
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.ABSTRACT
Eight artificially reared lambs were used to study locomotory and feeding activities in the first three weeks of life by actigraphy. Lambs were fitted with a Bluetooth-enabled (BT) accelerometer and data were downloaded as activity counts at 1-min intervals (Vector Magnitude, VM). Sensors were programmed to act as beacons, and two sensors programmed as receivers were installed next to the rubber nipples of the milk feeder and recorded the serial numbers and labels of other nearby beacons through the BT signals. Mean (±SE) VM was 140 ± 3 counts/min, and time of day and week had significant (P < 0.001) effects. Overall activity did not differ significantly between sexes (males: 139 ± 5; females: 142 ± 5). The proportion of lambs that exhibited a 24-h circadian rhythm decreased with age (week 1 = 75%, week 2 = 63%, week 3 = 50%). Mean number of suckling sessions/day was 3.7 ± 0.2, the mean number of minutes suckling/day was 12.5 ± 0.9, and mean number of minutes/meal was 3.5 ± 0.2. Males dedicated more time/meal than females (males: 4.1 ± 0.4; females: 3.0 ± 0.2 min; P < 0.05). In conclusion, actigraphy is a useful tool for investigating the locomotor and feeding behaviour of artificially reared lambs, which detected a reduction in the circadian rhythmicity and the number of suckling sessions with age.
KEYWORDS:
Introduction
Worldwide there are about 40 million milk-producing sheep (Morris Citation2017), and most lambs remain with their mothers for about one month if local dairy breeds are managed under semi-extensive systems; however, the spread of high-producing breeds such as Assaf and Lacaune that are bred under ultra-intensive management systems leads to the separation of lambs from their mothers at birth and its artificial rearing. Automatic feeders are used in large-scale lamb rearing systems to reduce labour costs and improve lamb growth by increasing milk allowance. An understanding of the development of feeding behaviour from birth to adulthood, and how this might affect lamb health and performance, is important for the future of sheep management systems and for the optimization of artificial lamb-rearing systems (Nieper Citation2017). The acclimation of the lambs to the automatic feeder is essential because suckling is an important behavioural trait that is suggestive of problems and is an early indicator of the lambs’ welfare. Improvements in the management of artificially reared lambs require detailed information about milk consumption and the behavioural patterns of lambs in the milking facility, and a large number of individual milk consumption profiles is necessary (David et al. Citation2014). In that case, the automatic continuous recording of feeding events is preferable to visual feeding inspections because examining visually many animals for an extended period is too time consuming. David et al. (Citation2014) presented an electronic automatic lamb feeder that records the milk consumption of individual lambs based on the detection of electronic ear tags and an antenna fitted to the feeder, which is used to calculate the quantity of milk consumed and the time spent suckling by the lamb. Based on accelerometers attached to lambs that suckled their mothers, Kuźnicka and Gburzyński (Citation2017) showed that suckling is indicated by a relatively simple and distinct acceleration signature, which makes it possible to detect the event based on relatively simple techniques that are easily implemented with low-end microcontrollers. In addition, they provided an algorithm that was based on the acceleration data that had been collected from animals on a farm.
Accelerometers have been extensively used to track everyday human behaviours such as sleep and exercise (Emery et al. Citation2010). The accelerometer provides information on the amount, frequency, and duration of the activity, as well as information on diurnal activity, and the intensity of the exercise. Significant advances have been made in triaxial accelerometers and they are widely used in veterinary medical research as a non-invasive way to record all small-scale body movements of an animal. Many of the traditional limitations on direct observations of animals in the field have been overcome because of the advantages of accelerometry over conventional means of tracking the activities of farm or companion animals (Brown et al. Citation2013). In previous studies, we have investigated the locomotor activity and circadian rhythmicity of lambs under natural lactation based on data collected by actigraphy, and have quantified the relationships between lambs and their twins by calculating their proximity and distance based on Bluetooth-enabled (BT) accelerometers (Abecia et al. Citation2022).
The aim of the present study was to quantify the locomotor and feeding activities of lambs in the first three weeks of artificial rearing, and their circadian rhythms based on data collected by actigraphy. Feeding activity was measured by calculating the proximity and distance to the milk-feeding machine by BT-enabled accelerometers.
Material and methods
Ethical approval
The study was performed at the experimental farm of the University of Zaragoza (Zaragoza, Spain; latitude 41°41’N) following a protocol that was approved by the Ethics Committee of the University of Zaragoza based on the requirements of the European Union for Scientific Procedure Establishments.
Animals
Eight Rasa Aragonesa twin lambs [4 males, 4 females; mean (±SD) birth live weight = 4.02 ± 0.35 kg] born from four ewes at the end of January were used in the study. Ewes and lambs were allocated to individual boxes (2 m x 2 m) immediately after lambing, where they remained for 48 h, which allowed the lambs to consume the necessary quantity of colostrum from their mothers. Thereafter, lambs were separated from their dams, housed in a straw-bedded pen (5 m x 3 m) with 12 other lambs of the same age, and artificially fed reconstituted milk replacer (Elvor Spécial Agneaux, Elvor, Maen Roch, France) (25% crude protein; 25% fat) by an automatic lamb feeder (TAPO-EZ1-32-M, Förster-Technik GmbH, Engen, Germany). In addition, the lambs had access to barley straw and clean water and, from the third week of age, a highly palatable starter diet (Ovirum Harina, Oviaragón, Zaragoza, Spain). Although the facility that housed the lambs was naturally-lit through through windows, the staff began its activity at 0800 h and the artificial lighting remained on until noon. As such the lambs were illuminated until sunset (1840h).
Accelerometer data collection
Lambs were fitted with commercially available sensors (46 mm × 33 mm × 15 mm in size, mass = 19 g) that record high resolution raw acceleration data and are Bluetooth-enabled (BT) (ActiGraph wGT3X-BT; ActiGraph, FL, USA), which were attached to the dorsal side of a neck collar that remained in place for 21 d (21 Jan-11 Feb) (). The sensors record accelerations (activity) based on the individual’s amplitude (g) and frequency (Hz) of movement along three axes (x for front-to-back, y for side-to-side, and z for up–down). Sensors were programmed to collect data at a rate of 30 Hz, which is equivalent to 30 samplings per second. Data from the sensors were used to quantify the proximity of the lambs to the milk feeders and to calculate the number of suckling sessions per day. The sensors on the lambs were programmed to act as beacons, and two sensors were installed close to the rubber nipples of the feeder (), which were programmed as receivers that recorded the serial numbers and labels of other beacons (lambs) nearby. Devices were programmed to scan for signals from the beacons once per minute. The beacons were programmed to send signals at 4 Hz (4 times per second).
Figure 1. a) Lambs housed in the artificial rearing facility wearing accelerometers, b) sensors (receivers) installed on the nipple of the milk feeder, c) a lamb suckling, and d) a lamb wearing an accelerometer that is sending a Bluetooth signal (beacon) to the sensor installed in the nipple (receiver).

The activity data were downloaded as activity counts per 1 min intervals by the ActiLife software (ActiGraph, LLC, Pensacola, FL). Actigraph provides three columns of data; i.e. activities in the x-, y-, and z-axes. The activity counts for the three axes were used to create minute-by-minute activity data values (Vector Magnitude, VM), which is the magnitude of the vector that is formed from the combination of the accelerations from the three axes on each device. VM is calculated as follows:
The BT signals received by the sensors on the lambs were downloaded to the ActiGraph software, and the distance between the beacon (lamb) and receiver (nipple) was calculated based on the Received Signal Strength Indicator (RSSI) received by the sensors and a polynomial equation (Ortmeyer et al. Citation2018) to convert the RSSI beacon data from the lamb's sensor to the distance to the feeder as follows:
Proximity of the lambs to the feeder was calculated based on a BT signal ≥ −57, which corresponds to a distance of ≤50 cm from the lamb to the feeder. A meal was indicated if the signal from at least one of the receptors was −57 and lasted for > 2 min. Ambient T (°C) and relative humidity (RH, %) were recorded by mini data-loggers (Testo 174H, Testo SE & Co. KGaA, Titisee-Neustadt, Germany).
Statistical analysis
For each of the three weeks of the study, mean (±S.E.) VM for the 7 d of each week that the lambs wore the sensors was calculated for each hour. The mean daily distribution of VM indicated two peaks in activity, and the mean of the peaks, the time of the peaks, the interval between peaks, and day (0800-1800h) and night (1900-0700 h) activities were calculated. The proportion of the records that had a value = 0, which indicated no activity, was calculated, and the significance of differences among groups was evaluated by a X2 Test. A lamb’s proximity to the feeder was calculated based on the total number of BT signals per hour, and the number of suckling sessions per lamb per day and the time spent suckling were calculated.
Circadian rhythms in VM were graphed by fitting the time-series measurements of each lamb to the cosine curve of a 24-h activity rhythm, which was obtained by the cosinor method at the Cosinor on-line platform (Molcan Citation2019). Midline Estimating Statistic of Rhythm (MESOR), the average around which the variable oscillates, amplitude (the difference between the peak and the mean value of a wave), and acrophase (the time of peak activity) were calculated for each individual. To test for rhythmicity, an F-test compared the (re-parameterized) cosine model and the non-rhythmic model. A P < 0.05 indicated that the time series fit a 24-h rhythm. Thereafter, the data were pooled and the mean 24-h cosinor curves for each of the three parameters were calculated, and the cosinor values of males and females, and among weeks were compared statistically by an ANOVA. A repeated-measures t-test was used to detect significant differences among the first three weeks of the lambs lives.
The effects of week, lamb sex, and time of day (day or night) on locomotor and feeding activity were evaluated statistically based on a multifactorial model that included week, lamb sex, and time of day (day or night) as fixed effects, and the Least Squares Method of the GLM procedure in SPSS v.26 (IBM Corp. Released Citation2019). Within fixed effects, significant differences were identified by an ANOVA. A general representation of the model is as follows: y = xb + e, where y is Nx1 vector of records, b denotes the fixed effect in the model within the association matrix x, and e is the vector of residual effects. Prior to those statistical analyses, a Kolmogorov–Smirnov Test confirmed whether the data were normally distributed.
Results
Ambient temperature and relative humidity
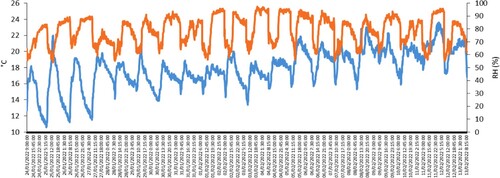
Mean (±SE) ambient T and RH were 17.83 ± 0.05°C and 78.79 ± 0.25%, respectively, and both differed significantly (P < 0.001) between day (18.17 ± 0.06°C, 71.27 ± 0.32%) and night (17.53 ± 0.09°C, 85.42 ± 0.22%) ().
Figure 2. Ambient temperature (T) and relative humidity (RH) in the 3-week experiment involving artificially reared suckling lambs.
Activity
Examples of the pattern of VM of an individual lamb in each of the three weeks are shown in . Mean (±SE) VM was 140 ± 3 counts/min, and time of day (daytime 180 ± 6; nighttime 107 ± 3) and week had significant (P < 0.001) effects, such that lambs exhibited the highest activity in week 2 and the least activity in week 3 (). Males and females did not differ significantly in activity in any of the three weeks () or overall (males: 139 ± 5; females: 142 ± 5).
Figure 3. Example of an actogram of a lamb in the first week of life as rendered by ActiLife software (ActiGraph, LLC) (activity counts per 1-min intervals).
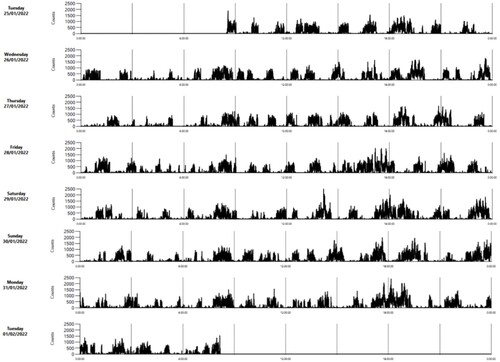
Figure 4. Example of an actogram of a lamb in the second week of life as rendered by ActiLife software (ActiGraph, LLC) (activity counts per 1 min intervals).
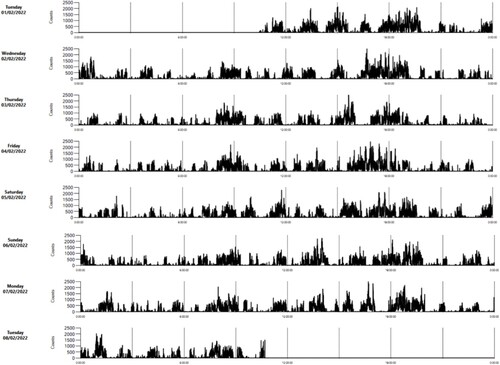
Figure 5. Example of an actogram of a lamb in the third week of life as rendered by ActiLife software (ActiGraph, LLC) (activity counts per 1-min intervals).
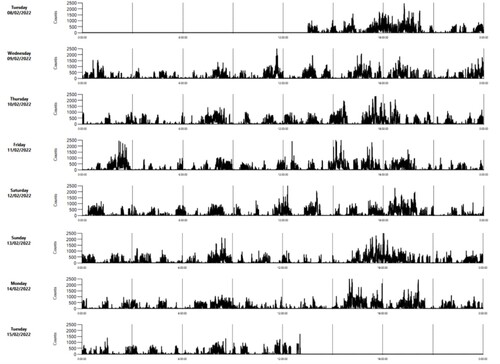
Table 1. Mean (±S.E.) activity (Vector Magnitude, VM) of lambs artificially fed reconstituted milk replacer by an automatic lamb feeder, in their first three weeks of life as measured by actigraphy, and their circadian rhythms (a,b, c; P < 0.05 among weeks; x,y; P < 0.05 between daytime and nighttime) (daytime: 8:00-18:00; nighttime: 19:00-7:00).
Table 2. Mean (±S.E.) activity (Vector Magnitude, VM) of male and female lambs artificially fed reconstituted milk replacer by an automatic lamb feeder in their first three weeks of life as measured by actigraphy (a,b within sex; P < 0.05).
In the three weeks of the study, the first peak in activity for each of the lambs occurred at 0800 h, but the timing of the second peak differed significantly (P < 0.01) among the three weeks (). The first peak was significantly (P < 0.05) higher than was the second, and activity at night was significantly (P < 0.001) higher in week 1 than it was in weeks 2 and 3. On average, lambs spent approx. 50% of their time inactive, and the time spent inactive was significantly (P < 0.05) lower in week 2 than it was in weeks 1 and 3 ().
Circadian rhythmicity
Most of the lambs exhibited the same pattern of activity; viz., two peaks in the day (at sunrise and sunset) and low activity at night. The curves of mean daily activity in weeks 1–3 are shown in . The proportion of lambs that exhibited a 24-h circadian rhythm decreased in the three weeks (wk 1 = 75%, wk 2 = 63%, wk 3 = 50%) (). The amplitude of the circadian rhythm was significantly (P < 0.05) higher in week 2 than it was in weeks 1 and 3, and acrophase did not differ significantly among weeks ().
Figure 6. Mean (±S.E.) daily activity (counts/min/h) of lambs in the first, second, and third weeks of life as measured by actigraphy (upper panel), and the corresponding cosinor curves of a 24-h activity rhythm (dark areas represent night) (lower panel).
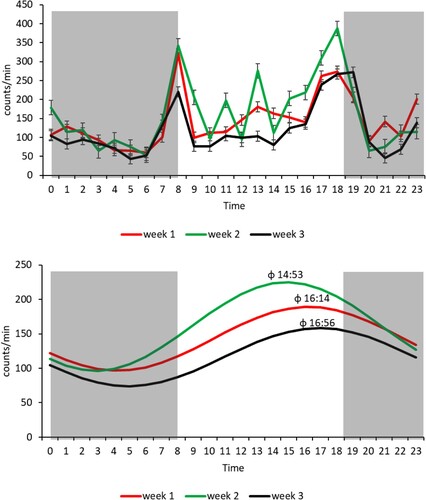
Males and females exhibited significantly (P < 0.05) higher activity in week 2 than they did in weeks 1 and 3, and VM did not differ significantly between the sexes in the first three weeks of life ().
Suckling activity
Mean (±S.E.) number of suckling sessions per lamb per day was 3.7 ± 0.2, mean total time was 12.5 ± 0.9 min, and mean time per meal was 3.5 ± 0.2 min. The number of suckling sessions per day and the total time per week were significantly (P < 0.05) lower in week 3 than they were in weeks 1 and 2 (). The time per meal did not differ significantly among weeks. Males and females did not differ significantly in the number of meals per day (males: 3.8 ± 0.3, females: 3.4 ± 0.3) or the time spent suckling per week (males: 14.9 ± 1.5; females: 10.1 ± 0.8 min); however, males (4.1 ± 0.4 min) spent significantly (P < 0.05) more time per suckling session than did females (3.0 ± 0.2 min).
Table 3. Mean (±S.E.) number of suckling sessions in the first week of life by lambs artificially fed reconstituted milk replacer by an automatic lamb feeder as measured by actigraphy (a,b; P < 0.05).
Discussion
In our study, artificially reared lambs exhibited a clear pattern in locomotor activity, with differences between day and night, a peak in activity at 0800 h, when the lights to the facility were turned on, and a second peak in the afternoon, which was identical to the patterns that we have documented in lambs reared by their mothers (Abecia et al. Citation2022), when the peaks in activity coincided with sunrise and sunset. If lambs are reared by their mothers under intensive management conditions, the morning peak in activity of lambs coincides with the time when dams go to feed. At that moment, the lambs normally rush rapidly back and forth in synchrony, and as the movement slows down, other behaviours take over (Sachs and Harris Citation1978). The second peak in activity of the lambs, in the evening, is influenced by their mother’s behaviour; e.g. she might be feeding on straw that has been offered ad libitum or ruminating, which often occurs in the evening, before dusk (Dulphy et al. Citation1997). In our experiment, however, the second peak of activity did not coincide with any other event associated with their management. Stephens and Baldwin (Citation1971) documented some ambling behaviour in the afternoon in lambs that had been artificially reared, which was interspersed with playing and suckling. Overall, lambs spent 50% of their time inactive. Veissier and Stefanova (Citation1993) reported that artificially reared lambs spent 59.7% and 15.8% with the chin resting and lying in the head-up position, respectively, and Steiber et al. (Citation2008) reported that lambs spent 50-60% of the time laying in the first three days of life.
In our study of artificially reared lambs, the largest proportion of lambs that exhibited a 24-h circadian rhythm occurred in the first week of life (75%), and the proportion decreased thereafter. McMillen and Nowak (Citation1989) found that, at three to ten weeks of age, dark-phase melatonin concentrations were significantly lower in lambs that suckled pinealectomized ewes than that in lambs that suckled pineal-intact ewes, which suggests that either melatonin was transferred to the suckling lamb from the ewe or the lack of exposure of the fetus to the maternal melatonin rhythm might have delayed the emergence of the melatonin rhythm in the newborn lamb. In our study, the lambs were gestated by pineal-intact ewes and, probably, the progressive decrease in the proportion of lambs that exhibited a 24-h locomotor rhythm was because of the loss of a 24-h melatonin secretion rhythm caused by the absence of maternal milk in the diet of artificial reared lambs. In fact, exogenously administered melatonin can entrain circadian activity rhythms in the adult rat (Redman et al. Citation1983), and our research has shown that the proportions of single and twin lambs that had been reared by their mothers exhibited a 24-h circadian rhythm were 25% and 50%, respectively, in the first week of life, which increased to 75% and 100%, respectively, in the fourth week of life (Abecia et al. Citation2022).
In the present experiment, locomotor activity did not differ significantly between the sexes. Some have reported sexual differences in postnatal motility in lambs reared by their mothers (Rurak et al. Citation2008) or differences between the sexes in the proportions of lambs that mounted, butted, or gambolled (Sachs and Harris Citation1978). May et al. (Citation2008) observed that, although there were no sexual differences in ewe–lamb distance or locomotor activity, female lambs synchronized their behaviour with their mother more than did males, which resulted in closer ewe–female lamb distances. Probably, those sex-specific behavioural differences are not expressed under artificial rearing because the role of the mother in the development of that behaviour is absent.
In our study, the number of suckling session per day recorded by the accelerometers was similar to that observed by Stephens and Baldwin (Citation1971), who observed 4–5 suckling session per day by each lamb in the first five weeks of life. In that study, lambs spent, on average, 13.1, 10.8, and 10.3 min suckling in the first three weeks of life, respectively, which were slightly higher than in our study. In an experiment that involved 95 artificially reared lambs under continuous lightning, David et al. (Citation2014) reported 9.5 suckling sessions per lamb per day that had a mean duration of 247 s (approx. 4 min), and a mean daily time spent feeding of 38 min. The presence of continuous lighting influenced the results, although suckling session duration was similar to that observed in our study. As in our experiment, Fletcher (Citation1971) observed that the frequency of suckling by lambs from their mothers was high in the first two weeks post-partum, decreased abruptly between the second and third weeks, and decreased more slowly and relatively uniformly up to the twelfth week of lactation. In our study, the lamb diet was supplemented with a starter feed in the third week of life, and straw was available from the beginning of the experiment. Although consumption of starter feed and straw was not measured, this additional food might have reduced milk intake. Sevi et al. (Citation2003) estimated that suckling lambs consume approx. 60 g of concentrate per day in the suckling period.
Although the sexes did not differ significantly in the number of suckling sessions per day or in the time spent suckling, male lambs spent more time per suckling session than did females. David et al. (Citation2014) reported that the number of meals per day was slightly higher for female lambs, but males consumed on average 161 ml more milk per day than did females; consequently, daily milk intake was similar in males and females. In lambs reared by their mothers, Teke and Akdag (Citation2011) reported significant differences between the sexes in suckling duration per bout, and female lambs exhibited a higher frequency of suckling. Stapleton et al. (Citation1980) found no sex differences in the frequency of, duration of, and interval between suckling periods.
Conclusion
In conclusion, actigraphy has been shown to be an effective tool for documenting the locomotor and feeding behaviour of artificially reared lambs, and detected a decrease in the circadian rhythmicity and the number of suckling sessions with lamb age.
Authors contribution
Conceptualization; Data curation; Formal analysis; Investigation; Methodology; Project administration; Resources; Software; Supervision, and writing of original draft were carried out by Jose A. Abecia and Francisco Canto.
Acknowledgments
We thank Bruce MacWhirter for the English revision of the manuscript. The authors acknowledge the use of Servicio General de Apoyo a la Investigación-SAI, Universidad de Zaragoza, and José Antonio Ruiz and Antonio Barrio for their help in the care of the animals. F. Canto was funded by the National Agency for Research and Development (ANID)/ Scholarship Programme / Doctorado Becas Chile/ 2020 - 72210031.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Abecia JA, Canudo C, Palacios C, Canto F. 2022. Measuring lamb activity during lactation by actigraphy. Chronobiol Int. 39:1368–1380.
- Brown DD, Kays RW, Wikelski M, Wilson RP, Klimley AP. 2013. Observing the unwatchable through acceleration logging of animal behavior. Anim Biotelemetry. 1:1–16.
- David I, Bouvier F, Ricard E, Ruesche J, Weisbecker JL. 2014. Feeding behaviour of artificially reared Romane lambs. Animal. 8:982–990.
- Dulphy JP, Martin-Rosset W, Dubroeucq H, Ballet JM, Detour A, Jailler M. 1997. Compared feeding patterns in ad libitum intake of dry forages by horses and sheep. Livest Prod Sci. 52:49–56.
- Emery EE, Woodhead EL, Molinari V, Hunt MG. 2010. Handbook of assessment in clinical gerontology. Lichtenberg (PA): Academic Press. Chapter, Treatment Adherence in Late-Life. 427–457.
- Fletcher IC. 1971. Relationships between frequency of suckling, lamb growth and post-partum oestrous behaviour in ewes. Anim Behav. 19:108–111.
- IBM Corp. Released. 2019. Ibm SPSS statistics for windows, version 26.0. Armonk, NY: IBM Corp.
- Kuźnicka E, Gburzyński P. 2017. Automatic detection of suckling events in lamb through accelerometer data classification. Comput Electron Agric. 138:137–147.
- May R, van Dijk J, Forland JM, Andersen R, Landa A. 2008. Behavioural patterns in ewe–lamb pairs and vulnerability to predation by wolverines. Appl Anim Behav Sci. 112:58–67.
- McMillen IC, Nowak R. 1989. The pre- and postnatal development of hormonal circadian rhythms. Baillieres Clin Endocrinol Metab. 3:707–721.
- Molcan L. 2019. Time distributed data analysis by cosinor. BioRxiv. 805960: Online Application.
- Morris S. 2017. 2 - Woodhead publishing series in food science, technology and nutrition, advances in sheep welfare. Woodhead Publishing. Chapter: Overview of Sheep Production Systems. 19–35.
- Nieper BA. 2017. The behaviour and health of dairy lambs reared artificially with and without early access to meal [master’s thesis]. Massey University.
- Ortmeyer HK, Robey L, McDonald T. 2018. Combining actigraph link and petpace collar data to measure activity, proximity, and physiological responses in freely moving dogs in a natural environment. Animals (Basel). 8:230.
- Redman J, Armstrong S, Ng KT. 1983. Free-running activity rhythms in the rat: entrainment by melatonin. Science. 219:1089–1091.
- Rurak DW, Fay S, Gruber NC. 2008. Measurement of rest and activity in newborn lambs using actigraphy: studies in term and preterm lambs. Reprod Fertil Dev. 20:418–430.
- Sachs BD, Harris VS. 1978. Sex differences and developmental changes in selected juvenile activities (play) of domestic lambs. Anim Behav. 26:678–684.
- Sevi A, Caroprese M, Annicchiarico G, Albenzio M, Taibi L, Muscio A. 2003. The effect of a gradual separation from the mother on later behavioral, immune and endocrine alterations in artificially reared lambs. Appl Anim Behav Sci. 83:41–53.
- Stapleton D, Hinch G, Thwaites C, Edey T. 1980. Effect of sex and litter size on the sucking behaviour of the lamb. Proceedings of the Australian. Society of Animal Production., Biennal Conference.
- Steiber O, Bodnár A, Kispál T. 2008. Impact of the artificial rearing system on Awassi lambs’ behaviour and growth performance – A new approach of sheep husbandry in subtropical countries. Proceedings of the conference on international research on food security, natural resource management and rural development; October 7-9; Tropentag, University of Hohenheim.
- Stephens DB, Baldwin BA. 1971. Observations on the behaviour of groups of artificially reared. lambs. Res Vet Sci. 12:219–225.
- Teke B, Akdag F. 2011. The effect of age, lactation number, sex and birth type on suckling and nursing behaviour of Karayaka lambs. Economic, Social and Environmental Sustainability in Sheep and Goat Production Systems. Options Mediterr. 100:323–327.
- Veissier I, Stefanova I. 1993. Learning to suckle from an artificial teat within groups of lambs: influence of a knowledgeable partner. Behav Processes. 30:75–82.