
Abstract
On Gökçeada Island, the Mezardere Formation is dominated by the prodelta facies composed mostly of light grey, thinly bedded, fissile calcareous mudstones and siltstones, which mostly display parallel lamination or rarely ripple lamination. These sediments are intercalated with isolated thin beds of very fine- or fine-grained slightly muscovitic sandstones, which display parallel lamination in the lower part and ripple lamination in the upper part, or only ripple lamination. Thicker sandstone beds, referred to as mouth bars, are present in places. The prodelta sediments are poorly bioturbated (ii = 0–1), but moderately diverse trace fossils occur on some bedding surfaces, including 22 ichnotaxa referred to 13 ichnogenera and three trace fossils left in open nomenclature. The impoverished trace fossil assemblages are typical of deltaic sediments, but paucity of the trace fossils and poor bioturbation of the prodelta sediments in the Mezardere Formation is striking. The typically marine ichnoataxa are rare or absent in most outcrops. This trace fossil assemblage, resembling the Mermia ichnofacies, suggests a strong brackishness of the palaeoenvironment; however, the occurrence of Bichordites kuzunensis in one outcrop suggests local and temporal normal marine conditions referred to as salinity fluctuations.
1. Introduction
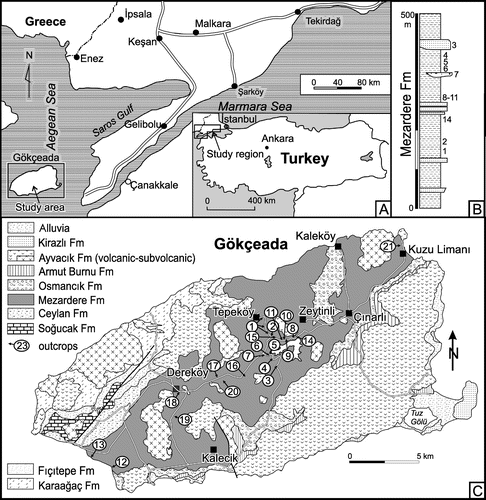
Deltaic sediments are usually characterised by more or less impoverished assemblages of shallow marine trace fossils and an overall low degree of bioturbation. These features are related to variable physicochemical stress factors influenced by high sedimentation rate, salinity, temperature and water turbidity fluctuations in deltaic environments (Gingras, MacEachern, & Pemberton, Citation1998; MacEachern, Bann, Bhattacharya, & Howell, Citation2005; Tonkin, Citation2012). Such ichnological features were expected in the Palaeogene Mezardere Formation, which on Gökçeada Island is dominated by prodelta mudstones and siltstones (Siyako & Huvaz, Citation2007; Temel & Çiftçi, Citation2002). However, it appears that the trace fossil association from this unit displays partial but distinct similarities to freshwater trace fossil assemblages, with only a few typically marine forms. Therefore, the main aim of this paper is to describe and interpret trace fossils from the prodelta deposits of the Mezardere Formation in the Gökçeada Island (Figure (A)–(C)) in the context of ichnology of the deltaic sediments.
Figure 1. Location maps. (A) General location of the study region and the study area. (B) Generalised section of the Mezardere Formation in the middle part of the island with approximate stratigraphic location of the outcrop partial sections (for reference numbers see Table ). (C) Geological map of the Gökçeada Island (modified from Temel & Çiftçi, Citation2002) showing locations of studied outcrops (for reference numbers see Table ).
Trace fossils of the Mezardere Formation are poorly known. Only Demircan and Uchman (Citation2008, Citation2010) described a new trace fossil Osculichnus labialis referred to hunting fishes, and mentioned about Planolites and the bivalve resting trace Lockeia siliquaria form the Mezardere Formation NW of İstanbul. They also described other new trace fossil Bichordites kuzunensis from Kuzulimanı in the Gökçeada Island (Demircan & Uchman Citation2012). Ichnology of the Mezardere Formation was also briefly presented during the 12th International Ichnofabric Workshop (Demircan & Uchman Citation2013a, Citation2013b).
2. Geological setting
The Mezardere Formation is a part of the up to 9000 m thick filling of the Thrace Basin (Eocene–Miocene). On the Gökçeada Island, the Thrace Basin filling is 2000 m thick and is composed of deep-sea to shallow-marine and non-marine, mostly clastic sediments (Figure ) which are cut by Late Oligocene and Late Miocene volcanic intrusions (Turgut & Eseller, Citation1999; Turgut, Siyako, & Dilki, Citation1983). The Yenimuhacır Group (Upper Eocene–Oligocene) composed mostly of deltaic sediments, is a part of the basin fill. It is subdivided into the Mezardere, Osmancık and Danişment formations (Siyako, Citation2006; Siyako & Huvaz, Citation2007). The first two formations are distinguished in the Gökçeada Island (Figure (B) and (C)).
Figure 2. Stratigraphic column of the units on Gökçeada Island (modified from Temel & Çiftçi, Citation2002, foremost in respect to the lithology of the Mezardere Formation).
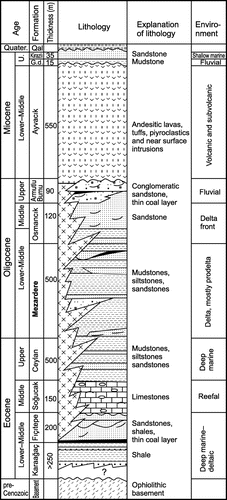
The Mezardere Formation (Gökçen, 1967; Saner, Citation1985; Sümengen et al., 1987), named also the Mezardere Shale by Ünal (Citation1967) or the Muhacır Formation by Lebküchner (Citation1974), is the lowest unit of Yenimuhacır Group. The Early Oligocene has been determined by dinocysts (Distatodinum craterum, Distatodinum ellipticum) and palynomorphs (Echites bifurcusnatospori, Baculoatisporites, Cingulatisporites macrospecious, Subtriporopollenites simplex (Temel & Çiftçi, Citation2002). The unit is composed mostly of mudstone, and siltstone, shales, siltstones and thin- or medium-bedded, poorly sorted sandstones (Figures (B), ). It is an important gas source-rock (Hoşgörmez & Yalçın, Citation2005; Karahanoğlu, Eder, & İlleez, Citation1995). The Mezardere Formation overlies sand and shale flysch of the Keşan Formation (Middle–Upper Eocene) or shale-sandy flysch with tuffites of the Ceylan Formation (Upper Eocene). The Mezardere Formation is overlain by delta front and delta plain sediments with coal, limestones and pyroclastics of the Osmancık Formation (Siyako, Citation2006).
Figure 3. Outcrop photographs illustrating facies of the Mezardere Formation. For outcrop numbers see Table . (A). Slightly metamorphosed mudstone shales intercalated with rippled, very fine-grained sandstones and siltstones of the prodelta facies, outcrop 8. (B) Prodelta facies dominated by mudstones; outcrop 19, about 20 m high. (C). Mudstones intercalated with fine-grained sandstones, prodelta facies; adjacent to outcrop 1. (D) Thickening- and coarsening-up package of sandstones intercalated with mudstones, which overlies mudstones; distal mouth bar facies overlying prodelta facies; outcrop 1; 1 m long measurement stick for scale in the red ellipse. (E) Isolated packages of thin-bedded sandstones in prodelta mudstones, probably very distal mouth bars; outcrop 20; the package of sandstone beds in the middle is about a metre thick. (F) Cross-bedded lenticular lithosoms of sandstones; proximal mouth bar facies; outcrop 16, about 20 m high.

On Gökçeada Island, the Mezardere Formation is 400–900 m thick (Kesgin & Varol, Citation2003; Temel & Çiftçi, Citation2002), and composed mostly of thinly bedded light grey calcareous mudstones and siltstones, which often display shale fissility, fine parallel lamination or, more rarely, ripple lamination (Figure (A) and (C)). Locally, carbonate concretions are present. Isolated thin beds, or packages of thin beds, of very-fine grained or fine-grained slightly muscovitic sandstones are locally present (Figure (E)). These beds display parallel lamination in the lower part and ripple lamination in the upper part, or only ripple lamination. Laterally discontinuous 3–12 m thick packages of thin- to thick-bedded, cross-laminated, fine- to medium-grained sandstones are also present in places. Some of them show clear thickening- and coarsening-up trends (Figure (D)). Very rarely thicker bodies of cross-laminated sandstones crop out (Figure (F)). Fine carbonised plant detritus dispersed in the sandstones is common. Occasionally, beds of coarse to very coarse sandstones and fine-grained conglomerates are present (see Figure (C)), outcrop 3). Some thicker sandstone beds show ball-and-pillow structures (see Figure (C)); outcrop 3, Table ).
Table 1. The studied outcrops of the Mezardere Formation in the Gökçeada Island.
Generally, the Mezardere Formation is interpreted as deltaic deposits, from delta front to prodelta (Temel & Çiftçi, Citation2002) or as prodelta sediments (Siyako & Huvaz, Citation2007). On Gökçeada Island, we interpret the Mezardere Formation to be dominated by the prodelta facies. Their depth of deposition was estimated to 50–80 m from the basin modelling (Hoşgörmez & Yalçın, Citation2005).
In this paper, the packages of coarsening and thickening upwards sandstones interbedded with siltstones and mudstones (Figure (D)) are interpreted as distal mouth bars, and the lenticular bodies composed of thick, thickening up, cross-bedded sandstones (Figure (F)) are considered proximal mouth bars. Locally, packages of lenticular sandstone beds suggest filling of small channels, which may have been formed on the slope of the delta front. The Mezardere Formation in the Gökçeada Island is likely composed of several superimposed deltaic cycles, of which upper parts are eroded in the majority. Therefore, the formation is dominated by the prodelta sediments typical of the lower part of the deltaic cycles.
3. Trace fossils
Trace fossils have been examined in 23 outcrops (Figure (C); Table ), which belong to all facies described above. Representative specimens are housed in the Natural History Museum of MTA in Ankara (institutional abbreviation TRGM), and single specimens in the Institute of Geological Sciences in the Jagiellonian University (institutional abbreviation INGUJ224P).
Archaeonassa cf. fossulata Fenton & Fenton, Citation1937a
Figure (A) and (B)
Figure 4. Photographs of beddings planes illustrating trace fossils in the Mezardere Formation. (A) Archaeonassa cf. fossulata, outcrop 3, specimen TRGM3/01. (B) Archaeonassa cf. fossulata, small form, outcrop 4, specimen TRGM1/01. (C) Archaeonassa isp. A, outcrop 4, specimen TRGM1/02. (D) Archaeonassa isp. B showing oblique marks (om); outcrop 4, specimen TRGM1/03. (E) ?Arenicolites isp., presumable pairs of structures related to limbs are in yellow ellipses; outcrop 21, specimen TRGM8/01. (F) ?Arenicolites isp., outcrop 3, specimen TRGM1/04. (G) Bichordites kuzunensis, outcrop 21, specimen INGUJ224P7. (H) ?Copeza isp., outcrop 7, specimen TRGM7/01. (I) G. indianensis, a specimen with short protrusions (p); outcrop 7, specimen TRGM6/01.

Archaeonassa cf. fossulata is an epichnial semi-circular looping furrow, 2–3 mm wide, excluding levees. The external width of this trace fossil is 5 mm, including levees which are up to 1.5 mm wide. Their external margin is uneven. The loops are 50–60 mm wide (Figure (A)). In a smaller, non-looping form, the trace fossil is only 2 mm wide (Figure (B)). In the type material from the middle Cambrian of British Columbia. Archaeonassa fossulata is not looping and its levees are even (Yochelson & Fedonkin, Citation1997). Archaeonassa is interpreted as a crawling trail produced mostly by gastropods (Buckman, Citation1994; Fenton & Fenton, Citation1937a; Stanley & Feldmann, Citation1998) or crustaceans (Mángano & Buatois, Citation2003; Yochelson & Fedonkin, Citation1997); however, flatworms and cerianthid anemones may also produce traces of this type, i.e. furrows bounded by levees (Collins, Lipps, & Valentine, Citation2000).
Archaeonassa isp. A and B
Figures (C) and (D), (E) and (F)
The variant A (Figures (C), (E) and (F)) is an epichnial, straight or curved, shallow V-shaped furrow with even levees, 3–5 mm wide, 20–30 mm long. Its surface is smooth or covered with perpendicular, irregular wrinkles (Figure (F)). The trace becomes gradually shallower and narrower at terminations. It co-occurs with crustacean trackways and resting traces of similar size, and probably is a preservational variant.
The variant B (Figure (D)) is epichnial, winding semi-circular groove, 2 mm wide with levees, which are 1 mm wide. When the furrow became shallower, the levees pass into a row of oblique marks, 1 mm wide, about 0.3 mm wide and about 0.3 mm apart. These marks are interpreted as traces of body appendages, probably arthropods. Melchor, Bellosi, and Genise (Citation2003) observed V-shaped Archaeonassa in Triassic lacustrine delta sediments and referred it to gastropods on the basis of neoichnological observations of Buccinanopsis in a tidal flat. They concluded that the cross-sectional profile of Archaeonassa depends on the substrate and depth of burrowing, with V-shaped pattern in subsurface burrows under a centimetre of sand.
?Arenicolites isp.
Figure (E) and (F)
An epichnial, oval depression or elevation, 1.5–2 mm in diameter that is (1) located in the centre of a shallow, flat-bottom depression or elevation, 3–14 mm in diameter; the depression/elevation is surrounded by an indistinct rim, which is 2 mm wide, or (2) without the depression or elevation, but surrounding irregular, convex up, low collar feature that is up to 2 mm wide. At least part of these structures seems to occur in pairs located 6–20 mm apart. These structures are interpreted to U-shaped vertical burrows typical of Arenicolites Salter. Arenicolites is regarded as a dwelling and feeding burrow of small crustaceans (Goldring, Citation1962) or suspension feeding annelids (Hakes, Citation1976), which occurs in different, but typically, shallow-marine environments (Crimes, Citation1977).
B. kuzunensis Demircan & Uchman, Citation2012
Figure (G)
This trace fossil an epichnial, gently convex, horizontal to sub-horizontal, winding ribbon-like structure, in a fine-grained sandstone bed, which is preserved in full relief. It is composed of a 1 mm thick, smooth central tunnel, which is enveloped symmetrically by a ribbed, slightly bilobate covering zone. Locally, the tunnel is exposed where the covering zone is removed by weathering. The covering zone is 4.5–10 mm wide, and of the constant width in each specimen. At the top, it displays tightly packed gently arcuate ribs, which are about 1 mm wide. Their concavity is unidirectional, both endings of the ribs narrow and are gently arcuate. The external part of the covering zone displays a thin mantle, which surface is indistinctly granulated. Crossings and overlaps between the burrows are present. This trace fossil is interpreted as the burrows of small echinoids (Demircan & Uchman, Citation2012).
?Copeza isp.
Figure (H)
This is an incomplete trackway (fragment of one flank) composed of two rows of long, and a row of short, narrow ridges. The longer ridges, 3–7 mm long, about 0.5 mm wide, 20–27 mm apart in each row, are obliquely arranged to the trackway course under an angle ca. 20° and ca. 50° in the external and the middle rows, respectively. The adjacent longer ridges from two rows touch each other tangentially or are up to 5 mm apart. The short ridges are up to 2 mm long, consequently obliquely arranged in the third row, 24–28 mm apart, 7–8 mm from the longer ridges of the middle row.
This trace fossil is most similar to Copeza triremis Hitchcock, Citation1858 from the Lower Jurassic continental deposits of Massachusetts (Rainforth, Citation2005). They were interpreted by Hitchcock (Citation1858) as the trackways of arthropods. Copeza is also noted in the Middle Jurassic supratidal sabkha deposits (Kilbourne, Curran, & Wilson, Citation1998).
Gordia indianensis (Miller, Citation1889)
Figures (I), (B)
Figure 5. Trace fossils of the Mezardere Formation. (A) ?Hamipes isp., outcrop 6, specimen TRGM7/02. (B) G. indianensis, outcrop 5, specimen TRGM5/01. (C) cf. Halimedides isp. (Hd) and Ptychoplasma isp. (Pt), outcrop 4, specimen TRGM1/05. (D) Helminthoidichnites isp., outcrop 5, specimen TRGM5/02. (E) Helminthoidichnites isp. on a rippled surface; outcrop 6, specimen TRGM1/06. (F) Helminthoidichnites isp., outcrop 5, specimen TRGM1/07. (G) Helminthoidichnites cf. ornatus, outcrop 8, specimen TRGM8/02. (H) Helminthoidichnites cf. ornatus, outcrop 6, specimen TRGM8/03.

This trace fossil (Gordia isp. in Demircan & Uchman, 2013) is a hypichnial, epichnial or endichnial, horizontal looping ridge or groove, 0.5–1.2 mm wide. The majority of the loops are 10–15 mm wide. The looping course of the trace is irregularly winding with common rapid changes of direction. In places, short, irregular protrusions of the kinks in the prolongation of a segment can be present (Figure (I)). Less looping forms, with a transition to Helminthoidichnites isp., are common. In dense occurrences, it is hard to distinguish between these two ichnotaxa.
Buatois, Mángano, Maples, and Lanier (Citation1997, Citation1998) made the transfer of Haplotichnus indianensis Miller to Gordia Emmons, but spelled as G. indianensis (Miller). Nevertheless, the original ichnospecies name is indianensis (Miller, Citation1889, p. 578). We follow the original spelling. Rindsberg and Kopaska-Merkel (Citation2005) argued that Haplotichnus should be conserved because (1) it is a surface trail or a shallow burrow while Gordia is a burrow only, and (2) the sharp turns in Haplotichnus are typical of short-bodied organisms, presumably arthropods, while gentle curves in Gordia can be referred to long-bodied ‘worms’. We find the first argument as very interpretative, vague (in some case both Gordia and Haplotichnus can be shallow burrows) and impractical in determination and the second as influenced by the presumable tracemakers, which cannot be accepted as the ichnotaxobase (Bertling et al., Citation2006). Therefore, we consider the array of loops and the tubular shape as the principle diagnostic features that join Haplotichnus and Gordia, with priority for the latter. G. indianensis (Miller) is known from lacustrine and marginal marine deposits of the Carboniferous (Buatois et al., Citation1997, Citation1998; Mángano & Buatois, Citation2004) and lacustrine Permian (Minter, Krainer, Lucas, Braddy, & Hunt, Citation2007) and Triassic (Melchor et al., Citation2003).
cf. Halimedides isp.
Figure (C)
The trace fossil cf. Halimedides is a hypichnial chain of oval mounds, 1–2 mm wide, joined by a thin string, about 0.5 mm wide. The mounds are 1.5–2 mm apart and elongated parallel to or perpendicular to the course of the chain. The trace fossil is very fragmentary preserved; therefore its determination is tentative. Taxonomy of this trace fossil was discussed by Uchman (Citation1999). Gaillard and Olivero (Citation2009) described Halimedides from a confined Early Cretaceous horizon in the Alps and regard it a deep-sea agrichnion, which chambers were used for food capture and storage, probably produced in stiff to firm substrates by a small crustacean.
?Hamipes isp.
Figure (A)
This is a trackway composed of two rows of hypichnial ridges aligned along almost straight line. The ridges are straight or slightly curved, 7–8 mm long, about 0.7 mm wide, pointed consistently from one side and obtuse from the other side. The stride (i.e. the distance between the relevant tips) is 28–32 mm. From the obtuse side, a knob-like ridge or a short, perpendicular ridge, 3 mm long, can be present. The rows are 7 mm apart, and the ridges from the two rows alternate.
This trace fossil is very similar to one flank of Hamipes didactylus Hitchcock, Citation1858 or to complete trackways of Acanthichnus cursorius Hitchcock, Citation1858 from the Lower Jurassic continental deposits of Massachusetts, both are referred to as an arthropod footprints (Rainforth, Citation2005), however, marks in H. didactylus are curved (Gibert, de Frenegal-Martínez, Buatois, & Mángano, Citation2000) and in both ichnotaxa, they are positioned symmetrically, differently from the described trace fossil in which the marks are located in the alternate position. A more alternated position of marks is displayed in Hamipes molassicus Bolliger & Gubler, Citation1997 from the Miocene freshwater molasse sediments in Switzerland. ?Hamipes isp. is also considered as a fragmentary preserved undertrack of Kouphichnium aspodon (Aldrich in Aldrich & Jones, Citation1930), in which some morphotypes display two rows of deep imprints on each side of the trackway (Buta, Kopaska-Merkel, Rindsberg, & Martin, Citation2005, plates 86(C)), 87(A, B)). Kouphichnium Nopsca, a limulid xiphosuran trackway, is known from shallow-marine and marginal-marine deposits, displays usually more morphological elements, with bifurcated branches in imprints of legs (Goldring & Seilacher, Citation1971).
Helminthoidichnites isp.
Figure (D)–(F)
Helminthoidichnites isp. is a horizontal, irregularly meandering, continuous or discontinuous hypichnial ridge or epichnial groove, 0.5–1.2 mm wide, which commonly shows rapid changes of course and occasional, short, irregular side protrusions.
Helminthoidichnites tenuis Fitch, Citation1850 shows more arcuate, curvatures, while H. isp. described herein shows the same type of rapid changes as in G. indianensis (Miller), to which it commonly transits and both ichnotaxa co-occur. This suggests that these trace fossils were produced by the same tracemaker. The main difference between them is that Helminthoidichnites Fitch, Citation1850 displays only occasional loops and is mostly locomotion trace, whereas in Gordia Emmons, Citation1844, loops are the most characteristic feature, pointing to a more localised activity, presumably referring to feeding. In contrast to these genera, Helminthopsis has no loops. The differences between Helminthoidichnites and Gordia have been analysed by Hofmann and Patel (Citation1989) and Hofmann (Citation1990), showing that Gordia has a distinctly stronger tendency to looping. Helminthoidichnites is known from marine Precambrian (Narbonne & Aitken, Citation1990) and marine-non-marine settings up to the Pleistocene (Uchman, Kazakauskas, & Gaigalas, Citation2009). In non-marine environments, it is interpreted as a probable nematophore or insect larvae trail (Buatois et al., Citation1997). However, arthropods can be producers on the basis of sets of oblique striae seen in some specimens (Uchman et al., Citation2009).
Helminthoidichnites cf. ornatus (Rindsberg & Kopaska-Merkel, Citation2005)
Figures (G) and (H),
This trace fossil is preserved as an epichnial, winding, V-shaped groove, 1–1.5 mm wide, with narrow levees and locally visible, perpendicular, hemispherical corrugations in 2 mm intervals. It can be abundant, with many crossings. This trace fossil is referred to Haplotichnus ornatus Rindsberg & Kopaska-Merkel, Citation2005, however the transverse corrugations in the latter look more like ribs covering flanks of the furrow. The type ichnospecies of Haplotichnus, H. indianensis Miller is involved in Gordia Emmons (see G. indianensis in this paper). Non-looping or occasionally looping traces of this sort are involved in Helminthoidichnites Fitch. H. ornatus is interpreted as a trace of marine arthropods (Rindsberg & Kopaska-Merkel, Citation2005).
Osculichnus cf. labialis Demircan & Uchman, Citation2010
Figure (A) and (B)
Figure 6. More trace fossils of the Mezardere Formation. (A), (B) Osculichnus cf. labialis, outcrop 4, specimen TRGM1/08. (C) P. tubularis, outcrop 1, specimen TRGM2/01. (D) P. beverleyensis, outcrop 8, specimen TRGM1/09. (E) P. montanus (Pl) and small burrows (sb), outcrop 12, specimen TRGM12/01. (F) P. montanus (Pl) and small burrows (sb), outcrop 1. (G) Ptychoplasma isp., outcrop 1, specimen TRGM12/02. (H) Treptichnus cf. apsorum, outcrop 7, specimen TRGM4/01. (I) Treptichnus cf. apsorum, outcrop 8, specimen TRGM4/02. (J) Treptichnus cf. apsorum, outcrop 4, specimen TRGM4/03. (K) Treptichnus cf. apsorum, outcrop 8, specimen TRGM4/04. (L) ?Treptichnus isp., outcrop 2, specimen TRGM12/03.

Osculichnus cf. labialis Demircan & Uchman, Citation2010 is an oval, shallow epichnial depression, 7–16 mm wide, 7–17 mm long, subdivided symmetrically or asymmetrically into two even or uneven lobes by a median ridge. The ridge is straight or curved, mostly wider towards both or one termination, well or poorly outlined, about 1–2 mm wide. The depressions are isometric or elongated according to the median ridge. Some of the depressions are rimmed by low levees. Specimens showing the typical, hypichnial preservation of O. labialis have not been found. This trace fossil is interpreted as a smaller, epichnial version of O. labialis Demircan & Uchman, Citation2010, which is produced by mouth of fishes hunting in the sediment.
Palaeophycus tubularis Hall, Citation1847
Figure (C)
This is a hypichnial and endichnial, slightly winding, slightly undulating, thinly lined, rarely branched, smooth, tubular ridge, 2–3 mm wide, which is traced up to about 100 mm on the bedding plane. P. tubularis is a facies-crossing form produced by carnivorous or omnivorous animals, mostly polychaetes (Pemberton & Frey, Citation1982). It is a trace fossil that occurs both in shallow and deep marine environments, though it is common in nearshore settings (Buatois & Mángano, Citation2011; Pemberton et al., Citation2001).
Planolites beverleyensis Billings, Citation1862
Figure (D)
Planolites beverleyensis is an endichnial, horizontal to oblique, winding, tubular, unbranched unlined, smooth structure, 2.5–4 mm wide, in which the filling is lighter in colour than the surrounding rock. It is visible in parting surface in siltstones. Planolites occurs in marine and non-marine facies. It is interpreted as a pascichnion, probably produced by a number of different organisms (Keighley & Pickerill, Citation1995; Pemberton & Frey, Citation1982).
Planolites montanus Richter, Citation1937
Figure (E) and (F)
This is a hypichnial and endichnial tubular burrow, 3–4 mm wide, which can be traced on the bedding sole for up to 5 mm in most cases, rarely in longer segments. It plunges into the bed and crosses the bedding surface under different angles. Its tendency to dipping in the bed expressed by short segments on the bedding surface is the main difference of P. montanus from P. beverleyensis, which points to a more horizontal burrowing.
Ptychoplasma isp.
Figures (C), (G)
Ptychoplasma isp. is a slightly curved row of hypichnial, elongated ridges. In one specimen (Figure (C)), the best preserved ridge is 10 mm long, 3 mm wide and pointed at the ends. Centres of adjacent ridges are located 10 mm apart, and the course of the trace fossils is marked by a discontinuous crest. In the second specimen (Figure (G)), the ridges are 3–4 mm long, 2 mm wide and the centres are located 6–9 mm apart. The best preserved ridge looks like Lockeia, which is a bivalve resting trace. The series of ridges proves locomotion of tracemakers interrupted by resting phases, which is a principle feature of Ptychoplasma Fenton and Fenton, Citation1937b (Uchman, Mikuláš, & Rindsberg, Citation2011). Ptychoplasma is a locomotion trace, interrupted by resting phases, produced by bivalves in continental, marginal marine, shallow-marine to deep-sea environments (Uchman et al., Citation2011).
Treptichnus cf. apsorum Rindsberg & Kopaska-Merkel, Citation2005
Figure (H)–(K)
This is a hypichnial, horizontal to sub-horizontal trace fossil (Treptichnus isp. in Demircan & Uchman, 2013) composed of almost straight or slightly winding, thinly lined segments. The direction shows rapid changes at an angle up to 30°, rarely more. The segments, 1.5–2.5 mm (rarely 1 mm wide), 10–20 mm, (occasionally up to 45 mm long), are preserved in full relief in fine-grained sandstone beds. Some of them plunge gently into the bed or in the underlying mudstone. Treptichnus apsorum known from Pennsylvanian estuarine tidal flat sediments of Alabama and interpreted as an insect trace (Rindsberg & Kopaska-Merkel, Citation2005), displays more curved and more irregular segments. Cited authors discussed the taxonomic and environmental problems of Treptichnus Miller and like to restrict this ichnogenus mostly to non-marine forms, however on the basis of interpretative criteria.
?Treptichnus isp.
Figure (L)
A hypichnial zigzag ridge, 2–3 mm wide. The segments of the ridge are 6–9 m long, and are at an angle of less than 90° in respect to the adjacent segment. Only one, short protrusion of the segment from the kink of the zigzag is seen. The zigzag course and the protrusion points to the ichnogenus Treptichnus, but the incomplete preservation prevents a more detailed determination.
?Undichna isp.
Figure (A) and (B)
Figure 7. ?Undichna isp., outcrop 8. (A) Plate with ?Undichna isp., and Helminthoidichnites cf. ornatus (upper left detail in Figure (G)) and Helminthoidichnites isp, specimen TRGM8/02. (B) The same plate with ?Undichna isp. traced by lines, specimen TRGM8/02.
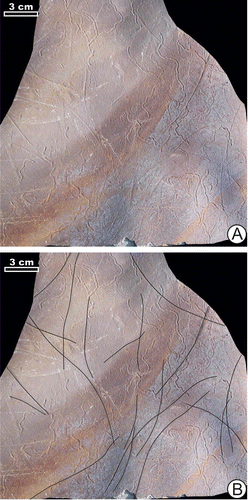
This trace fossil appears as an even, gently curved, thin hypichnial ridges or epichnial furrows, up to 1 mm wide, of which curvatures are at least 50 mm wide and amplitude is up to 15 mm. It co-occurs with abundant Helminthoidichnites. Undichna Anderson is interpreted as the traces by fins of swimming fish touching the sediment surface (Gibert, Citation2001; Trewin, Citation2000).
Small burrows
Figure (E) and (F)
Hypichnial, differently oriented tubular burrows, 0.5 mm in diameter, visible in 1–2 mm long, rarely longer segments, which show their straight or slightly winding course. The variable orientation resembles P. montanus, with which this trace fossil co-occurs, but P. montanus is a magnitude larger. It is not excluded that they can be produced by the juvenile tracemakers of P. montanus.
Arthropod trackway
Figure (A)–(F)
Figure 8. Some arthropod trace fossils from Mezardere Formation, outcrop 4. (A) Arthropod trackways, specimen TRGM1/10. (B) Resting traces, specimen TRGM1/10. (C) Detail of B, with indication of examples of oblique furrows (f) inclined in opposite direction in respect to the longer trace axis, specimen TRGM1/10. (D) Resting traces. (E), (F) Arthropod trackways and Archaeonassa isp. A (Ar), including specimen showing irregular wrinkles (w), specimen TRGM1/10.

Arthropod trackways are epichnial paths, 3–3.5 mm wide, composed of a central uneven, slightly winding median ridge and side, narrow, distally curved imprints. The median ridge is up to 1 mm wide. The imprints join the ridge tangentially under a 45° angle. They are 1–1.5 mm apart. Along the path, the median ridge can disappear and only the oblique imprints are visible. This trackway displays the same, basic morphological elements as the resting trace. Probably, the same tracemaker produced both of them as wells the co-occurring Archaeonassa isp. A and B.
Resting trace
Figure (B)–(D)
Resting traces are epichnial concave structures, 8–11 mm long, 3.5–4.5 mm wide, composed of elevated axial part, and thin, less than 0.5 m wide, up to 2 mm long straight or curved imprints (furrows), which run obliquely from the axial part, in generally opposite direction on each side. Some of the furrows criss-cross in the middle part. The axial part is about 1 mm wide. The morphology suggests an animal with appendages on each side, rear and front of elongated body, most probably an isopod, which used pereopod, antennae and uropods.
4. Discussion
The trace fossil assemblage is moderately diverse (22 different ichnotaxa referred to 13 ichnogenera and 3 forms left in open nomenclature). It is dominated by locomotion and feeding traces (pascichnia), e.g. Helminthoidichnites, Gordia, Bichordites. Other categories are rare and they are represented by domichnia (?Arenicolites), natichnia (?Undichna), cubichnia (resting trace) and praedichnia (Osculichnus). Only a few trace fossils are more frequent, including Gordia, Helminthoidichnites and Palaeophycus (Table ). B. kuzunensis occurs abundantly, but only in a single bed in one outcrop. The majority of other trace fossils was found only on one or two outcrops as a single or a few specimens. Very rarely, more than one ichnotaxon can be found in the same bed. Most beds do not contain any trace fossils. Even the beds containing trace fossils are poorly bioturbated, usually only at the top of ripple laminated sandstones. Their sedimentary structures are almost undisturbed in cross section. These features suggest a shallow, occasional, opportunistic colonisation, usually by one tracemaking organism.
Table 2. Occurrences of trace fossils in outcrops.
The features mentioned above fit well with the general ichnological characteristics of deltaic sediments, which display impoverished ichnodiversity, low degree of bioturbation, sporadic colonisation and recolonization, and paucity of trace fossils produced by suspension feeders. All of these features are a consequence of variable environmental stress factors, caused mainly by high sedimentation rate and input of fresh, commonly turbid waters, which eliminate, in particular, stenohaline and filter-feeding organisms (Gingras et al., Citation1998; MacEachern et al., Citation2005; Tonkin, Citation2012).
According to Tonkin (Citation2012), distal delta front to prodelta sediments contain trace fossils of the distal Cruziana and the Zoophycos ichnofacies, with common contribution of Thalassinoides, Phycosiphon, Chondrites, Zoophycos, Scolicia and Palaeophycus, and common bioturbational disturbance of sedimentary structures. However, the Mezardere Formation trace fossil assemblage is not a typical one. It displays more similarities to ichnoassemblages strongly influenced by freshwaters, for instance these from the Upper Carboniferous estuarine tidal rhythmites of Kansas (Buatois et al., Citation1997, Citation1998; Mángano and Buatois, Citation2004) as well as from the Upper Carboniferous estuarine tidal flat clastic sediments of Alabama (Rindsberg & Kopaska-Merkel, Citation2005) and lacustrine deltas (Melchor et al., Citation2003). These units show abundance of G. indianensis, Helminthoidichnites and arthropod trackways. Therefore, it is proposed that the prodelta sediments described herein were deposited in mainly strongly brackish waters, with only short episodes of normal salinity marked by the occurrence of B. kuzunensis, which was produced probably by irregular echinoids – a generally stenohaline organism (Demircan & Uchman, Citation2010). Palynomorphs also point to a marine influence.
Buatois and Mángano (Citation2011) presented examples of well bioturbated proximal prodelta sediments in a Middle Jurassic river dominated delta. However, the prodelta sediments of the Mezardere Formation are poorly bioturbated (BI = 0–1). Sediments of the Baltic Sea (salinity about 1%) are better bioturbated than the studied deposits (Bentley & Nittrouer, Citation1999; Virtasalo, Kotilainen, & Gingras, Citation2006). In the Danish Straits (salinity 1–1.5%), bioturbation is stronger (Winn, Citation2006). However, the mentioned Baltic deposits accumulated beyond the deltaic influences. Virtasalo et al. (Citation2011) presented ichnofabrics from strongly brackish Holocene sediments of the Baltic Sea, in which trace fossils resemble very much the typically lacustrine Mermia ichnofacies. In fact, the trace fossils from the Mezardere Formation display the same similarity by the common occurrence of Gordia, Helminthoidichnites, Treptichnus and arthropod traces (see Buatois & Mángano, Citation2011). The freshwater influences can be enhanced by possible hyperpycnal flows, which depauperate trace fossil assemblages in river-dominated deltas (Tonkin, Citation2012). It is possible that the thin sandstone beds in the Mezardere Formation are hyperpycnites.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Aldrich, T. H., & Jones, W. B. (1930). Footprints from the coal measures of Alabama. Alabama Museum of Natural History, Museum Paper, 9, 1–64.
- Bentley, S. J., & Nittrouer, C. A. (1999). Physical and biological influences on the formation of sedimentary fabric in a fjord-like depositional environment: Eckernförde Bay, southwestern Baltic Sea. Palaios, 16, 469–485.
- Bertling, M., Braddy, S. J., Bromley, R. G., Demathieu, G. R., Genise, J., Mikuláš, R., Nielsen, J. K., & Uchman, A. (2006). Names for trace fossils: A uniform approach. Lethaia, 39, 265–286.10.1080/00241160600787890
- Billings, E. (1862). New species of fossils from different parts of the Lower, Middle and Upper Silurian rocks of Canada. In Palaeozoic Fossils, vol. I. Geological Survey of Canada (pp. 96–168). Montreal: Dawson Brothers.
- Bolliger, T., & Gubler, T. (1997). Arthropodenlaufspuren von Hochdorf (LU) und weitere Arthropodennachweise aus der Obern Süsswassermolasse der Schweiz [Arthropod-walking traces from Hochdorf (LU) and further evidences of arthropods from the Upper Freshwater Molasse of Switzerland]. Eclogae Geologicae Helvetiae, 90, 617–627.
- Buatois, L. A., & Mángano, M. G. (2011). Ichnology: Organism-substrate interactions in space and time. Cambridge: Cambridge University Press. 358 pp.10.1017/CBO9780511975622
- Buatois, L. A., Mángano, M. G., Maples, C. G., & Lanier, W. P. (1997). The paradox of nonmarine ichnofaunas in tidal rhythmites: Integrating sedimentologic and ichnologic data from the late Carboniferous of eastern Kansas, USA. Palaios, 12, 467–481.10.2307/3515384
- Buatois, L. A., Mángano, G. M., Maples, C. G., & Lanier, W. P. (1998). Ichnology of an Upper Carboniferous fluvio-estuarine paleovalley: The Tonganoxie Sandstone, Buildex quarry, eastern Kansas, USA. Journal of Paleontology, 72, 152–180.10.1017/S0022336000024094
- Buckman, J. O. (1994). Archaeonassa Fenton & Fenton 1937 reviewed. Ichnos, 3, 185–192.10.1080/10420949409386387
- Buta, R. J., Kopaska-Merkel, D. C., Rindsberg, A. K., & Martin, A. J. (2005). Atlas of Union Chapel Mine invertebrate trackways and other traces. In R. D. Buta, A. K. Rindsberg, & D. C. Kopaska-Merkel (Eds.), Pennsylvanian footprints in the Black Warrior Basin of Alabama. Alabama Paleontological Society Monograph, 1, 277–337.
- Collins, A. G., Lipps, J. H., & Valentine, J. W. (2000). Modern mucociliary creeping trails and the bodyplans of Neoproterozoic trace-makers. Paleobiology, 26, 47–55.10.1666/0094-8373(2000)026<0047:MMCTAT>2.0.CO;2
- Crimes, T. P. (1977). Trace fossils of an Eocene deep-sea fan, northern Spain. In T. P. Crimes, T. P. & J. C. (Eds.), Trace fossils 2, Geological Journal Special Issue, 9, 71–90.
- Demircan, H., & Uchman, A. (2008). A mortal kiss of hunting fish: Trace fossil evidence from Late Eocene – Early Oligocene prodelta sediments of the Mezardere Formation, Thrace Basin, NW Turkey. In A. Uchman (Ed.), The second international congress on ichnology, Cracow, Poland, August 29 – September 8, 2008, Abstract book and the intra-congress field trip guidebook (p. 32). Warszawa: Polish Geological Institute.
- Demircan, H., & Uchman, A. (2010). Kiss of death of a hunting fish: Trace fossil Osculichnus labialis igen. et isp. nov. from late Eocene – early Oligocene prodelta sediments of the Mezardere Formation, Thrace Basin, NW Turkey. Acta Geologica Polonica, 60, 29–38.
- Demircan, H., & Uchman, A. (2012). The miniature echinoid trace fossil Bichordites kuzunensis isp. nov. from early Oligocene prodelta sediments of the Mezardere Formation, Gökçeada Island, NW Turkey. Acta Geologica Polonica, 62, 205–215.
- Demircan, H., & Uchman, A. (2013a, June). Ichnological features of prodelta sediments from the Mezardere Formation (Late Eocene – Early Oligocene), Gökçeada Island, NW Turkey. Paper presented during conference: H. Demírcan (Ed.), XII. International ichnofabric workshop, 30th June–5th July 2013, Çanakkale – Türkiye, Abstracts (pp. 24–25).
- Demircan, H., & Uchman, A. (2013b, June). 4th July 2013, Gökçeada Island. In H. Demírcan & A. Uchman (Eds.), Ichnological sites of the Gelibolu Peninsula and the Gökçeada Island, Thrace, NW Turkey. XII. International Ichnofabric Workshop, 30th June–5th July 2013, Çanakkale-Türkiye. Field Guidebook (pp. 69–82).
- Emmons, E. (1844). The Taconic system based on observations in New-York, Massachusetts, Maine, Vermont, and Rhode-Island. Albany: Caroll and Cook. 63 pp.
- Eocene-Oligocene sedimentation in Keşan area, southwestern Thrace. Turkey. The Bulletin of Mineral Research and Exploration, 69, 1–10
- Fenton, C. L., & Fenton, M. A. (1937a). Archaeonassa: Cambrian snail trails and burrows. American Midland Naturalist, 18, 454–458.10.2307/2420587
- Fenton, C. L., & Fenton, M. A. (1937b). Burrows and trails from Pennsylvanian rocks of Texas. American Midland Naturalist, 18, 1079–1084.10.2307/2420606
- Fitch, A. (1850). A historical, topographical and agricultural survey of the County of Washington. Part 2–5. Transactions of the New York Agricultural Society, 9, 753–944.
- Gaillard, C., & Olivero, D. (2009). The ichnofossil Halimedides in Cretaceous pelagic deposits from the Alps: Environmental and ethological significance. Palaios, 24, 257–270.10.2110/palo.2008.p08-055r
- Gibert, J. M., de, (2001). Undichna gosiutensis, isp. nov.: A 115 new fish trace fossil from the Jurassic of Utah. Ichnos, 8, 15–22.
- Gibert, J. M., de, Frenegal-Martínez, M. A., Buatois, L. A., & Mángano, M. G. (2000). Trace fossils and their palaeoecological significance in Lower Cretaceous lacustrine conservation deposits, El Montsec, Spain. Palaeogeography, Palaeoclimatology, Palaeoecology, 156, 89–101.10.1016/S0031-0182(99)00133-9
- Gingras, M. K., MacEachern, J. A., & Pemberton, S. G. (1998). A comparative analysis of the ichnology of wave- and river-dominated allomembers of the Upper Cretaceous Dunvegan Formation. Bulletin of Canadian Petroleum Geology, 46, 51–73.
- Gökçen, L. S. (1967). Eocene-Oligocene sedimentation in Keşan area, southwestern Thrace. Turkey. The Bulletin of Mineral Research and Exploration, 69, 1–10 (in Turkish).
- Goldring, R. (1962). The trace fossils of the Baggy Beds (Upper Devonian) of north Devon, England. Paläontologische Zeitschrift, 36 232–251.10.1007/BF02986976
- Goldring, R., & Seilacher, A. (1971). Limulid undertracks and their sedimentological implications. Neues Jahrbuch für Geologie und Paläontologie, Abhandlungen, 137, 422–442.
- Hakes, W. G. (1976). Trace fossils and depositional environment of four clastic units, Upper Pennsylvanian megacyclothems, northeast Kansas. The University of Kansas, Palaeontological Contributions, 63, 1–46.
- Hall, J. (1847). Paleontology of New York (Vol. 1, pp. 362). Albany: C. Van Benthuysen.
- Hitchcock, E. (1858). Ichnology of New England. A Report on the Sandstone of the Connecticut Valley, Especially its Fossil Footmarks (220 pp.). Boston, MA: William White.
- Hofmann, H. J. (1990). Computer simulation of trace fossils with random patterns, and the use of goniograms. Ichnos, 1, 15–22.10.1080/10420949009386327
- Hofmann, H. J., & Patel, I. M. (1989). Trace fossils from the type “Etchemian Series” (Lower Cambrian Ratcliffe Brook Formation), Saint John area, New Brunswick, Canada. Geological Magazine, 126, 139–157.10.1017/S0016756800006294
- Hoşgörmez, H., & Yalçın, M. N. (2005). Gas-source rock correlation in Thrace basin, Turkey. Marine and Petroleum Geology, 22, 901–916.10.1016/j.marpetgeo.2005.04.002
- Karahanoğlu, N., Eder, A., & İlleez, A. I. (1995). Mathematical approach to hydrocarbon generation history and source rock potential in the Thrace Basin, Turkey. Marine and Petroleum Geology, 12, 587–596.10.1016/0264-8172(95)98086-K
- Keighley, D. G., & Pickerill, R. K. (1995). The ichnotaxa Palaeophycus and Planolites: Historical perspectives and recommendations. Ichnos, 3, 301–309.10.1080/10420949509386400
- Kesgin, Y., & Varol, B. (2003). Gökçeada and Bozcaada Tertiary Geology. The Bulletin of Mineral Research and Exploration, 126, 49–67.
- Kilbourne, K. H., Curran, H. A., & Wilson, M. A. (1998). Ichnocoenoses and paleoenvironments of the Middle Jurassic Carmel-Twin Creek Seaway (Carmel Formation, southwestern Utah). Society of Economic Paleontologists and Mineralogists – AAPG Annual Meeting Expanded Abstracts Book (pp. 217–220). AAPG, Salt Lake City.
- Lebküchner, R. F. (1974). The Geology of the middle Thrace Oligocene. The Bulletin of Mineral Research and Exploration, 83, 1–29. (in Turkish).
- MacEachern, J. A., Bann, K. L., Bhattacharya, J. P., & Howell, C. D., Jr (2005). Ichnology of deltas; Organism responses to the dynamic interplay of rivers, waves, storms, and tides; River deltas; concepts, models, and examples. Special Publication-Society for Sedimentary Geology, 83, 49–85.
- Mángano, M. G., & Buatois, L. A. (2003). Trace fossils. In J. L. Benedetto (Ed.), Ordovician Fossils of Argentina (pp. 507–534). Córdoba: Universidad Nacional de Córdoba, Secrtetaria de Ciencia y Tecnologia.
- Mángano, M. G., & Buatois, L. A. (2004). Ichnology of Carboniferous tide-influenced environments and tidal flat variability in the North American Midcontinent. In D. McIlroy (Eds.), The application of ichnology to palaeoenvironmental and stratigraphic analysis. Geological Society of London, Special Publication, 228, 157–178.10.1144/GSL.SP.2004.228.01.09
- Melchor, R. N., Bellosi, E. S., & Genise, J. F. (2003). Invertebrate and vertebrate trace fossils from a Triassic lacustrine delta: The Los Rastros Formation, Ischigualasto Provincial Park, San Juan, Argentina. Asociación Paleontológica Argentina, Publicación Especial, 9, 17–33.
- Miller, S. A. (1889). North American geology and paleontology for the use of amateurs, students and scientists (664 pp.). Cincinnati, OH: Western Methodist Book Concern.
- Minter, N. J., Krainer, K., Lucas, S. G., Braddy, S. J., & Hunt, A. P. (2007). Palaeoecology of an Early Permian playa lake trace fossil assemblage from Castle Peak, Texas, USA. Palaeogeography, Palaeoclimatology, Palaeoecology, 246, 390–423.10.1016/j.palaeo.2006.10.009
- Narbonne, G. M., & Aitken, J. D. (1990). Ediacaran fossils from the Sekwi Brook area, Mackenzie Mountains, northwestern Canada. Palaeontology, 33, 945–980.
- Pemberton, G. S., & Frey, R. W. (1982). Trace fossil nomenclature and the Planolites-Palaeophycus dilemma. Journal of Paleontology, 56, 843–881.
- Pemberton, S. G., Spila, M., Pulham, A. J., Saunders, T., Maceachern, J. A., Robbins, D., & Sinclair, I. (2001). Ichnology and sedimentology of shallow to marginal marine systems. Geological Association of Canada, Short Course, 15, 1–343 pp.
- Rainforth, E. C. (2005). Ichnotaxonomy of the fossil footprints of the Connecticut Valley (Early Jurassic, Newark Supergroup, Connecticut and Massachusetts) (1302 pp.). ( PhD thesis), Columbia University, New York.
- Richter, R. (1937). Marken und Spuren aus allen Zeiten. I-II. [Marks and traces of all ages. 1–2]. Senckenbergiana, 19, 150–163.
- Rindsberg, A. K., & Kopaska-Merkel, D. C. (2005). Treptichnus and Arenicolites from the Steven C. Minkin Paleozoic Footprint site (Langsettian, Alabama, USA). In R. J., Buta, A. K. Rindsberg, & D. C. Kopaska-Merkel (Eds.), Pennsylvanian footprints in the Black Warrior Basin of Alabama: Alabama Paleontological Society Monograph, 1, 121–141.
- Saner, S. (1985). Sedimentary sequences and tectonic setting of Saros Gulf area-Northeast Agean Sea, Turkey. Bulletin of the Geological Society of Turkey, 28, 1–10.
- Siyako, M. (2006). Trakya havzası Tersiyer kaya birimleri [Thrace Basin Tertiary rock units]. In M. Üzer (Ed.), Trakya Bölgesi Litostratigrafi Birimleri [Lithostratigraphic units of the Thrace region] (pp. 43–83). Ankara: Maden Tetkik ve Arama Genel Müdürlüğü. ( In Turkish).
- Siyako, M., & Huvaz, O. (2007). Eocene stratigraphic evolution of the Thrace Basin, Turkey. Sedimentary Geology, 198, 75–91.10.1016/j.sedgeo.2006.11.008
- Stanley, T. M., & Feldmann, R. M. (1998). Significance of nearshore trace-fossil assemblages of the Cambro-Ordovician Deadwood Formation and Aladdin Sandstone, South Dakota. Annals of Carnegie Museum, 67, 1–51.
- Sümengen, M., Terlemez, İ., Şentürk, K., Karaköse, C., Erkan,N. E., Ünay, E., Atalay, Z. (1987).Gallipoli Peninsula and stratigraphy of the Tertiary basin Southwestern Thrace, sedimentology and tectonics. Unpublished MTA report. Ankara, 337 pp. (In Turkish).
- Temel, R. Ö., & Çiftçi, N. B. (2002). Stratigraphy and depositional environments of the Tertiary sedimentary units in Gelibolu peninsula and islands of Gökçeada and Bozcaada (Turkey). Turkish Association of Petroleum Geologists Bulletin, 14, 17–40.
- Tonkin, N. S. (2012). Deltas. In R. G. Bromley, & D. Knaust (Eds.), Trace fossils as indicators of sedimentary environments. Developments in Sedimentology, 64, 507–528. 10.1016/B978-0-444-53813-0.00017-4
- Trewin, N. H. (2000). The ichnogenus Undichna, with examples from the Permian of the Falkland Islands. Palaeontology, 43, 979–997.10.1111/pala.2000.43.issue-6
- Turgut, S., & Eseller, G. (1999). Sequence stratigraphy, tectonics and depositional history in eastern Thrace Basin, NW Turkey. Marine and Petroleum Geology, 17, 61–100.
- Turgut, S., Siyako, M., & Dilki, A. (1983). Geology and hydrocarbon potential of the Thrace Basin. Turkish Geological Congress Bulletin, 4, 35–46.
- Uchman, A. (1999). Ichnology of the Rhenodanubian flysch (Lower Cretaceous-Eocene) in Austria and Germany. Beringeria, 25, 65–171.
- Uchman, A., Kazakauskas, V., & Gaigalas, A. (2009). Trace fossils from Late Pleistocene lacustrine varved sediments in eastern Lithuania. Palaeogeography, Palaeoclimatology, Palaeoecology, 272, 199–211.10.1016/j.palaeo.2008.08.003
- Uchman, A., Mikuláš, R., & Rindsberg, A. K. (2011). Mollusc trace fossils Ptychoplasma Fenton & Fenton, 1937 and Oravaichnium Plička & Uhrová, 1990: Their type material and ichnospecies. Geobios, 44, 387–397.10.1016/j.geobios.2010.08.001
- Ünal, O. T. (1967). Thrace geology and oil facilities. (TPAO, Turkish Petroleum Corporation archives no. 391) (pp. 1–80). Ankara. (In Turkish).
- Virtasalo, J. J., Kotilainen, A. T., & Gingras, M. K. (2006). Trace fossils as indicators of environmental change in Holocene sediments of the Archipelago Sea, northern Baltic Sea. Palaeogeography, Palaeoclimatology, Palaeoecology, 240, 453–467.10.1016/j.palaeo.2006.02.010
- Virtasalo, J. J., Bonsdorff, E., Moros, M., Kabel, K., Kotilainen, A. T., Ryabchuk, D., & Hämäläinen, K. (2011). Ichnological trends along an open-water transect across a large marginal-marine epicontinental basin, the modern Baltic Sea. Sedimentary Geology, 241, 40–51.10.1016/j.sedgeo.2011.09.010
- Winn, K. (2006). Bioturbation structures in marine Holocene sediments of the Great Belt (Western Baltic). Meyniana, 58, 157–178.
- Yochelson, E. L., & Fedonkin, M. A. (1997). The type specimens (Middle Cambrian) of the trace fossil Archaeonassa Fenton & Fenton. Canadian Journal of Earth Sciences, 34, 1210–1219.10.1139/e17-097