan et al. Citation2018; Zhang et al. Citation2019; Bekhet Citation2020) they stated that a study of infection of the developing chick embryos might yields interesting results as regards to pathogenicity of disease, because preliminary experiments showed that the embryo was extremely sensitive to infection and that definite and pronounced lesions were easily recognizable in the tissues of the embryo itself. Necrosis of affected liver tissues occurs within 72 h, and subsequent resorption of hepatocytes results in degeneration or loss of already formed structures, which may conclude as malformation which coincide with Cloney and Franz–Odendaal (Citation2015 and Zhang et al. (Citation2019). Similar results were also reported by (Gardinar Citation2006) which showed a reduction of 50% in cardiac defects with herbal oils during incubation. Furthermore, such embryonic heart defects were reported by (Zhang et al. Citation2019) caused by a variety of environmental toxins and maternal diseases.
We investigated the modulatory effect of oregano oil on the destructive effect of LPS on chick embryo and newly hatched chick from 0 d up to 15 days. Oregano oils are an essential oil from different species of the family Lamiaceae and have from 20 to 60 active components found in a mixture with different concentrations. Oregano oils have anti-inflammatory, anti-oxidant, anti-fungal and analgesic medicinal properties a long time ago (Fournomiti et al. Citation2015). Oregano oil has significant anti-bacterial properties including those gram-negative and gram-positive bacteria (De Oliveira et al. Citation2010). Furthermore, there is no potent information about the toxicity of oregano oils (Sturchio et al. Citation2016). We investigated the fluctuation of mRNA expression rate in different organs as shown in ((a,c) and (a,c)). We prepared mRNA from brain, liver, kidney, heart, lung, spleen, thymus and bursa respectively ((a,c) and (a,c)). For both Daam2 and Tbx2 genes as they are very important in the development of the chick embryo before and after hatching (Esteban et al. Citation1999; Li et al. Citation2011). Our results showed that there is a significant increase in Daam2 mRNA expression rate in LPS treated group in both RT-PCR ((a), and (a)) and in situ hybridization studies ((b) and (b)) in spleen and bursa. The expression rate of Daam2 mRNA in LPS treated group appears to be very high as shown by band densities in RT-PCR amplification and in the strength of the signal in in situ hybridization ((a, bii, biv) and (a, bii, biv)). In the OEO treated group mRNA expression was near to that control in RT-PCR ((a) and (a)) and in situ hybridization studies ((biii, bvi) and (biii, bvi)) indicating that the modulating effect of OEO on the LPS-induced oxidative stress. LPS cause an oxidative stress starts from weak inflammation to cell destruction (Cheng et al. Citation2018). It releases many pro-inflammatory cytokines such as IL-1 beta, IL-6 and TNF (Park et al. Citation2020) that cause oxidative stress for different types of cells. OEO modulate bacterial, viral and fungal harmful effect in accordance with our results (PuškárováCitation1 et al. Citation2017).
With regard to Tbx2 mRNA expression analysis, we found that the expression rate in the same line as that of Daam2. The LPS injection leads to the increasing of Tbx2 mRNA in RT-PCR amplification ((c) and (c)) and in situ hybridization slides ((dii, div) and (dii, div)). Tbx2 gene controls the development of heart. Histopathological results showed that alteration of the heart as a whole in the LPS injected group. The expression of this gene in different organs showed also a significant difference between the LPS injected group and control. Using the OEO in treatment showed that a modulation effect against LPS effect where it is mentioned.
Our results showed that the effect of LPS before hatching is more than that after hatching and this may be due to the challenge of the tissue during this period but still affected by LPS. So if eggs are treated with a small concentration of OEO before incubation may lead to fiving more healthy chicks.
5. Conclusion
Due to the scarcity of antioxidants for embryo protection against anti-neoplastic agents, newer and better herbal drugs need to be developed with anti-oxidant potentials that are efficient in maintaining peroxidant/anti-oxidant balance. Hence, in the present investigation pretreatment with Oregano oil proved beneficial by restoring anti-oxidant balance and biochemical variables in developing chick embryos. Nonetheless, further characterization of herbal drugs necessitates efficacy studies. Larger trials are obligatory before it is recommended for clinical exercise in future, though it carries immense prospective to be developed as a drug for pharmaceutical diligence.
Author contribution
G.B: Rearing eggs in the experiment, numerical data analysis and histopathological work, A.A.S: Numerical data analysis, RT-PCR work and in situ hybridization work. Both authors shared preparing the manuscript.
Acknowledgment
The authors extend their appreciation to the Deputyship for Research & Innovation, Ministry of Education in Saudi Arabia for funding this research work through the project number IFT20171.
Disclosure statement
No potential conflict of interest was reported by the author(s).
References
- Adaszyńska-Skwirzyńska M, Szczerbińska D. 2017. Use of essential oils in broiler chicken production – a review. An Anim Sci. 17(2):317–335. DOI:10.1515/aoas-2016-0046.
- Alessandra P, Paola S. 2014. The Lipopolysaccharide export pathway in Escherichia coli: structure, organization and regulated assembly of the LPT machinery: Mar. Drugs. 12:1023–1042. DOI:10.3390/md12021023.
- Altop A, Earner G, Durum ME, Isik K. 2018. Effects of essential oils from liquidambar orientalis mill. leaves ongrowth performance, carcass and some organ traits, some blood metabolites and intestinal microbiota in broilers. Br Poult Sci. 59:121–127.
- Baylan M, Akpınar GC, Canogullari SD, Ayasan T. 2018. The effects of using garlic extract for quail hatching egg disinfection on hatching results and performance. Revista Brasileira de Ciencia Avicola 20 (2), pp. 343–350. DOI:10.1590/1806-9061-2017-0693
- Bekhet GA. 2020. Impact of hatching Egg disinfection on hatching characteristics and chick embryos Indian. J Anim Res, B. 1129:1–6.
- Bokariya P, Umarji BN. 2012. The estimation of lethal doses for Acyclovir, insulin and ondansetron on developing chick embryos. IOSR J Pharm. 2:29–31.
- Cetin E, Yibar A, Yesilbag D, Cetin I, Cengiz SS. 2016. The effect of volatile oil mixtures on the performance and ilio-caecal microflora of broiler chickens. Br Poult Sci. 57:780–787.
- Cheng C, Zou Y, Peng J. 2018. Oregano essential oil attenuates RAW264.7 cells from lipopolysaccharide-induced inflammatory response through regulating NADPH oxidase activation-driven oxidative stress. Molecules. 23(8):1857.
- Cloney K, Franz–Odendaal TA. 2015. Optimized ex–ovo culturing of chick embryos to advanced stages of development. J Vis Exp. 95:e52129.
- Cook MI, Beissinger SR, Toranzos GA, Arendt WJ. 2005. Microbial infection affects egg viability and incubation behavior in a tropical passerine. Behav Ecol. 16:30–36.
- Copur G, Arslan M, Duru M, Baylan M, Canogullari S, Aksan E. 2010. Use of oregano (Origanum onites L.) essential oil as hatching egg disinfectant. Afr J Biotechnol. 8:2531–2538.
- De Oliveira MM, Brugnera DF, Cardoso MG, Alves E, Piccol RH. 2010. Disinfectant action of cymbopogon sp. essential oils in different phases of biofilm formation by Listeria monocytogenes on stainless steel surface. Food Control. 21:549–553.
- Esteban RC, Tsukui T, Yonei S, Magallon J, Tamura K, Belmonte IJC. 1999. The T-box genes Tbx4 and Tbx5 regulate limb outgrowth and identity. Nature. 398(6730):814–818.
- Fournomiti M, Kimbaris A, Mantzourani I, Plessas S, Alexopoulos A. 2015. Antimicrobial activity of essential oils of cultivated oregano (Origanum vulgare), sage (salvia officinalis), and thyme (thymus vulgaris) against clinical isolates of Escherichia coli, Klebsiella oxytoca, and Klebsiella pneumoniae. Microb Ecol Health Dis. 26:23289–23295.
- Gabe M. 1976. Histological techniques. Paris: Masson.
- Gardinar HM. 2006. Keeping abreast of advances in fetal cardiology. Early Hum Dev. 82:415–419.
- Giriwono P, Shirakawa H, Hokazono H, Goto T, Komai M. 2011. Fermented barley extract supplementation maintained anti-oxidative defence suppressing lipopolysaccharide-induced inflammatory liver injury in rats. Biosci, Biotech, and Biochem. 75:1971–1976.
- Hang CT, Amack JD, Whipps CM. 2016. Zebrafish embryo disinfection with povidone–iodine. Evaluat Alter Chlorine Bleach Zebrafish. 13:96–101.
- Jones DR, Curtis PA, Anderson KE, Jones FT. 2004. Microbial contamination in inoculated shell eggs: II. Poultry Sci. 83:95–100.
- Kusstatscher P, Cernava T, Liebminger S, Berg G. 2017. Replacing conventional decontamination of hatching eggs with a natural defense strategy based on antimicrobial, volatile pyrazines. Sci Rep. 7:13253. DOI:10.1038/s41598-017-13579-7.
- Leyva LN, Gutiérrez EP, Vazquez OG, Heredia JB. 2017. Essential oils of oregano: biological activity beyond their antimicrobial properties. Molecul. 22(6):989.
- Li D, Hallett MA, Zhu W, Rubart M, Liu Y, Yang Z, Chen H, Haneline LS, Chan RJ, Schwartz RJ, et al. 2011. Dishevelled-associated activator of morphogenesis 1 (Daam1) is required for heart morphogenesis. Develop. 138:303–315.
- Marrelli M, Araniti F, Abenavoli MR, Statti G, Conforti F. 2018. Potential health benefits of Origanum heracleoticum essential oil: phytochemical and biological variability among Di_erent calabrian populations. Nat Prod Commun. 13. 1183–1187.
- Meijerhof R. 2000. Embryo temperature as a tool in the incubation process. Incubation and Fertility Research Group (WPSA Working Group 6 (Reproduction)), St. Edmund’s Hall, Oxford, UK.
- Melo EF, Clímaco WLS, Triginelli MV, Vaz DP, de Souza MR, Baião NC, Pompeu MA, Lara LJC. 2019. An evaluation of alternative methods for sanitizing hatching eggs. Poult Sci. 98(6):2466–2473. DOI:10.3382/ps/pez022.
- Meneses EA, Durango DL, García CM. 2009. Antifungal activity against postharvest fungi by extracts from Colombian propolis quimica. Nova. 32 (8):2011–2017. DOI:10.1590/S0100-40422009000800006. http://www.scielo.br/pdf/qn/v32n8/v32n8a06.pdf.
- Miyake K. 2006. Roles for accessory molecules in microbial recognition by toll-like receptors. J Endotoxin Res. 12:195–204.
- Mousa B, Mohamed TM, Al-Midani RA, El-Samad SA. 2016. Impact of boiler breeders hatching eggs disinfection time on some hatchability parameters. Int J Sci Basic Appl Res. 2016(30):230–240.
- Olivier B, Walter W. 1998. A simplified In situ hybridization protocol using Non-radioactively labeled probes to detect abundant and rare mRNAs on tissue sections. Bio chemica. (1). 10–16.
- Park HJ, Jo SM, Seo HS, Lee M, Lee Y, Kang I. 2020. Anti-Inflammatory potential of cultured ginseng roots extract in lipopolysaccharide-stimulated mouse macrophages and adipocytes. Int J Environ Res Public Health. 17(13):4716. DOI:10.3390/ijerph17134716. Published online 2020 Jun 30.
- Puškárová1 A, Bučková1 M, Kraková1 L, Pangallo1 D, Kozics K. 2017. The antibacterial and antifungal activity of six essential oils and their cyto/genotoxicity to human HEL 12469 cells. Sci Rep. 7:8211. DOI:10.1038/s41598-017-08673-9.
- Rath NC, Huff GR, Huff WE, Balog JM. 2000. Factors regulating bone maturity and strength in poultry. Poultry Sci. 79:1024–1032.
- Roofchaee A, Irani M, Ebrahimzadeh MA, Akbari MR. 2011. Effect of dietary oregano (origanum vulgare L.) essential oil on growth performance, cecal microflora and serum antioxidant activity of broiler chickens. Afric J Biotech. 10(32):6177–6183.
- SAS, Institute. 1998. SAS users guide: statistics. Cary, NC, USA: SAS Institute Inc.
- Sevim B, Gümüş E, Harman H, Ayaşan T, Başer E, Altay Y, Akbulut K. 2020. Effects of dietary rosemary essential oil on growth performance, carcass traits and some hematological values of chukar partridge. Turk J Agri - Food Sci Tech. 8(2):430–435.
- Sturchio E, Boccia P, Zanellato M, Meconi C, Donnarumma L, Mercurio G, Mecozzi M. 2016. Molecular and structural changes induced by essential oils treatments in vicia faba roots detected by genotoxicity testing. J Toxicol Environ Health A. 79(4):143–152.
- Tonelli LH, Holmes A, Postolache TT. 2008. Intranasal immune challenge induces sex-dependent depressive-like behavior and cytokine expression in the brain. Neuropsychopharmacology. 33:1038–1048.
- Vidács A, Kerekes FB, Rajko R, Krisch J. 2017. Optimization of essential oil-based natural disinfectants against Listeria monocytogenes and Escherichia coli biofilms formed on polypropylene surfaces. J Mol Liq. 255. DOI:10.1016/j.molliq.2018.01.179.
- Wang P, Yang FJ, Du H, Guan YF, Xu TY, Xu XW, Su DF, Miao CY. 2011. Involvement of leptin receptor long isoform (LepRb)-STAT3 signaling pathway in brain fat mass- and obesity-associated (FTO) downregulation during energy restriction. Mol Med. 17:523–532.
- Zeweil HS, Rizk RE, Bekhet GM. 2015. Comparing the effectiveness of egg disinfectants against bacteria and mitotic indices of developing chick embryos. J Bas Appl Zoo. 70:1–15.
- Zhang W, Zhang SS, Huang HL, Song BJ, Liu X, Qi Z. 2019. Enhanced endothelin A and B receptor expression and receptor-mediated vasoconstriction in Rat mesenteric arteries after Lipopolysaccharide challenge. Mediators Inflamm. 14(2019):6248197. DOI:10.1155/2019/6248197.