
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Substantial metabolic and oxidative stress and a decrease in immune function have been associated with increased sensitivity to several diseases in dairy cows during the transition period (before and after calving). We investigated the effect of supplementation with linseed (LS), rich in omega-3 polyunsaturated fatty acids (n-3 PUFAs), alone, or in combination with an organic source of selenium, on the plasma metabolic parameters during the transition (10 ± 3 days before to 42 days after calving). A portion of the total mixed ration (TMR) from the control group (C) was replaced with n-3 PUFA-rich LS in both experimental groups (LS and LS + Se), keeping the diets iso-energetic and iso-nitrogenous. In addition, in the LS + Se group, inorganic sodium selenite was replaced by organic selenium. On day 42, we found (a) higher (P < 0.05) LDL level and Sodium ion concentration in the LS group than in the LS + Se and C groups; (b) lower (P < 0.05) concentrations of triglycerides and lactate in the LS and LS + Se groups than in the C group; and (c) higher (P < 0.05) urea concentration in the LS + Se group than in the C and LS groups. In conclusion, n-3 PUFA supplementation acted positively on lipid profile and no adverse effect on plasma metabolic parameters were found.
KEYWORDS:
Introduction
The transition period of dairy cows can be broadly defined as the period from three weeks before to three weeks after calving (Fiore et al. Citation2017a). During this time, cows undergo substantial metabolic and physiological changes in preparation for lactation (Tessari et al. Citation2020; Boushehri et al. Citation2021). Around the parturition, calf nutritional requirements reach a peak, and the mother’s dry matter intake (DMI) drops, resulting in a negative energy balance (NEB) after the onset of lactation (Esposito et al. Citation2014; Sayiner et al. Citation2021). Negative energy balance can be defined as a state in which cows are not consuming adequate calories to cover expenditure for basal needs and milk production, resulting in the mobilization of fat reserves (Delfino et al. Citation2021). This can lead to substantial metabolic and oxidative stress, as well as impaired immune function, which have been associated with an increased incidence of several diseases, including ketosis (Sordillo and Raphael Citation2013; Zachut et al. Citation2020), lipidosis and hormone deficits (Fiore et al. Citation2017b). To ameliorate this effect, numerous additives have been evaluated as supplements to the diets of dairy cows during the transition period (Pulina et al. Citation2020). For example, diets supplemented with omega-3 polyunsaturated fatty acids (n-3 PUFAs) have been shown to improve immune response by modulating the production of cytokines and changing the ratio of immune cells (Agazzi et al. Citation2004). Oxidative stress, which reaches its peak at the time of parturition (Đidara et al. Citation2015), contributes to reduced cellular immunity during the transition period. Supplementation with dietary antioxidants can reduce oxidative stress. For example, selenium (Se), a precursor of at least 25 selenoproteins, plays an important role in the regulation of vital pathways, and contributes substantially to the antioxidant defence network (Surai et al. Citation2019).
In a previous study, we found that feeding transition dairy cows with linseed increases α-linolenic fatty acid content in their milk; furthermore, we found that combining linseed with organic selenium amplifies this effect (Đidara et al. Citation2017). However, it remains unclear whether supplementation with linseed and organic selenium is effective in the prevention of postpartum disease in transition dairy cows. The relationship between the incidence of postpartum diseases in dairy cows and plasma metabolites has been previously demonstrated by Paiano et al. (Citation2019). In this study, we have chosen a number of parameters which helps dairy practitioners to assess the energy and protein status, liver health and mineral status of dairy cows. Thus, the present study aims to determine whether supplementation with linseed, alone, or in combination with organic selenium, has any influence on the plasma metabolic parameters indicative of disease in transition dairy cows.
Material and methods
Experimental design and diets
The experiment is a part of larger research in which the effect of supplementation of diet with linseed, alone, or in combination with organic selenium, upon oxidative stress (Đidara et al. Citation2015) and milk fatty acid composition (Đidara et al. Citation2017) in transition dairy cows have been already tested. The experiment was conducted with 30 Holstein dairy cows at their second parity with an average body weight of 635 ± 37 kg. The cows were randomly allocated into three groups of ten, which were held in separate pens under the same environmental conditions during their transition period (10 ± 3 days before calving to 42 days after calving). All cows were fed the total mixed ration (TMR), which was offered ad libitum, and refreshed daily. A portion of dietary n-6 fatty acids in the control (C) group’s TMR was replaced with n-3 fatty acids in two experimental groups (LS and LS + Se), keeping the diets iso-energetic and iso-nitrogenous. The TMR contained 16.00% crude protein (CP) and net energy for lactation (NEL) values were 6.59, 6.53 and 6.52 MJ/kg dry matter (DM) for the C, LS and LS + Se groups respectively. The TMR was composed of corn silage 8.4 kg DM, grass silage 5 kg DM, grass hay 0.85 kg DM, wet beet pulp 1.7 kg DM and experimental concentrate 9.57 kg DM per cow per day. The experimental concentrate for the control (C) group consisted of ground corn, sunflower, barley, mineral premix and toasted soy. In both experimental groups (LS and LS + Se), a part of toasted soy was replaced with ground linseed. In addition, in the LS + Se group, sodium selenite from the mineral premix was replaced by organic selenium (B-Traxim®, Pancosma, Switzerland). The ingredients and chemical compositions of the experimental diets are shown in . Experimental and animal management procedures were conducted following the ‘Directive for the Protection of Vertebrate Animals used for Experimental and other Purposes’ (86/609/EEC) and Croatian regulation including Animal Protection Act (NN 102/17, NN32/19).
Table 1. Ingredients and chemical composition of the diet supplemented with linseed, alone, or in combination with organic selenium.
Chemical analyses of diets
The analyses of the diets were carried out in duplicate using the AOAC (Citation2000) method to determine: dry matter (934.01); nitrogen and crude protein content by destruction in block (992.23); crude fibre by a ceramic fibre filter method (962.09); and ether extract by the Soxhlet method (991.36 AOAC, Citation2000).
Blood sample collection and analytical procedure
At the same time of the day (three hours after feeding), on days −10 ± 3, 1, 21 and 42 relative to parturition, blood samples of all cows were collected from the coccygeal vein and stored in tubes containing lithium heparin anticoagulant (Becton Dickinson, Plymouth, England, UK). Within 2 h, samples were centrifuged (1.500 g, 10 min at 4°C) and plasma was separated and frozen at −80°C until analysis. Biochemical parameters were determined using an automatic clinical chemistry analyser (Beckman Coulter AU400, Beckman Coulter, Germany). Globulin concentrations were calculated by subtracting albumin levels from the total protein concentration. Low-density lipoprotein (LDL) concentrations were calculated from total cholesterol, triglyceride and high-density lipoprotein (HDL) values (cholesterol – HDL – triglycerides/2.2). The concentrations of non-esterified fatty acid (NEFA) and β-hydroxybutyrate (BHB) were determined using commercial kits (Randox Laboratories Ltd., Crumlin, UK) by an enzymatic colorimetric method.
Statistical analysis
Descriptive statistics for plasma biochemical parameters were performed using STATISTICA (TIBCO Software Inc. 2018). The assumption of normality was checked using the Shapiro–Wilk test. For the evaluation of the effect of treatment and days relative to parturition on the variability of biochemical parameters in the plasma of individual animals, the following fixed statistical model was used:
where yijk = estimated trait (biochemical parameter in plasma); μ = intercept; Ti = fixed effect of diet i (groups = C, LS and LS + Se); Dj = fixed effect of day j (j = −10, 1, 21 and 42 days relative to parturition); eijky = residual.
The significance of the differences between the analysed traits due to the fixed effect of treatment and time, as well as any interactions, was tested by post hoc Fischer's test at the level of P < 0.05; PROC MIXED procedure for repeated measurements over animals in STATISTICA (TIBCO Software Inc. 2018).
Results
Day × diet interaction
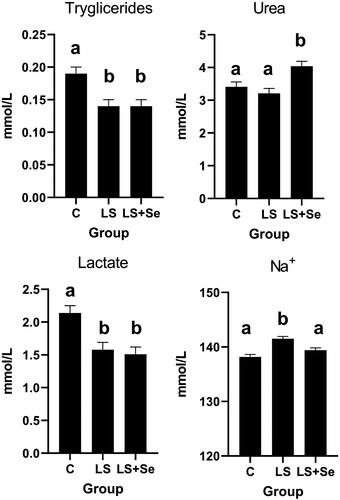
The plasma metabolic parameters are shown in . A significant Day × Diet interaction was observed for LDL. Specifically, on day 42, LDL concentration was significantly higher (P < 0.05) in the LS group than in the LS + Se and Control groups. For the other parameters, no significant Day × Diet interaction was found.
Table 2. Plasma metabolic parameters in dairy cows fed total mixed ration supplemented with linseed and organic selenium during the experimental period.
Effect of diet
When a significant effect of diet existed, a post hoc Fisheŕs test was used to determine differences among diets (). A significant difference among diets was observed for triglyceride, urea and lactate concentrations (). Specifically, a higher (P < 0.05) concentration of triglycerides and lactate was observed in the control group than in to the LS and LS + Se groups. On the other hand, significantly higher (P < 0.05) urea concentrations were observed in the LS + Se group than in the C and LS groups. Concentration of Na+ was significantly higher (P < 0.05) in the LS group than in the C and LS + Se groups.
Figure 1. Some metabolic parameters in dairy cows fed total mixed ration supplemented with linseed and organic selenium during the experimental period (C – Control group; LS – Linseed group; LS + Se – Linseed + Selenium group; a, b – values with different letters differ significantly at P <0.05).
Effect of day
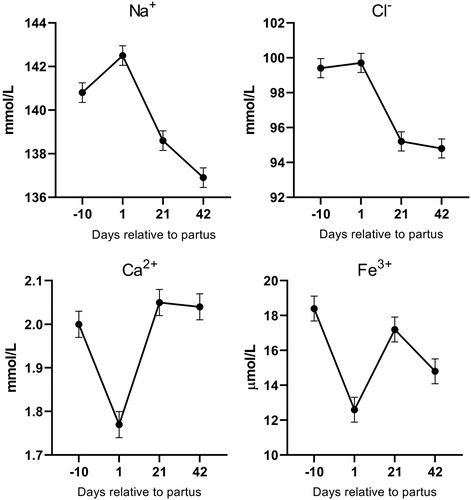
Similarly to diet, a post hoc Fisheŕs test was used when a significant effect of day relative to parturition on blood plasma parameters existed (). From the energy and protein parameters, a significant effect of day (P < 0.05) was observed for total protein, globulin, urea, triglycerides, NEFA, BHB and Lactate (). Most of these parameters except triglycerides rise during the post-partum period, with NEFA and BHB concentrations reaching peak at day 1 postpartum and falling afterword’s. From the enzyme and hepatic parameters, a significant increase (P < 0.05) was observed in the post-partum period for, AST, cholesterol, HDL, LDL (). Finally, from mineral parameters, a significant effect (P < 0.05) was observed for Na+, Cl−, Ca2+ and Fe3+ concentrations (). A significant decrease in Fe3+ and Ca2+ was observed on day 1 post-partum.
Figure 2. Protein and energy parameters in plasma of dairy cows during the experimental period (NEFA – non-esterified fatty acids; BHB – β-hydroxybutyrate).
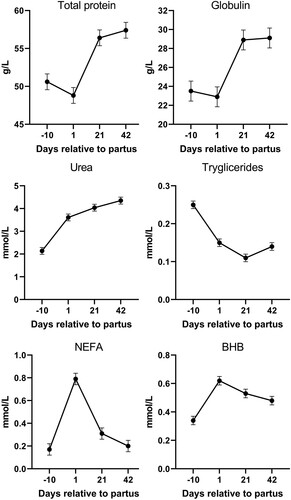
Figure 3. Enzymes and hepatic parameters in plasma of dairy cows during the experimental period (AST – aspartate aminotransferase; ALT – alanine aminotransferase; LDL – Low-density lipoprotein; HDL – high-density lipoprotein).
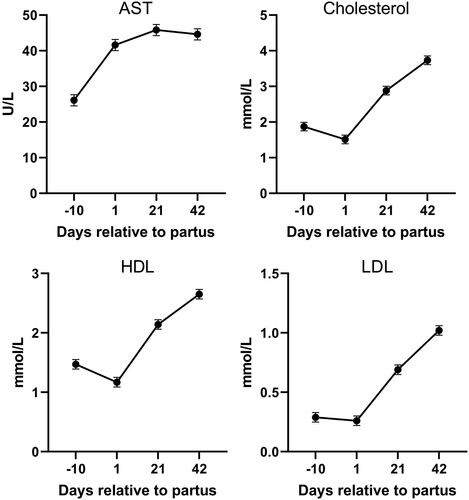
Figure 4. Mineral parameters in plasma of dairy cows during the experimental period.
Discussion
On day 42, LDL concentration was significantly higher (P < 0.05) in the LS group, and triglyceride and lactate levels were significantly lower (P < 0.05) in both experimental (LS and LS + Se) groups compared to the controls during the whole transition period. This observation is in agreement with Harris (Citation1997), who observed reduced serum triglyceride concentrations and increased serum LDL cholesterol levels, with no significant effect on total cholesterol, after n-3 fatty acid supplementation in humans. The beneficial effects of n-3 PUFA supplementation on dyslipidaemia have been described in the literature. Pizzini et al. (Citation2017) summarized the mechanism by which n-3 PUFAs promote intracellular catabolism of apolipoprotein B-100-containing lipoproteins, suppress their hepatic production, stimulate triglyceride clearance from plasma via lipoprotein lipase, increase the conversion rate of very-low-density lipoprotein (VLDL) to LDL, and reduce LDL synthesis. Docosahexaenoic acid (DHA), one of the most important n-3 fatty acids, reduces triglyceride concentrations but increases LDL serum concentrations (Balk et al. Citation2006; Wei and Jacobson Citation2011; Bernstein et al. Citation2012). In contrast to these findings, Chang et al. (Citation2009) argue that n-3 rich diets decrease arterial total LDL delivery.
A significantly higher (P < 0.05) urea concentration was observed in the LS + Se group than in the C and LS groups. This could be a consequence of increased intake of Se, which can lead to an imbalance of ruminally degradable and undegradable proteins (Roseler et al. Citation1993; Amirteymoori et al. Citation2021). In an in vitro experiment, Kim et al. (Citation1997) concluded that Se supplementation can influence rumen microbial fermentation. Supplementation with selenomethionine (an organic form of selenium) altered rumen fermentation by linearly increasing total volatile fatty acids and the molar proportions of propionate and butyrate, but rumen pH, ammonia content and the ratio of acetate to propionate were decreased. Compared with sodium selenite, selenomethionine enhanced the molar proportion of propionate in the rumen and the apparent digestibility of crude protein, neutral detergent fibre, acid detergent fibre and selenium (Wei et al. Citation2019). Thus, a higher plasma urea concentration in the LS + Se group could be the consequence of increased crude protein degradation, higher ammonia flux to the liver, and consequently greater metabolism of ammonia to urea.
Significantly lower (P < 0.05) blood lactate concentrations were observed in both experimental groups. This observation agrees with positive effect of feeding n-3 fatty acids on lowering lactate levels found by other authors. In the study of Pomposelli et al. (Citation1991) dietary treatment with n-3 fatty acids ameliorated the lactic acidosis induced by endotoxin. The reduction in lactate concentration was associated with improved aerobic metabolism that would limit lactate production or/and increase lactate clearance rate. Mickleborough (Citation2013) found that a fish oil supplemented diet in rats significantly decreased the activity of lactate dehydrogenase in some muscles, an enzyme that catalyses the conversion of lactate to pyruvate and back.
Sodium is one of the major electrolytes in the extracellular fluid and is responsible for maintaining its volume and osmotic pressure. Sodium concentration in cow blood plasma ranges within 134 and 156 mmol/L (Skrzypczak et al. Citation2014). In this study, Na+ concentration was significantly higher (P < 0.05) in the LS group than in the C and LS + Se groups. In men with moderate cardiovascular risks, the consumption of fish oil capsules for 12 weeks caused a reduction in urinary sodium excretion (Vandongen et al. Citation1993). In our experiment, we have not measured urinary sodium excretion, but we can assume that lower excretion of sodium due to the higher n-3 dietary levels can increase sodium in the blood.
There was a significant effect (P < 0.05) of day relative to parturition on multiple energy and protein parameters (), enzyme and hepatic parameters (), and mineral parameters ().
A drop of the Fe3+ on 1st day post-partum should be interpreted in light of the immune system physiology. Changes in total protein concentration corresponded with changes in globulin levels. Both innate and acquired immune responses are weaker around calving (Sordillo et al. Citation2009) which is reflected in lower globulin levels; a major portion of globulins consist of immunoglobulins, which provide humoral immunity. A weakened immune system during the transition period makes cows more prone to bacterial infection. Within hours of infection or other inflammatory stimuli, plasma iron concentrations decrease, often below 10 µM. A common mechanism of hypoferremia of inflammation is a cytokine-driven increase in hepcidin, which downregulates ferroportin and thereby decreases iron flow into the extracellular fluid from all sources. Thus, under the influence of increased concentrations of hepcidin, recycled iron is retained in the macrophages of the liver and spleen, and iron absorption is decreased (Ganz Citation2017).
Urea plasma concentration steadily increased from day −10 pre-calving in all groups (). This was most likely due to changes of diet composition from dry to lactating period, specifically a higher proportion of crude protein. NEFA and BHB concentrations were higher (P < 0.05) on day one after calving, with NEFA levels higher than physiological levels (>0.7 mmol/L) (Mulligan et al. Citation2006). This indicates that animals are in a negative energy balance (Knob et al. Citation2021). When in NEB, energy from fat reserves is mobilized (ketosis), causing plasma NEFA concentrations to rise (Antunović et al. Citation2021). Low glucose and gluconeogenic substrate supply slows down the Krebs cycle in the liver cells and increases the production of ketone bodies, including BHB (White Citation2015). Although BHB levels were higher (P < 0.05) on day 1 after parturition, it remained within the physiologically recommended level (<1.40 mmol/L; Fiore et al. Citation2018).
In our study, plasma triglyceride concentration was lower (P < 0.05) after parturition compared to day −10. Van den Top et al. (Citation2005) found that fatty liver in dairy cows postpartum is associated with decreased plasma triglyceride concentration and decreased lipoprotein lipase activity in adipocytes. A decrease in adipose tissue lipoprotein lipase activity possibly helps to channel fatty acids away from adipose tissue and into the milk gland, which requires large quantities of triglycerides for milk synthesis. Lower lipoprotein lipase activity during the postpartum period could therefore be the reason for the significantly higher (P < 0.05) total cholesterol, LDL, and HDL in the postpartum period compared to day −10 prepartum.
A significant (P < 0.05) decrease in plasma calcium concentration on day 1 postpartum, below physiological values of 1.8 mmol/L could be interpreted as subclinical hypocalcaemia (Thilsing-Hansen et al. Citation2002). The risk of postparturient hypocalcaemia occurs due to the high demand for calcium for milk synthesis, low gut absorption capacity and bone tissue mobilization.
Similarly to our results, Skrzypczak et al. (Citation2014) reported an increase in plasma concentrations of sodium in the last week of pregnancy, with the highest concentration on the day of delivery, and a statistically significant drop during the first two postnatal weeks. An increase in sodium concentration immediately before parturition may be related to an increase in aldosterone concentration observed immediately before calving and during the first week of lactation. The observed drop in sodium concentration during the first weeks of lactation may be a consequence of decreased plasma renin activity, which can cause higher sodium loss through urine (Skrzypczak et al. Citation2014). Furthermore, regulation of chloride in blood is tightly associated with regulation of sodium, which is reflected in our results; peak chloride concentrations were observed on the day of parturition, with lower concentrations during the first week of lactation.
Conclusions
A diet rich in n-3 PUFAs acts positively on lipid profile and does not exacerbate the negative energy balance, or further reduce immune response during the transition period. Furthermore, we found that linseed supplementation increases triglyceride and lactate clearance, demonstrated by lower triglyceride and lactate content in blood plasma. Cows supplemented with linseed showed no change in hepatic enzyme production, demonstrating a minimal impact of the dietary supplementation on liver function. We found no adverse effects of n-3 PUFA-rich linseed supplementation, alone, or in combination with organic selenium, on the blood plasma parameters of dairy cows during the transition period.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
The data that support the findings of this study are available from the corresponding author, M.Đ., upon reasonable request.
References
- Agazzi A, Cattaneo D, Dell’Orto V, Moroni P, Bonizzi L, Pasotto D, Bronzo V, Savoini G. 2004. Effect of administration of fish oil on aspects of cell-mediated immune response in periparturient dairy goats. Small Rumin Res. 55:77–83.
- Amirteymoori E, Khezri A, Dayani O, Mohammadabadi M, Khorasani S, Mousaie A, Kazemi-Bonchenari M. 2021. Effects of linseed processing method (ground versus extruded) and dietary crude protein content on performance, digestibility, ruminal fermentation pattern, and rumen protozoa population in growing lambs. Ital J Anim Sci. 20(1):1506–1517.
- Antunović Z, Mioč B, Klir ŠŽ, Širić I, Držaić V, Đidara M, Novoselec J. 2021. The effect of lactation stage on the hematological and serum-related biochemical parameters of the Travnik Pramenka ewes. Poljoprivreda. 27:56–62. doi:10.18047/poljo.27.2.7.
- AOAC. 2000. Official methods of analysis. 17th ed. Vol. 2. Arlington (VA): Association of Official Analytical Chemist Inc.
- Balk EM, Lichtenstein AH, Chung M, Kupelnick B, Chew P, Lau J. 2006. Effects of omega-3 fatty acids on serum markers of cardiovascular disease risk: a systematic review. Atherosclerosis. 189:19–30.
- Bernstein AM, Ding EL, Willett WC, Rimm EB. 2012. A meta-analysis shows that docosahexaenoic acid from algal oil reduces serum triglycerides and increases HDL-cholesterol and LDL-cholesterol in persons without coronary heart disease. J Nutr. 142:99–104.
- Boushehri M, Sadeghi AA, Chamani M, Aminafshar M. 2021. Effects of antioxidants and prebiotics as vegetable pellet feed on production performance, hematological parameters and colostrum immunoglobulin content in transition dairy cows. Ital J Anim Sci. 20(1):1863–1869.
- Chang CL, Seo T, Matsuzaki M, Worgall TS, Deckelbaum RJ. 2009. n-3 fatty acids reduce arterial LDL-cholesterol delivery and arterial lipoprotein lipase levels and lipase distribution. Arterioscler Thromb Vasc Biol. 29:555–561.
- Delfino NC, Silva RDG, Alba HDR, Oliveira MXS, de Carvalho GGP, Araújo MLGML, Pina DS, de Freitas Júnior JE. 2021. Milk yield and composition, blood, and urinary parameters of Murrah buffaloes in different maturity stages during the transition period and early lactation. J Appl Anim Res. 49(1):247–256.
- Đidara M, Poljičak-Milas N, Milinković-Tur S, Mašek T, Šuran J, Pavić M, Kardum M, Šperanda M. 2015. Immune and oxidative response to linseed in the diet of periparturient Holstein cows. Animal. 9:1349–1354.
- Đidara M, Šperanda M, Domaćinović M, Novoselec J, Mašek T, Balenović M, Poljičak Milas N, Vukšić N. 2017. Plasma and milk fatty acid composition as a response to dietary n-3 fatty acids and selenium in periparturient Holstein cows. Mljekarstvo. 67:123–129.
- Esposito G, Irons PC, Webb EC, Chapwanya A. 2014. Interactions between negative energy balance, metabolic diseases, uterine health and immune response in transition dairy cows. Anim Reprod Sci. 144:60–71.
- Fiore E, Giambelluca S, Morgante M, Piccione G, Vazzana I, Contiero B, Orefice T, Arfuso F, Gianesella M. 2017a. Changes in thyroid hormones levels and metabolism in dairy cows around calving. Acta Vet. 67:318–330.
- Fiore E, Piccione G, Perillo L, Barberio A, Manuali E, Morgante M, Gianesella M. 2017b. Hepatic lipidosis in high-yielding dairy cows during the transition period: haematochemical and histopathological findings. Anim Prod Sci. 57:74–80.
- Fiore E, Piccione G, Rizzo M, Morgante M, Barberio A, Giudice E, Gianesella M. 2018. Adaptation of some energetic parameters during transition period in dairy cows. J Appl Anim Res. 46:402–405.
- Ganz T. 2017. Iron and infection. Int J Hematol. 107:7–15.
- Harris WS. 1997. N-3 fatty acids and serum lipoproteins: human studies. Am J Clin Nutr. 65:1645S–1654S.
- Kim J, Van Soest PJ, Combs GF. 1997. Studies on the effects of selenium on rumen microbial fermentation in vitro. Biol Trace Elem Res. 56:203–213.
- Knob DA, Scholz AM, Kappes R, Rodrigues WB, Alessio DRM, Perazzoli L, Mendes BPB, Neto AT. 2021. Dry matter intake, body condition score, beta-hydroxy-butyrate, milk yield, and composition of Holstein and crossbred Holstein x Simmental cows during the transition period. J Appl Anim Res. 49(1):6–14.
- Mickleborough TD. 2013. Omega-3 polyunsaturated fatty acids in physical performance optimization. Int J Sport Nutr Exerc Metab. 23:83–96.
- Mulligan FJ, O’Grady L, Rice DA, Doherty ML. 2006. A herd health approach to dairy cow nutrition and production diseases of the transition cow. Anim Reprod Sci. 96:331–353.
- Paiano RB, Lahr FC, Silva LSB, Marques DS, Ferreira CA, Birgel DB, Birgel EH. 2019. Haematological and biochemical profiles during the puerperium in dairy cows. Acta Vet Hung. 67:377–384.
- Pizzini A, Lunger L, Demetz E, Hilbe R, Weiss G, Ebenbichler C, Tancevski I. 2017. The role of omega-3 fatty acids in reverse cholesterol transport. Nutrients. 9:1099.
- Pomposelli JJ, Flores EA, Blackburn GL, Zeisel SH, Bistrian BR. 1991. Diets enriched with N-3 fatty acids ameliorate lactic acidosis by improving endotoxin-induced tissue hypoperfusion in Guinea pigs. Ann Surg. 213(2):166–176.
- Pulina G, Tondo A, Danieli PP, Primi R, Crovetto GM, Fantini A, Macciotta NPP, Atzori AS. 2020. How to manage cows yielding 20,000 kg of milk: technical challenges and environmental implications. Ital J Anim Sci. 19(1):865–879.
- Roseler DK, Ferguson FD, Sniffen CF, Herrema I. 1993. Dietary protein degradability effects on plasma and milk urea nitrogen and milk nonprotein nitrogen in Holstein cows. J Dairy Sci. 76:525–534.
- Sayiner SD, Ergene OI, Aslan S. 2021. Changes in antioxidant enzyme activities and metabolic parameters in dairy cows during different reproductive periods. Theriogenology. 159:116–122.
- Skrzypczak W, Kurpińska A, Stański Ł, Jarosz A. 2014. Sodium, potassium and chloride homeostasis in cows during pregnancy and first months of lactation. Acta Biol Cracoviensia s. Zool. 55/56:58–64.
- Sordillo LM, Contreras GA, Aitken SL. 2009. Metabolic factors affecting the inflammatory response of periparturient dairy cows. Anim Health Res Rev. 10:53–63.
- Sordillo LM, Raphael W. 2013. Significance of metabolic stress, lipid mobilization, and inflammation on transition cow disorders. Vet Clin N Am Food Anim Pract. 29:267–278.
- Surai PF, Kochish II, Fisinin VI, Juniper DT. 2019. Revisiting oxidative stress and the use of organic selenium in dairy cow nutrition. Animals. 9:462.
- Tessari R, Berlanda M, Morgante M, Badon T, Gianesella M, Mazzotta E, Contiero B, Fiore E. 2020. Changes of plasma fatty acids in four lipid classes to understand energy metabolism at different levels of non-esterified fatty acid (NEFA) in dairy cows. Animals. 10:1410.
- Thilsing-Hansen T, Jørgensen R, Østergaard S. 2002. Milk fever control principles: a review. Acta Vet Scand. 43:1.
- Van den Top AM, Van Tol A, Jansen H, Geelen MJ, Beynen AC. 2005. Fatty liver in dairy cows postpartum is associated with decreased concentration of plasma triacylglycerols and decreased activity of lipoprotein lipase in adipocytes. J Dairy Res. 72:129–137.
- Vandongen R, Mori TA, Burke V, Beilin LJ, Morris J, Ritchie J. 1993. Effects on blood pressure of n-3 fats in subjects at increased risk of cardiovascular disease. Hypertension. 22:371–379.
- Wei JY, Wang J, Liu W, Zhang KZ, Sun P. 2019. Effects of different selenium supplements on rumen fermentation and apparent nutrient and selenium digestibility of mid-lactation dairy cows. J Dairy Sci. 102:3131–3135.
- Wei MY, Jacobson TA. 2011. Effects of eicosapentaenoic acid versus docosahexaenoic acid on serum lipids: a systematic review and meta-analysis. Curr Atheroscler Rep. 13:474–483.
- White HM. 2015. The role of TCA cycle anaplerosis in ketosis and fatty liver in periparturient dairy cows. Animals. 5:793–802.
- Zachut M, Šperanda M, de Almeida AM, Gabai G, Mobasheri A, Hernandez-Castellano LE. 2020. Biomarkers of fitness and welfare in dairy cattle: healthy productivity. J Dairy Res. 87:4–13.