
Abstract
The Upper Cretaceous Gaogou, Majiachun and Sigou formations in the Xixia Basin are composed of terrestrial clastic deposits, which are dominated by red-violet mudstones, muddy siltstones, siltstones, grayish fine-grained sandstones, intercalated with grayish, off-white medium- and coarse-grained sandstones or conglomerates. They contain 10 ichnogenera and 22 ichnospecies and Rhizoliths, including Arenicolites isp., Archaeonassa isp., Beaconites antarcticus, B. capronus, B. coronus, Cylindricum isp., Gastruichnus xixiaensis, Palaeophycus megas, P. tubularis, P. heberti, P. sulcatus, P. cf. sulcatus, P. striatus, Planolites beverleyensis, P. isp., Scoyenia gracilis, Skolithos verticalis, S. linearis, S. isp., Taenidium irregularis, T. serpentinum and T. barretti. From north to south across the basin, alluvial fan, braided and meandering fluvial and fluvio-lacustrine systems are recognised on the basis of sedimentary features. In reference to them, four ichnoassemblages are recognised: (1) Skolithos-Gastruichnus ichnoassemblage in the channel environment of braided and meandering river, (2) Beaconites-Taenidium ichnoassemblage in the natural levee and crevasse splays, (3) Beaconites-Palaeophycus ichnoassemblage in the flood plain and (4) Taenidium-Scoyenia ichnoassemblage in the periodically exposed fluvio-lacustrine (pond or very shallow lacustrine) sedimentary environments. The present work not only enrich the trace fossils database of terrestrial origin but also offer new data to complete further continental ichnofacies mode and also provide the constraints on palaeoenvironments of the Xixia Basin.
1. Introduction
The Cretaceous strata of the Xixia Basin have recently received greater attention because of the discoveries of dinosaur fossils and dinosaur eggs (Wang et al., Citation1996, Citation2008). In addition, invertebrate trace fossils and assemblages (Hu, Citation2014; Hu, Wang, & Song, Citation2014; Hu, Zhong, & Song, Citation2010; Zhang, Guo, & Zeng, Citation2004) have been reported. However, their more detailed study are necessary for a better recognition of palaeoenvironment of these palaeontologically important deposits. This paper is focused on recently collected specimens of invertebrate trace fossils from the Upper Cretaceous Gaogou, Majiacun and Sigou formations. The objectives of this paper are: (1) to document trace fossils and assemblages recorded in studied fluvial deposits, (2) to discuss the sedimentary features and sedimentary environment and (3) to discuss ichnoassemblages in alluvial fan, braided and meandering fluvial and fluvio-lacustrine systems.
2. Geologic setting
The Xixia Basin is located in southwest Henan Province, China; it occupies the area of 1000 km² (Figure a), on the south-facing slope of the east Qinling Orogenic Belt. It is a rifted, transtensional basin controlled by northwest-trending strike-slip faults. The basin, wider from the eastern side, is fulfilled asymmetrically (Figure b). During the Late Cretaceous, it was filled by red molasses sediments under semi-arid climatic conditions (Wang et al., Citation1996). The deposits have been subdivided, in ascending order, into the Gaogou, Majiacun and Sigou formations according to the body fossils, lithology and lithofacies (Zhu, Citation1997).
Figure 1a. Location of the study area in Henan Province, China
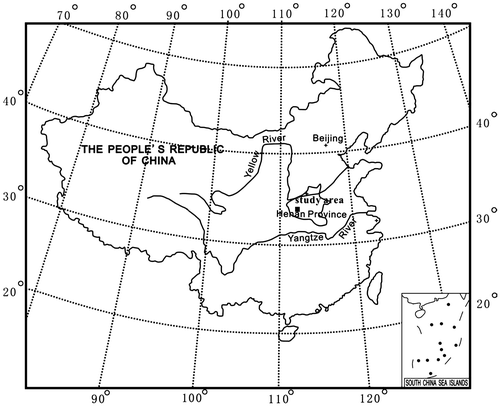
Figure 1b. Location and sketch structural map of the study area in Henan Province, China (■: location of the measured section).
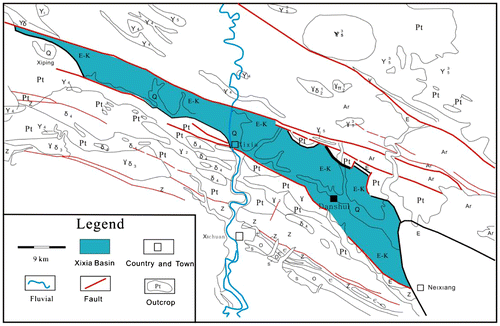
The Gaogou Formation is composed of conglomerates, medium- to coarse-grained sandstones mixed with conglomerates, and coarse- to fine-grained sandstones, alternating with layers of purplish-red, silty mudstones and siltstones (Figure ). These deposits rest with an unconformity on the Devonian Nanwan Formation. The 690-m thick succession is interpreted mainly as alluvial fan, braided fluvial, floodplain and fluvio-lacustrine deposits (Song et al., Citation2008).
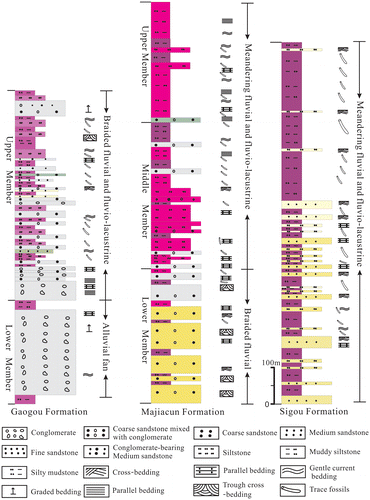
Figure 2. Stratigraphic sections and sedimentary facies of the Upper Cretaceous in Xixia Basin, southwestern Henan Province, China.
The Majiacun Formation, 935 m thick, is consists of coarse-grained sandstones mixed with conglomerates, medium- to coarse-grained sandstones and fine-grained sandstones, interlayered with purplish-red mudstones with abundant calcareous concretions. It rests conformably on the underlying Gaogou Formation (Figure ). The Majiacun Formation was deposited in braided fluvial, meandering fluvial and fluvio-lacustrine systems, with two kinds of ichnofabrics produced in the hard and soft substrates (Zhang et al., Citation2004).
The Sigou Formation comprises mainly red silty mudstones with abundant calcareous concretions, siltstones and grayish-yellow, grayish-green fine-grained sandstones, intercalated with grayish-yellow, medium- to coarse-grained sandstones, and sandstones mixed with conglomerates. The unit is 900 m (Figure ), and deposited in fluvial and fluvio-lacustrine systems (The Henan Geology and Mineral Bureau, Citation1989).
Dinosaurs eggs have been systemically researched since firstly discovered in Majiagou Formation in the spring of 1974. So far, about 15 types of dinosaur eggs are recognised in the Cretaceous of the Xixia Basin, including Faveoloolithus sp., Longiteresoolithus xixiaensis, Dendroolithus sp., Paraspheroolithus sp., P. irenensis, Dictyoolithus neixiangensis, Youngoolithus xiaguanensis, Dendroolithus sp. (Wang et al., Citation1996, Citation2008; Zhou, Feng, & Hui, Citation2001). Ornithopoda dinosaurs skeleton, bivalves, conchostracans, ostracods, gastropods, macrophyte and charophytes previously been reported from the Xixia Basin (Pan, Wang, & Feng, Citation2007; Pan, Xu, & Xi, Citation2008; Wang et al., Citation2006, Citation2008; Xu et al., Citation2000). However, the trace fossils have not been studied in detail. The aim of this paper is to describe the ichnology and sedimentary features, and to interpret the palaeoenvironment of the Upper Cretaceous dinosaur eggs-bearing strata.
3. Depositional environment
3.1. Gaogou Formation
The Gaogou Formation can be divided into two lithologically distinct members that are referred to as the Lower and Upper Members (Figure ). The Lower Member consists of conglomerates, fine- to coarse-grained sandstones mixed with conglomerates and interbedded with silty mudstones. The upper member is dominated by fine- to coarse-grained sandstones with siltstones and mudstones, and the rare occurrence of pebble conglomerates.
The lower part of the Lower Member is mainly conglomerates, and sometimes mixed with brownish red coarse- to fine-grained sandstones containing gravel. The conglomerates are matrix-, clast-supported or rarely cross-stratified. These conglomerates occur as large lenses, which can be traced laterally up to few hundred metres. The pebble grains range from angular to sub-round usually of size between 5 and 15 cm with a maximum of 60 cm. The compositions of the pebbles are complex. Thick, laterally continuous conglomerate beds with erosional scour structures reflect the basal erosion surface of major channels. The upper part of sequence is brownish-red, purplish red muddy siltstones and silty mudstones containing calcarious concretions, sometimes interbedded with coarse-grained sandstones mixed with conglomerates, pebble-bearing medium- to fine-grained sandstones. The sedimentary features include horizontal bedding and trough cross-bedding. Invertebrate trace fossils have not been discovered. Thin, discontinuous conglomerates most likely record minor channel bases. The occurrence of thick mudstone layers reflects a transport by mud flows through torrential streams. Generally, outcrop profiles display a sequence of graded beds that become thinner and finer upwards. All these characteristics point to an alluvial fan.
From the bottom to the top of Gaogou Formation, the lithology association and sedimentary sequences are following: the lower part is predominantly composed of fine to medium, rarely coarse-grained sands and poorly to well-sorted, sub-rounded granule to pebble conglomerates that are moderately to well-sorted massive, with large trough cross-bedding, wedge cross-bedding, graded bedding and massive bedding. Invertebrate trace fossils (Skolithos) are rarely diverse in this association, and burrows are restricted to the uppermost bed.
The upper part of the sequence corresponds to a widespread of muddy siltstones and mudstones, with horizontally stratified, gentle current bedding, trough cross-bedding and parallel bedding. Invertebrate trace fossils are few in these clastic rocks. These deposits corresponds to braided river which proximate to the alluvial fan facies. In addition, the lithofacies associations displayed by alternating layers of brownish-, purplish-red silty mudstones and muddy siltstones, alternating layers of fines-grained sandstones and siltstones, and thick layer calcareous siltstones. Invertebrate trace fossils are abundant in this association. Such as: Palaeophycus, Beaconites, Gastruichnus and Rhizoliths are common. Facies analysis reveals the presence of sandy fluvial channels associated with extensive floodplains. Ponds are developed in the floodplain.
The alluvial fan facies is widely distributed in the lower member of Gaogou Fm., in which scarce sediment was disturbed by invertebrate trace fossils and plant root system. Upwards to the formation, our facies descriptions and the recognition are agreement with the braided-river interpretation. This is consistent with work by Song et al. (Citation2008) and Zhang (Citation2002) who interpreted the depositional environment of Gaogou Formation.
The Calcareous concretion, fibrous gypsum and palygorskite indicates the dry climate during the deposition of the Gaogou Formation, which coincides with the return of warm period after the second temperature-falling event in the world (Wang et al., Citation2008). In a word, the Gaogou Formation was developed in a semi-arid to arid setting typified by alluvial fan and braided fluvial systems.
3.2. Majiacun Formation
The Majiacun Formation can be divided into three lithologically distinct members referred to informally as the Lower, Middle and Upper Members (Figure ).
The lower part of this sedimentary sequence, which is interpreted to form in channel deposition, is characterised by medium- to coarse-grained sandstones with trough and wedge cross-bedding, mixed with pebble-bearing sandstones. The upper part is alternating layers of brownish red siltstones and silty mudstones, in which the silty mudstone beds are commonly thicker than the siltstone beds, which represent the overbank deposition. The fluvial architecture is characterised by high channel/overbank ratio. Trace fossils Beaconites, Palaeophycus and Skolithos were found in the sandstones, siltstones and mudstones. Based on these results, the Gaogou Formation is interpreted to have been deposited in braided fluvial system.
The Middle to Upper Members of the Majiacun Formation to be the deposit of a sandy meandering fluvial system that was affected by extreme flooding events. The lower part of this sedimentary sequence is comprised by coarse-grained sandstones mixed with conglomerates, pebble-bearing medium- to fine-grained sandstones, invertebrate trace fossils are scarce, planar cross-bedding, trough cross-bedding and parallel bedding are displayed, which is interpreted to form in channel deposit systems. The upper part is alternating layers of brownish-, purplish- red mudstones, silty mudstones and siltstones, sometimes intercalated grayish-yellow, offwhite fine- to medium-grained sandstones, with coal streak, asymmetric gentle current bedding and parallel bedding. Sandstone and mudstone display trace fossils including, Palaeophycus, Beaconites, Scoyenia, Planolites, Cylindricum and Skolithos. This facies is interpreted as meandering floodplain deposits, including overbank, crevassing fan and levee settings.
The Upper Member of the Majiacun Formation displayed in extreme shallow lake. The lithology is alternating layers of fine-grained sandstones and silty mudstones with parallel and current bedded, purplish red mudstones with horizontal to gentle current bedding. The thickness of silty mudstones and mudstones is greater than sandstones. The occurrence of black calcareous mudstones, and invertebrate trace fossils Beaconites, Scoyenia, Taenidium and Palaeophycus are discovered in this facies. These elements are interpreted to record deposition within shallow lakes and ponds.
In conclusion, the depositional environments of the Sigou Formation are braided and meandering fluvial system and fluvio-lacustrine systems are distributed spatially. This is consistent with work by Zhang (Citation2002), Zhang et al. (Citation2004) who interpreted the fluvial systems of the Majiacun Formation, and based on numerous paleocurrent measurements, a recognised shift in fluvial drainage patterns is observed in the Middle Member. These data demonstrate a change in paleoflow directions between the middle and upper unit from north and northeastward to northwestward direction (Zhang, Citation2002). Proxies for a semi-arid interpretation for the Majiacun Formation include the presence of calcareous concretions and crumb, the desiccation cracks are common on the bedding plane of layer (Figure ), the palygorskite and fibrous gypsum discovered in the Majiacun Formation (Wang et al., Citation2008).
3.3. Sigou Formation
The sequence characteristic of Sigou Formation is corresponds to fine-grained sandstones and siltstones with subordinate amount of granules and pebbles, which is always present as the lowermost deposits at the base of each complete fining upward cycle. It consists of wedge cross-bedding, parallel bedded and deformed bedding. The thin layers of muddy siltstones, silty mudstones and fine-grained sandstones deposited by repeated episodes of flooding, which represent the uppermost part of the underlying depositional cycle. The sedimentary features includend massive, horizontal bedding, gentle current, flaser bedding and desiccation cracks. Abundant invertebrate trace fossils have been discovered from the Sigou Formation, including Taenidium, Beaconites, Palaeophycus, Scoyenia, Planolites, Cylindricum, Arenicolites, Archaeonassa and Skolithos. Interbedded and finely laminated sandstone and siltstone units of variable geometry are interpreted to represent levee and flood-basin deposits that flanked the major channel. Lateral accretion surfaces suggest high-sinuosity rivers. The presence of shrinkage cracks and caliche nodules indicates subaerial exposure during sedimentation. The mud cracks, absence of root system and presence of palygorskite indicate the dry climate during the deposit period of the Sigou Formation (Wang et al., Citation2008).
Based on lithology, physical sedimentary structures, sedimentary sequences, bed boundaries and trace fossil contents of the Late Cretaceous of Xixia Basin, Henan Province, three kinds of depositional environment were recognised in the succession studied (Figure ): (1) alluvial fan system, (2) braided fluvial and fluvio-lacustrine system and (3) meandering fluvial and fluvio-lacustrine system.
4. Trace fossil descriptions
4.1. Ichnogenus Skolithos Haldemann, Citation1840
The systematics of Skolithos has been reviewed by Alpert (Citation1974, Citation1975), and six valid ichnospecies can be considered, including Skolithos annulatus, S. bulbus, S. ingens, S. linearis, S. magnus and S. verticalis. However, as is claimed, Skolithos needs a detailed ichnospecific revision (Gillette, Pemberton, & Sarjeant, Citation2003; Gregory, Campbell, Zuraida, & Martin, Citation2006; Melchor, Bedatou, de Valais, & Genise, Citation2006; Schlirf & Uchman, Citation2005).
Figure 3. Occurrence characteristics of Skolithos and Palaeophycus in the Upper Cretaceous of Xixia Basin. (A) S. linearis, (B) S. verticalis, (C, D) S. magnus, (E-a) P. tubularis, (E-b) P. heberti.

4.1.1. S. linearis Haldemann, 1840 (Figure (A))
Vertical or subvertical, steeply inclined and slightly curved cylindrical burrows, preserved as endichnia in very fine-grained to silty sandstones. Its diameters ranges from 3 to 6 mm, and the length varied from 3 to 12 cm. The fill is massive mudstones, different than the host rock. Burrow margins are distinct. S. linearis is common in the Gaogou and Majiacun formations.
4.1.2. S. verticalis Hall, 1843 (Figure (B))
Vertical or steeply inclined cylindrical burrows, 3–5 mm in diameter and taper downward, the length varied from 1 to 1.4 cm. Burrow wall is smooth and margin are indistinct and irregular, filling is brownish-red massive mudstones. The burrow preserved as endochnia in siltstones, and was only observed in Majiacun Formation.
Inverted cone to cylindrical burrow (Gao et al., Citation2013), the longer and sandy type of fill distinguishes S. linearis from S. verticalis (Carmona, Buatois, Mángano, & Bromley, Citation2008).
4.1.3. S. magnus Howell, Citation1944 (Figure (C) and (D))
Cylindrical to subcylindrical, vertical and slightly curved burrows, preserved as endochnia in fine- to coarse-grained sandstones. Diameters range from 7 to 21 mm, and length is 5–18 cm. Burrow fill is brownish-red massive mudstones. These burrows were observed in Gaogou and Majiacun formations.
4.2. Ichnogenus Palaeophycus Hall, Citation1847
The systematics of Palaeophycus has been reviewed by Pemberton and Frey (Citation1982), who interpreted Palaeophycus as a passively filled dwelling burrow of a worm-like organism. Palaeophycus is distinguished from Planolites by the presence of a burrow lining in the former ichnogenus, which is lacking in Planolites (Keighley & Pickerill, Citation1995).
4.2.1. P. tubularis Hall, Citation1847 (Figure (E))
The ichnospecie is a simple, unbranched, straight to curved burrow with a distinct lining, 5–30 mm in diameter, visible length ranges from 2.8 to 15 cm, having different or identical infill to the host rock. Burrow wall is smooth with no ornamentations. These burrows are parallel or inclined to the bedding plane. Overcrossing of different specimens is common. Preserved on the bedding plane and intrastratal sandstones and mudstones. Thinly lined burrows are present as convex epirelief and endochnia in the Upper Cretaceous of Xixia Basin.
P. tubularis is distinguished from the other ichnospecies of Palaeophycus by its thin burrow wall and lack of conspicuous wall ornamentation (Pemberton & Frey, Citation1982).
4.2.2. P. heberti de Saporta, Citation1872 (Figure (E))
The burrows are straight to curved, horizontal to vertical, unbranched and lined. Burrow wall is smooth with no ornamentations. The wall lining is 1–5 mm in thickness. Diameters range from 4 to 30 mm and the length varies from 5 to 10 cm. Burrow infill material is different to the host rock. Burrows are present as convex epireliefs and endochnia in the Upper Cretaceous Gaogou and Majiacun formations.
Figure 4. Occurrence characteristics of Palaeophycus in the Upper Cretaceous of Xixia Basin. (A) P. sulcatus, (B) P. striatus, (C-a) Palaeophycus cf. sulcatus, (C-b) Rhizoliths, (D) P. megas.

4.2.3. P. sulcatus Miller and Dyer, Citation1878 (Figure (A))
Horizontal or high dipping, thin lined, straight to curved, unbranched cylindrical burrows, 5–10 mm in diameter, 11 cm long burrows. Anastomosing wrinkles are moderately well preserved. The burrow was filled with material different to the host rock and discovered only in the Upper Cretaceous Sigou Formation.
4.2.4. Palaeophycus cf. sulcatus (Figure (C))
Vertical to high dipping, thin lined, curved, unbranched cylindrical burrows. Anastomosing wrinkles are less preserved in the burrow wall. Diameter varies from 14 to 30 mm diameter, maximum length is 41 cm, which are filled with material different to the host rock and discovered only in the Upper Cretaceous Sigou Formation.
4.2.5. P. striatus Hall, Citation1852 (Figure (B))
The cylindrical burrow is vertical, slightly curved, thin lined and unbranched. Continuously parallel transverse striations are moderately well preserved in burrow wall. Burrow diameter ranges from 12 to 18 mm and nearly 10.6 cm long. They are filled with material different to the host rock and found only in the Upper Cretaceous Sigou Formation.
Palaeophycus striatus differs from P. alternatus the presence of fine, longitudinal striae on the burrow wall (Pemberton & Frey, Citation1982).
4.2.6. P. megas Hu, Citation2014 (Figure (D))
The burrows are curved, horizontal to vertical, unbranched and lined. Burrow wall is smooth without ornamentation. Diameters range from 13 to 55 mm and the length is 9–50 cm. Burrow filling is different to the host rock. The burrows are present as convex epireliefs and endochnia in the Upper Cretaceous Gaogou and Majiacun formations.
The ichnospecie is bigger, slightly curved to curved, and with variable dimensions, which differed from the other ichnospecies of Palaeophycus (Hu, Citation2014).
4.3. Ichnogenus Beaconites Vialov, Citation1962
Revision and relationship of the ichnogenus Beaconites can be found in several papers (Keighley & Pickerill, Citation1994, Citation1995; Schlirf, Citation2000; Vialov, Citation1962). Beaconites is interpreted as feeding structure of a worm-like anima (Schlirf, Citation2000). The presence of a lining distinguishes Beaconites from Taenidium (Keighley & Pickerill, Citation1994; Schlirf, Citation2000).
Figure 5. Occurrence characteristics of Beaconites and Gastruichnus in the Upper Cretaceous of Xixia Basin. (A) B. coronus, (B) B. antarcticus, (C-a) B. coronus, (C-b), B. capronus, (D, E) G. xixiaensis.

4.3.1. B. coronus Vialov, Citation1962 (Figure (A))
They are horizontal to vertical, straight to slightly curved, unbranched, lined, meniscate burrow, 4–22 mm in diameter, 4–25 cm in length. The lining is ~1 mm in thickness. The width of meniscate is lesser than the diameter. They are preserved as convex epirelief and full relief structures, and having identical or different infill material to the host rock. The abundant specimens are discovered in the Upper Cretaceous of Xixia Basin.
4.3.2. B. antarcticus Vialov, Citation1962 (Figure (B))
The specimens are horizontal to overtical, straight to slightly curved, unbranched, lined, meniscate burrows. Burrow diameter varies from 10 to 20 mm and maximum length is 33 cm. The meniscate packets are wider than overall burrow width, they are preserved as convex epireliefs and full relief structures, and having identical or different infill material to the host rock. The abundant specimens are discovered from the Upper Cretaceous in Xixia Basin.
4.3.3. B. capronus Howard et Frey, 1984 (Figure (C))
The specimens are horizontal to vertical, straight to slightly curved, lined, unbranched burrows, 15–20 mm in diameter, 4–10 cm in length. They are characterised by the chevron-shaped meniscates and preserved as convex epirelief and full relief structures, and having identical infill material to the host rock. The abundant specimens are discovered from the Upper Cretaceous in Xixia Basin.
4.4. Ichnogenus Gastruichnus
4.4.1. G. xixiaensis Hu, Citation2014 (Figure (D) and (E))
The burrow is a larger, high dipping, slightly curved and unbranched, with nodular and annular grains preserved in burrow wall, and showing partial distension and upward to the largest in terminal. The burrow is stomach-shaped and preserved as endochnia in siltstones. The burrow diameter varies, and 5–34 cm in length observed. Burrow filling is massive mudstone that is different to the host rock (siltstone). Burrow distribution is relatively concentrated in the Gaogou and Sigou formations, they preserved as full relief structures, and may be used to reproduction of mollusc.
G. xixiaensis is interpreted as produced by deposit-feeders, probably mollusc breed in terrestrial settings (Hu, Citation2014). In contrast to Macanopsis, the burrows are circular to elliptical in cross section, displaying diameter diminishing downward.
4.5. Ichnogenus Taenidium Heer, Citation1877
Taenidium is distinguished from Scoyenia by the absence of wall striations and lining (Keighley & Pickerill, Citation1994). The unwalled nature of the specimen differentiates it from Beaconites (Goldring & Pollard, Citation1995; Keighley & Pickerill, Citation1994). Taenidium ichnospecies can be differentiated by the variation within the meniscate infill. Such as, Menisci of T. serpentinum (Heer, Citation1877) are much more widely spaced. T. barretti (Bradshaw, Citation1981) is much larger, with hemispherical to deeply arcuate menisci, commonly filled with coarser material. Menisci of T. irregularis are more arcuate and backfill organised in nested series, the burrow diameter is uneven (Li, Citation1993).
Figure 6. Occurrence characteristics of Taenidium and Scoyenia in the Upper Cretaceous of Xixia Basin. (A-a) T. serpentinum, (A-b) T. irregularis, (B) T. barrette, (C) S. gracilis.

4.5.1. T. serpentinum Heer, Citation1877 (Figure (A))
Horizontal to oblique, straight to curved, unbranched, unwalled, meniscate burrows. Meniscates are generally arc-shaped and merge laterally at the burrow boundary. Burrow diameter is 4–16 mm, and maximum length is 15 cm. The overlap of the sample is common. Sometimes, intersection part is radially arranged from the centre of the burrow. The burrow is preserved as convex epireliefs in the Sigou Formation.
4.5.2. T. irregularis Li, Citation1993 (Figure (A))
The meniscate burrows are horizontal to oblique, straight to curved, unbranched, and unwalled. Diameter ranges from 4 to 16 mm, maximum length is 25 cm. The Menisci is generally arc-shaped and merge laterally at the burrow boundary. The space between menisci is ~1 mm. The shape and size of menisci is irregular, and can be encircled by the latter. The abundant specimens are discovered from the Upper Cretaceous in Xixia Basin.
4.5.3. T. barretti Bradshaw, Citation1981 (Figure (B))
Horizontal to oblique, straight to meandering trace, unbranched, unwalled, meniscate burrows which range from 6 to 15 mm in diameter. Menisci are generally hemispherical and not full merge laterally at the burrow boundary. Maximum length is 25 cm. Preserved as convex epireliefs. The abundant specimens are discovered from the Upper Cretaceous in Xixia Basin.
4.6. Ichnogenus Scoyenia White, Citation1929
4.6.1. S. gracilis White, Citation1929 (Figure (C))
It is a slightly curved, tubular burrow with a crescent backfill structure, up to 12 cm in length and 7 mm in width. The surface longitudinal striations are only partly observed, because the burrow is not well preserved in all specimens observed. The burrows occur in red-silty mudstones and infill material is identical to the host rock. They are oriented parallel to the bedding plane. S. gracilis is preserved in the Upper Cretaceous Majiacun and Sigou formations.
The ichnospecie is usually regarded as burrow made by deposit-feeding organisms in different non-marine environments, and well-developed longitudinal striations of the wall distinguish Scoyenia from otherwise similar meniscate backfilled structures, such as the smooth-walled Beaconites and Taenidium (Frey, Pemberton, & Fagerstrom, Citation1984).
Figure 7. Occurrence characteristics of Planolites in the Upper Cretaceous of Xixia Basin. (A) P. beverleyensis, (B) Planolites isp.

4.7. Ichnogenus Planolites Nicholson, 1873
4.7.1. P. beverleyensis Billing, 1862 (Figure (A))
Abundant specimens from all sections comprise smooth, straight to curved, horizontal to slightly inclined, unbranched and cylindrical burrows. Burrows are 6–14 mm in diameter and up to 12 cm in length. Burrow filling is massive mudstone and darker than host rock. They are preserved as convex epirelief and endoreliefs in the Upper Cretaceous Majiacun and Sigou formations.
Planolites was reviewed by Pemberton and Frey (Citation1982) and Fillion and Pickerill (Citation1990), which was interpreted as fodinichnion and referred to polyphyletic vermiform deposit-feeders producing active backfills (Rodríguez-Tovar & Uchman, Citation2004). P. beverleyensis is very similar to Planolites montanus, but the former being larger in size and more tortuous (Pemberton & Frey, Citation1982). In this sense, Keighleyn and Pickerill (Citation1997) analyzed the problems of differentiating between the two ichnospecies.
4.7.2. Planolites isp. (Figure (B))
Simple, straight to gently curved, unbranched burrow, 3–10 mm in diameter and up to 14 cm in visible length. The burrow has pinstripe in wall surface and overlaps each other in some specimen, which is distributed parallel to bedding plane and preserved as convex epirelief in the Upper Cretaceous Majiacun and Sigou formations.
Figure 8. Occurrence characteristics of the trace fossils in the Upper Cretaceous of Xixia Basin. (A, B) Cylindricum isp., (C) Arenicolites isp.

4.8. Ichnogenus Cylindricum Linck, Citation1949
4.8.1. Cylindricum isp. (Figure (A) and (B))
Vertical, unbranched, unwalled burrows, preserved as endochnia in mudstones. Diameters range from 5 to 12 mm, and the length is 10–18 cm. Burrow filling is massive mudstones and having identical or different infill material to the host rock. The burrow is discovered in the Upper Cretaceous Majiacun and Sigou formations.
Relatively large vertical shafts such as the forms from the Xixia Basin were described as Cylindricum (Linck, Citation1949). The ichnogenus was partly included in Skolithos by Alpert (Citation1974). However, much greater width:length ratio of Cylindricum is distinct from Skolithos, such differences in morphology are significant to maintain Cylindricum as a valid ichnotaxon (Hasiotis, Citation2008).
4.9. Ichnogenus Arenicolites Salter, Citation1857
4.9.1. Arenicolites isp. (Figure (C))
This specimen is a simple ‘U’-shaped tube that is vertical or steeply inclined to the bedding plane, without spreite structure between two limb tubes. The diameter of burrow tube is 4 mm, width between two limb tubes is 35 mm and the length (or vertical depth) is 4.1 cm. Burrow filling material is identical to the overlying sediments. The right tube is overcrossed by other trace fossil. The burrow preserved as full relief in the Upper Cretaceous Sigou Formation.
Arenicolites consists of vertical U-tubes without spreite (Salter, Citation1857), occasionally differentiated as vertical to slightly oblique-paired burrows (Gillette et al., Citation2003), which is distinguished from the morphologically similar Diplocraterion by the absence of a spreite (Bromley, Citation1996).
Figure 9. Occurrence characteristics of the trace fossils in the Upper Cretaceous of Xixia Basin. (A) Archaeonassa isp. with abundant P. tubularis. (B) Rhizoliths.

4.10. Ichnogenus Archaeonassa Fenton and Fenton, Citation1937
4.10.1. Archaeonassa isp. (Figure (A))
Simple, straight, bilateral symmetry rige structures with marginal smooth and a wide central groove. Width of the rige and groove are 5 and 6 mm, respectively, and maximum length recorded is 56 mm. Scratch marks absent. Preserved as positive hyporelief in the Upper Cretaceous Sigou Formation.
The ichnotaxonomic is still uncertain (Buckman, Citation1994; Yochelson & Fedonkin, Citation1997). Only A. fossulata (Fenton & Fenton, Citation1937) is known. In continental environments, this ichnotaxa may be produced by annelids or mollusks (Buatois & Mángano, Citation2002).
4.11. Rhizoliths (Figure (B))
The root traces observed here, widely occur in the Upper Cretaceous Gaogou Formation, are 2–18 cm long (or deep) and 2–19 mm wide, and are oriented vertically or slightly inclined to bedding. They are filled with fine-grained oxidised sediments, and are preserved in silty mudstones that is explained to produce in floodplain environment.
5. Trace fossils assemblages and their sedimentary environments
The fluvial depositional system is well developed in the Upper Cretaceous of Xixia Basin, which includes channel subfacies (containing channel lag, point bar or heart bar microfacies), embankment subfacies (composed of natural levee and crevasse splay microfacies), floodplain subfacies (made up of alluvial flat and back-swamp microfacies) and fluvio-lacustrine subfacies. We recorded 10 ichnogenera and 22 ichnospecies in the Upper Cretaceous terrestrial deposits of the Xixia Basin including S. verticalis, S. linearis, S. magnus, Arenicolites isp., G. xixiaensis, B. antarcticus, B. capronus, B. coronus, P. megas, P. tubularis, P. heberti, P. sulcatus, P. cf. sulcatus, P. striatus, T. irregularis, T. serpentinum, T. barretti, S. gracilis, P. beverleyensis, P. isp., Cylindricum isp., Archaeonassa isp. and Rhizoliths. Based on their distribution, occurrences and sedimentary environments, four ichnoassemblage are recognised.
5.1. Skolithos-Gastruichnus ichnoassemblage
This ichnoassemblage consists of trace fossils with higher abundance and diversity, common ichnospecies are S. linearis, S. magnus, G. xixiaensis, Arenicolites isp., P. tubularis, P. megas, B. capronus and B. coronus, which mainly developed in the medium- to thick-bedded fine-grained sandstones. Among these trace fossils, S. linearis, Skolithos isp., G. xixiaensis, Arenicolites isp. and B. coronus are dwelling (associated with feeding activity) burrows, arranged almost perpendicular to the bedding plane, revealing the medium energy water current. The ichnogenus Palaeophycus is a kind of large feeding burrows distributed along the bedding planes of fine-grained sandstones with parallel bedding, which indicates the water body is stable and beneficial for the trace maker’s activity. This ichnoassemblage is commonly generated in the channel environment (Figure ).
Figure 10. A palaeogeographical model of the sedimentary and ichnological characteristics of fluvial deposits in the Upper Cretaceous of Xixia Basin, Southwest Henan Province.
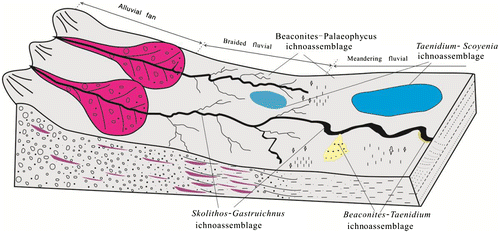
Comparable assemblages were described by Kim, Kim, and Pickerill (Citation2002), who recorded an ichnofauna from the Cretaceous Hasandong and Jinju formations of the southeast Korea. The ichnocoenose dominated by S. magnus, P. montanus and Chondrites isp., containing a small number of S. verticalis, T. barrette. Compare with the ichnofossil assemblage of ephemeral channels, the arthropod trackways and vertebrate footprints were not found in this assemblage. Such as, Melchor et al. (Citation2006) documented that ephemeral channels are dominated by S. linearis and rare Diplichnites isp., P. tubularis, T. barretti and vertebrate footprints (Dicynodontipus isp.), which are likely produced during the abandonment phase. Another relatively similar assemblage was described by Demathieu and Wycisk (Citation1990), Keighley and Pickerill (Citation2003), Schlirf, Uchman, and Kümmel (Citation2001). The ichnofaunas described are from the stratigraphic record that may be comparison with the channel top deposits (emergent sandy bars), the ichnocoenosis composed mostly of tetrapod footprints and very few specimens of P. tubularis and Helminthopsis hieroglyphica (Krapovickas, Ciccioli, Mángano, Marsicano, & Limarino, Citation2009). These trace fossil assemblages are comparable with Scoyenia ichnofacies (Buatois & Mángano, Citation1995, Citation1998; Frey & Pemberton, Citation1987; Frey et al., Citation1984).
Other ichnofaunas recorded from active fluvial channels are remarkably different, which documented low diversity and often monospecific traces, such as Skolithos (Buatois & Mángano, Citation2007; Buatois, Mangano, Genise, & Taylor, Citation1998; Minter, Krainer, Lucas, Braddy, & Hunt, Citation2007), Beaconites (Keighley & Pickerill, Citation2003). These trace fossil assemblages are defined Skolithos ichnofacies, which is characteristic of high-energy shifting sand substrates (Buatois & Mángano, Citation2004).
5.2. Beaconites-Taenidium ichnoassemblage
The ichnoassemblage is marked by B. coronus, B. antarcticus, T. irregularis and P. tubularis. These trace fossils, with high abundance and low diversity, are mostly burrows with meniscus backfill structures and are preserved in the purplish thin muddy siltstones and off-white fine-grained sandstones intercalated with reddish mudstones, and also, associated with cross-bedding, parallel bedding and wavy bedding. Based on these deposit characteristics, this ichnoassemblage is explained to occur in natural levee and crevasse splay sedimentary environments (Figure ).
In composition, this ichnoassemblage can be compared with the Cenozoic Toro Negro Formation of Argentina, which includes abundant meniscate trace fossils and dwelling tubes (e.g. Taenidium, Scoyenia and Palaeophycus) (Krapovickas et al., Citation2009). Likewise, this ichnoassemblage is similar to the assemblages from the fluvial deposits of the Middle Triassic Youfangzhuang Formation in western Henan of China (Wang, Qi, Li, Dai, & Chang, Citation2014).
5.3. Beaconites-Palaeophycus ichnoassemblage
This ichnoassemblage is mainly composed of P. tubularis, P. heberti, P. sulcatus, P. cf. sulcatus, P. striatus, B. coronus, B. antarcticus, S. gracilis, T. barretti, Planolites isp., Cylindricum isp., G. xixiaensis, S. verticalis, Archaeonassa isp. and Rhizoliths. These trace fossils with high abundance and diversity occur in the fine-grain sandstones and siltstones, red-violet silty mudstones and mudstones, locally containing calcareous concretions and desiccation cracks reflected a semi-arid climatic environment. The sediment-filled Rhizoliths associated with this assemblage indicate the floodplain and extremely shallow water conditions. Above lithologic characteristics display that the assemblage is compatible with periods of stability in hydrologic surface, less stressed depositional environment (Figure ).
There are several floodplain ichnoassemblages described from the stratigraphic record that favour comparison with the one analyzed in this paper. Such as, Morrissey, Hillier, and Marriott (Citation2012) described the assemblage in floodplain deposits from the Late Silurian and Early Devonian of the Anglo-Welsh Basin, UK; Buatois and Mángano (Citation2002) reviewed the ichnofaunas of the Carboniferous desiccated floodplain in western Argentina; Wang et al. (Citation2014) documented the assemblage in floodplain deposits from the Middle Triassic Youfangzhuang Formation in western Henan of China; Hu and Wu (Citation1993) discussed Scoyenia-Steinichnus-Rusophycus assemblage of fluvial deposits from the Upper Cretaceous Jiaguan Formation in Sichuan Basin of China.
5.4. Taenidium-Scoyenia ichnoassemblage
The main trace fossils in this ichnoassemblage are the feeding traces like T. serpentinum, T. irregularis, B. coronus, Planolites isp., P. tubularis, P. sulcatus, Palaeophycus cf. sulcatus and S. gracilis. Generally, the trace fossils with high abundance and middle diversity are arranged along or inclined at high angle to the bedding plane. This ichnoassemblage is mainly developed in the thin bedded siltstones, silty mudstones or muddy siltstones, and always associated with horizontal bedding and gentle current bedding, which reflects low energy conditions of water. The desiccation cracks indicate subaerial exposure during sedimentation. So, this ichnoassemblage is commonly explained to appear in the periodically exposed fluvio-lacustrine (pond or extremely shallow lacustrine) sedimentary environment (Figure ).
Compared to other ichnofaunas recorded in ephemeral water bodies deposits, a relatively diverse assemblage was described by Abdelkbir, Sebastian, Hafid, Jörg, and Driss (Citation2012) from the Permian Ikakern Formation of Morocco, and reviewed by Minter et al. (Citation2007) from the overfilled overbank deposits in Castle Peak of Texas, USA. The traces fossil assemblages include abundant arthropod grazing traces and tetrapod footprints.
A remarkably different ichnofauna from the Carboniferous Tupe Formation of western Argentina was described by Buatois and Mángano (Citation2002). As well as, Morrissey et al. (Citation2012) documented ephemeral lakes deposits from the Late Silurian and Early Devonian of the Anglo-Welsh Basin, UK. The ichnofauna is characterised by low diversity, dominance of very simple forms, superficial or very shallow trace fossils as combination of locomotion, grazing, and dwelling structures.
As mentioned above, the lithofacies and ichnoassemblages in the Upper Cretaceous Gaogou, Majiachun and Sigou formations in Xixia Basin of Western Henan province indicate that they were generated in channel, levee and crevasse splay, floodplain and fluvio-lacustrine depositional environments (Figure ).
All of ichnoassemblages described above could be classified under the Scoyenia ichnofacies, which is generally characterised (cf. Buatois & Mángano, Citation1998, Citation2002, Citation2004) by: (1) meniscate backfilled burrows (Beaconites, Taenidium, Scoyenia), (2) simple horizontal traces (Planolites, Palaeophycus), (3) trackways and trails (Archaeonassa), (4) vertical dwelling burrows (Skolithos, Cylindricum, Arenicolites, Gastruichnus), (5) calcareous concretions and desiccation cracks, (6) a mixture of invertebrate and plant traces, (7) low to moderate ichnodiversity, and localised high abundance.
6. Conclusion
| (1) | Based on lithology, physical sedimentary structures, sedimentary sequences, bed boundaries and trace fossil contents, this paper studied in detail the paleodepositioal setting of the Cretaceous succession of Xixia Basin, Henan Province and proposed a palaeoenvironment model, i.e. from north to the south along the Basin, in turn, alluvial fan system, braided fluvial system, meandering fluvial system and fluvio-lacustrine system are distributed spatially. | ||||
| (2) | Trace fossils from the study area can be assigned to Arenicolites isp., Archaeonassa isp., B. antarcticus, B. capronus, B. coronus, Cylindricum isp., G. xixiaensis, P. megas, P. tubularis, P. heberti, P. sulcatus, P. cf. sulcatus, P. striatus, P. beverleyensis, P. isp., S. gracilis, S. verticalis, S. linearis, S. isp., T. irregularis, T. serpentinum, T. barretti and Rhizoliths. | ||||
| (3) | Based on the composition, occurrence and distribution of trace fossils in the subfacies of fluvial and extremely shallow lacustrine facies, four types of ichnoassemblages are recognised, including the Skolithos-Gastruichnus, Beaconites-Taenidium, Beaconites-Palaeophycus and Taenidium-Scoyenia ichnoassemblages that are recorded in the Upper Cretaceous in Xixia Basin of southwestern Henan Province, China. | ||||
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Abdelkbir, H., Sebastian, V., Hafid, S., Jörg, W. S., & Driss, H. (2012). On a moderately diverse continental ichnofauna from the Permian Ikakern Formation (Argana Basin, Western High Atlas, Morocco). Journal of African Earth Sciences, 68, 15–23.10.1016/j.jafrearsci.2012.03.011
- Alpert, S. P. (1974). Systematic review of the genus Skolithos. Journal of Paleontology, 48, 661–669.
- Alpert, S. P. (1975). Planolites and Skolithos from the Upper Precambrian–Lower Cambrian White–Inyo Mountains, California. Journal of Paleontology, 49, 508–521.
- Bradshaw, M. A. (1981). Paleoenvironmental interpretations and systematics of Devonian trace fossils from the Taylor Group (lower Beacon Supergroup), Antarctica. New Zealand Journal of Geology and Geophysics, 24, 615–652.10.1080/00288306.1981.10421537
- Bromley, R. G. (1996). Trace fossils: Biology, taphonomy and applications (2nd ed.). London: Chapman and Hall.10.1007/978-1-4899-2875-7
- Buatois, L. A., & Mángano, M. G. (1995). The paleoenvironmental and paleoecological significance of the lacustrine Mermia ichnofacies: An archetypical subaqueous nonmarine trace fossil assemblage. Ichnos, 4, 151–161.10.1080/10420949509380122
- Buatois, L. A., & Mángano, M. G. (1998). Trace fossil analysis of lacustrine facies and basins. Palaeogeography, Palaeoclimatology, Palaeoecology, 140, 367–382.10.1016/S0031-0182(98)00020-0
- Buatois, L. A., & Mángano, M. G. (2002). Trace fossils from Carboniferous floodplain deposits in western Argentina: Implications for ichnofacies models of continental environments. Palaeogeography, Palaeoclimatology, Palaeoecology, 183, 71–86.10.1016/S0031-0182(01)00459-X
- Buatois, L. A., & Mángano, M. G. (2004). Animal-substrate interactions in freshwater environments: Applications of ichnology in facies and sequence stratigraphic analysis of fluvio-lacustrine successions. In D. McIlroy (Ed.), The application of ichnology to palaeoenvironmental and stratigraphic analysis (pp. 311–333). London: The Geological Society.
- Buatois, L. A., & Mángano, M. G. (2007). Invertebrate ichnology of continental freshwater environments. In W. Miller III (Ed.), Trace fossils: Concepts, problems, prospects (pp. 285–323). Amsterdam: Elsevier.10.1016/B978-044452949-7/50143-1
- Buatois, L. A., Mangano, M. G., Genise, J. F., & Taylor, T. N. (1998). The ichnologic record of the continental invertebrate invasion: Evolutionary trends in environmental expansion, ecospace utilization, and behavioral complexity. Palaios, 13, 217–240.10.2307/3515447
- Buckman, J. O. (1994). Archaeonassa Fenton and Fenton 1937 reviewed. Ichnos, 3, 185–192.10.1080/10420949409386387
- Carmona, N. B., Buatois, L. A., Mángano, M. G., & Bromley, R. G. (2008). Ichnology of the Lower Miocene Chenque Formation, Patagonia, Argentina: Animal – substrate interactions and the Modern Evolutionary Fauna. Ameghiniana, 45, 1–32.
- Demathieu, G. R., & Wycisk, P. (1990). Tetrapod trackways from Southern Egypt and Northern Sudan. Journal of African Earth Sciences, 10, 435–443.10.1016/0899-5362(90)90096-W
- de Saporta, G. (1872). Paléontologie française ou description des fosiles de la France. 2 série Végétaux. Plantes Jurassiques (Vol. 1, 432 pp., 60 pls). Paris: G. Masson
- Fenton, C. L. & Fenton, M. A. (1937). Burrows and trails from Pennsylvanian rocks of Texas. American Midland Naturalist, 18, 1079–1084.
- Fillion, D., & Pickerill, R. K. (1990). Ichnology of the Upper Cambrian? To Lower Ordovician Bell Island and Waban groups of eastern Newfoundland, Canada. Palaeontographica Canadian, 7, 1–119.
- Frey, R. W., & Pemberton, S. G. (1987). The Psilonichnus ichnocoenose, and its relationship to adjacent marine and nonmarine ichnocoenoses along the Georgia coast. Bulletin of Canadian Petroleum Geology, 35, 333–357.
- Frey, R. W., Pemberton, S. G., & Fagerstrom, J. A. (1984). Morphological, ethological and environmental significance of the ichnogenera Scoyenia and Ancorichnus. Journal of Paleontology, 58, 511–528.
- Gao, Z. H., Li, R. Y., Wang, Q. J., Dong, C., Yan, D. F., & Sun, B. N. (2013). Trace fossil assemblages in the Miocene from northeast margin of Qaidam Basin and its sedimentary environment. Acta Sedimentologica Sinica, 31, 413–420. (in Chinese with English abstract).
- Gillette, L., Pemberton, S. G., & Sarjeant, W. A. S. (2003). A late Triassic invertebrate ichnofauna from Ghost Ranch, New Mexico. Ichnos, 10, 141–151.10.1080/10420940390255493
- Goldring, R., & Pollard, J. E. (1995). A re-evaluation of Ophiomorpha burrows in the Wealden Group (Lower Cretaceous) of southern England. Cretaceous Research, 16, 665–680.10.1006/cres.1995.1041
- Gregory, M. R., Campbell, K. A., Zuraida, R., & Martin, A. J. (2006). Plant traces resembling Skolithos. Ichnos, 13, 205–216.10.1080/10420940600843617
- Haldeman, S. S. (1840). Supplement to number one of “a monograph of the Limniades, or freshwater univalve shells of North America”, containing descriptions of apparently new animals in different classes, and the names and characters of the subgenera in Paludina and Anculosa. Philadelphia, PA: J. Dobson.
- Hall, J. (1847). Containing descriptions of the organic remains of the lower division of the New York System (equivalent of the lower Silurian Rocks of Europe). Palaeontology of New York (Vol. 1). Albany: C. van Benthuysen.
- Hall, J. (1852). Palaeontology of New York (Vol. 2). Albany: State of New York.
- Hasiotis, S. T. (2008). Reply to the Comments by Bromley et al. of the paper “Reconnaissance of the Upper Jurassic Morrison Formation ichnofossils, Rocky Mountain Region, USA: Paleoenvironmental, stratigraphic, and paleoclimatic significance of terrestrial and freshwater ichnocoenoses” by Stephen T. Hasiotis. Sedimentary Geology, 208, 61–68.10.1016/j.sedgeo.2008.04.004
- Heer, O. (1877). Flora fossilis Helvetiae. Die vorweltliche Flora der Schweiz [Flora fossilis Helvetiae. The vorweltliche flora of Switzerland]. Zürich: Verlag J. Wurster and Co.
- Howell, B. F. (1944). A new Skolithos from the Cambrian Hardyston Formation of Pennsylvania. Wagner Free Institute of Science of Philadelphia Bulletin, 19, 41–44.
- Hu, B. (2014). The Mesozoic ithofacies-paleogeography of Henan Province. Beijing: Coal industry Press (in Chinese).
- Hu, B., Wang, Y. Y., & Song, H. B. (2014). The ichnofacies and ichnoassemblages in terrestrial deposits of China. Journal of Palaeography, 3, 61–73.
- Hu, B., & Wu, X. T. (1993). Ichnocoenosis of alluvial Jiaguan Formation (Upper Cretaceous), Emei, Sichuan, China. Acta Palaeontologica Sinica, 32, 478–489 (in Chinese with English abstract).
- Hu B., Zhong M. Y., Song H. B. (2010). Zhaoying village: Beaconites, Scoyenia and dinosaur eggs ichnofabrics in fluvial and extremely shallow lacustrine deposits from the Upper Cretaceous Zhaoying village, Xixia town of Nanyang city. Progress in ichnofabric and ichnocoenosis – The field trip guidebook and abstracts of XIIW, Henan, Jiaozuo of Henan Province. Beijing: Coal industry Press.
- Keighley, D. G., & Pickerill, R. K. (1994). The ichnogenus Beaconites and its distinction from Ancorichnus and Taenidium. Palaeontology, 37, 305–337.
- Keighley, D. G., & Pickerill, R. K. (1995). The ichnotaxa Palaeophycus and Planolites: Historical perspectives and recommendations. Ichnos, 3, 301–309.10.1080/10420949509386400
- Keighleyn, G. D., & Pickerill, R. K. (1997). Systematic ichnology of the Mabou and Cumberland groups (Carboniferous) of western Cape Breton Island, eastern Canada, 1: Burrows, pits, trails, and coprolites. Atlantic Geology, 33, 181–215.
- Keighley, D. G., & Pickerill, R. K. (2003). Ichnocoenoses from the Carboniferous of eastern Canada and their implications for the recognition of ichnofacies in nonmarine strata. Atlantic Geology, 39, 1–22.
- Kim, J. Y., Kim, K.-S., & Pickerill, R. K. (2002). Cretaceous nonmarine trace fossils from the Hasandong and Jinju Formations of the Namhae Area, Kyongsangnamdo, Southeast Korea. Ichnos, 9, 41–60.10.1080/10420940216414
- Krapovickas, V., Ciccioli, P. L., Mángano, M. G., Marsicano, C. A., & Limarino, C. O. (2009). Paleobiology and paleoecology of an arid-semiarid Miocene South American ichnofauna in anastomosed fluvial deposits. Palaeogeography, Palaeoclimatology, Palaeoecology, 284, 129–152.
- Li, R. H. (1993). Trace fossils and ichnofacies of middle Ordovician Gongwusu Formation, Zhuozishan, Inner Mongolia. Acta Palaeontologica Sinica, 32, 88–108 (in Chinese with English abstract).
- Linck, O. (1949). Lebens-spuren aus dem Schilfsandstein (Mittl. Keuper km 2) NW-Wurttembergs und ihre Bedeutung fur die Bildungsgeschichte derr Stufe [Living tracks of the schilfsandstein (m. keuper km) northwest wurttembergs and their significance for the history of education to the level]. Verin Vateral. Naturkd. Wurttemberg, Jahresh, 97–101, 1–1000.
- Melchor, R. N., Bedatou, E., de Valais, S., & Genise, J. F. (2006). Lithofacies distribution of invertebrate and vertebrate trace-fossil assemblages in an Early Mesozoic ephemeral fluvio-lacustrine system from Argentina: Implications for the Scoyenia ichnofacies. Palaeogeography, Palaeoclimatology, Palaeoecology, 239, 253–285.10.1016/j.palaeo.2006.01.011
- Miller, S. A., & Dyer, C. B. (1878). Contributions to Paleontology (Vol. 2). Cincinnati, OH: Private Publication.
- Minter, N. J., Krainer, K., Lucas, S. G., Braddy, S. J., & Hunt, A. P. (2007). Palaeoecology of an Early Permian playa lake trace fossil assemblage from Castle Peak, Texas, USA. Palaeogeography, Palaeoclimatology, Palaeoecology, 246, 390–423.10.1016/j.palaeo.2006.10.009
- Morrissey, L. B., Hillier, R. D., & Marriott, S. B. (2012). Late Silurian and Early Devonian terrestrialisation: Ichnological insights from the Lower Old Red Sandstone of the Anglo-Welsh Basin, U.K. Palaeogeography, Palaeoclimatology, Palaeoecology, 337–338, 194–215.10.1016/j.palaeo.2012.04.018
- Pan, H. Z., Wang, D. Y., & Feng, J. C. (2007). Late Cretaceous non-marine gastropoda from the dinosaur egg-bearing strata of Xixia Basin, Henan. Acta Palaeontologica Sinica, 46, 293–298 (in Chinese with English abstract).
- Pan Z. C., Xu L., Xi Y. H. (2008). Stratigraphic paleontology research of Henan Province: The fifth volume of Mesozoic Era. Zhengzhou: Yellow River Water Conservancy Press (in Chinese).
- Pemberton, S. G., & Frey, R. W. (1982). Trace fossil nomenclature and the Planolites-Palaeophycus dilemma. Journal of Paleontology, 56, 843–881.
- Rodríguez-Tovar, F. J., & Uchman, A. (2004). Trace fossils after the KT boundary event from the Agost section, SE Spain. Geological Magazine, 141, 429–440.10.1017/S0016756804009410
- Salter, J. W. (1857). On annelide-burrows and surface markings from the Cambrian rocks of the Longmynd. Quarterly Journal of the Geological Society of London, 13, 199–206.10.1144/GSL.JGS.1857.013.01-02.29
- Schlirf, M. (2000). Upper Jurassic trace fossils from the Boulonnais (Northern France). Geologica et Palaeontologica, 34, 145–213.
- Schlirf, M. & Uchman, A. (2005). Revision of the ichnogenus Sabellarifex Richter, 1921 and its relationship to Skolithos Haldeman, 1840 and Polykladichnus Fursich, 1981. Journal of Systematic Palaeontology, 3, 115–131.10.1017/S1477201905001550
- Schlirf, M., Uchman, A., & Kümmel, M. (2001). Upper Triassic (Keuper) non-marine trace fossils from the Haßberge area (Franconia, south-eastern Germany). Paläontologische Zeitschrift, 75, 71–96.10.1007/BF03022599
- Song, H. B., Hu, B., Wang, D. Y., Liu, S. X., Gao, Y., & Zhong, M. Y. (2008). Sedimentary characteristics and environments of the Gaogou Formation of Upper Cretaceous in Xixia Basin, Henan. Petroleum Geology and Engineering, 22(4), 1–5 (in Chinese with English abstract).
- The Henan Geology and Mineral Bureau. (1989). Regional geology of Henan Province. Beijing: Geology Press (in Chinese).
- Vialov, O. S. (1962). Problematica of the Beacon Sandstone at Beacon Height West, Antarctica. New Zealand Journal of Geology and Geophysics, 5, 718–732.10.1080/00288306.1962.10417633
- Wang, D. Y., Chen, P. J., Chen, J. H., Cao, M. Z., Pan, H. Z., & Li, G. (2006). Discovery of invertebrate fossils from the dinosaurian egg-bearing strata in the Xixia Basin of Henan. China. Acta Palaeontologica Sinica, 45, 494–497 (in Chinese with English abstract).
- Wang, D. Y., Feng, J. C., Zhu, S. C., Wu, M., Fu, G. H., He, P., … Wang B. X. (2008). Dinosaur eggs and skeletons from Henan province in China. Beijing: Geological Publishing House (in Chinese with English abstract).
- Wang, D. Y., Luo, M. J., Zhou, S. Q., Zhang, K. W., Li, J. F., & He, P. (1996). Dinosaur egg fossils and paleoecologic characterics in Henan Province. In Progress in geology of china (1993–1996). Papers to 30 th IGC. Beijing: China Ocean Press.
- Wang, M., Qi, Y. A., Li, D., Dai, M. Y., & Chang, Y. G. (2014). Ichnofabrics and their environmental interpretation from the fluvial deposits of the Middle Triassic Youfangzhuang Formation in Western Henan, Central China. Journal of Earth Science, 25, 648–661.10.1007/s12583-014-0454-2
- White, C. D. (1929). Flora of the Hermite Shale, Grand Canyon, Arizona. Washington: Publications of the Carnegie Institution.
- Xu, X., Zhao, X. J., Lü, J. C., Huang, W. B., Li, Z. Y., & Dong, Z. M. (2000). A new iguanodontian from sangping Formation of neixiang, Henan and its stratigraphical implication. Vertebrata Palasiatica, 38, 176–191 (in Chinese with English abstract).
- Yochelson, E. L., & Fedonkin, M. A. (1997). The type specimens (Middle Cambrian) of the trace fossil Archaeonassa Fenton and Fenton. Canadian Journal of Earth Sciences, 34, 1210–1219.10.1139/e17-097
- Zhang, G. C. (2002). Evolution of the Late Cretaceous depositional systems in the Xixia Basin, Henan province. Journal of Jiaozuo Institute of Technology (Natural Science), 21, 85–90 (in Chinese with English abstract).
- Zhang, G. C., Guo, W. X., & Zeng, Y. F. (2004). Ichnofabrics from the upper Cretaceous fluvial and lacustrine deposits in Xixia Basin. Henan. Journal of Palaeogeography, 6, 434–441 (in Chinese with English abstract).
- Zhou, S. Q., Feng, Z. J., & Hui Y. X. (2001). A study of dinosaurian fossil eggs from the Xixia Basin Henan. Jiangxi Geology, 15, 96–101 (in Chinese with English abstract).
- Zhu, G. Y. (1997). Stratigraphy and sedimentary facies of red beds in Xixia Basin of Henan province. Journal of the University of Petroleum, China, 21, 110–114 (in Chinese with English abstract).