
Abstract
Orthophragminids from the Bartonian Fulra Limestone in Kutch, India and the coeval units in Sulaiman Range in Pakistan suggest the establishment of a significant number of endemic species in the Indian subcontinent (Eastern Tethys). Among a total of fifteen species of Discocyclina, Orbitoclypeus and Asterocyclina, six of them appear to be confined to Indian subcontinent while seven species are common both to the peri-Mediterranean/Europe region (Western Tethys) and Indian subcontinent. Two species, Asterocyclina sireli, a four-ribbed species of possibly Indo-Pacific origin, and Orbitoclypeus haynesi that form large populations in Fulra Limestone, appear to have spread into North Africa and Turkey but not into European platforms as a response to Middle Eocene Climatic Optimum (MECO). The lack of Lutetian and Priabonian fauna in the studied sections, either due to a hiatus or unsuitable depositional environments, hampers the establishment of the actual stratigraphic ranges of the identified taxa. Our record provides us to characterize the orthophragminids in shallow benthic zone (SBZ) 17 for Eastern Tethys in detail by comparing the data from the above localities with those from the North Africa, Europe and Turkey, showing the change in diversity.
1. Introduction
Paleocene and Eocene shallow-marine deposits in Indian subcontinent were traditionally regarded to have formed under three major transgressions, Ranikot (Paleocene), Laki (early Eocene) and Kirthar (middle Eocene), the latter being the most significant one owing to deposition of highly fossiliferous beds in India and Pakistan (Eames, Citation1952a, 1952b; Nagappa, Citation1959; Nuttall, Citation1926). The Fulra Limestone in Kutch Basin in western India and the correlative units in Pakistan, the Pirkoh and Drazinda formations in Sulaiman Range correspond to Bartonian Kirthar transgression coeval with the Middle Eocene Climatic Optimum (MECO) (Khanolkar, Saraswati, & Rogers, Citation2017). The Bartonian shallow-marine deposits are exposed in Kutch, Cambay and Jaisalmer basins in western India, in the Sulaiman and Kirthar ranges in Pakistan, and in the Assam-Meghalaya regions in eastern India. Generally referred to as ‘Nummulitic limestone’, these sequences contain abundant orthophragminids that are locally more common than nummulitids (Ali et al., Citation2018; Özcan, Saraswati, Hanif, & Ali, Citation2016b; Samanta & Lahiri, Citation1985). Our studies on the taxonomy and paleobiogeography of orthophragminids from the Fulra Limestone and Sulaiman Range in Pakistan revealed a significant number of new species that appear to be confined to Indian subcontinent (Ali et al., Citation2018; Özcan et al., Citation2016b). This suggests a major faunal differentiation between peri-Mediterranean/Europe region (Western Tethys) and Indian subcontinent (Eastern Tethys) in the middle Eocene. As opposed to Western Tethyan orthophragminid assemblages, Discocyclina, along with subordinate Asterocyclina, is the most common group in Fulra Limestone, whereas Orbitoclypeus is represented by a single species, O. haynesi. Pirkoh and Drazinda formations from Pakistan yielded predominantly Discocyclina consisting of some new species including Discocyclina pseudodispansa Özcan, Ali & Yücel, 2018, D. sulaimanensis Özcan, Ali & Hanif, 2016, D. rakhinalaensis Özcan, Ali & Yücel, 2018, D. zindapirensis Özcan, Ali & Yücel, 2018, and D. kutchensis Özcan & Saraswati, 2016, the latter one occurring also in the Fulra Limestone (Ali et al., Citation2018; Özcan et al., Citation2016b). A new morphological structure, bulges, observed at the test surface as semi-rounded to rounded thickening of the lateral layers, in D. kutchensis was introduced (Özcan et al., Citation2016b).
We aim to describe here in detail the orthophragminids from the Fulra Limestone to complement our findings from the Pirkoh and Drazinda formations in Pakistan (Ali et al., Citation2018). We make a general synthesis of Bartonian orthophragminids from the Indian subcontinent by incorporating and comparing the data from Western Tethys (Europe, north Africa and Turkey). A synthesis for shallow benthic zone (SBZ) 17 for Indian subcontinent is made in the light of new orthophragminid data from this region.
2. Geological setting and stratigraphy
2.1. Kutch Basin
In Kutch the Eocene succession represents the first marine transgression following the Deccan volcanism and consists of Naredi and Harudi formations and Fulra Limestone (Biswas, Citation1992; Saraswati, Khanolkar, & Banerjee, Citation2017) ((A), (B)). The Fulra Limestone conformably overlying the Harudi Formation is interpreted to have been deposited in a warm, oligotropic sea during the globally recognized warming period called Middle Eocene Climatic Optimum (MECO) (Khanolkar et al., Citation2017). It comprises exclusively of shallow marine carbonates consisting of various fossil groups including planktonic foraminifera at its lower part (Khanolkar et al., Citation2017; Samanta, Citation1970, 1993; Samanta, Bandopadhyay, & Lahiri, Citation1990; Samanta & Lahiri, Citation1985; Saraswati, Patra, & Banerjee, Citation2000; Saraswati et al., Citation2017; Sen Gupta, Citation1963a, 1963b). The unit is characterized by six facies, as orthophragminid mudstone (sample FUL1, (A)), orthophragminid wackestone-mudstone alternation, orthophragminid wackestone-packstone alternation, nummulitic grainstone, nummulitic wackestone-packstone alternation, and Alveolina wackestone-packstone alternation (samples FUL2–15) corresponding to various depositional settings on a carbonate ramp (Figures (C), (C)–(E)) (Banerjee, Khanolkar, & Saraswati, Citation2017). These facies indicate a depositional spectrum ranging from bar-lagoon to mid-ramp depositional setting. Orthophragminid mudstone in the lower part of the Fulra succession contains planktonic foraminifera including Orbulinoides beckmanni and records the deepest bathymetry, up to 60 m, corresponding to mid-ramp setting. The cross-stratified Nummulitic grainstone beds, with slightly convex-up geometry and common palaeokarst surfaces, represent a high-energy deposit in inner-ramp setting (Banerjee et al., Citation2017; Chattoraj, Sarkar, Chakraborty, Banerjee, & Saraswati, Citation2012). These beds record the shallowest deposits in the ramp indicating low-relief bars in shoal environment. Nummulitic wackestone-packstone alternation represents a shoal flank environment. Orthophragminid wackestone-packstone alternation and orthophragminid wackestone-mustone alternations form more distally. The repetetion of above facies record repeated phases of shallowing and deepening. The Fulra Limestone is overlain unconformably by the Maniyara Fort Formation.
Figure 1. Location of Kutch Basin and Sulaiman Range in W India and Pakistan (A), simplified geological map of the Kutch and position of the studied sections (B), and composite stratigraphic section of the Fulra Limestone (C).
Note: Geological map is after Biswas (Citation1992).
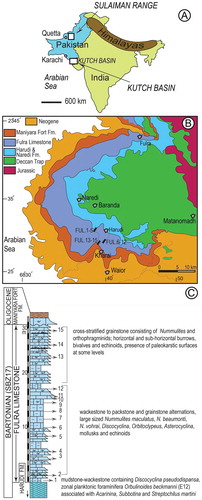
Figure 2. Field aspects of the Fulra Limestone: Transition from Harudi Formation to the Fulra Limestone (A, B), the lower beds of the Fulra Limestone (sample FUL1) with planktonic foraminifera and LBF (B), close-up view of sample FUL6 with predominantly Discocyclina discus and D. dispansa (C), a shallowing upward cycle with transition from wackestone-packstone alternation (FUL6–10) to grainstone facies (FUL11–12) in the middle part of the unit (D), the upper carbonates of the Fulra Limestone of predominantly grainstone facies (E).

The age of the Fulra Limestone is firmly constrained as Bartonian by the occurrence of key planktonic species identified at its lower part and in the upper part of the underlying Harudi Formation (see Saraswati et al., Citation2017 for details). Samanta (Citation1970) described planktic foraminifera from the Fulra Limestone and assigned it to the Orbulinoides beckmanni and Truncorotaloides (Acarinina) rohri zones. Saraswati, Khanolkar, Raju, Dutta, and Banerjee (Citation2014) identified Orbulinoides beckmanni, Acarinina rohni, A. topilensis, Streptochilus martini, Pseudohastigerina micra, and Jenkinsina columbiana in shale beds immediately below the Harudi Formation-Fulra Limestone boundary and assigned P13 Zone to this level. According to the scheme of Berggren and Pearson (Citation2005) the unit corresponds to the Zones E12 and E13. The Fulra Limestone was assigned to shallow benthic zone (SBZ) 17 based on orthophragminids (Ben Ismail-Lattrache et al., Citation2014; Özcan et al., Citation2016b) and benthic foraminiferal assemblages supplemented by the planktonic foraminifera (Khanolkar et al., Citation2017).
2.2. Sulaiman range
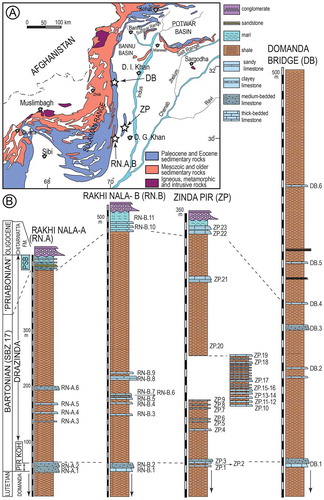
The Middle to Upper Eocene sedimentary sequence in Sulaiman Range in Pakistan consists of Habib Rahi, Domanda, Pirkoh and Drazinda formations that were collectively named as ‘Kirthar’ by Blanford (Citation1879) (). The term ‘Drazinda Shale Member’ of the ‘Kirthar’ Formation was introduced by Hemphill and Kidwai (Citation1973) to replace the ‘Upper Chocolate Clays’ of Eames (Citation1952a, 1952b). The Drazinda Formation, more than 380 meters in thickness consists of dark-brown to greenish gray shale and subordinate marl and limestone beds containing abundant LBF, bivalves, bryozoans and echinoids in its lower and middle, and pale yellowish green Pellatispira-bearing marls in upper part. The previous studies suggested a middle and/or late Middle Eocene age for the Drazinda Formation based on calcareous nannofossils (Köthe, Khan, & Ashraf, Citation1988), planktonic foraminifera (Afzal, Asrar, & Naseer, Citation1997; Samanta, Citation1973; Warraich & Nishi, Citation2003) and orthophragminids (Ali et al., Citation2018). According to Köthe et al. (Citation1988), the age of this unit in Rakhi Nala section is middle Eocene based on the presence of nannoplankton zones NP 16 and 17. Afzal et al. (Citation1997) identified P14 and dated the lower part of Drazinda Formation (below the ‘Pellatispira beds’) as late middle Eocene (Bartonian) in age. The Pellatispira-beds containing Heterostegina, reticulate Nummulites and Pellatispira is considered Priabonian (Özcan et al., Citation2016a). The Pirkoh Formation was deposited in the inner to outer shelf, and the bulk of the Drazinda Formation has been deposited in the shelf lagoon and shoal environments. The ‘Pellatispira beds’ of the Drazinda Formation have been deposited in the outer shelf (Abbas, Citation1999). The Drazinda Formmation is unconformably overlain by coastal deltaic to fluviatile deposits of Oligocene Chitarwatta Formation (Shah, Citation2009).
Figure 3. Location of Sulaiman Range in Pakistan (A), simplified geological map of the Sulaiman Range (B), and stratigraphic sections and sampling points from the Pirkoh and Drazinda formations (C).
Notes: Geological map is simplified from Kazmi and Rana (Citation1982). PSB: Pellatispira beds (from Ali et al., Citation2018).
3. Material and methods
3.1. Material
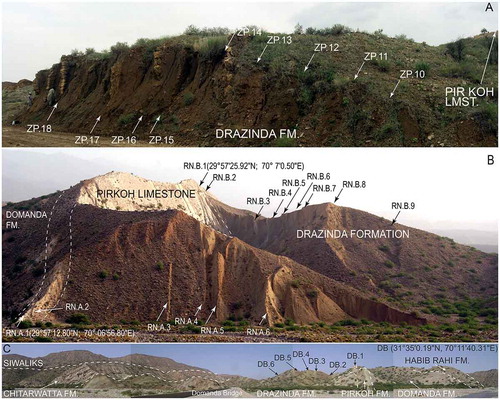
The samples of the Fulra Limestone were collected from three sections, near Kharai and Harudi villages, 100 km northwest of Bhuj, Kutch (23°30′55.97″N, 68°40′5.98″E; 23°28′45.32″N, 68°40′49.23″E) (Figures ). We have sampled 15 levels representing the unit. The Pirkoh and Drazinda formations have been sampled in three different outcrop areas, Rakhi Nala, Zinda Pir and Domanda Bridge, in the Sulaiman Range to provide the widest coverage of the orthophragminids (Figures ). The Rakhi Nala A 29°57′12.80″N, 70°06′56.80″E; 29°57′13.51″N, 70°7’1.74″E), and Rakhi Nala B (29°57′25.92″N, 70°7′0.50″E; 68°40′49.23″E, 29°57′16.10″N,70°7′11.20″E) sections are from the same area west of Dera Ghazi Khan in the Punjab province. The Zinda Pir section (30°20′2.38″N, 70°29′32.54″E; 30°19′56.65″N, 70°29′39.48″E), located in the Zinda Pir anticline, is ca. 55 km north-east of the Rakhi Nala section. The Domanda Section (31°35′0.19″N, 70°11′40.31″E; 31°35′14.43″N; 70°11′28.78″E) is located in the north-west of both Rakhi Nala and Zinda Pir sections, and is about 73 km west of Dera Ismail Khan in the Federally Administered Tribal Areas (FATA) (Ali et al., Citation2018).
Figure 4. Field aspects of Pirkoh Limestone and Drazinda Formation from Sulaiman Range in Pakistan and sampling points (from Ali et al., Citation2018).
3.2. Sample preparation
Specimens extracted from the shale, marl and limestone beds were studied for their external features, features in the equatorial layer and axial sections. The oriented sections of 507 megalospheric and 70 microspheric specimens (A and B-forms respectively) have been prepared through their equatorial layer. The morphometric measurements and counts were carried out on equatorial sections of the megalospheric specimens. Axial sections of 83 megalospheric and microspheric forms have been prepared for the comparisons in axial sections and also in order to facilitate specific recognition in rock thin sections.
4. Test features of the Tethyan orthophragminids
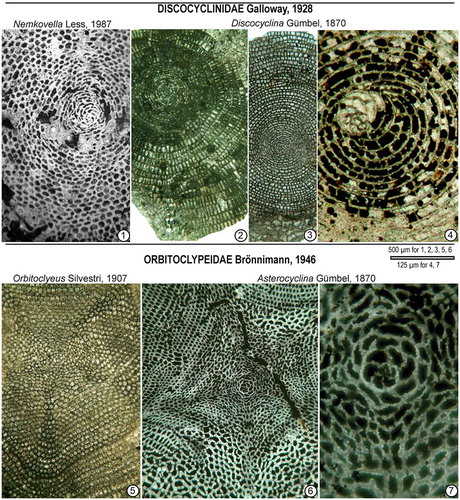
Orthophragminids are bilamellar, perforate foraminifera characterized by a discoidal, lenticular test with an equatorial layer consisting of cyclically arranged equatorial chambers and lateral layers composed of lateral chambers and pillars on either side of the equatorial layer ((A)). Externally, the test surface is either smooth, occasionally with an inflated or depressed central part, or it is characterized by radially developed ribs and bulges (Özcan et al., Citation2016b). Based on the relationship of protoconch and deuteroconch in equatorial sections, about ten configurations are used in the description of the embryon ((B)). The equatorial chambers are divided into chamberlets of different shapes as observed in equatorial sections. Tethyan orthophragminids consist of five genera; Discocyclina Gümbel, 1870, Nemkovella Less, Citation1987 and poorly known Asterophragmina Rao, 1942 placed in Discocyclinidae Galloway, 1928 and two genera, Orbitoclypeus Silvestri, 1907 and Asterocyclina Gümbel, 1870 placed in Orbitoclypeidae Brönnimann, 1946 (Ferràndez-Cañadell, Citation1998; Less, Citation1987) (). The obsolete genus Actinocyclina Gümbel 1870, characterized by the presence of numerous ribs, is assigned to genus Discocyclina after their discocyclinid-type microspheric juvenarium (Ferràndez-Cañadell, Citation1997; Less, Citation1987) (.1). The microspheric forms of some of the Tethyan genera are illustrated in .
Figure 5. General test features in Tethyan orthophragminid genera (A) (after Less, Citation1987; Ferràndez-Cañadell, Citation1997; Özcan et al., Citation2016b), qualitative parameters (B): a- types of embryon configurations, b- types of the adauxiliary chamberlets, c- different growth patterns of the equatorial annuli, d- types of granules and lateral chamberlets on the test surface, and parameters used in the morphometric description of orthophragminids as illustrated in D. pseudodispansa from Sulaiman Range (C). pac: principal auxiliary chamberlets.
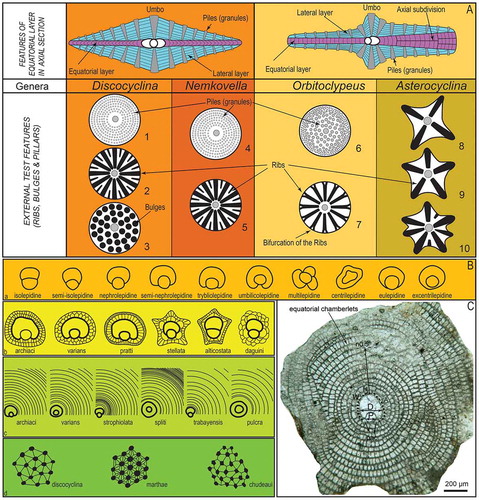
Figure 6. Microspheric juvenarium representative of some Tethyan orthophragminid genera (1–4: family Discocyclinidae Galloway, 5–7: family Orbitoclypeidae Brönnimann). 1: Nemkovella evae Less, Horsarrieu, SW France, early Cuisian. 2: Discocyclina nandori Less, (a species with ribs), Cambay Basin (W India), late middle Eocene. 3–4: Discocyclina archiaci (Schlumberger), Patala Formation (Thal, Pakistan), early Eocene. 5: Orbitoclypeus haynesi (Samanta & Lahiri, Citation1985), Fulra Limestone (Kutch, India), late middle Eocene. 6–7: Asterocyclina stellata (d’Archiac), Soğucak Formation (Thrace Basin, Turkey), middle-late Eocene transition.
5. The orthophragminids from the Fulra Limestone and Pirkoh and Drazinda formations
The LBF in Fulra Limestone are characterized by common occurrence of orthophragminids, nummulitids and alveolinids accompanied by rare rotaliids, and some stratigraphically important taxa as Dictyoconoides, Linderina, Calcarina, and smaller benthic foraminifera (Khanolkar et al., Citation2017; Samanta, Citation1993; Samanta & Lahiri, Citation1985; Samanta et al., Citation1990; Saraswati et al., Citation2000; Sen Gupta, Citation1963a, 1963b). Orthophragminids belong mainly to Discocyclina Gümbel, 1870, a single species of Orbitoclypeus Silvestri, 1907 and three species of Asterocyclina Gümbel, 1870 were also recognized. The following species are identified; Discocylina dispansa (Sowerby, Citation1840), Discocyclina pseudodispansa Özcan, Ali & Yücel, 2018, Discocyclina kutchensis Özcan & Saraswati, 2016, Discocyclina praeomphalus Samanta & Lahiri, Citation1985; Discocyclina augustae van der Weijden, Citation1940; Discocyclina discus (Rütimeyer, Citation1850), Discocyclina pratti (Michelin, Citation1846), Orbitoclypeus haynesi (Samanta & Lahiri, Citation1985), Asterocyclina sireli Özcan & Less, 2006, Asterocyclina alticostata (Nuttall, Citation1926), Asterocyclina stellata (d’Archiac, Citation1846) ((A)). The nummulitids, such as Nummulites beaumonti (.1–2), Nummulites sp. 1 (.3–4), Nummulites maculatus (.5–6), Nummulites gr. bullatus (.7–8), Operculina gr. gomezi (.9–10), Linderina sp. (.11–12), Calcarina sp. (.13–14), and Dictyoconoides sp. (.15). Operculina gr. gomezi, a diagnostic Bartonian-Priabonian species in Western Tethys (Less & Özcan, Citation2012), is identified for the first time from the Fulra Limestone.
Figure 7. Distribution of orthophragminids and associated LBF from the Fulra Limestone (A) and Pirkoh and Drazinda formations (B).
Notes: The synthetic stratigraphic columnar section representing Pirkoh and Drazinda formations in Sulaiman Range includes data from Rakhi Nala A and B, Zinda Pir, and Domanda Bridge sections.
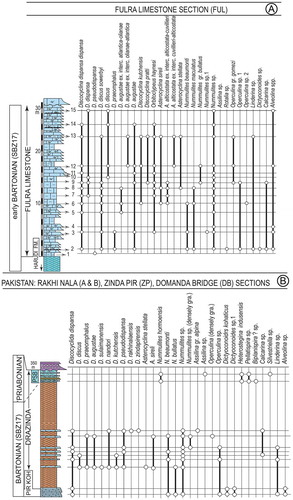
Figure 8. Common LBF associated with orthophragminids in the Fulra Limestone. 1–2: Nummulites beaumonti, 1: external view, FUL3–27, 2: equatorial section, FUL3–29. 3–4: Nummulites sp. 1, external view and equatorial section, FUL3–28. 5–6: Nummulites maculatus, external view and equatorial section, FUL2–33. 7–8: Nummulites gr. bullatus, external view and equatorial section, FUL9–2. 9–10: Operculina gr. gomezi, 9: external view, FUL9–5, 10: equatorial section, FUL12–16. 11–12: Linderina sp., external view and equatorial section, FUL2–32. 13–14: Calcarina sp., external view and equatorial section, FUL9–3. 15: Dictyoconoides sp., vertical section, FUL2–26. 16–26: Alveolina spp., 16–21: FUL15, 22: FUL4, 23–26: FUL12–62.

Orthophragminids in the Pirkoh and Drazinda formations are represented by Discocyclina and scanty Asterocyclina while Orbitoclypeus and Nemkovella are absent ((B)). The Pirkoh Formation yielded only discocyclinids, represented by D. dispansa (Sowerby, Citation1840), D. discus (Rütimeyer, Citation1850), D. praeomphalus Samanta & Lahiri, Citation1985 accompanied by Dictyoconoides, Nummulites, Assilina, Operculina, Linderina and alveolinids, found only in one level (sample ZP.2) in the Zinda Pir section. Orthophragminids in the Drazinda Formation are more diverse and are characterized by D. dispansa (Sowerby, Citation1840), D. discus (Rütimeyer, Citation1850), D. praeomphalus Samanta & Lahiri, 1985; D. augustae van der Weijden, 1940; ‘D.’ sulaimanensis Özcan, Ali & Hanif, 2016, D. kutchensis Özcan & Saraswati, 2016, D. nandori Less, 1987; D. pseudodispansa Özcan, Ali & Yücel, 2018, D. rakhinalaensis Özcan, Ali & Yücel, 2018, D. zindapirensis Özcan, Ali & Yücel, 2018, Asterocyclina sireli Özcan & Less, 2006, and A. stellata (d’Archiac, Citation1846). The associated LBF are represented by Nummulites sp., Assilina sp., Operculina sp., Calcarina sp., and Linderina sp. The Pellatispira-beds in the upper part of the Drazinda Formation contain scarce unidentified Discocyclina and other LBF, such as Pellatispira, reticulate Nummulites, Heterostegina, Silvestriella and Operculina (Ali et al., Citation2018).
6. Systematic paleontology
Detailed description of some of the orthophragminids from the Fulra Limestone was given by Sen Gupta (Citation1963a) and Samanta and Lahiri (Citation1985). We here focus on the general features important for the genus and morphometry of these taxa.
Order Foraminiferida Eichwald, 1830
Family Discocyclinidae Galloway, 1928
Genus Discocyclina Gümbel, 1870
Type-species: Orbitolites pratti Michelin, Citation1846
Discocyclina dispansa ( Citation Sowerby, 1840 )
(Figures (B), )
Figure 9. External test features of orthophragminids from the Fulra Limestone (B, F, I-N) and Drazinda Formation (A, C-E, G-H). A: Discocyclina discus, B: D. dispansa, C: D. pseudodispansa, D: D. sulaimanensis, E: D. zindapirensis, F: D. augustae, G–H: D. nandori, I: D. praeomphalus, J–K: D. kutchensis, L: Asterocyclina alticostata, M: A. sireli, N: Orbitoclypeus haynesi.

Figure 10. Equatorial and axial sections of D. dispansa dispansa from the Fulra Limestone. 1: sample FUL12–72, 2: FUL8–23, 3: FUL7–5, 4: FUL13–208.

1840 Lycophris dispansus n. sp., Sowerby, p. 327, pl. 24, Figs. 16, 16a–b.
1926 Discocyclina undulata sp. nov., Nuttall, p. 160–151, pl. 7, Figs. 8–9, pl. 8, Fig. 5.
2018 Discocyclina dispansa (Sowerby, Citation1840), Ali et al., Figs. 9, 12, 15, 17.
Diagnosis. Discocyclina dispansa is a small to large sized, ‘flat to saddle’ shaped, unribbed form. The small to medium-sized megalospheric embryon is semi-nephrolepidine in the earliest representatives of its lineage (e.g. D. d. broennimanni and D. d. taurica) and is trybliolepidine in the phylogenetically advanced members. The adauxiliary chamberlets are moderately wide and high, and of the ‘archiaci’ type. The equatorial chamberlets are also moderately wide and high. This species includes six subspecies in Western Tethys: D. d. broennimanni Less, Citation1987 (Dmean < 160 μm); D. d. taurica Less, Citation1987 (Dmean = 160–230 μm); D. d. hungarica Kecskeméti, 1959 (Dmean = 230–290 μm); D. d. sella (d’Archiac, Citation1850) (Dmean = 290–400 μm); D. d. dispansa (Sowerby, Citation1840) (Dmean = 400–520 μm); and D. d. umbilicata (Deprat, Citation1905) (Dmean > 520 μm) (Less, Citation1998).
Remarks. Discocyclina dispansa, one of the most common orthophragminid species in the Fulra Limestone, first described from this unit by Sowerby (Citation1840). In Fulra Limestone and Drazinda Formation, the species is represented by a flat test with a thick umbo, surrounded by a thin flange. In equatorial layer, a characteristic feature is the presence of the high early chambers followed by low chambers (.1–2). It appears that in the Western Tethys, saddle-shaped and flat forms exhibiting the ‘similar’ embryonic configuration and development of equatorial chambers appear to have been lumped under this species as evidenced by notable overlaps in the ranges of some subspecies such as D. dispansa hungarica and D. dispansa sella in the Lutetian and Bartonian (Zakrevskaya, Beniamovsky, Less, & Báldi-Beke, Citation2011), and D. dispansa sella and D. dispansa dispansa in the Bartonian (Less, Özcan, & Okay, Citation2011). We think that a revision of this group is necessary focusing on the distinction between the flat and saddle forms. Based on biometry (), the species is represented by D. d. dispansa (Sowerby, Citation1840) in the Fulra Limestone. This species may externally be confused with D. pseudodispansa.
Table 1. Statistical data of orthophragminids from the Fulra Limestone.
Discocyclina pseudodispansa Özcan, Ali & Yücel, 2018
(Figures (C), )
Figure 11. Equatorial and axial sections of D. pseudodispansa from the Fulra Limestone. 1: FUL1–3, 2: FUL1–9, 3: FUL1–7, 4: FUL1–17, 5: FUL1–19.
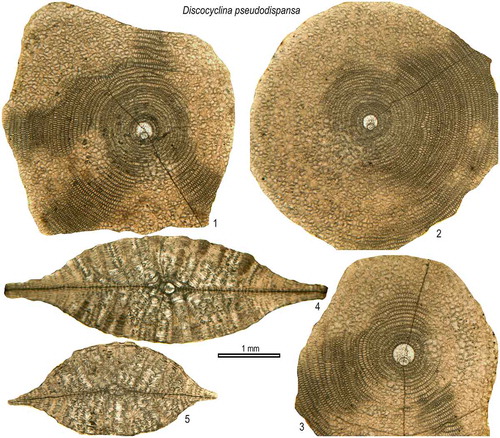
2018 Discocyclina pseudodispansa n. sp. Özcan, Ali, & Yücel, Ali et al., Figs. 9, 12, 15, 16.
Diagnosis. Flat test, small-to medium-sized, strongly inflated with a thick umbo. The small embryon consists of a rounded to nearly-rounded protoconch and larger deuteroconch displaying semi-nephrolepidine to trybliolepidine type configuration in equatorial sections. The annuli are typically narrow, presenting ‘archiaci’ type growth pattern. The piles are coarser in the umbonal part, compared to flange. Moderately high and wide lateral chamberlets are arranged in irregular rows.
Description.
External features. The test is typically flat, small to medium sized (1.35–6.8 mm in diameter) with a thick umbo, surrounded by a narrow, thin flange (). The thickness of the test varies between 600 and 2100 μm. The piles are relatively larger along the rims of the umbo (80–160 μm) compared to those in the flanges (50 μm), and are evenly distributed over the test surface. A detailed description of the species is given in Ali et al. (Citation2018).
Figure 12. Equatorial and axial sections of D. augustae ex. interc. olianae- atlantica from the Fulra Limestone. 4, microspheric; others, megalospheric. FUL13–170, 2: FUL13–171, 3: FUL13–41, 4: FUL7–3, 5: FUL13–102, 6: FUL13–209, 7: FUL13–213, 8: FUL.13–210.

Remarks. Discocyclina pseudodispansa, a common species in the Sulaiman Range, occurs only in the lowermost sample from Fulra Limestone. It could be easily confused with D. dispansa on the basis of external characters. The axial sections of both species are also similar to each other in terms of development of lateral chamberlets and their size (compare Figures 10.4, 11.4–5) (Ali et al., Citation2018). This species, however, differs from D. dispansa in having a much smaller embryon and tighter early chambers (n0.5 parameters in ). In equatorial sections, some of the specimens exhibiting semi-nephrolepidine type of embryon in the Sulaiman Range may be confused with D. augustae. The external features and axial sections of both species are totally different as the lateral chamberlets in D. augustae are very low (slit-like) and small in size.
Discocyclina augustae Citation van der Weijden, 1940
(Figures (F), )
1940 Discocyclina augustae n. sp. van der Weijden, p. 23−26, pl. 1, Figs. 4, 5, 7, 8; pl. 2, Figs. 1, 2, 11.
2018 Discocyclina augustae van der Weijden, Citation1940, Ali et al., Figs. 10–11, 12, 21.
Diagnosis. Discocyclina augustae is an unribbed form having a very small to small, semi-iso- to nephrolepidine embryon, narrow and low, ‘archiaci’ type adauxiliary chamberlets and also narrow and relatively low equatorial chamberlets mostly with ‘strophiolata’ type growth pattern. This species includes four subspecies in Western Tethys: D. a. sourbetensis Less, Citation1987 (Dmean < 145 μm); D. a. atlantica Less, Citation1987 (Dmean = 145–180 μm); D. a. olianae Almela & Rios, 1942 (Dmean = 180–225 μm); D. a. augustae van der Weijden, 1940 (Dmean > 225 μm).
Remarks. Discocyclina augustae is easily differentiated externally by its flat test with small central umbo and uniformly distributed piles ((F)). It can be, however, easily confused with D. pseudodispansa in equatorial sections, whereas their axial sections are completely different. This species has a much larger embryon than that of D. rakhinalaensis (Ali et al., Citation2018). In the studied material, D. augustae is represented by transitional stages between D. a. atlantica and D. a. olianae, according to the evolutionary scheme of the genus in the Western Tethys ().
Discocyclina praeomphalus Citation Samanta & Lahiri, 1985
(Figures (I), )
Figure 13. Equatorial and axial sections of D. praeomphalus from the Fulra Limestone. 8–9, microspheric; others, megalospheric. 1: FUL2–6, 2: FUL3–41, 3: FUL2–3, 4: FUL2–20, 5: FUL2–19, 6: FUL3–7, 7: FUL2–46, 8–9: FUL.3–8.

1985 Discocyclina praeomphalus n. sp. Samanta & Lahiri, p. 272–275. pl. 5, Figs. 1–6, text Figures 5–7, 12.
2018 Discocyclina praeomphalus, Ali et al., Figs. 10, 14, 23–24.
Diagnosis. Discocyclina praeomphalus is a medium sized (2.4–4 mm), saddle shaped, unribbed form with a slight depression at the center of the test. The megalospheric embryon (average 222.0–223.7 μm in diameter) exhibits semi-nephrolepidine to trybliolepidine type configuration. The adauxiliary chamberlets are moderately wide and high, and of the ‘archiaci’ type. The equatorial chamberlets are low and narrow in the early stage and are rectangular in shape in the later stages. The lateral chambers are typically low, numerous and not aligned in regular rows.
Remarks. This saddle-shaped, omphaloid species, first described from the Fulra Limestone (Samanta & Lahiri, Citation1985), occurs abundantly in the Drazinda Formation in the Sulaiman Range (Ali et al., Citation2018). It differs externally from other orthophragminids from the Drazinda Formation by its characteristic saddle shape and the central depression of the test. Discocyclina sulaimanensis, another omphaloid species from the Drazinda Formation, has a flat and smaller test than that of D. praeomphalus. The axial sections of D. praeomphalus are easily differentiated from that of the saddle-shaped D. discus in having very low (slit-like) chamberlets. This is a common discocyclinid in the middle Eocene of the Indian Subcontinent.
Discocyclina kutchensis Özcan & Saraswati, 2016
(Figures (J), (K), )
Figure 14. Equatorial and axial sections of D. kutchensis from the Fulra Limestone. 5, microspheric; others, megalospheric. 1: FUL6–51, 2: FUL6–131, 3: FUL6–132, 4: FUL13–145, 5: FUL6–59, 6: FUL6–123, 7: FUL6–127, 8: FUL6–102.
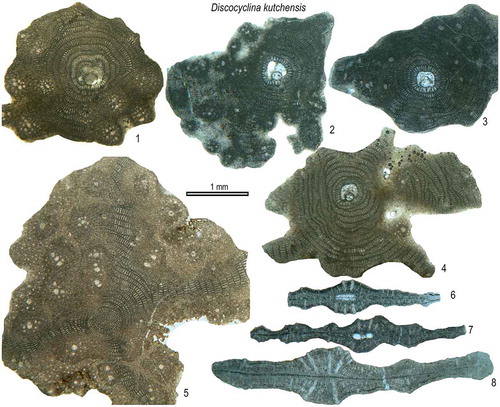
2016b Discocyclina kutchensis sp. nov. Özcan & Saraswati, p. 267–274, Figs. 5L, 6A–I, 7A, 8A–D, 9, 10A–D.
2018 Discocyclina kutchensis Özcan, Ali, & Yücel, Ali et al., Figs. 9, 11, 14, 22–23.
Diagnosis. Medium sized (2 to 6 mm), flat forms with numerous bulges. The bulges, uniformly distributed over the test surface, are semi-rounded and rounded in shape and range in size between 250 and 350 μm. The embryon is large, with an average diameter of the deuteroconch ranging between 410 and 478 μm, trybliolepidine to umbilicolepidine in configuration. The adauxiliary chamberlets (37–53 in number) are high and archiaci-type. The equatorial chambers are typically high, narrow and rectangular in shape.
Remarks. This species, originally described from the Fulra Limestone and Drazinda Formation, differs from most orthophragminids in Tethys by having the bulges uniformly distributed on the test surface. The bulges are semi-rounded to rounded, localized thickenings, homogenously distributed over the test surface (Özcan et al., Citation2016b). These structures are about 250–350 μm in diameter, 100–150 μm high and 100–300 μm apart from each other, and form an uneven test surface. Internally, the bulges consist of lateral chamberlets and coarse piles of up to 100 μm in diameter at its center, surrounded by smaller piles (25–50 μm in diameter), forming a circular pattern. These piles are semi-circular to polygonal in shape in transversal sections. The lateral surfaces of the bulges are rather sharp. The axial sections show no variation in the thickness of the equatorial layer adjacent to the bulges and inter-bulge areas. This implies that bulges are formed solely by the thickenings in the lateral layers (). The Bartonian ribbed orthophragminids in Indo-Pakistan region are represented by four species; Discocyclina nandori Less, Citation1987; Asterocyclina sireli Özcan & Less, 2006, A. alticostata (Nuttall, Citation1926) and rare A. stellata (d’Archiac, Citation1846). While they have different surface features, internally, D. nandori is differentiated from D. kutchensis by having a small, semi-nephrolepidine- to trybliolepidine type embryon and by low equatorial chamberlets (Özcan et al., Citation2016b). D. kutchensis is similar to D. radians, a Western Tethyan species, in having same type of embryon configuration, development of equatorial chambers and their shape. Both species are phylogenetically linked and D. kutchensis replaces D. radians in Eastern Tethys.
Discocyclina pratti
()
Figure 15. Equatorial sections of Discocyclina pratti from the Fulra Limestone. 1: FUL9–4, 2: FUL13–174, 3: FUL13–188, 4: FUL13–172, 5: FUL13–192.
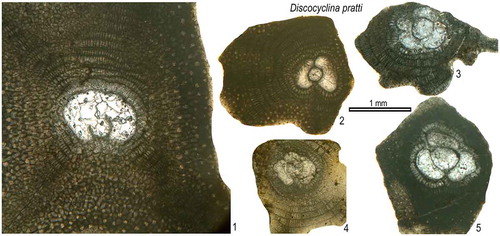
1846 Orbitolites pratti n. sp. Michelin, p. 278, pl. 63, Fig. 14.
Diagnosis. Discocyclina pratti is an unribbed species having a medium-sized to large, tryblio- to excentrilepidine-type embryon, numerous moderately wide and high, ‘pratti’ type adauxiliary chamberlets and narrow but high equatorial chamberlets with ‘pulcra’ type growth pattern. This species includes three subspecies: D. p. montfortensis Less, Citation1987 (Dmean < 510 μm), D. p. pratti (Michelin, Citation1846) (Dmean = 510–700 μm), D. p. minor Meffert, 1931 (Dmean > 700 μm) after the analysis of rather rich assemblages from Turkish sections (Özcan, Less, & Kertész, Citation2007).
Remarks. Some specimens with thin, fragile and undulated test revealed embryons ranging from 600 to 865 μm. The embryons display various embryonic configurations ranging from umbilicolepidine to centrilepidine where the wall of deutroconch may merge to the protoconchal wall at some points. The equatorial chambers are relatively high compared to those of D. discus. The embryon appears to be smaller than of D. discus. This species is rare in the studied material.
Discocyclina discus ( Citation Rütimeyer, 1850 )
((A), )
Figure 16. Equatorial and axial sections of D. discus sowerbyi from the Fulra Limestone. 1: FUL3–26, 2: FUL3–32, 3: FUL6–12, 4: FUL6–14, 5: FUL8–57, 6: FUL6–44, 7: FUL13–44, 8: FUL13–42, 9: FUL3–31.

1850 Orbitolites discus n. sp. Rütimeyer, p. 116, pl. 5, Figs. 70−71, 78, 80−81.
1926 Discocyclina sowerbyi nom. nov., Nuttall, p. 149−150, pl. 3, Figs. 1–3.
1985 Discocyclina adamsi n. sp., Samanta & Lahiri, p. 229−242, pl. 1, Figs. 6–10, pl. 5, Fig. 9, text –Figs. 5, 8.
2010 Lepidocyclina sp. (L. pustulosa), Matsumaru & Sarma, p. 550, pl. 3, Fig. 11.
2018 Discocyclina discus (Rutimeyer), Ali et al., Figs. 11, 14, 24–25.
Diagnosis. Discocyclina discus is a large, saddle-shaped and unribbed form with a giant, mostly umbilicolepidine (rarely also tryblio- or excentrilepidine) embryon, numerous, wide and high, ‘archiaci’ or transitional ‘archiaci-pratti’ type adauxiliary chamberlets and wide and high equatorial chamberlets with ‘archiaci’ or transitional ‘archiaci-pulcra’ type growth pattern. This species includes two subspecies in Tethys: D. d. discus (Rütimeyer, Citation1850) (Dmean < 1350 μm) and D. d. sowerbyi Nuttall, Citation1926 (Dmean > 1350 μm) after our present data from Fulra Limestone.
Remarks. Discocyclina discus, widely reported from the Fulra Limestone and the Drazinda Formation as D. sowerbyi, is distinguished from other orthophragminids by its thick, saddle-shaped test and its large embryon with irregular wall. D. praeomphalus, another saddle-shaped species, is much smaller and thinner than D. discus (compare the axial sections in Figures and ). In addition, the equatorial chambers of D. discus are much higher than those of D. praeomphalus. Our data show that the thick, saddle-like forms described as Discocyclina sowerbyi (Nuttall, Citation1926) by Nuttall (Citation1926) and Sen Gupta (Citation1963a) from the type locality of Lycophris ephippium of Sowerby (Citation1840) in the Kutch Basin, present the same morphological features as D. discus (Rütimeyer, Citation1850). Nuttall (Citation1926) proposed the specific name ‘sowerbyi’ to replace ‘ephippium’, which was preoccupied by another species. We maintain that D. sowerbyi, widely recorded from the middle Eocene of the Indian Subcontinent, is a junior synonym of D. discus. Discocyclina adamsi, established by Samanta and Lahiri (Citation1985) from the Fulra Limestone, is however, a junior synonym of D. sowerbyi, which was established from the same unit by Nuttall (Citation1926). We emend here the subdivision of D. discus lineage by differentiating two subspecies, D. discus discus and D. discus sowerbyi. This requires the replacement of ‘adamsi’ by ‘sowerbyi’. In Fulra Limestone the species belongs to D. discus sowerbyi. Matsumaru and Sarma (Citation2010) illustrated an axial section of a saddle-shaped specimen with a large and thick embryon from the late Eocene of Meghalaya (NE India) and identified it as a Caribbean Lepidocyclina sp. (L. pustulosa) without any illustration and description of the equatorial layer (Pl. 3, Fig. 11). This was interpreted to be the first record of the genus in Meghalaya and significant for the occurrence of Caribbean lepidocyclinids in the Eocene of Indian Subcontinent. In our opinion, the illustrated axial section has the same morphological features of D. discus in having a large embryon, high and wide lateral chamberlets and saddle-shaped test.
Family Orbitoclypeidae Brönnimann 1946
Genus Orbitoclypeus Silvestri, 1907
Orbitoclypeus haynesi ( Citation Samanta & Lahiri, 1985 )
((N), )
Figure 17. Equatorial and axial sections of Orbitoclypeus haynesi from the Fulra Limestone. 5–6, microspheric; others, megalospheric.1: FUL8–55, 2: FUL13–30, 3: FUL12–44, 4: FUL8–17, 5–6: FUL13–57, 7: FUL9–30.

1985 Discocyclina haynesi n. sp. Samanta & Lahiri, p. 262−272, pl. 4, Figs. 1−6; pl. 12, Figs. 9−14; text-Figures 5−7.
2010 Orbitoclypeus haynesi (Samanta & Lahiri), Özcan et al., Citation2010, p. 59−61, Fig. 29i−n.
2013 Orbitoclypeus haynesi (Samanta & Lahiri), Ben Ismail-Lattrache et al., 2013, p. 12, pl. 2, Fig. 9−15, fig. 8.
2017 Discocyclina dispansa Sowerby, BouDagher-Fadel and Price (Citation2017), Fig. 5−9.
Diagnosis. Orbitoclypeus haynesi is an unribbed species with ‘marthae’-type rosette, small eulepidine (rarely excentrilepidine) embryon, adauxiliary chamberlets of ‘varians’-type of average size and shape, moderately wide and high equatorial chamberlets arranged into strongly undulated annuli with ‘varians’-type growth pattern.
Remarks. Orbitoclypeus haynesi is the only orbitoclypeid in the Bartonian of Indian subcontinent. This species, referred to Discocyclina by Samanta and Lahiri (Citation1985), was later assigned to Orbitoclypeus based on the embryonic configuration, presence of undulated annuli and shape of the equatorial chamberlets (Özcan et al., Citation2010). We here show a microspheric form showing orbitoclypeid type juvenarium justifying the generic assignment (.5–6). Although similar to O. varians in external and internal test features, O. haynesi possesses a much smaller embryon, tighter annuli (parameter n0.5) and also commonly eulepidine-type embryonic configuration. Some specimens may exhibit excentrilepidine-type configuration. O. haynesi, also recorded from Turkey (Özcan et al., Citation2010) and northern margin of Africa (Ben Ismail-Lattrache et al., Citation2014) but not known from Europe, might be immigrant from the Indian subcontinent because of the expansion of the Eastern Tethyan fauna during Middle Eocene Climatic Optimum (MECO).
Genus Asterocyclina Gümbel, 1870
Type species Asterocyclina stellata ( Citation d’Archiac, 1846 )
Asterocyclina alticostata ( Citation Nuttall, 1926 )
(Figures (L), )
Figure 18. Equatorial and axial sections of A. alticostata ex. interc. alticostata-cuvillieri (2–5) and A. alticostata ex. interc. cuvillieri-alticostata (1) from the Fulra Limestone. 1: FUL13–9, 2: FUL12–11, 3: FUL12–5, 4: FUL6–24, 5: FUL6–30.
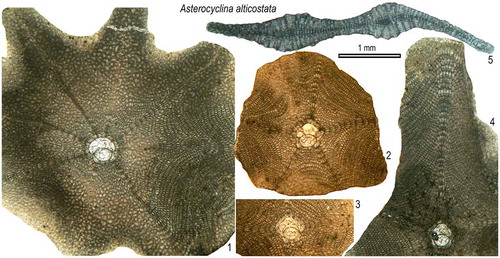
1926 Actinocyclina alticostata sp. nov., Nuttall, p. 151, pl. 8, Figs. 6−8.
Diagnosis. Asterocyclina alticostata is a star-shaped species usually with five to ten rays and ‘chudeaui’ type rosette. It has a medium-sized to relatively large isolepidine-type embryon, very few, very wide and moderately low, ‘alticostata’ type adauxiliary chamberlets and also wide and moderately high equatorial chamberlets arranged into asteroidal annuli with ‘strophiolata’ or ‘varians’ type growth pattern. This species includes four subspecies as; A. a. gallica Less, Citation1987 (Dmean < 275 μm); A. a. cuvillieri (Neumann, Citation1958) (Dmean = 275–350 μm); A. a. alticostata (Nuttall, Citation1926) (Dmean = 350–450 μm); A. a. danubica Less, Citation1987 (Dmean > 450 μm).
Remarks. Nuttall (Citation1926) erected this species from Kutch and noted the presence of about eight to twelve fairly wide prominent ribs in twelve specimens. He assigned these ribbed specimens to Actinocyclina (Gümbel) based on numerous ribs. We have collected a large number of specimens from the Fulra Limestone while this species is extremely rare in the Drazinda Formation. The number of ribs counted in fifty-four specimens from the Fulra Limestone is given in . This suggests that A. alticostata commonly develops more than five ribs, the maximum number being ten ribs in our material. Some of these ribs develop only in the adult stage of the test and they look like as if they are ‘secondary ribs’ on the test surface. The consistent occurrence of more than five ribs in this species in Indian Subcontinent as well as lower Bartonian Reineche Limestone in North Africa and coeval deposits in Turkey and Europe (Neumann, Citation1958; Köhler, Citation1967; Less, Citation1987; Less et al., Citation2011; Zakrevskaya et al., Citation2011; Ben Ismail-Lattrache et al., Citation2014) also points that this is not an ecological feature, but a specific character. The same feature is also observed in A. schweighauseri, a rare species recorded in the Cuisian and lower-middle Lutetian in Europe and Israel (Less, Citation1987, Citation1998). According to Less (Citation1987), always five rays start from the embryon of A. alticostata with further development of other rays in the adult stage (up to 10 rays), and many ribs start directly from the embryon in A. schweighauseri. We think that this is not a valid criterion for the distinction of these species since in some specimens of A. alticostata more than five rays may start to develop from the embryon (.1). In Fulra Limestone, this species is represented by the transitional developmental stages of A. alticostata cuvillieri and A. alticostata alticostata ().
Table 2. The variation in the number of ribs in A. alticostata (Nuttall) and A. sireli Özcan & Less.
Asterocyclina sireli Özcan & Less, 2006
(Figures (M), )
Figure 19. Equatorial and axial sections of A. sireli (1–11) and A. stellata (12–14) from the Fulra Limestone. 1: FUL13–71, 2: FUL13–152, 3: FUL13–92, 4: FUL6–66, 5: FUL6–107, 6: FUL6–135, 7: FUL8–2, 8: FUL6–138, 9: FUL6–136, 10: FUL8–1, 11: FUL8–3, 12: FUL8–20, 13–14: FUL13–153.
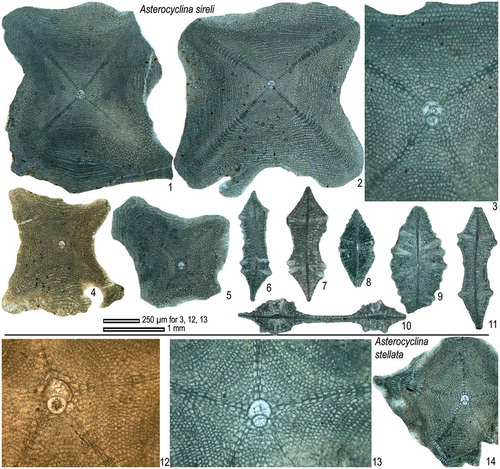
2006 Asterocyclina sireli n. sp., Özcan et al., p. 506−507, pl. 3, Fig. 32; pl. 4, Figs. 1−3; pl. 5, Figs. 1−5; text-Figure 12.
Diagnosis. Medium to large, flat forms with mostly four radial ribs and ‘marthae’ type rosette. The embryon is small, iso- to nephrolepidine. The deuteroconchal wall corresponding to the position of the successive stage of the developing rib is mostly depressed. The adauxiliary chamberlets are few (2–4) in number, low and moderately wide. The equatorial annuli are arranged usually in four rays.
Remarks. This species was first introduced from the Upper Lutetian of the Sivas Basin (Turkey) for asterocyclinid specimens displaying mostly four ribs and an embryon different from contemporaneous asterocyclinids in Western Tethys (Özcan, Less, Báldi-Beke, Kollányi, & Kertesz, Citation2006). It was later recorded from the Bartonian Reineche Limestone in Tunisia and the Fulra Limestone in India (Ben İsmail-Lattrache et al., Citation2014). In its type material (sample ALM.6) from Sivas, twenty-two specimens out of twenty-five have four ribs (). In lower Bartonian Reineche Limestone in Tunisia, only three specimens out of twenty-three specimens have five ribs. In the Fulra Limestone, only one out of seventy-nine specimens has five ribs and in the Drazinda Formation all specimens are with four ribs. The consistent occurrence of four ribs and the wide geographic range of this feature support that A. sireli is essentially a four-ribbed species. In contrast to its common occurrence in the Fulra Limestone, A. sireli is very rare in the Drazinda Formation.
Asterocyclina stellata ( Citation d’Archiac, 1846 )
()
Diagnosis. Asterocyclina stellata is a star-shaped form usually with five rays and ‘marthae’ type rosette. It has a small semi-iso- to nephrolepidine-type embryon, few, wide and low, ‘stellata’ type adauxiliary chamberlets and also narrow and low equatorial chamberlets arranged into asteroidal annuli with ‘strophiolata’ type growth pattern. This species includes four subspecies in Western Tethys: A. s. adourensis Less, Citation1987 (Dmean < 150 μm); A. s. stellata (d’Archiac, Citation1846) (Dmean = 150–190 μm); A. s. stellaris (Brünner, 1848 in Rütimeyer, Citation1850) (Dmean = 190–240 μm); A. s. buekkensis Less, Citation1987 (Dmean > 240 μm).
Remarks. A. stellata, recorded only from the Priabonian of Assam as A. matanzensis Cole by Samanta (Citation1965) from the Indian subcontinent, occurs sporadically in the Fulra Limestone. A subspecies designation is not possible because of the rare specimens in the studied material.
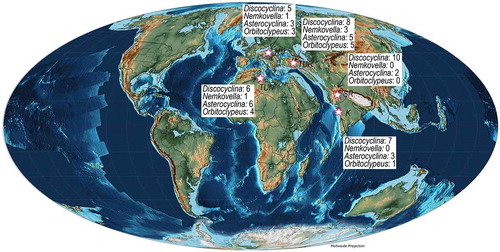
7. A synthesis of orthophragminids characterizing SBZ 17 in Tethys
The orthophragminids occurring in SBZ 17 in the first scheme of shallow benthic zonation (SBZ) in Tethys included thirteen species and Discocyclina pulcra baconica was marked as the key orthophragminid species for this zone (Serra-Kiel et al., Citation1998). This scheme was subsequently improved in Western Tethys as to include twenty-three species in SBZ 17 (Özcan et al., Citation2006, Citation2010; Less et al., Citation2011). Because of the recalibration of orthophragminid zones across the Lutetian-Bartonian boundary with respect to the boundaries of SBZ 17, distribution of these taxa has also changed with respect to SBZ zonation (Özcan et al., Citation2006). In updated scheme, stratigraphic range of no any subspecies corresponds to the full range of this zone. With the integration of six species confined to Indian subcontinent (species shown by yellow colour in ) and two species common to Indian subcontinent, Turkey and North Africa (A. sireli and O. haynesi), a total of twenty-nine species are now recorded in SBZ 17 in Tethys. In Indian subcontinent, this zone is characterized by the occurrence of fifteen species; Discocylina dispansa, Discocyclina pseudodispansa, Discocyclina kutchensis, Discocyclina pratti, Discocyclina praeomphalus, Discocyclina augustae, Discocyclina discus, Discocyclina sulaimanensis, Discocyclina rakhinalaensis, Discocyclina zindapirensis, Discocyclina nandori, Orbitoclypeus haynesi, Asterocyclina sireli, Asterocyclina stellata, and Asterocyclina alticostata. Among these only nine species (species shown by green colour in ) are common in Western Tethys and Indo-Pakistan region.
Figure 20. Updated scheme of orthophragminids showing the occurrence of twenty-nine species (lineages) in SBZ 17 (Bartonian) in Tethys. Yellow color shows those taxa confined to Indian subcontinent, green color shows taxa common to Turkey and Indian subcontinent and blue color shows taxa common to Turkey and Europe. Orbitoclypeus sp. 1 is a new orbitoclypeid from Baskil, Turkey, not yet formally described.
As opposed to Western Tethyan orthophragminids, the orthophragminid fauna in Eastern Tethys is dominantly characterized by genus Discocyclina (). The genus Orbitoclypeus, represented by a single species, O. haynesi, was only recorded from the Fulra Limestone and it is not present in the Sulaiman Range. The true Nemkovella does not occur in both regions. Nemkovella daguini, which displays a different peri-embryonic chamber arrangement than the true nemkovellids, occurs only in the Bartonian deposits of Meghalaya, India (unpublished data of E. Özcan). At present it is the only nemkovellid species in Eastern Tethys, where generic affiliation is also not certain. Asterocyclina sireli presents a wide geographic distribution in Tethys, from Turkey and Tunisia to the Indian subcontinent and probably its range extends at least to the western Pacific region. According to Less (Citation1987), the asterocyclinids in Western Pacific domain are characterized by the dominance of four-ribbed specimens. Less (Citation1987) considers that these four-ribbed asterocyclinids are of ‘Caribbean- Pacific affinity. Due to lack of detailed stratigraphic and taxonomic works from this area, a reliable correlation of A. sireli with Pacific Asterocyclina is presently not possible. It is highly probable that this species has migrated to Indian subcontinent and Western Tethys during the Middle Eocene Climatic Optimum that resulted in the expansion of Pacific fauna to higher latitudes in Bartonian. A similar migration scenario is also proposed here for O. haynesi, which occurs abundantly in Fulra Limestone, in North Africa and Turkey but not recorded in European platforms.
Figure 21. Orthophragminid species diversity in some Western to Eastern Tethys deposits corresponding to SBZ 17. K: Kutch, SR: Sulaiman Range, Pakistan, TUN: Tunisia, TUR: Turkey, HU: Hungary. The section in Hungary is from Dudar (sample DUDAR in Less, Citation1998). The combined sections from Turkey correspond to Gizliliman (Gizliliman section in Özcan et al., 2011), Çevirme (Çevirme section in Gül et al., Citation2012) and Baskil (Keçili section in Özcan et al., Citation2006). The section in Tunisia is from Damouss Quarry in Cap Bon Peninsula (Ben İsmail-Lattrache et al., Citation2014).
8. Conclusions
The Bartonian orthophragminids characterizing the SBZ 17 from the Fulra Limestone belong to Tethyan genera Discocyclina, Asterocyclina and Orbitoclypeus. Among a total of eleven species, seven of them belong to Discocyclina, one to Orbitoclypeus and three to Asterocyclina. Nemkovella does not occur in Indian subcontinent. Taking into account the twelve species from the Drazinda Formation, some are common to the Fulra Limestone, a total of fifteen species [Discocylina dispansa (Sowerby), Discocyclina pseudodispansa Özcan, Ali & Yücel, Discocyclina kutchensis Özcan & Saraswati, Discocyclina pratti (Michelin), Discocyclina praeomphalus Samanta & Lahiri, Discocyclina augustae van der Weijden, Discocyclina discus (Rütimeyer), Discocyclina sulaimanensis Özcan, Ali & Hanif, Discocyclina rakhinalaensis Özcan, Ali & Yücel, Discocyclina zindapirensis Özcan, Ali & Yücel, Discocyclina nandori Less, Orbitoclypeus haynesi (Samanta & Lahiri), Asterocyclina sireli Özcan & Less, Asterocyclina stellata (d’Archiac), and Asterocyclina alticostata (Nuttall)] are recognized from the Indian subcontinent. Six of them are confined only to Indo-Pakistan region (). Two species, A. sireli and O. haynesi, common in the Fulra Limestone and also occurring in North Africa (Tunisia) and Turkey but not in Europe, appear to have migrated to Western Tethys during Middle Eocene Climatic Optimum (MECO).
In addition to ribs, bulges, rounded surface structures corresponding to thickening of the lateral layers, are recognized in D. kutchensis, occurring abundantly in the Fulra Limestone as well as in the Drazinda Formation in Pakistan. This species is associated with D. nandori in the Drazinda Formation, while the latter species has not been recorded from the Fulra Limestone.
We recorded two common asterocyclinid species, A sireli and A. alticostata, and scanty A. stellata from the Fulra Limestone. A sireli and A. alticostata display the consistent characteristic features by developing four and many ribs respectively in the Tethys. This is a feature not observed in most of the asterocyclinid species, predominantly developing five ribs. Except for the above taxa and A. schweighauseri, a Cuisian to middle Lutetian species, other common asterocyclinid species, such as A. stellata and A. stella consistently develop mainly five ribs. Our specific examples from A. sireli and A. alticostata from geographically distant locations of Tethys suggest that certain asterocyclinid taxa consistently develop a specific number of ribs. Thus, the traditional concept of Asterocyclina with ‘five-ribs’ is to be abandoned.
We observed that the shape of the test (e.g. saddle-shaped vs flat tests) is a consistent character for each species. The saddle-shape test, for instance, is a specific feature of D. discus and D. praeomphalus. D. dispansa has always a flat test as D. augustae. This further requires re-evaluation of some D. dispansa populations in Western Tethys, which include both saddle and flat tests, interpreted to be ecological variations of the same species.
Funding
This work was supported by TUBA (Turkish Academy of Sciences); and INSA (Indian National Science Academy).
Disclosure statement
No potential conflict of interest was reported by the authors.
Acknowledgements
We thank Gyorgy Less (Miskolc) and an anonymous reviewer for helpful suggestions in the review of the manuscript. Fieldwork in India was possible due to funding by TUBA (Turkish Academy of Sciences) and INSA (Indian National Science Academy) Exchange of Scientists to E. Özcan and funding for laboratory work to P. K. Saraswati.
References
- Abbas, G. (1999). Microfacies, depositional environments, diagenesis and porosity development in Limestone horizons of Kirthar Formation (Middle-Late Eocene) in frontal parts of Sulaiman fold belt and adjoining areas, Pakistan ( Ph.D. thesis). University of Punjab, 112 pp.
- Afzal, J., Asrar, M. K., & Naseer, A. S. (1997). Biostratigraphy of the Kirthar Formation (Middle to Late Eocene), Sulaiman Basin, Pakistan. Pakistan Journal of Hydrocarbon Research, 9, 15–33.
- Ali, N., Özcan, E., Yücel, A. O., Hanif, M., Hashmi, S. I., Ullah, F., … Pignatti, J. (2018). Bartonian orthophragminids with new endemic species from the Pirkoh and Drazinda formations in the Sulaiman Range, Indus Basin, Pakistan. Geodinamica Acta, 30(1), 31–62.10.1080/09853111.2017.1419676
- d’Archiac, E. J. A. (1846). Description des fossiles recueillis par M. Thorent dans les couches à Nummulites des environs de Bayonne. Mémoires de la Société geologique de France, 2, 190–217.
- d’Archiac, E. J. A. (1850). Description des fossiles du groupe nummulitique recueillis par M. S.-P. Pratt et M. J. Delbos aux environs de Bayonne et de Dax. Mémoires de la Société géologique de France, 2, 397–456.
- Banerjee, S., Khanolkar, S., & Saraswati, P. K. (2017). Facies and depositional settings of the Middle Eocene-Oligocene carbonates in Kutch. Geodinamica Acta. doi:10.1080/09853111.2018.1442609
- Ben İsmail-Lattrache, K., Özcan, E., Boukhalfa, K., Saraswati, P. K., Soussi, M., & Jovane, L. (2014). Early Bartonian orthophragminids (foraminiferida) from Reineche Limestone, north African platform, Tunisia: Taxonomy and paleobiogeographic implications. Geodinamica Acta, 26, 94–121.
- Berggren, W. A., & Pearson, P. N. (2005). A revised tropical to subtropical paleogene planktonic foraminiferal zonation. Journal of Foraminiferal Research, 35, 279–298.10.2113/35.4.279
- Biswas, S. K. (1992). Tertiary stratigraphy of Kutch. Journal of the Palaeontological Society of India, 37, 1–29.
- Blanford, W. T. (1879). The Geology of Western Sind. Memoirs of the Geological Survey of India, 17, 1–210.
- BouDagher-Fadel, M. K., & Price, G. D. (2017). The paleogeographic evolution of the orthophragminids of the Paleogene. Journal of Foraminiferal Research, 47, 337–357.
- Chattoraj, S. L., Sarkar, U., Chakraborty, S., Banerjee, S., & Saraswati, P. K. (2012). Facies and palaeogeography of the Middle Eocene Fulra limestone, Kutch. Journal Indian Association Sedimentologists, 31, 1–9.
- Deprat, J. (1905). Les d.pots eocenes neo-cal.doniens. Bulletin de la Societé géologique de France, 4, 485–516.
- Eames, F. E. (1952a). A contribution to the study of the Eocene in western Pakistan and western India: A. The geology of standard sections in the western Punjab and in the Kohat district. Quarterly Journal of the Geological Society, 107, 159–171.
- Eames, F. E. (1952b). A contribution to the study of the Eocene in western Pakistan and western India: D. discussion of the faunas of the certain standard sections, and their bearing on the classification and correlation of the Eocene in western Pakistan and western India. Quarterly Journal of the Geological Society, 107, 173–200.
- Ferràndez-Cañadell, C. (1997). A new, ribbed species of Nemkovella 1987 (Discocyclinidae), and discussion on the genus Actinocyclina Gümbel, 1870. Journal of Foraminiferal Research, 27, 175–185.10.2113/gsjfr.27.3.175
- Ferràndez-Cañadell, C. (1998). Morphostructure and paleobiology of Mesogean orthophragminids (Discocyclininidae and Orbitoclypeidae, Foraminifera). Acta Geologica Hispanica, 31, 183–187.
- Gül, M. A., Ercan, A., Sinanli, M., Özkan, R., Alay, Z., Dinç, T., & Özcan, E. (2012). Eocene lithostratigraphy, biostratigraphy and geological evolution of northern part of internal Anotolian basins. Unpublished TPAO report, 308p ( In Turkish).
- Hemphill, W. R., & Kidwai, A. H. (1973). Stratigraphy of the Bannu and Dera Ismail Khan areas, Pakistan. United States Geological Survey Professional Paper, 716-B, 36p.
- Kazmi, A. H., & Rana, R. A. (1982). Tectonic map of Pakistan, 1: 2,000,000. Quetta: Geological Survey of Pakistan.
- Khanolkar, S., Saraswati, P. K., & Rogers, K. (2017). Ecology of foraminifera during the middle Eocene climatic optimum in Kutch, India. Geodinamica Acta. doi:10.1080/09853111.2017.1300846
- Köhler, E. (1967). Grossforaminiferen und Stratigraphie des Paläogens des Rajec− und Turiec−Kessels (Westkarpaten). Nauka o zemi, Geologica, 5, 1–87.
- Köthe, A., Khan, A. A., & Ashraf, M. (1988). Biostratigraphy of the Surghar Range, Salt Range, Sulaiman Range and the Kohat area, Pakistan, according to Jurassic through Paleogene calcareous nannofossils and Paleogene dinoflagellates. Geologisches Jahrbuch, B71, 1–87.
- Less, Gy. (1987). Paleontology and stratigraphy of the European Orthophragminae. Geologica Hungarica series Palaeontologica, 51, 1–373.
- Less, G. (1998). Zonation of the Mediterranean Upper Paleocene and Eocene by Orthophragminae. Opera Dela Slovenska Akademija Znanosti in Umetnosti (4), 34(2), 21–43.
- Less, Gy, & Özcan, E. (2012). Bartonian-Priabonian larger benthic foraminiferal events in the Western Tethys. Austrian Journal of Earth Sciences, 105(1), 129–140.
- Less, Gy, Özcan, E., & Okay, A. I. (2011). Stratigraphy and larger foraminifera of the Middle Eocene to Lower Oligocene shallow-marine units in the northern and eastern parts of the Thrace Basin, NW Turkey. Turkish Journal of Earth Sciences, 20, 793–845.
- Matsumaru, K., & Sarma, A. (2010). Larger foraminiferal biostratigraphy of the lower Tertiary of Jaintia Hills, Meghalaya, NE India. Micropaleontology, 56, 539–565.
- Michelin, H. (1846). Iconographie zoöphytologique. Paris: P. Bertrand.
- Nagappa, Y. (1959). Foraminiferal biostratigraphy of the Cretaceous- Eocene succession in the India-Pakistan-Burma region. Micropaleontology, 5, 145–192.10.2307/1484208
- Neumann, M. (1958). Révision des Orbitoididés du Crétacé et de l’Eocène en Aquitaine occidentale. Mémoires de la Societé géologique de France (n. sér.), 37, 1–174.
- Nuttall, W. L. F. (1926). The zonal distribution and description of the larger foraminifera of the Middle and Lower Kirthar series (Middle Eocene) of parts of Western India. Records of the Geological Survey of India, 59, 115–164.
- Özcan, E., Less, Gy, Báldi-Beke, M., Kollányi, K., & Kertesz, B. (2006). Biometric analysis of middle and upper Eocene Discocyclinidae and Orbitoclypeidae (Foraminifera) from Turkey and updated orthophragmine zonation in the Western Tethys. Micropaleontology, 52, 485–520.
- Özcan, E., Less, Gy, & Kertész, B. (2007). Late Ypresian to Middle Lutetian orthophragminid record from central and northern Turkey: Taxonomy and remarks on zonal scheme. Turkish Journal of Earth Sciences, 16, 281–318.
- Özcan, E., Less, G., Okay, A. I., Báldi-Beke, M., Kollányi, K., & Yilmaz, I. Ö. (2010). Stratigraphy and Larger Foraminifera of the Eocene Shallow-marine and Olistostromal Units of the Southern Part of the Thrace Basin, NW Turkey. Turkish Journal of Earth Sciences, 19, 27–77.
- Özcan, E., Ali, N., Hanif, M., Hashmi, S. I., Khan, A., Yücel, A. O., & Abbasi, I. A. (2016a). New Priabonian Heterostegina from the Eastern Tethys (Sulaiman fold belt, West Pakistan): Implications for the development of Eastern Tethyan heterostegines and their paleobiogeography. Journal of Foraminiferal Research, 46(4), 393–408.
- Özcan, E., Saraswati, P. K., Hanif, M., & Ali, N. (2016b). Orthophragminids with new axial thickening structures from the Bartonian of the Indian subcontinent. Geologica Acta, 14(3), 261–282.
- Rütimeyer, L. (1850). Ueber das schweizerische Nummulitenterrain mit besonderer Berücksichtigung des Gebirges zwischen den Thunersee und der Emme. Denkschriften der Schweizerischen Naturforschende Gesellschaft, 11, 1–120.
- Samanta, B. K. (1965). Asterocyclina from the Upper Eocene of Assam, India. Contributions from the Cushman Foundation for Foraminiferal Research, 16, 22–27.
- Samanta, B. K. (1970). Middle Eocene planktonic foraminifera from Lakhpat, Cutch, western India. Micropaleontology, 16, 185–215.10.2307/1485114
- Samanta, B. K. (1973). Planktonic foraminifera from the Palaeocene-Eocene succession in the Rakhi Nala, Sulaiman Range, Pakistan. Bulletin of the British Museum (Natural History), 22, 421–482.
- Samanta, B. K. (1993). Foraminiferal genus Alveolina d’Orbigny from the Middle Eocene Fulra Limestone of Kutch, Gujarat, Western India with observations on the distribution of the genus in the Indo-Pacific region. The Geological, Mining & Metallurgical Society of India, 57, 1–92.
- Samanta, B. K., & Lahiri, A. (1985). The occurrence of Discocyclina Gümbel in the middle Eocene Fulra limestone of Cutch, Gujarat, Western India, with notes on species reported from the Indian region. Bulletin Geological Mining Metallurgical Society of India, 52, 211–295.
- Samanta, B. K., Bandopadhyay, K. P., & Lahiri, A. (1990). The occurrence of Nummulites Lamarck (Foraminiferida) in the Middle Eocene Harudi Formation and Fulra Limestone of Cutch, Gujarat, Western India. Bulletin Geological Mining Metallurgical Society of India, 55, 1–66.
- Saraswati, P. K., Patra, P. K., & Banerjee, R. K. (2000). Biometric study of some Eocene Nummulites from Kutch and Jaisalmer, India. Journal of Palaeontological Society of India, 45, 91–122.
- Saraswati, P. K., Khanolkar, S., Raju, D. S. N., Dutta, S., & Banerjee, S. (2014). Foraminiferal biostratigraphy of lignite mines of Kutch, India: Age of lignite and fossil vertebrates. Journal of Palaeogeography, 3(1), 90–98.
- Saraswati, P. K., Khanolkar, S., & Banerjee, S. (2017). Paleogene stratigraphy of Kutch, India: An update about the progress in foraminiferal biostratigraphy. Geodinamica Acta. doi:10.1080/09853111.2017.1408263
- Sen Gupta, B. K. (1963a). A restudy of two common species of discocyclina from India. Micropaleontology, 9, 39–49.10.2307/1484604
- Sen Gupta, B. K. (1963b). Structure of Asterocyclina alticostata (Nuttall). Contributions from the Cushman Foundation for Foraminiferal Research, 14, 95–98.
- Serra-Kiel, J., Hottinger, L., Caus, E., Drobne, K., Ferràndez, C., Jauhri, A. K., … Zakrevskaya, E. (1998). Larger foraminiferal biostratigraphy of the Tethyan Paleocene and Eocene. Bulletin de la Societé géologique de France, 169, 281–299.
- Shah, S. M. I. (2009). Stratigraphy of Pakistan. Geological Survey of Pakistan Memoirs, 22, 381 pp.
- Sowerby, J. de C. (1840). Systematic list of organic remains. Appendix to: Grant, C.W., Memoir to illustrate a geological map of Cutch. Transactions of the Geological Society of London, 5(2), 317–329.
- Warraich, M. Y., & Nishi, H. (2003). Eocene planktic foraminiferal biostratigraphy of the Sulaiman Range, Indus Basin, Pakistan. Journal of Foraminiferal Research, 33(3), 219–236.10.2113/33.3.219
- van der Weijden, W. J. M. (1940). Het genus Discocyclina in Europa. Een monografie naar aanleiding van een heronderzoek van het Tertiair-profiel van Biarritz ( Thesis), Rijkuniversiteit Leiden, 116p.
- Zakrevskaya, E., Beniamovsky, V., Less, Gy, & Báldi-Beke, M. (2011). Integrated biostratigraphy of Eocene deposits in the Gubs section (Northern Caucasus) with special attention to the Ypresian/Lutetian boundary and to the Peritethyan-Tethyan correlation. Turkish Journal of Earth Sciences, 20, 753–792.