
Abstract
Nummulites with reticulate septal filaments stratigraphically span from the Bartonian to the Rupelian Stages. The size of the proloculus of the megalospheric forms of reticulate species helped recognise N. fabianii – N. fichteli lineage in western Tethys. Unlike the species of this lineage, N. ptukhiani, described from Armenia, is characterised by an unusually large proloculus. A possibly second lineage of reticulate species comprising N. ptukhiani is recently reported from Lutetian – Bartonian succession of Tanzania. The present study examines reticulate species from palaeogeographically adjacent Bartonian succession of Kutch. The statistical analysis of the biometric data suggests the presence of three distinct species, referred to Nummulites ptukhiani, N. aff. hormoensis and N. acutus. The reticulation starts developing in N. acutus that ranges from P13 to P14 in its type locality, Kutch. We infer that Nummulites ptukhiani and N. aff. hormoensis possibly evolved from N. acutus in Zone P14. A binary tree model based on Classification and Regression Tree is proposed to statistically discriminate the three reticulate species.
Introduction
In the planispirally coiled foraminiferal genus Nummulites the septa continue over the lateral surfaces of the test as septal filaments. The septal filament traces are categorised as primary (arising directly from the septa), secondary (originating from the primary septa) and tertiary (derived from both primary and secondary septa) (Adams, Citation1988). The 3-D reconstruction by micro-CT however has revealed that all three originate similarly as traces of the primary septal filaments (Renema & Cotton, Citation2015). In radiate or striate forms of the genus, the septal filaments are primary, straight to curved and converge towards the poles. The secondary filaments occur in sub-reticulate to reticulate forms. In reticulate Nummulites there is a complex network of secondary filaments and the primary filaments are partially or wholly lost. The reticulate species of Nummulites are of chronostratigraphic value in shallow marine successions of Bartonian – Rupelian age. The N. garganicus – N. hormoensis – N. fabianii – N. fichteli – N. bormidiensis lineage is well established in the western Tethys (Less & Özcan, Citation2012). The lineage, extending from the Zone SBZ17 to SBZ22, is characterised by progressively increasing diameter of the proloculus from 100 – 140 μm in N. garganicus to 200 – 300 μm in N. fabianii – N. fichteli and 300 – 450 μm in N. bormidiensis. Apart from this lineage is N. ptukhiani described from Armenia by Kacharava (Citation1969), having a large proloculus of up to 600 μm. It is further recognised in eastern Turkey (Less & Özcan, Citation2012) and very recently in Tanzania (Cotton, Pearson, & Renema, Citation2016). In India, Nummulites fabianii and N. fichteli are widely reported from Priabonian – Rupelian sections (Dasgupta, Citation1970; Nuttall, Citation1925; Shukla, Citation2008). Samanta, Bondhopadhaya, and Lahiri (Citation1990) reported Nummulites hormoensis for the first time from the late Middle Eocene of Kutch. Shukla (Citation2008) reported two more reticulate forms, Nummulites sp. cf. N. ptukhiani from uppermost Middle Eocene of Surat-Broach area and N. fabianii forma primitive from the Middle Eocene of Kutch and Cambay Basin. Biometric data of the species from India are very limited, however, the size of proloculus recorded in some of the studies suggest that the embryons of the recorded species are larger compared with that of the corresponding species in western Tethys. The proloculii of the reported N. hormoensis and N. fichteli are two times that of the respective species in the western Tethys (Table ). Does it imply that (i) the eastern Tethys developed an independent lineage of reticulate Nummulites and, (ii) there are multiple lineages evolving contemporaneously in a region? There is a need to re-examine the reticulate species of this region in the biometric framework of the modern taxonomy to compare with the taxa of the western Tethys. We therefore attempt this study to examine the earliest reticulate forms occurring in the Bartonian succession of Kutch and carry out a detailed biometric analysis to distinguish them by multivariate statistical analysis.
Table 1. Reticulate species of Nummulites reported from India.
Samples and methods
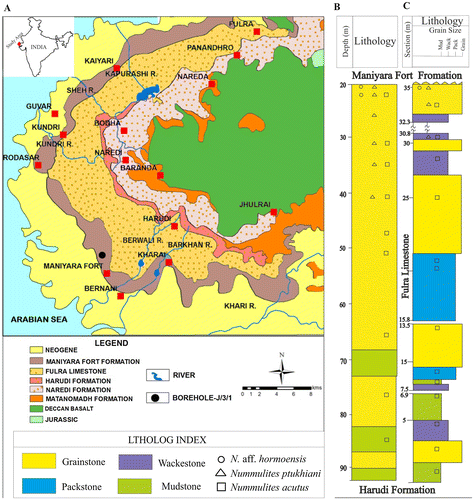
The samples for the present study are from a borehole and an outcrop section in Kutch (Figure ). Stratigraphically, they belong to Fulra Limestone of Bartonian age, referred by Samanta (Citation1970) to Zones Orbulinoides beckmanni – Truncorotaloides rohri (P13-P14). The planktonic foraminifera comprise Acarinina rohri, A. topilensis, Globigerina linaperta, G. officinalis, Globorotalia centralis, G. lehneri, Globoturborotalita ouachitaensis and Orbulinoides beckmanni among others (Mohan & Soodan, Citation1970; Samanta, Citation1970; Saraswati, Khanolkar, Raju, & Banerjee, Citation2016). The larger benthic foraminiferal assemblage includes Alveolina elliptica, Assilina exponens, Asterocyclina alticostata, A. sireli, Calcarina sp., Dictyoconoides cooki, Discocyclina dispansa, D. sowerbyi, Nummulites acutus, N. maculatus, N. stamineus and Orbitoclypeus haynesi, (Ismail-Lattrache et al., Citation2013; Özcan, Saraswati, Hanif, & Ali, Citation2016; Samanta & Lahiri, Citation1985; Samanta et al., Citation1990; Saraswati, Patra, & Banerji, Citation2000). The two types of reticulate species reported in this paper are confined to the upper part of the Fulra Limestone (Figure ). It may be mentioned that the reticulate species are low in abundance and not-so-well preserved. A relatively good number of specimens could be obtained only from borehole samples. We also re-examined our previous collection of Nummulites from Fulra Limestone in Berwali river section (Saraswati et al., Citation2000) for the morphometric data of N. acutus.
Figure 1. Locations of samples on the geological map of the study area (A), and the lithologs of the borehole (B) and outcrop sections (C).
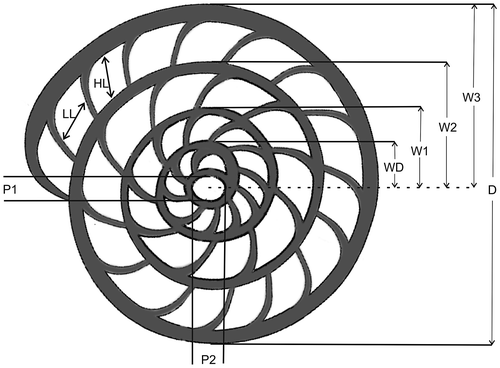
The reticulate Nummulites were separated into morphotypes based on external characters. Diameter and thickness of selected specimens were measured prior to preparations of their equatorial sections for biometric measurements (Figure ). We tried to study them by micro-CT but except for a few specimens the results were not satisfactory due to calcite infilling.
Figure 2. Schematic section of Nummulites to illustrate biometric measurements carried out in this study: P1, vertical proloculus height; P2, horizontal proloculus height; WD, distance from centre of proloculus to edge of deuteroloculus; W1, W2, W3, successive whorl radii; LL, HL, length and height of the chamber in last whorl; D, Diameter (schematic drawing of Nummulites after Drooger, Marks, & Papp, Citation1971).
The biometric data were subject to cluster analysis by paired group method using the PAST package (Hammer, Harper, & Ryan, Citation2001). We attempted to apply discriminant function, successfully used in an earlier study to discriminate species of Nummulites (Saraswati & Patra, Citation2000), but the biometric data of the studied populations were not multivariate normal (as per Mardia’s multivariate normality test). A non-parametric method, Classification and Regression Tree (CART), was therefore employed using Statistical Analysis Software. Saraswati and Sabnis (Citation2006) discussed the methodology of CART and its application in the taxonomy of Nummulites.
Results
The following two morphotypes of reticulate Nummulites are visually distinguished on their external characters (Pls.1-2):
Type-1: small and lenticular; reticulate; granular, granules spirally arranged and some granules merging at the poles.
Type-2: small and inflated (compared with Type-1); reticulate; coarsely granulated and a prominent polar knob.
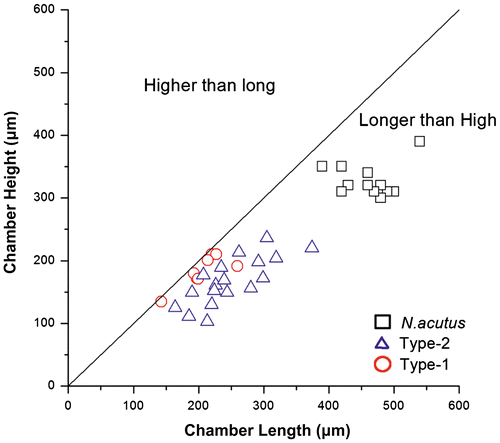
The diameter and thickness of the tests and the internal measurements in equatorial sections of the two morphotypes and that of N. acutus are given in Table . The bivariate plot of diameter and thickness of the tests shows that all the three examined taxa have inflated lenticular tests (Figure ). The bivariate plots of length and height, respectively, of the chambers in the last whorl express the shape of the chambers that are longer than high (Figure ).
Table 2. Statistical summary of the morphometric parameters of the reticulate Nummulites, Type-1, Type-2 and N. acutus from the Bartonian section of Kutch.
Figure 3. Bivariate plots of diameter and thickness of the three species of Nummulites. The tests of Type-1 and Type-2 samples fall in the field of inflate lenticular tests and N. acutus are inflate lenticular to sub-globular.
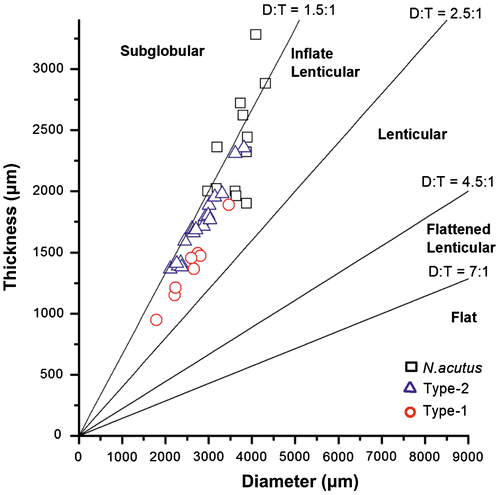
Figure 4. Bivariate plots of length and height of chambers in last whorl of the three species of Nummulites. All the samples have longer than high chambers.
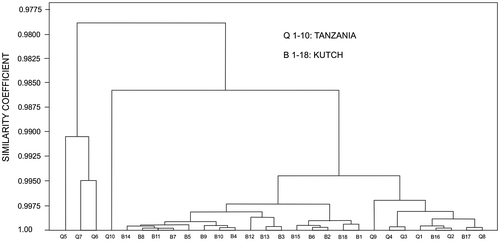
The cluster analysis of the biometric data distinctly classifies all the specimens of the two reticulate species in to their respective morphogroups (identified on external characters only), except for misclassification of a specimen of Type-2 with Type-1 (Figure ). It signifies the two types of reticulate taxa are not only externally distinguishable but also quantitatively distinct in terms of their various internal morphological characters. The phenogram of the two types and N. acutus (that somewhat resemble externally) is presented in Figure . The spiral diagrams (Figure ) suggest that spires are tighter in Type-1 compared with Type-2. The coiling of N. acutus falls between the two types. Although not all comparable parameters of the Tanzania samples are available, cluster analysis on common data in both Kutch (Type-2 populations) and Tanzania (data from stratigraphically equivalent Zone P14) was carried out for comparison (Figure ). Notwithstanding few data, Kutch and Tanzania populations of reticulate Nummulites are closely clustered, sharing high similarity coefficients.
Figure 5. Cluster analysis of the biometric data of Type-1 and Type-2 Nummulites. Except for one mismatch (B10), the specimen of Type-1 and Type-2 are distinctly clustered at high similarity coefficients.
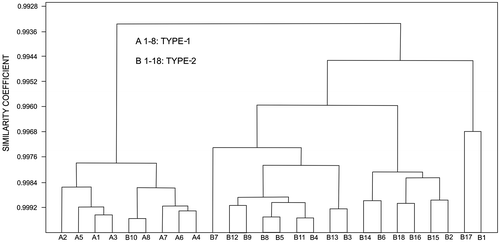
Figure 6. Cluster analysis of biometric data of Type-1, Type-2 and N. acutus. The phenogram shows that Type-1, Type-2 and N. acutus are closely clustered at high similarity coefficient, some of the specimen of N. acutus are however clustered with Type-1 and some with Type-2.
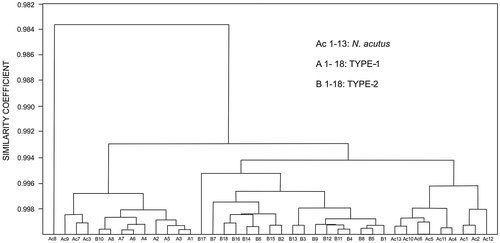
Figure 7. Spiral diagram of the three species. It shows tighter coiling in Type-1 compared with Type-2. Coiling of N. acutus falls between the two types. The sample from Tanzania (TDP 4; Cotton et al., Citation2016) falls in the broader field Type-2.
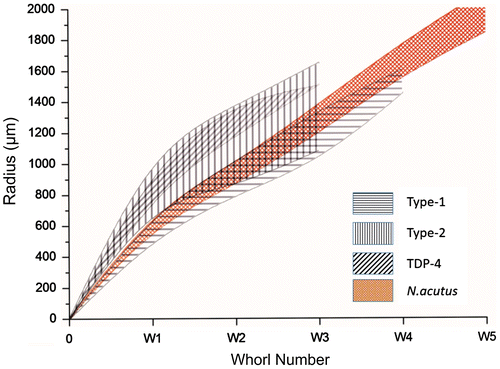
Figure 8. Cluster analysis of Type-2 and Tanzania samples suggesting close similarities of most specimens from the two regions.
Discussion
The Middle Eocene reticulate Nummulites of Kutch were reported for the first time in Bartonian Fulra Limestone by Samanta et al. (Citation1990), and referred them to N. hormoensis. Both A and B- forms were reported, characterised by coarse, spirally arranged granules and sinuous septal filaments in A forms and crude reticulate structure in B forms. Biometric data are not given in the publication to statistically compare it with the Type-1 in the present study. Roveda (Citation1970) provided some measurements on diameter and thickness of the tests and diameter of the proloculus of the topotypes of the species in Sedgwick Museum (UK). The Type-1 is smaller, inflated and have less number of whorls and significantly larger size of the proloculus compared with N. hormoensis (Table ). At this stage, we refrain from giving a new name to the Kutch species and assign Type-1 to N. aff. hormoensis.
Table 3. Comparison of morphometric data of Type-1 reticulate species of Kutch with N. hormoensis. The data of N. hormoensis are after Roveda (Citation1970).
The Type-2 in the studied assemblage differs distinctly from the Type-1 both qualitatively and statistically. The Type-1 tends to be lenticular, have more reticulation as well as granulation and smaller proloculus compared with Type-2. The Type-2 is more inflated and has a prominent polar knob. The average diameter of the proloculus in Type-2 is 600 μm, significantly larger than the average diameter ~400 μm in Type-1. The spire is high and opens rapidly in Type-2 compared with tighter coiling in Type-1 (Figure ). Statistically, the specimens of the two groups cluster separately in the phenogram (Figure ). This validates the differentiation of the two types based on external characters. The inflated tests, pronounced polar knobs and large proloculii (P2: 465–750 μm, average 600 μm) in Type-2 are comparable with Armenian N. ptukhiani (A-form) as discussed by Papazzoni (Citation1998). The Type-2 form closely resembles the recently reported Nummulites ptukhiani (population TDP4) from Tanzania (Cotton et al., Citation2016). The inflated lenticular test, comparable size of the proloculus, number of whorls, coarse granulations, presence of polar knob and similar nature of spire opening (Figure ) make the two populations from India and Tanzania remarkably similar. This is also reflected in cluster analysis (Figure ) in spite of limited quantitative data. We therefore assign Type-2 to Nummulites ptukhiani. The species chronostratigraphically lies at similar position in Zone P14 in India and Tanzania.
Sowerby (Citation1840) described Nummulites acutus from Lakhpat (Kutch). Samanta (Citation1982) studied the only remaining syntype specimen of the species at NHM, London, and observed gently wavy and irregularly branching septal filaments producing a subreticulate septal pattern on the surface of the microspheric form. We have re-examined our previous collections (Saraswati et al., Citation2000) and find that the A-forms of the species are characterised by reticulate pattern on the peripheral parts and somewhat resemble Nummulites ptukhiani (Type-2) in their inflated tests, large proloculii and polar knob. N. acutus however is comparatively larger and thicker, have sharp margin and distinctly larger chambers compared with N. ptukhiani. The cluster analysis of the combined data distinguishes the three species and, as explained below, N. acutus is clustered partially with both Type-1 and Type-2 (Figure ). Nummulites acutus occurs throughout the Fulra Limestone (Zones P13–P14).
There are general external similarities in the Bartonian reticulate Nummulites discussed in the present study. The small size, inflated lenticular tests, granular surface, and reticulation of varying degree are common external features of the three species. The internal morphology in equatorial sections varies. It is reflected in the cluster analysis as discussed before. The phenogram unambiguously distinguishes N. ptukhiani from N. aff. hormoensis. Interestingly, some specimens of N. acutus cluster at higher similarity coefficient with N. ptukhiani while the others cluster with N. aff. hormoensis. It is likely indicator of evolution of both N. ptukhiani and N. aff. hormoensis from N. acutus in Zone P14. In the suggested N. ptukhiani lineage of Tanzania, the Lutetian population does not show reticulation (Cotton et al., Citation2016). The reticulation develops in the population of Zone P13 in Tanzania and the reticulated N. acutus in Kutch ranges down to Zone P13. A future study may be important and interesting to compare the materials of the two regions to find out if this part of the eastern Tethys was the cradle of evolution of N. ptukhiani lineage.
We have attempted to statistically distinguish the Bartonian reticulate Nummulites of Kutch by CART. The majority of tree models in CART analysis were built on two key variables, height of chambers in the first whorl (HI) and horizontal proloculus height (P2). A binary tree model (Figure ) of the training data-set assigned all the specimens to the correct groups as N. acutus and N. ptukhiani but two (of the nine) specimens of N. aff. hormoensis were wrongly assigned to N. ptukhiani. All specimens in the validation data-set were correctly referred to their respective groups.
Figure 9. A binary tree model of N. acutus, N. ptukhiani and N. aff. hormoensis discriminating the three taxa on the basis of height of chambers in the first whorl (HI) and diameter of the protoconch (P2).
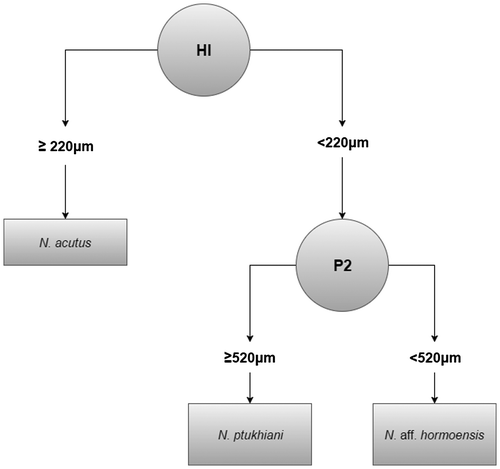
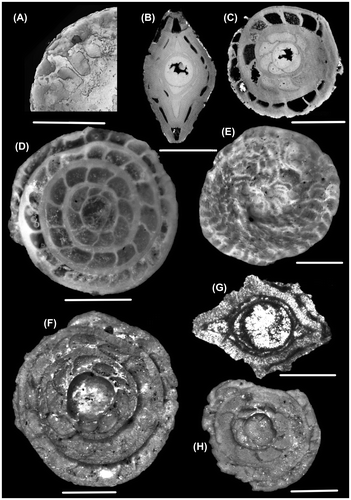
Plate 1. Photomicrographs of N. aff. hormoensis (external view: A, B, C; side view: E; equatorial section: D, F, G; axial section: I) and N. ptukhiani (external view: H). Scale bar: 1000 μm. A, B, D, E, I: borehole 21.20 m;; F,G: borehole 21.55 m; H: borehole 31.88 m depth; C: outcrop sample.

Plate 2. Photomicrographs of N. ptukhiani (external view: A; equatorial section: C, D; axial section: B, D) and N. acutus (external view: E; equatorial section: F, H; axial section: G). Scale bar: 1000 μm. A, B, C: borehole 21.55 m; D, E, F, G, H: outcrop.
Conclusions
Three species of reticulate Nummulites are recognised in the Bartonian succession of Kutch. These are referred to Nummulites ptukhiani, N. aff. hormoensis and N. acutus. It is observed that reticulation starts developing in Nummulites acutus, particularly visible in the A-forms of the species. We infer that Nummulites ptukhiani and N. aff. hormoensis possibly evolved from N. acutus in Zone P14. A binary tree model based on CART is proposed to statistically discriminate the three reticulate species.
Disclosure statement
No potential conflict of interest was reported by the authors.
Acknowledgement
We gratefully acknowledge the help of Prof. Sanjeev V. Sabnis in statistical analysis. We are thankful to Prof. George Less for reading the first draft of this paper and to the reviewer, Prof. Ercan Özcan, for his constructive suggestions that immensely improved the manuscript.
References
- Adams, C. G. (1988). Septa, septal traces and septal filaments in the foraminifera genus Nummulites Lamarck. Journal of Micropalaeontology, 7, 89–102.10.1144/jm.7.1.89
- Cotton, L. J., Pearson, P. N., & Renema, W. (2016). A new Eocene lineage of reticulate Nummulites (Foraminifera) from Kilwa district, Tanzania; a place for Nummulites ptukhiani? Journal of Systematic Palaeontology, 14, 569–579. doi:10.1080/14772019.2015.1079562
- Dasgupta, A. (1970). Further study in reticulate Nummulites (Foraminifera) from Oligocene beds of Cutch, India. Quarterly Journal of the Geological Mining and Metallurgical Society of India, 62, 159–172.
- Drooger, C. W., Marks, P., & Papp, A. (1971). Small radiate Nummulites of northwestern Europe. Utrecht Micropalaeontological Bulletin, 5, 1–137.
- Hammer, Ø., Harper, D. A. T., & Ryan, P. D. (2001). PAST: Paleontological statistics software package for education and data analysis. Palaeontologia Electronica, 4, 1–9.
- Ismail-Lattrache, K. B., Ozcan, E., Boukhalfa, B., Saraswati, P. K., Soussi, M., & Jovane, L. (2013). Early Bartonian orthophragminids (Foraminiferida) from Reineche Limestone, north African platform, Tunisia: Taxonomy and paleobiogeographic implications. Geodinamica Acta, 26, 94–121. doi:10.1080/09853111.2013.858950
- Kacharava, Z. D. (1969). O filogeneticheskom ryade gruppy Nummulites fabianii. Bulletin of the Academy of Sciences of the Georgian SSR, 55, 497–500.
- Less, G., & Özcan, E. (2012). Bartonian-Priabonian larger benthic foraminiferal events in the western Tethys. Austrian Journal of Earth Sciences, 105, 129–140.
- Mohan, M., & Soodan, K. S. (1970). Middle Eocene planktonic foraminiferal zonation of Kutch, India. Micropaleontology, 16, 37–46.10.2307/1484845
- Nuttall, W. L. F. (1925). Indian reticulate Nummulites. Annals and Magazine of Natural History, Ser 9, 9, 661–667.
- Özcan, E., Saraswati, P. K., Hanif, M., & Ali, N. (2016). New orthophragminid species with novel axial thickening from the Bartonian shallow marine of Indian Subcontinent. Geologica Acta, 14, 261–282.
- Papazzoni, C. A. (1998). Biometric analysis of Nummulites “ptukhiani” Z.D. Kacharava, 1969 and Nummulites fabianii (Prever in Fabiani, 1905). Journal of Foraminiferal Research, 28, 161–176.
- Renema, W., & Cotton, L. (2015). Three dimensional reconstructions of Nummulites tests reveal complex chamber shapes. PeerJ, 3, e1072. doi:10.7717/peerj.1072
- Roveda, V. (1970). Revision of the Nummulites (Foraminiferida) of the N. fabianii-fichteli group. Revista Italiana di Paleontologia, 76, 235–324.
- Samanta, B. K. (1970). Middle Eocene planktonic foraminifera from Lakhpat, Cutch, western India. Micropaleontology, 16, 185–215.10.2307/1485114
- Samanta, B. K. (1982). A restudy of two Middle Eocene species of Nummulites (Foraminiferida) from Cutch, Gujarat. Bulletin of Indian Geological Association, 15, 21–50.
- Samanta, B. K., & Lahiri, A. (1985). The occurrence of Discocyclina Gümbel in the middle Eocene Fulra limestone of Cutch, Gujarat, Western India, with notes on species reported from the Indian region. Bulletin Geological Mining Metallurgical Society of India, 52, 211–295.
- Samanta, B. K., Bondhopadhaya, K. P., & Lahiri, A. (1990). The occurrence of Nummulites Lamarck (Foraminiferida) in the Middle Eocene Harudi formation and Fulra Limestone of Kutch. Bulletin of Geological Mineralogical and Metallurgical Society of India, 55, 1–66.
- Saraswati, P. K., & Patra, P. K. (2000). Multivariate analysis of three closely resembling species of Nummulites from Middle Eocene of India. Revue de Micropalaeontology, 43, 319–326.10.1016/S0035-1598(00)90161-2
- Saraswati, P. K., & Sabnis, S. V. (2006). Comparison of CART and discriminant analysis of morphometric data in foraminiferal taxonomy. FORAMS 2006, Anuario do Instituto de Geociencias – UFRJ, Brazil, 29, 153–162.
- Saraswati, P. K., Patra, P. K., & Banerji, R. K. (2000). Biometric study of some Eocene Nummulites and Assilina from Kutch and Jaisalmer, India. Journal of the Palaeontological Society of India, 45, 91–122.
- Saraswati, P. K., Khanolkar, S., Raju, D. S. N., & Banerjee, S. (2016). An updated Eocene stratigraphy of Kutch. In M. G. Thakkar (Ed.), Recent studies on the geology of Kachchh. Special Publications 6 (pp. 25–31). Bengaluru: Geological Society of India.
- Sengupta, S. (2000). Problems of classifying Early Oligocene reticulate Nummulites (Foraminiferida) from southwestern Kutch, Gujarat. Journal of the Geological Society of India, 56, 673–677.
- Shukla, S. (2008). Atlas of taxonomic and bio-chronostratigraphic studies on Paleogene larger benthic foraminifera from Indian sedimentary basins. In Paleontographica Indica (Vol. 9, 183 pp). Dehradun: KDMIPE, ONGC.
- Sowerby, J. de C. (1840). Systematic list of organic remains. Appendix to grant C.W., Memoir to illustrate a geological map of Cutch. Transaction Geological Society of London Sr 2, 5, 317–329.