
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Traditionally, population models distinguish individuals on the basis of their current state. Given a distribution, a discrete time model then specifies (precisely in deterministic models, probabilistically in stochastic models) the population distribution at the next time point. The renewal equation alternative concentrates on newborn individuals and the model specifies the production of offspring as a function of age. This has two advantages: (i) as a rule, there are far fewer birth states than individual states in general, so the dimension is often low; (ii) it relates seamlessly to the next-generation matrix and the basic reproduction number. Here we start from the renewal equation for the births and use results of Feller and Thieme to characterize the asymptotic large time behaviour. Next we explicitly elaborate the relationship between the two bookkeeping schemes. This allows us to transfer the characterization of the large time behaviour to traditional structured-population models.
Mathematics Subject Classification:
1. Introduction
In the pioneering paper [Citation7], Jim Cushing and Zhou Yicang
defined the net reproductive number (aka the basic reproduction number and in the present paper denoted by
) in the context of linear discrete time population models, while highlighting its interpretation as the expected lifetime number of offspring,
showed that the stability of the extinction state (i.e. the trivial steady state) is governed by the sign of
demonstrated, by way of examples, that often
By making systematic use of Perron-Frobenius theory, Chi-Kwong Li and Hans Schneider streamlined the proofs and extended these results in [Citation22]. For far-reaching generalizations we refer to recent papers of Horst Thieme, in particular [Citation28, Citation29]. And for similar continuous time results in the context of epidemic models to [Citation9, Citation14, Citation15, Citation27]. Also see [Citation6].
Over the years (and in close collaboration with various other authors) we have developed theory for continuous time structured-population models [Citation10–12, Citation24]. These papers deal with rather general models and may, as a consequence, not be easily accessible. The aim of the present paper is to formulate the discrete time version of the linear theory in a reader-friendly fashion. To this end, we limit our attention to population models in which individuals are distinguished by a number of traits (e.g. species, age, size, spatial location, infection status), but such that the set of conceivable individual states (shortly, i-states) is finite.
In our approach, the renewal equation (RE) for the birth rate takes centre stage. Once one solves the RE constructively, all other relevant quantities are given by explicit expressions in terms of the initial condition and the birth rate. As a consequence, we can deduce the asymptotic large time behaviour of every quantity of interest from the asymptotic large time behaviour of the birth rate. And to determine the latter, we can rely on Feller's celebrated Renewal Theorem for scalar RE and on Thieme's generalization for systems of RE (see Section XIII.10 in [Citation16] and [Citation25]; continuous time results can be found in [Citation4, Citation8, Citation17]).
The key feature that makes this approach both attractive and efficient is that, as a rule, there are far fewer individual birth states than individual states in general. In particular, the RE is a scalar equation when all individuals are identical at birth, either in the literal sense (like, for instance, in the case of age as the only structuring trait) or in the stochastic sense, when states-at-birth have a fixed probability distribution (for example, in an epidemiological model where individuals begin their infected life (i.e. are born epidemiologically) as symptomatic with probability p or asymptomatic with probability 1−p, regardless of the i-state of the individual that infected them).
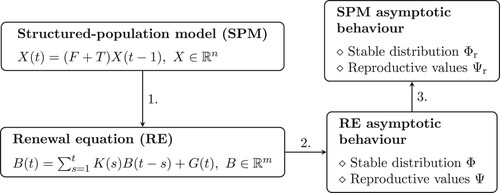
We begin by preparing the ground: in Section 2, we formulate the RE for the birth rate and determine the asymptotic large time behaviour using the theorems of William Feller (for scalar RE) and Horst Thieme (for systems of RE). In Section 3 we first show how classical discrete time structured-population models (SPM) give rise to (typically much smaller) systems of RE for the birth rate and discuss how the two bookkeeping schemes relate to each other in terms of dynamics and next-generation matrices. Next we show how, after having deduced the asymptotic behaviour of solutions of the RE using the results of Section 2, one can determine the asymptotic i-state distribution and the reproductive values in SPM from the ones obtained in the corresponding RE (see Figure ).
Figure 1. Deducing the asymptotic large time behaviour in (linear discrete time) structured-population models via the corresponding renewal equation for the birth rate.
In Section 6 of [Citation10] and Section 3.2 of [Citation12] it was shown that, for the special situation that , not only the spectral equivalence
holds, but that one can also compute the asymptotic distribution (i.e. the right eigenvector) in the real time setting from that in the generation bookkeeping framework. In Section 4 we supplement this result with a corresponding result for the reproductive values (or, in other words, for the left eigenvector).
In [Citation12] it was demonstrated that for a large class of nonlinear models the condition arises when looking for steady states. This makes the results about connecting the eigenvectors useful for nonlinear models as well.
In Appendix we present a brief outline of continuous time analogues of various equations and identities derived in Sections 2, 3 and 4.
2. The renewal equation
The first step in the formulation of renewal equations is the identification of states-at-birth. We may interpret this literally in the sense of determining all the i-states in which individuals may begin their lives. For example, think of a model where individuals are structured by age a and size s and where the set of i-states is Assuming that newborns may be either small or large, there are two states-at-birth:
and
However, when small and large offspring is produced at a fixed ratio
(independently of the i-state of the parent) we can exploit the fact that there is a single state-at-birth in the stochastic sense: all individuals are born with the same probability distribution of their i-state. Another example is an epidemiological model where individuals may begin their infected life (i.e. are born epidemiologically) as either asymptomatic or symptomatic. There are again two states-at-birth in the literal sense, but when new cases arise at a fixed ratio there is but one state-at-birth in the stochastic sense.
Both interpretations of the term state-at-birth are valid. However, while the literal interpretation may be a natural starting point, understanding the term in the stochastic sense allows us to formulate RE systems of minimal size (we elaborate this point in Section 3).
We begin by considering the case where all individuals are identical at birth, i.e. are born with the same probability distribution of their i-state.
2.1. The scalar equation
Imagine that immediately after the yearly breeding season a census is made of the subpopulation of newborn females. Let denote that number in year t. If we ignore density dependence and adopt a deterministic point of view at the population level (i.e. we assume that the numbers are so large that it makes sense to focus on expected values and to ignore the effects of demographic stochasticity) then
(1)
(1) where
is the expected number of daughters born to a female in year s after her own birth. We consider the sequence
as the basic model ingredient and make the following assumptions:
| (A1) |
| ||||
| (A2) |
| ||||
Note that
(2)
(2) is the basic reproduction number, i.e. the expected lifetime number of (female) offspring.
Equation (Equation1(1)
(1) ) is linear and translation invariant, so we expect it to have geometric solutions. If we substitute the Ansatz
(3)
(3) into (Equation1
(1)
(1) ) and divide both sides by
we obtain the Euler–Lotka characteristic equation
(4)
(4) for the unknown z. Note that
is the (one-sided) z-transform of
(with
) and that the z-transform is the discrete time analogue of the Laplace transform.
For real z, is a strictly decreasing function with limit zero at infinity. So if
takes, for a real z, a value larger than one, Equation (Equation4
(4)
(4) ) has precisely one real solution. We denote this solution by ρ and observe that there are precisely three possibilities:
whenever
for a value
(in the degenerate case that only
we have
). To see this, note that
if
if
If z is complex then
Consequently the solutions of (Equation4
(4)
(4) ) are contained within the closed disc of radius ρ.
We say that is periodic with period p>1 if
except when s is an integer multiple of p. One easily verifies that in that case z is a solution of (Equation4
(4)
(4) ) if z is the product of ρ and a pth root of unity. So for periodic
there are non-real solutions of (Equation4
(4)
(4) ) on the circle of radius ρ in
. We make a further assumption:
| (A3) |
| ||||
So far we focused on special solutions of (Equation1(1)
(1) ) of the form (Equation3
(3)
(3) ). Now, in the spirit of [Citation13], we prescribe the history of b up to t = 0 by putting
(5)
(5) for a given function
such that
(6)
(6) is finite for
. This allows us to rewrite (Equation1
(1)
(1) ) in the form
(7)
(7) with the convolution product interpreted as zero for t = 0, i.e.
(8)
(8) Let
and
denote the (one-sided) z-transforms of, respectively,
and
, i.e.
The z-transform has the useful property that it turns a convolution product into an ordinary product. So in terms of the z-transforms, the convolution equation (Equation7
(7)
(7) ) can be written as
If follows that
and next, by taking the inverse z-transform, that
where the closed contour encircles the origin counter-clockwise at a distance great enough to enclose all singularities of the integrand. The observations made above concerning the solutions of (Equation4
(4)
(4) ) now suggest that
for
. Theorem 2.1 in Section XIII.10 of [Citation16] gives a far more informative characterization of the asymptotic large time behaviour of b.
Theorem 2.1
Feller's Renewal Theorem
Consider the renewal Equation (Equation7(7)
(7) ) and assume that (A1), (A2) and (A3) hold. Furthermore, assume that
for all
and that
. For
let
denote the unique real solution of the Euler-Lotka Equation (Equation4
(4)
(4) ).
| (i) | If | ||||
| (ii) | If | ||||
| (iii) | If | ||||
Remark 2.2
To facilitate comparison with Feller's formulation in [Citation16], we present the notation-translation table:
More importantly, note that Feller uses the generating function rather than the z-transform. IfIf (Equation6(6)
(6) ) holds we deduce from the second identity that
(9a)
(9a) with
(9b)
(9b) The coefficients
thus tell us how the various components of the initial condition (Equation5
(5)
(5) ) contribute to the ultimate geometric population growth with multiplication factor ρ when
(for the time being, we do not normalize these contributions).
We conclude that Theorem 2.1 gives a complete description of the large time behaviour of solutions of the renewal Equation (Equation7(7)
(7) ) when we prescribe as an initial condition the history of b up to time zero as in (Equation5
(5)
(5) ).
2.2. Systems of renewal equations
Suppose that individuals begin their lives in one of m>1 states-at-birth. The renewal equation now reads
(10)
(10) where the components
of the vector
denote the number of females born at time t with state-at-birth j and
is a positive
matrix with elements
Remark 2.3
A matrix A is called positive if for all i and j. In this case, we write
. We write A>0 when
and
. A matrix A is called strictly positive if
for all i and j. In this case, we write
. In Appendix we use that a matrix A is called positive-off-diagonal if
for
and
[Citation1].
The sequence of matrices (with
) forms a kernel. We assume
| (A4) |
| ||||
| (A5) |
| ||||
Remark 2.4
In a population dynamical context, it is preferable to equip with the
-norm, i.e. to define
In A5, we then use the corresponding operator norm,
If we now look for solutions of the RE (Equation10(10)
(10) ) in the form
for some m-vector Φ we observe that the Euler-Lotka equation takes the form
(11)
(11) where
denotes the spectral radius and where, as before,
(12)
(12) We call
the next-generation matrix and define
(13)
(13) Note (by exploiting the fact that the spectral radius of
is a decreasing function of z) that the Euler-Lotka Equation (Equation11
(11)
(11) ) has a unique real solution ρ when
is larger than or equal to one. In that case, we necessarily have
or
Furthermore,
and
are positive matrices, and therefore for both matrices the spectral radius is a dominant eigenvalue with a positive corresponding eigenvector [Citation1, Citation2].
The key reference concerning the asymptotic behaviour of the solutions of the renewal equation (Equation10(10)
(10) ) is now [Citation25]. In order to present Thieme's renewal theorem, we again prescribe the history of B up to t = 0,
(14)
(14) for given positive
such that
(15)
(15) defines a vector with finite components for all
. This allows us to write (Equation10
(10)
(10) ) as
(16)
(16) with the convention
(17)
(17) Next, we introduce the resolvent of the kernel K. The resolvent
is defined by the equation
where * denotes the convolution product, i.e.
(18)
(18)
In our case and consequently
, meaning that we can write (Equation18
(18)
(18) ) as
The results in [Citation25] concern a rather general setting in terms of positive linear operators on ordered Banach spaces. When dealing with spaces of functions defined on non-compact domains, it often helps to define positivity in terms of a special element w of the positive cone, with w possibly tending to zero for the argument tending to infinity. Here, however, we only deal with the standard positive cone in
and when invoking the results of [Citation25] we simply take the m-vector
.
We make further assumptions
| (A6) | There exists | ||||
| (A7) | There exist | ||||
Theorem 2.5
Thieme's Renewal Theorem
Consider the renewal equation (Equation16(16)
(16) ) and assume that (A4), (A5), (A6) and (A7) hold. Furthermore, assume that there exists M>0 such that
for all
. Assume that
. The Euler–Lotka equation (Equation11
(11)
(11) ) has a unique real solution ρ with either
or
. Furthermore let Φ, with
and Ψ, with
, denote, respectively, the right and the left normalized positive eigenvector of
corresponding to eigenvalue 1.
Then for
(19a)
(19a) with
(19b)
(19b)
Remark 2.6
The following table aims to facilitate comparison with Thieme's results in [Citation25]:
Table
Note that one can normalize Ψ in any way one wants, since in (Equation19b
3. Structured-population models
When density dependence is ignored and the set of i-states is finite, a discrete time structured-population model leads to the system of linear recursion relations
(20)
(20) Here the components
of the vector
correspond to the density of females with i-state j at time t, while F and T are positive
matrices that describe, respectively, reproduction and survival
development, cf. [Citation3, Citation5]. More precisely,
and
where
and
Since no individual is immortal we assume that
.
The aim of this section is twofold: (i) we show how the recurrence (Equation20(20)
(20) ) and the renewal equation (Equation10
(10)
(10) ) relate to each other and (ii) we discuss how the next-generation matrix and the basic reproduction number corresponding to (Equation20
(20)
(20) ) are connected with (Equation13
(13)
(13) ). We begin with the former.
Suppose there are m states-at-birth in the literal sense (). There exist a positive
matrix V (normalized such that all m columns of V have
-norm equal to one) and a positive
matrix U such that
(21)
(21) In particular, if the states-at-birth are
we define V as the matrix with columns
and U as the fertility matrix obtained by taking the rows
from F.
Directly from the interpretation we discover the first relation, namely,
(22)
(22) Using the generation expansion we then deduce from (Equation20
(20)
(20) ), (Equation21
(21)
(21) ) and (Equation22
(22)
(22) ) that
leading in the limit to the second relation
(23)
(23) The two Equations (Equation22
(22)
(22) ) and (Equation23
(23)
(23) ) can both be understood on the basis of their interpretation and together they provide a complete description of the dynamics. By substituting (Equation23
(23)
(23) ) into (Equation22
(22)
(22) ) one obtains (Equation10
(10)
(10) ) with
(24)
(24) Likewise one recovers (Equation20
(20)
(20) ) from (Equation22
(22)
(22) ) and (Equation23
(23)
(23) ) by splitting off the
term in (Equation23
(23)
(23) ) to obtain
(25)
(25) and next use (Equation22
(22)
(22) ) and (Equation21
(21)
(21) ).
The two Equations (Equation22(22)
(22) ) and (Equation23
(23)
(23) ) relate the time course of B to that of X and vice versa, when defined for all t, not just for
. To deal with the initial value problem for X, we can rewrite (Equation20
(20)
(20) ) in the variation-of-constants form
(26)
(26) By acting on both sides of this identity with F we obtain
(27)
(27) where
Once we solve the RE (Equation27
(27)
(27) ), we can rewrite (Equation26
(26)
(26) ) as the explicit expression
(28)
(28) Here Y takes values in
, but whenever F has the form (Equation21
(21)
(21) ), we know that
(29)
(29) for a function B taking values in
. We can then rewrite (Equation27
(27)
(27) ) in the form
(30)
(30) that is, we get (Equation16
(16)
(16) ) with
(31a)
(31a)
(31b)
(31b) Note that when the rank of U is not maximal, there are fewer states-at-birth in the stochastic sense than there are in the literal sense. In such a case, a further reduction in RE system size is possible with a different choice of U and V (see one such example below).
Hence, if the conditions of Theorem 2.5 (or Theorem 2.1 in the case where (Equation30(30)
(30) ) amounts to a scalar equation) are satisfied, we can first deduce the asymptotic behaviour of
from that theorem and then use (Equation28
(28)
(28) ), with Y given by (Equation29
(29)
(29) ), to determine the large time behaviour of the population state
. We work out the details of this procedure in the two subsections that follow. But first, we discuss how the next-generation matrix and the basic reproduction number corresponding to (Equation20
(20)
(20) ) are connected with (Equation13
(13)
(13) ).
For , the
th element of
is the probability that an individual with i-state j is alive t units of time later and has then i-state i. If
then
exists and the
th element of
equals the expected time an individual starting with i-state j will spend in i-state i.
The matrix
(32)
(32) is the next-generation matrix (NGM) with large domain, with the
th element of
specifying the expected number of future offspring with i-state i produced by an individual presently having i-state j. Often one defines
(33)
(33) The elements of the NGM with large domain (Equation32
(32)
(32) ) specify the expected numbers of future offspring of individuals for all i-states. But, with the factorization of F as described directly after (Equation21
(21)
(21) ), we can focus on the true NGM
(34)
(34) We invite the reader to verify that the
th element of
gives the expected lifetime number of offspring with state-at-birth i produced by a newborn individual with state-at-birth j.
Note that when the rank of U is smaller than m (which happens whenever there are fewer states-at-birth in the stochastic sense than there are in the literal sense), a further reduction to the NGM with small domain is possible by altering U and V (see one example below and also [Citation14] for the derivation of NGM with small and large domains in the context of epidemiological models in continuous time).
One easily verifies that
the NGM with large domain (Equation32
If Φ is a right eigenvector of
If
Furthermore, when we deduce from (Equation12
(12)
(12) ) and (Equation31a
(31a)
(31a) ) that
(36)
(36) That is, when U and V are defined as described after (Equation21
(21)
(21) ) then
is the NGM and we recover (Equation13
(13)
(13) ).
3.1. One state-at-birth
The simplest situation arises when there is only one state-at-birth, either in the deterministic sense where all individuals are identical at birth (e.g. when age is the only structuring variable) or in the stochastic sense where states-at-birth have a given probability distribution. In that case F has one dimensional range.
Then F = V U, where (with
) spans the range of F and U corresponds to the row n-vector (i.e. a
matrix) of fertility rates. In the notation of Section 2.1 we then define
and consider the scalar RE (Equation7
(7)
(7) ) with
(37a)
(37a)
(37b)
(37b) Let's assume that
and
thus defined satisfy the conditions in Theorem 2.1.
When we have
(38)
(38) for
and in particular
(39)
(39) If
then
as
and consequently
as
.
Let's focus on the case . We can then compute the real time growth rate ρ as the unique solution of the Euler–Lotka equation
with necessarily either
or
. Since (Equation38
(38)
(38) ) can be written as
(40)
(40) we observe that the population growth rate ρ is the value of z for which both the generation and the real time process become stationary when we multiply all fertilities and all transition probabilities by
. When
the latter can be interpreted as introducing an additional, state-independent, death probability per time step
.
Then for
for some constant c and from (Equation28
(28)
(28) ) and (Equation29
(29)
(29) ) it now follows that
That is, the solution
of (Equation20
(20)
(20) ) with
grows, for large t, geometrically with rate ρ while converging to the stable distribution
(41)
(41) Here the index r refers to ‘real time’ (as opposed to ‘generation’). By rewriting (Equation41
(41)
(41) ) as
(42)
(42) we can (with the
norm) interpret the denominator as the life expectancy of a newborn individual when we introduce into the system an additional, state-independent, death probability per time step
(or, in other words, discount again and again the next year by a factor
).
In the spirit of (Equation9b(9b)
(9b) ) we next ask: how do the components of
contribute to future population sizes, i.e. to the constant c? From (Equation37b
(37b)
(37b) ) it follows that
(43)
(43) Now recall Theorem 2.1(iii) and conclude that the j-th component of the row vector
(44)
(44) specifies the (relative) contribution of individuals with i-state j to future population size. Or, in the now generally accepted terminology introduced by Fisher [Citation18], specifies the reproductive value of state j.
It is straightforward to check that
F + T has dominant eigenvalue ρ with the stable distribution
We normalize the vector of reproductive values by requiring
(45)
(45) and obtain
(46)
(46) If we now observe that
we can rewrite
as
(47)
(47) and interpret the denominator as
times the expected discounted age of the parent of a newborn individual.
Example 3.1
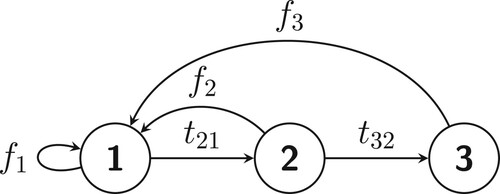
Consider the Leslie model with three age classes, i.e.
(see Figure for the schematic representation of the model). All individuals are born into the first age class so F = V U for
A straightforward calculation reveals that
which yields the Euler–Lotka equation
Hence
Using (Equation42
(42)
(42) ), we obtain the asymptotic age distribution
while (Equation47
(47)
(47) ) gives the row vector of reproductive values
where the constant c is determined such that
.
Figure 2. A schematic representation of the Leslie model in Example 3.1.
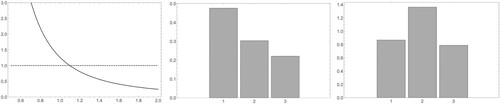
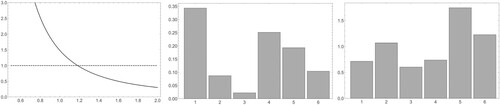
For the specific case with we find that
and
(see Figure (a)). The asymptotic age distribution is
while the normalized vector of reproductive values is
(see panels (b) and (c) of Figure ). Note that if we define
we can write
which clearly exposes the Leslie matrix model as the analogue of the PDE formulation of a continuous age and time model. The RE (Equation7
(7)
(7) ) is, of course, the analogue of Lotka's RE in the continuous age and time setting. Note also that
Figure 3. The plots of (a) , (b) the asymptotic age distribution and (c) the reproductive values for the (concrete example of the) Leslie model in Example 3.1.
3.2. More than one state-at-birth
Even when individuals may be born in different i-states, the number of individual states-at-birth is typically significantly lower than the total number of i-states, thus making the much smaller system sizes of RE an attractive alternative to (Equation20(20)
(20) ).
Suppose there are m states-at-birth with 1<m<n (this can now be understood literally, or, if aiming for the minimal RE system size, in a stochastic sense). We then write F = V U for some positive matrix V (normalized such that all m columns of V have
-norm equal to one) and a positive
matrix U and consider the RE (Equation16
(16)
(16) ) with
and
as in (Equation31a
(31a)
(31a) ). Next, require that the assumptions of Theorem 2.5 hold.
If the asymptotic dynamics of (Equation30
(30)
(30) ) is completely determined by the second term. Furthermore,
(48)
(48) for
. In particular,
is the next-generation matrix (possibly with small domain) and
(49)
(49) When
we have
as
and consequently
as well.
Suppose now that . The real time growth rate ρ is determined as the unique solution of the Euler-Lotka equation
where again either
or
. Note that rewriting (Equation48
(48)
(48) ) as
(50)
(50) we can once more observe that the population growth rate ρ is the value of z for which both the generation and the real time process become stationary when we multiply all fertilities and all transition probabilities by
(and when
the latter can be interpreted as introducing an additional, state-independent, death probability per time step
). Note also that the matrix
is the NGM (or NGM with small domain) when we discount fertilities and transition rates by a factor
,
The m-vector
will for large t grow geometrically with rate ρ while converging to the distribution Φ, where Φ is the normalized positive right eigenvector of
corresponding to eigenvalue 1. We can then deduce from (Equation28
(28)
(28) ) and (Equation29
(29)
(29) ) that, for
and a non-trivial initial condition
, the vector
grows geometrically with rate ρ while converging to the asymptotic distribution
(51a)
(51a)
(51b)
(51b) When using the
norm, we can interpret the denominator in (Equation51b
(51b)
(51b) ) as the expected duration of life of a newborn when newborns are sampled from the asymptotic distribution Φ and with additional, state-independent, death probability
introduced into the system (or alternatively, when we discount again and again the next year by
).
To see how the components of contribute to future population growth (i.e. to the constant c in (Equation19b
(19b)
(19b) )) we compute
(52)
(52) Using (Equation19b
(19b)
(19b) ) we now conclude that, if Ψ is a positive left eigenvector of
corresponding to eigenvalue 1 then the j-th element of the row n-vector
(53)
(53) specifies the (relative) contribution of individuals with i-state j to future population growth. Again, we normalize this vector by requiring
. Thus we obtain
(54a)
(54a)
(54b)
(54b) With the same reasoning as in the previous subsection, we can now interpret the j-th component of
in the denominator of (Equation54b
(54b)
(54b) ) as
times the expected age of the parent of a newborn individual with i-state j when we sample parents according to Φ and take into account an additional, state-independent, death probability
.
Again, one easily verifies that
F + T has dominant eigenvalue ρ with the stable distribution
Note that the conditions and
imposed on the resolvent can be interpreted biologically. Indeed, by definition the kernel
contains information about the expected numbers of daughters produced by one female at various ages. The matrix
yields the expected numbers of granddaughters (i.e. second generation offspring) and the j-th convolution of K with itself,
, yields the expected number of j-th generation offspring. Summing over all generations of offspring we obtain the clan kernel,
Since clan members of every female are either her daughters or clan members of one of her daughters, or, alternatively, either her daughters or daughters of a member of the clan, we must have
That is, the clan kernel
is the resolvent of K. The assumptions
and
can therefore be interpreted as follows:
| (A6) | Consider a newborn individual. At some time | ||||
| (A7) | Consider two newborn individuals. There exist | ||||
As a summary of the results, we provide at the end of this Section an algorithm for deducing the large time behaviour of (discrete time) structured-population models (SPM) via the renewal equation formulation (and for an application see the example below).
Example 3.2
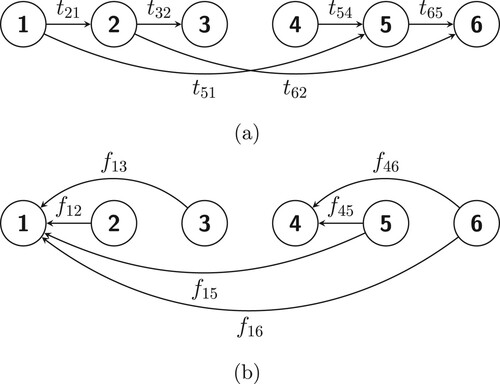
Consider a population in which individuals are characterized by both age a and some indicator of size s (for example, both age and size are important determinants of population dynamics of Rhododendron maximum shoots, see [Citation3, Citation23]) and let
be the set of i-states. We enumerate the i-states in the following way:
Let us first consider the case where all the s and the
s are strictly positive. Using (Equation31a
(31a)
(31a) ) we find
and using (Equation31b
(31b)
(31b) ) that
for
. Therefore
The positivity of the resolvent for
can also be deduced by interpretation. Indeed, if all the t's and the f's are strictly positive then individuals with either birth state will produce offspring of either size at ages 2 and 3. For
there will therefore be born clan members with any state-at-birth, irrespective of the birth state of the mother.
Figure 4. A schematic representation of (a) the state transitions and (b) fertilities in the model described in Example 3.2.
All the assumptions of Theorem 2.5 are therefore satisfied. We have
Let's assume that
. We can then compute the growth rate
as the unique real solution z of the Euler-Lotka equation
As a concrete example consider
Then
We can find the solution of the Euler-Lotka equation numerically and get
, while
(see Figure (a)). The asymptotic distribution of states-at-birth is found as the ( normalized) positive eigenvector of
corresponding to eigenvalue 1. We obtain
and then from (Equation51b
(51b)
(51b) ) conclude that the asymptotic population distribution is given by
We furthermore find that the left eigenvector of
corresponding to eigenvalue 1 (normalized such that
) is
from which we conclude using (Equation54b
(54b)
(54b) ) that the normalized vector of reproductive values is
(see also panels (b) and (c) of Figure ).
Figure 5. The plots of (a) , (b) the asymptotic i-state distribution and (c) the reproductive values for (the specific example of) the age-size structured-population model in Example 3.2.
Note that the condition of strict positivity of t's and f's can be relaxed somewhat. For example, if we assume that (i) individuals of size 1 only reproduce at age 3 (and then only produce offspring of size 1) and (ii) individuals of size 2 reproduce at age 2 (then producing size 1 individuals) and 3 (when they produce offspring of either size) then it is clear that is strictly positive for
.
In some situations, a further reduction of the NGM and the RE system is possible. Such is the case when we assume that (i) small individuals do not reproduce (i.e. ) and that (ii) large individuals produce size 1 and size 2 offspring at a given ratio
, regardless of the age of the parent. That is,
and
for some 0<p<1 and
, leading to
In this case, F (or U in (Equation56
(56)
(56) )) has rank 1. There are still two states-at-birth in the literal sense. However, if we interpret the term state-at-birth in the stochastic sense then all individuals are identical at birth in the sense that they are all born with i-state distribution
Then F = V U for
and the NGM with the small domain amounts to the scalar
In this special case, the basic reproduction number
can also easily be deduced from interpretation (and we invite the reader to do so).
We now have
and the scaler Euler-Lotka equation takes the form
(57)
(57) If
we can then first find the growth rate ρ as the unique real solution of the Euler–Lotka equation (Equation57
(57)
(57) ) and then use (Equation42
(42)
(42) ) and (Equation47
(47)
(47) ) to determine the asymptotic i-state distribution and the asymptotic reproductive values, respectively.
Deducing the asymptotic behaviour in (linear discrete time) SPM from the corresponding RE
Starting with a structured-population model
the recipe is as follows.
| Step 1. | Identify states-at-birth and write the fertility matrix in the form F = V U for some normalised Remark. For practical purposes, it may be handy to order the i-states such that the birth states come first. Then in the case of m states-at-birth (in the literal sense) V is the matrix with columns | ||||
| Step 2. |
| ||||
| Step 3. |
| ||||
1 A sufficient condition is that all column sums of T are strictly below 1.
4. The case
We now show directly that in the special case when we can compute
the asymptotic distribution (i.e. the right eigenvector) and
the reproductive values (i.e. the left eigenvector)
in the real time setting from those obtained in the generation-bookkeeping framework. Note that the former result has been demonstrated in a more general setting in [Citation10, Citation12].
Suppose that there are m states-at-birth in the literal sense and as before write F = V U for some normalized birth-state matrix V and the
fertility matrix U. Then
is the next-generation matrix.
Let and
denote the normalized positive right eigenvector in, respectively, generation and real time framework, corresponding to eigenvalue 1. That is
(58a)
(58a) and
(58b)
(58b) Now apply V from the left on both sides of (Equation58a
(58a)
(58a) ) and write
If we now denote
then clearly
. We thus find that the asymptotic distributions in real time and in the generation setting are related by
(59)
(59) as indeed follows from (Equation51b
(51b)
(51b) ) when we observe that with
,
is the NGM and
. When using the
norm, the denominator in (Equation59
(59)
(59) ) is the expected duration of life of a newborn when we sample newborns according to
. As observed before,
is the asymptotic distribution in the generation-bookkeeping framework when we consider the NGM with large domain
.
Now let and
denote the normalized positive left eigenvector in, respectively, generation and real time framework, corresponding to eigenvalue 1, i.e.
(60a)
(60a) and
(60b)
(60b) Acting with U from the right on both sides of (Equation60a
(60a)
(60a) ) we find that
Then for we have
. We thus find that the reproductive values in the real time setting relate to the ones in the generation-bookkeeping by
where the constant c is chosen such that with
in (Equation59
(59)
(59) ) we have
. We obtain
(61)
(61) Alternatively, we can deduce this from (Equation54b
(54b)
(54b) ) by taking into account that with
we have
. The j-th component of
in the denominator of (Equation61
(61)
(61) ) is the expected age of the parent of a newborn individual with i-state j when we sample parents according to
. As observed before,
is also the vector of generation-based reproductive values when we consider the NGM with large domain
.
5. Concluding remarks
The specification of a linear physiologically structured population model involves two rules, one for reproduction and one for development/maturation/movement and survival. Once these ingredients are specified, one can constructively define next-population-state operators. For constant environments, i.e. autonomous dynamics, a first question is: will the population ultimately grow exponentially (in which case one has to ponder the issue of density dependence) or decline and go extinct? As emphasized by Jim Cushing and Zhou Yicang [Citation7], an efficient way of answering the question is to adopt a generation perspective, by focusing on expected lifetime offspring production by newborn individuals, and to characterize the appropriate average as the dominant eigenvalue of the next-generation matrix. On the other hand, people familiar with the Perron+-Frobenius theory of positive dynamical systems (as presented in, for instance, Berman and Plemmons [Citation2], Bátkai et al. [Citation1]) will be inclined to compute the spectral bound of the generator of the real time dynamics. Here, in the spirit of the elegant Li-Schneider paper [Citation22], we have uncovered how these two approaches relate to each other. Our main contribution has been to highlight the connecting role of the Renewal Equation and to revive the powerful results of Feller and Thieme that describe the asymptotic large time behaviour of its solutions. As a one sentence summary of the present paper we offer: use the ingredients to formulate the RE, apply Feller/Thieme, and everything you might be interested in can next be computed explicitly.
Acknowledgments
Hans Metz benefited from the support from the ‘Chaire Modélisation Mathématique et Biodiversité of Veolia Environnement-École Polytechnique-Museum National d'Histoire Naturelle-Fondation X’.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- A. Bátkai, M. Kramar Fijavž, and A. Rhandi, Positive Operator Semigroups: From Finite to Infinite Dimensions, Vol. 257, Birkhäuser, Basel, 2017.
- A. Berman and R.J. Plemmons, Nonnegative Matrices in the Mathematical Sciences, SIAM, Philadelphia, 1994.
- H. Caswell, Matrix Population Models, Sinauer Sunderland, MA, 2000.
- K.S. Crump, On systems of renewal equations, J. Math. Anal. Appl. 30 (1970), pp. 425–434.
- J.M. Cushing, An Introduction to Structured Population Dynamics, SIAM, Philadelphia, 1998.
- J.M. Cushing and O. Diekmann, The many guises of R0 (a didactic note), J. Theor. Biol. 404 (2016), pp. 295–302.
- J.M. Cushing and Y. Zhou, The net reproductive value and stability in matrix population models, Nat. Resour. Model. 8 (1994), pp. 297–333.
- B. De Saporta, Renewal theorem for a system of renewal equations, Ann. Inst. Henri Poincare (B) Probab. 39 (2003), pp. 823–838.
- O. Diekmann, J.A.P. Heesterbeek, and J.A.J. Metz, On the definition and the computation of the basic reproduction ratio R0 in models for infectious diseases in heterogeneous populations, J. Math. Biol. 28 (1990), pp. 365–382.
- O. Diekmann, M. Gyllenberg, J.A.J. Metz, and H.R. Thieme, On the formulation and analysis of general deterministic structured population models I. Linear theory, J. Math. Biol. 36 (1998), pp. 349–388.
- O. Diekmann, M. Gyllenberg, H. Huang, M. Kirkilionis, J.A.J. Metz, and H.R. Thieme, On the formulation and analysis of general deterministic structured population models II. Nonlinear theory, J. Math. Biol.43 (2001), pp. 157–189.
- O. Diekmann, M. Gyllenberg, and J.A.J. Metz, Steady-state analysis of structured population models, Theor. Popul. Biol. 63 (2003), pp. 309–338.
- O. Diekmann, P. Getto, and M. Gyllenberg, Stability and bifurcation analysis of Volterra functional equations in the light of suns and stars, SIAM J. Math. Anal. 39 (2008), pp. 1023–1069.
- O. Diekmann, J.A.P. Heesterbeek, and M.G. Roberts, The construction of next-generation matrices for compartmental epidemic models, J. R. Soc. Interface 7 (2010), pp. 873–885.
- O. Diekmann, J.A.P. Heesterbeek, and T. Britton, Mathematical Tools for Understanding Infectious Disease Dynamics, Princeton University Press, Princeton, 2013.
- W. Feller, An Introduction to Probability Theory and Its Applications, Vol. 1, John Wiley & Sons, New York, 1968.
- W. Feller, An Introduction to Probability Theory and Its Applications, Vol. 2, John Wiley & Sons, New York, 1971.
- R. Fisher, The Genetical Theory of Natural Selection, 2nd ed., Dover Publications, New York, 1958.
- E. Franco, O. Diekmann, and M. Gyllenberg, Modelling physiologically structured populations: Renewal equations and partial differential equations, J. Evol. Equ. 23 (2023), pp. 23–46.
- H.J.A.M. Heijmans, The Dynamical Behaviour of the Age-Size-Distribution of a Cell Population, Springer, Berlin, 1986.
- H. Inaba, Age-Structured Population Dynamics in Demography and Epidemiology, Springer, Berlin, 2017.
- C.-K. Li and H. Schneider, Applications of Perron–Frobenius theory to population dynamics, J. Math. Biol. 44 (2002), pp. 450–462.
- J. McGraw, Effects of age and size on life histories and population growth of Rhododendron maximum shoots, Am. J. Bot. 76 (1989), pp. 113–123.
- J.A.J. Metz and O. Diekmann, The Dynamics of Physiologically Structured Populations, Lecture Notes in Biomathematics Vol. 68, 1986. http://webarchive.iiasa.ac.at/Research/ADN/Metz2Book.html.
- H.R. Thieme, Renewal theorems for linear discrete Volterra equations, J. Für Die Reine und Angew. Math. 353 (1984), pp. 55–84.
- H.R. Thieme, Spectral bound and reproduction number for infinite-dimensional population structure and time heterogeneity, SIAM. J. Appl. Math. 70 (2009), pp. 188–211.
- H.R. Thieme, Mathematics in Population Biology, Princeton University Press, Princeton, 2018.
- H.R. Thieme, Discrete-time population dynamics on the state space of measures, Math. Biosci. Eng. 17 (2020), pp. 1168–1217.
- H.R. Thieme, Reproduction number versus turnover number in structured discrete-time population models, in Advances in Discrete Dynamical Systems, Difference Equations and Applications: 26th ICDEA, Sarajevo, Bosnia and Herzegovina, July 26–30, 2021, Springer, 2023, pp. 495–539.
Appendix. Concise summary of the continuous time formalism
As the analogue of (Equation20(20)
(20) ) we consider the linear ODE system
(A1)
(A1) where X takes values in
, F is a positive
matrix describing reproduction and T is a positive-off-diagonal
matrix describing survival and development. We assume that the spectral bound
is negative, allowing us to write
(A2)
(A2) The variation-of-constants version of (EquationA1
(A1)
(A1) ) reads
(A3)
(A3) Putting
(A4)
(A4) and applying F to both sides of (EquationA3
(A3)
(A3) ) we obtain the RE
(A5)
(A5) and, once we have constructed Y by solving this RE, we can interpret (EquationA3
(A3)
(A3) ) as an explicit expression for X:
(A6)
(A6) Whenever F has the form (Equation21
(21)
(21) ), we can introduce
. Then
and we can rewrite (EquationA5
(A5)
(A5) ) as
(A7)
(A7) The translation invariant version of (EquationA7
(A7)
(A7) ) is
(A8)
(A8) Solutions of (EquationA1
(A1)
(A1) ) and (EquationA8
(A8)
(A8) ) defined for all times, i.e. for
, are related to each other by
(A9)
(A9) Clearly (EquationA1
(A1)
(A1) ) has a solution of the form
(A10)
(A10) if and only if r is an eigenvalue of F + T and
is a corresponding eigenvector, i.e.
(A11)
(A11) By substitution we see that (EquationA8
(A8)
(A8) ) has a solution of the form
(A12)
(A12) if and only if
has eigenvalue 1 and Φ is a corresponding eigenvector, i.e.
(A13)
(A13) According to (EquationA9
(A9)
(A9) ) we have
(A14)
(A14) We recall how this can be used in practice:
| Step 1. | Find the real number r for which the spectral radius of | ||||
| Step 2. | Find Φ as the dominant positive eigenvector of | ||||
| Step 3. | Compute | ||||
For the corresponding left eigenvectors we have
(A15)
(A15) Rewriting the first of these as follows:
we see that, modulo normalization,
and Ψ are identical. Note that the NGM with large domain is now given by
(A16)
(A16) and that, with
defined as the spectral radius of
, the relation
(A17)
(A17) holds. In the special case
, Ψ and Φ are, respectively, the left and the right eigenvector of
corresponding to its dominant eigenvalue.
The formalism easily extends to an i-state space that is a continuum and to measures see [Citation10, Citation19, Citation20, Citation26]. A convenient starting point is the following generalization of (EquationA9(A9)
(A9) ):
(A18)
(A18) By substituting the second of these identities into the first we obtain the RE
(A19)
(A19) with
(A20)
(A20) Clearly, (EquationA19
(A19)
(A19) ) has a solution of the form
(A21)
(A21) if and only if
(A22)
(A22) We refer to [Citation19] for a detailed analysis.