
ABSTRACT
The concepts of integration and modularity refer to the degree of covariation between the components of a biological structure. Components that covariate strongly, but are relatively independent of other modules, are called morphological modules. Morphological integration is understood as the coordinated morphological variation of these components as a functional unit. In mandible case, four modules have been defined based on their differential embryonic origin: alveolar, basal body, coronoid and condylar. The aim was to test these four modular mandible bases using geometric morphometric techniques. The hypothesis of modularity of mandibular development, based on differential embryonic origins, was tested using digital pictures of hemimandibles in lateral form from 26 Araucan horses for those modules. Subsequently, the level of morphological integration between the modules defined above was evaluated using eight landmarks and 65 semi-landmarks, through an analysis of paired blocks of least squares. The divisions between those units presented a modular foundation and the respective level of morphological integration between all considered units. The results of our modular and integrative analysis can be very useful for the comparative study of adaptive processes of other equine breeds.
Introduction
Modularity is an important organizational principle in biological systems, manifesting itself at a morphological level (Klingenberg Citation2014). A module has been defined as a unit within which there is a high integration of many or strong interactions but relatively independent of other such units (Klingenberg Citation2014). Depending on the problem of study and the level of organization studied, the nature of the interactions may vary, which may be genetic, developmental and/or functional (Segura et al. Citation2020). From a morphometric perspective, these interactions are manifested as a strong covariation of the components or parts within a module, and a weak covariation between modules (Curth et al. Citation2017; Heck et al. Citation2018; Romaniuk Citation2018)
Integration refers to the level of cohesion between different structures due to the interactions between the various biological processes that generate the phenotype under study (Klingenberg Citation2009). Morphological integration is usually inferred from the study of covariation between multiple traits (Püschel Citation2014). Therefore, morphological integration usually describes how one structure is linked to another due to various processes (e.g. common developmental cascades; coordinated biomechanical functions; pleiotropies) and/or how two structures vary in coordination (Klingenberg Citation2009; Bertossa Citation2011). In any case, both concepts refer to the degree of covariation between the components of a unit: on the one hand, modularity refers to the relative independence of certain structures, while morphological integration concerns how these units collide with each other (Klingenberg Citation2009; Goswami and Polly Citation2010), so both concepts are related on how the different traits of an organism are correlated.
The mandible is an odd bone, medium and symmetrical, which articulates to the temporal bone, the only mobile bone of the head (Barone Citation2000). The study of the morphology and morphometry of the mandibles is important since it supports most of the structures of the base of the mouth so that it could present differences according to different alimentary adaptations (Barone Citation1999). Despite the growing interest in morphological integration between the different regions of the mandible, the reliability of other modules, defined on functional criteria and/or differential embryonic origins, has not been tested. Moreover, although integration and modularity are studied with morphological data, combining multivariate statistical methods and morphometry, it is interesting to test the hypothesis of integration and modularity of development in animal breeds to contrast different types of development.
Geometric morphometrics (GM) is the study of form covariation with underlying geometric factors, and it is used to determine the ‘shape’ of an individual or structure using discrete and well-defined points in the space (landmarks) (Adams et al. Citation2013). This type of analysis is not affected by the size of the individual and by its rotation or scale, so the shape of the object under study is not described in terms of its dimensions but of the spatial relationship between its parts. This is a fundamental difference from traditional morphometrics (TM), which represents the ‘total form"(Richtsmeier et al. Citation2002). In recent times, GM has gained particular importance, mainly due to the interest of researchers in answering complex questions, which relate morphological variation with functional, developmental or evolutionary norms, among others. GM studies the changes in the displacement in the plane (2D) or space (3D) of the landmarks used to describe the object, thus preserving geometric information. This is a significant advantage for TM, since graphical reconstruction is possible of the morphological change between the cases analyzed from the visualization of the displacements of the landmarks (Toro et al. Citation2010).
Methods based on landmarks analyze morphological changes based on the relative positions of a set of homologous morphometric points (landmarks (points that are present in all individuals in the sample and that correspond in their location, presenting a morphological or functional equivalence). The incorporation of semi-landmarks allows us to include information about curvatures and surfaces using anatomical points. These are arbitrary points distributed along a defined outline between two landmarks. Thus, the GM using semi-landmarks allows representing statistical results as a more real form (and their variations).
The application of GM is very appropriate for studying the phenomena of modularity and morphological integration. These methods show high flexibility, statistical power, and well-developed and coherent mathematical foundations. The combined geometry and multivariate statistical methods ensured a complete and non-redundant characterization of the forms under study, therefore, an adequate representation of the morphologies of interest, allowing this way is the quantitative and multivariate analysis of the phenomena of morphological integration and modularity in mandibles. The study of the morphology and morphometry of the mandibles is important since it supports most of the structures of the base of the mouth, so it could present differences according to different alimentary adaptations (Barone Citation2000).
Araucan horse is adapted to the adverse climatic conditions of the savannah region, and it has not been subjected to any artificial selection process in the environment where it lives. It is an animal of small body size () and resistant to travelling long distances in drought and flood conditions (Salamanca et al. Citation2016, Citation2017). It is used for the management of extensive livestock, for transportation and as a work animal (Salamanca-Carreño et al. Citation2020). Morphological reports on the Araucan horse are available. Biometric studies have shown that the Araucan horse has uniformity in its body measurements as a dolichocephalic and fast-type animal (Salamanca et al. Citation2017; Salamanca-Carreño et al. Citation2020). The study of the Araucan horse breed is justified because it offers information on a breed adapted to environmental conditions of flooded savannah, so with natural adaptive and evolutionary traits, which must be known. Similarly, the breed represents a genetic and cultural heritage that must be studied. In no case, we intend to compare with other breeds.
Figure 1. Araucan horses represent a breed that is adapted to the harsh environmental conditions of flooded savanna in eastern Colombia.
The present research aimed to test a four modular basis in mandibles using GM techniques. Subsequently, an evaluation of the morphological integration between the different modules defined above was carried out. In this sense, the question that followed this research was to describe the level of morphological integration between four mandibular modules (condylar, coronoid, alveolar and basal). In the present work, we use GM using landmarks and semi-landmarks, considering that the latter can help define the mandibular curvilinear contour.
Materials and methods
Sample
We obtained lateral digital records of mandibles from different Araucan horses (Equus caballus L.). Bones were collected from different areas of the Araucan floodplain, eastern Colombia. The final sample consisted of 27 hemi-mandibles, because of the difficulty of collecting bones in the Araucan plains. Mandibles belong to adult animals as they have all their molar series erupted. Lateral plane photographs of the hemi-mandible (right or left side) were taken with a Nikon P530 42X Optical Zoom camera®. Mandibles were aligned to a measurement pattern. The camera was placed in a parallel position to the lateral mandibular face. In order not to have distortions or deformations associated with alignment problems of the specimens, all the recommendations were described in Zelditch et al. (Citation2004). These photographic documents are currently kept in the digital collection of the Facultad de Medicina Veterinaria y Zootecnia, Universidad Cooperativa de Colombia, Arauca, Colombia.
The inclusion criteria were considered according to Püschel (Citation2014):
Samples that present sufficient conservation of mandibular morphological relationships.
Samples that present adequate integrity of the morphological structures that define the homologous anatomical landmarks in the lateral norm.
The exclusion criteria were
Those samples that contradict any of the inclusion criteria, such as foals, with a solution of continuity in any of their anatomical structures, or fragmented.
Photographs of low sharpness to distinguish homologous landmarks or radiographic images with obvious distortions due to the enlargement or deformation.
Individuals who show signs of having been deformed.
Age and sex data since most of the mandibles were collected in the field and unknown information. The decision was made by all authors.
Only a specimen was excluded due to an extensive bone fracture, bringing the final sample size to 26 hemi-mandibles corresponding to 26 animals.
Processing of the primary data
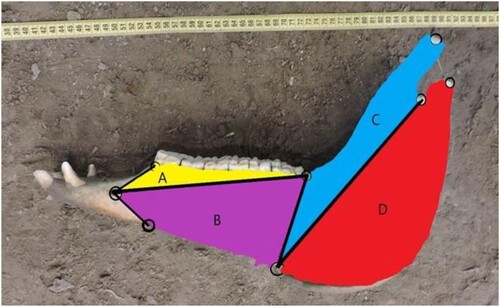
Morphometric data were collected as 2D coordinates of homologous anatomical landmarks using the tpsDig v. 1.40 program (Rohlf Citation2015). In each mandible, eight anatomical discrete landmarks were digitized (), and a set of four groups of 65 semi-landmarks was established considering the following units: condylar (25 semi-landmarks), coronoid (17 semi-landmarks), alveolar (8 semi-landmarks) and basal body (15 semi-landmarks) ().
Figure 2. Map of homologous landmarks (eight) used on lateral hemi-mandible. 1. Mental foramen; 2. Rostral border of second premolar alveolus; 3. Caudal border of molar series; 4. Coronoid process; 5. Mandibular notch; 6. Mandibular condyle; 7. Mandibular angle; 8. Orthogonal projection of landmark 2 onto mandibular body ventral edge.

Figure 3. Modules considered: alveolar (A), basal body (B), coronoid (C) and condylar (D).
We eliminated differences due to rotation, translation and scale through a Generalized Procrustes Analysis (GPA) (Mitteroecker and Gunz Citation2009). The size of the centroid was used as a size estimator. For semi-landmarks, since their location between the landmarks is arbitrary (they are deficient in a coordinate), they are slipped iteratively on the line tangent to the curve to minimize the difference between the configurations and the average shape and thus achieve optimal alignment along the contour (optimizes spacing between semi-landmarks along the curve in relation to the average way to reduce variations due to spacing arbitrary) (Rohlf Citation2010). By this process, the semi-landmarks acquire a geometric correspondence (they become homologous);therefore, it is possible to treat them as typical landmarks (Rohlf Citation2010). This procedure was done using the ‘append tps curve to landmarks’ option of the tpsUtil v. 1.70 software (Rohlf Citation2015). Geometric and statistical morphometric analyses were carried out using the MorphoJ v. 1.07a software (Klingenberg Citation2011).
Coefficient RV
To evaluate a modularity hypothesis, the partition of the homologous landmark configuration into two sub-sets that corresponded to the hypothetical modules (body and branch) was specified. Subsequently, the strength of the covariation between the modules was compared against several possible alternative partitions with the same number of homologous landmarks as the hypothetical modules by calculating the RV coefficient. This coefficient can be considered a multivariate analogue of a correlation and was calculated between the hypothetical modules and between the set of alternative partitions, generating a distribution of values.
Partial least squares
The partial Least Squares (PLS) method is used to study the patterns of covariation between two or more sets of variables. Its usefulness in geometric morphometry lies in the fact that at least one of these two or more sets of variables contain components of the form. Therefore, PLS can be used to relate these form components to other types of data such as experimental conditions, ecological information, or other shape variables. Two-block PLS analysis is based on the single value decomposition method of the covariance matrix between the two sets of variables. This analysis finds correlated pairs of linear combinations (singular vectors) between two sets (or blocks) of variables, which account for the greatest possible covariation between the two blocks of original variables. The aim is to maximize the representation in a few dimensions of the covariance structure between sets of variables (or blocks).
Results
Modularity hypothesis
The modularity of the mandible was analyzed based on four developmental modules (condylar, coronoid, alveolar and basal body) using the RV coefficient. The distribution of the RV coefficients shows that the a priori hypothesis is at the left end of the distribution curve, with the partition with the lowest RV coefficient (RV = 0.521789) ().
Figure 4. Histogram of the distribution of the 1000 RV coefficients calculated from randomly generated modules and with the same number of homologous milestones as the modules hypothesized a priori. The arrow indicates the value of the RV coefficient calculated for the proposed modules. The a priori hypothesis is at the left end of the distribution curve, with the partition with the lowest RV coefficient (RV = 0.521789).
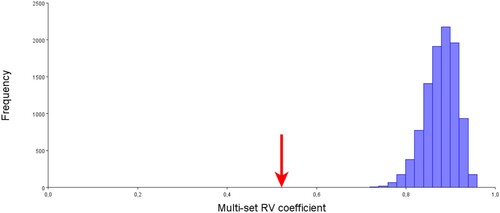
Of a 1,000 alternative partitions to which the RV coefficient was calculated, none has an RV coefficient lower than the one hypothesized in advance. The analysis presented here is good evidence to support the modular organization by differential developmental origins. The result presented here shows a correspondence between the modules defined in advance and those defined as the alternative partitions of lower covariation, which is enough evidence to support the modular division based on differentiated developmental origins.
Partial least squares
The results of the PLS analysis showed a highly significant RV value (p < 0.05), indicating that there is a high integration between different portions of the mandible.
Discussion
Morphological integration of the mandible parts of the Araucan horse was detected due to their spatially close location. In recent years, morphological modularity studies have been developed in several species of mammals (including humans) (Veneziano et al. Citation2018). However, there are a few studies for animal breeds (Parés-Casanova Citation2014; Parés-Casanova and Cabello Citation2020), so this is the first research of morphological integration of the mandible in a locally adapted breed, the Araucan horse.
The different parts of the mandible form a functional unit that is mutually docked. Each of the modules forms a functional unit influenced by a functional matrix acting on the whole bone (Barone, Citation2000). How these processes of gene-environmental interaction occur predominantly determines the observable mandibular morphological variation.
Within the craniofacial growth, that mandible is the last to start and is also the last to finish completing, at least among humans (Thilander Citation1995). The mandible bone is within the craniofacial complex, the bone with the greatest growth potential after birth (Mizoguchi et al. Citation2013). The mandible is a bone whose origin is very different from the rest of the components of the craniofacial complex. It is a mainly membranous bone structure where the development of the condyle responds to the need for adaptation to the environmental changes that can be generated. After birth, only endochondral growth occurs at the level of the articular surface of the condyle, so the pressure to which the mandible is subjected during its function generates a chondrogenic rather than osteogenic response. But based on the evidence presented here, it is feasible to argue that most of the components of the mandible are integrated between them.
Changes in modularity and integration alter the evolutionary limitations of a lineage, and some phenotypes will not be accessible if modularity skews the directions of response to selection. It has been pointed out that changes in modularity and integration will alter the evolutionary limitations of a lineage, and some phenotypes will not be accessible if modularity skews the directions of response to selection. Therefore, the integration and modularity of evolutionary changes in morphometric traits must be quantified to understand how the forms of organisms evolve (Klingenberg Citation2005).
Strong morphological integration between units could lead to more conservative mandibles (e.g. a high degree of morphological change as a whole) and reduced use of resources, while a weak morphological integration between units (e.g. high modularity) could transmit more different regional morphological changes, allowing a great diversification (Vizcaíno et al. Citation2016). When a complex structure meets these modularity requirements, adaptive analysis can be done based on various levels of organization (Dressino and Lamas Citation2010). The present work could test the hypothesis of modularity quantitatively. The degree of modularity on the mandibles of the Araucan horse allows us to infer a diversification in them. In this way, it was evident that all four modules exist but behave relatively dependently. As there are no similar works from other breeds, it is difficult to study breed differences, but our results can be used as a reference for this comparative future research.
In conclusion, detected morphological integration of the Araucan horse mandible parts seemed to indicate that mandibular functions affect them as a whole. But according to the objectives of this study, we cannot elucidate the modular ontogenetic sequence during the mandibular conformation. Anyway, this followed modular and integrative approach could be used as a tool for analyzing adaptive processes for the study of other horse breeds, so our obtained results can be used as a reference for these future comparative research.
Acknowledgements
It was not a necessary approval by the Animal Ethics Committee since animal remains were used in the field.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
Data are available upon request to the second author.
Additional information
Funding
References
- Adams DC, Rohlf FJ, Slice DE. 2013. A field comes of age: geometric morphometrics in the 21st century. Hystrix. 24:7–14. doi:10.4404/hystrix-24.1-6283.
- Barone R. 1999. Anatomie comparée des mamifères domestiques. Tome 1. Vigot Fréres, 5ta. ed.
- Barone R. 2000. Anatomie comparée des mammifères domestiques. Tome 2. Arthrologie et myologie. Vigot.
- Bertossa RC. 2011. Morphology and behaviour: functional links in development and evolution. Phil Trans Royal Soc B: Biol Sc. 366:2056–2068. doi:10.1098/rstb.2011.0035.
- Curth S, Fischer MS, Kupczik K. 2017. Patterns of integration in the canine skull: an inside view into the relationship of the skull modules of domestic dogs and wolves. Zoology. 125:1–9. doi:10.1016/j.zool.2017.06.002.
- Dressino V, Lamas SG. 2010. Una perspectiva modular de la adaptación biológica. Ludus Vitalis. 18:13–24.
- Goswami A, Polly PD. 2010. Methods for studying morphological integration and modularity. (J. Alroy, & G. Hunt, Edits.). Paleontol Soc Pap. 16:213–243. doi:10.1017/S1089332600001881.
- Heck L, Wilson LA, Evin A, Stange M, Sánchez-Villagra MR. 2018. Shape variation and modularity of skull and teeth in domesticated horses and wild equids. Frontiers Zool. 15:1–17. doi:10.1186/s12983-018-0258-9.
- Klingenberg CP. 2005. Developmental constraints, modules and evolvability. In: En BK, editor. Variation. San Diego (CA): Academic Press; p. 219–247.
- Klingenberg CP. 2009. Morphometric integration and modularity in configurations of landmarks: tools for evaluating a priori hypotheses. Evol Develop. 11:405–421. doi:10.1111/j.1525-142X.2009.00347.x.
- Klingenberg CP. 2011. Morphoj: An integrated software package for geometric morphometrics. Mol Ecol Res. 11:353–357. doi:10.1111/j.1755-0998.2010.02924.x.
- Klingenberg CP. 2014. Studying morphological integration and modularity at multiple levels: concepts and analysis. Phil Trans Royal Soc B: Biol Sc. 369:33–35. doi:10.1098/rstb.2013.0249.
- Mitteroecker P, Gunz P. 2009. Advances in geometric morphometrics. Evol Biol. 36:235–247. doi:10.1007/s11692-009-9055-x.
- Mizoguchi I, Toriya N, Nakao Y. 2013. Growth of the mandible and biological characteristics of the mandibular condylar cartilage. Jap Dental Sc Review. 49:139–150. doi:10.1016/j.jdsr.2013.07.004.
- Parés-Casanova PM. 2014. Age-dependent mandibular asymmetries in domestic pigs. Research. 1:4–7. doi:10.13070/rs.en.1.797.
- Parés-Casanova PM, Cabello M. 2020. Patterns of mandibular asymmetries in two types of companion rabbits. J Vet Medic Series C: Anat Histol Embryol. 49:227–232. doi:10.1111/ahe.12517.
- Püschel T. 2014. Modularidad e Integración Morfológica en Cráneos Humanos: un Enfoque Morfométrico Geométrico. Int J Morphol. 32:299–304. doi:10.4067/S0717-95022014000100048.
- Richtsmeier JT, DeLeon VB, Lele SR. 2002. The promise of geometric morphometrics. Y Phys Anthropol. 45:63–91. doi:10.1002/ajpa.10174.
- Rohlf FJ. 2010. Digitalized landmarks and outlines (2.26). Stony Brook. Department of Ecology and Evolution. State University of New York.
- Rohlf FJ. 2015. The tps series of software. Hystrix. 26(1):9–12.
- Romaniuk A. 2018. Functional and phylogenetic aspect in modularity of Palearctic mustelids (Carnivora, Mustelidae) mandible. Vestnik Zool. 52:165–176. doi:10.2478/vzoo-2018-0018.
- Salamanca CA, Parés-Casanova PM, Crosby RA, Monroy N. 2017. Biometric analysis of Araucano Criollo horse. Arch. Zootec. 66:267–278.
- Salamanca CA, Parés-Casanova PM, Vélez TM, Bentez MJ. 2016. Uso de índices zoométricos en la diferenciación racial del caballo criollo de las sabanas inundables araucanas (Colombia). Actas Iberoam Conserv Anim. 7:1–7.
- Salamanca-Carreño A, Jordana J, Crosby-Granados RA, Bentez-Molano J, Parés-Casanova PM. 2020. Lineal discrimination of horses and mules. A sympatric case from Arauca, Colombia. Animals (Basel). 10:1–6. doi:10.3390/ani10040679.
- Segura V, Cassini GH, Prevosti FJ, Andrade MF. 2020. Integration or modularity in the mandible of canids (Carnivora: Canidae): a geometric morphometric approach. J Mammal Evol. 28:145–157. doi:10.1007/s10914-020-09502-z.
- Thilander B. 1995. Basic mechanisms in craniofacial growth. Acta Odontol Scandin. 53:144–151. doi:10.3109/00016359509005964.
- Toro M, Manriquez SG, Suazo GI. 2010. Morfometría geométrica y el estudio de las formas biológicas: de la morfología descriptiva a la morfología cuantitativa geométrica. Int J Morphol. 28:977–990. doi:10.4067/S0717-95022010000400001.
- Veneziano A, Meloro C, Irish JD, Stringer C, Profico A, De Groote I. 2018. Neuromandibular integration in humans and chimpanzees: implications for dental and mandibular reduction in homo. Am J Phys Anthropol. 167:84–96. doi:10.1002/ajpa.23606.
- Vizcaíno SF, Bargo MS, Cassini GH, Toledo N, Vizca SF, Bargo S, Cassini GH, Toledo S. 2016. Forma y función en paleobiología de vertebrados, primera ed. EDULP.
- Zelditch ML, Swiderski DL, Sheets HD. 2004. Geometric morphometrics for biologists: a primer. Elsevier Academic Press. doi:10.1016/B978-0-12-778460-1.X5000-5.