
Abstract
A growing body of work has found that a mismatch between early and recent life stress, more than a cumulative influence of stress, contributes to detrimental stress-related health outcomes. To date, however, no work has examined how such a mismatch might relate to stress-related cognitive outcomes. We addressed this gap in the current study by assessing participants’ (N = 154, Mage = 18.7, 104 female) early and recent life stress using the same inventory, and subsequently assessing their inhibitory control in a hybrid stop-signal/flanker task. Surprisingly, we found that a greater degree of stressor mismatch was associated with better response inhibition (i.e. smaller stop-signal reaction time) across a number of analytic approaches. Cognitive inhibition (i.e. the flanker interference effect) was not associated with stressor mismatch. These results thus show that a greater degree of mismatch between early and recent life stress is related to response inhibition in the same way as acute stress affects response inhibition, suggesting that response inhibition may be an important cognitive process for navigating both acute stress and general environmental conditions that do not match the conditions in which expected stress occurrence was established.
More stress produces worse outcomes, or at least that’s how the story goes. Recent work, however, has suggested that distinct patterns of stress exposure might relate to distinct health outcomes (Nederhof & Schmidt, Citation2012). To date, no work has examined how distinct patterns of time-dependent stressor exposure might relate to cognitive functioning. This study helps to address that gap.
Although the predominant theory of stress and health—allostatic load, or cumulative exposure effects (McEwen, Citation1998)—has best explained most findings related to stress and health, it has recently been challenged. In particular, various theories of stress, including the adaptive calibration hypothesis and the mismatch hypothesis (Del Giudice et al., Citation2011; Nederhof & Schmidt, Citation2012), have noted that a mismatch between the frequency and/or severity of early life and recent stress exposure is important to some stress-related outcomes. The mismatch hypothesis proposes that individuals are adaptively programmed to best survive in the environment present during sensitive developmental periods in childhood, and that individuals experience dysfunction when their current environment does not match the environment to which they adapted in early life (Nederhof & Schmidt, Citation2012). This hypothesis thus carries with it the interesting implication that individuals with greater early adversity should be less impacted by high stress in adulthood than individuals with little early adversity. A fair amount of evidence supports this hypothesis with respect to health outcomes (e.g. Santarelli et al., Citation2014). For example, a mismatch between the frequency of early and recent life stress is associated with greater rates of depression and substance use (McMullin et al., Citation2021; Santarelli et al., Citation2014). These findings suggest that patterns of stressor exposure, more than overall stressor exposure, might be important in some stress-related outcomes, such as cognitive function.
To date, no work has examined how a mismatch between early and recent life stress might relate to cognitive processes. Prior work has suggested that the differential effects of acute stress on inhibitory control processes (i.e. response inhibition and cognitive inhibition) may be the strongest effect of stress on executive functions (e.g. Shields et al., Citation2016), making inhibitory control an ideal first step to examine in relation to stress mismatch.
Current research
The current study addressed the above gap by examining associations between early and recent life stress in relation to inhibitory control. Participants (N = 154 with usable data) first completed a modified form of the Childhood Trauma Questionnaire, which assessed both recent (i.e. over the last year) and early (i.e. before 13 years of age) exposure to each stressor. Next, participants completed a hybrid stop-signal/flanker task, which permitted assessment of both response inhibition and cognitive inhibition.
We expected to find that early and recent life stress would be associated with poorer response inhibition and/or cognitive inhibition. In addition, and consistent with the mismatch hypothesis, we expected to find that a mismatch between early and recent life stress would be particularly strongly associated with poorer inhibitory control.
Method
Participants
We recruited 154 participants (Mage = 18.7, SDage = 0.9, rangeage : [18, 24]; 67.5% female) with usable data according to our a priori exclusion criteria (see Inhibitory Control section) for this study. Participants were undergraduate students enrolled in a general introductory or other psychology course that offered extra credit for participation. This sample size represented all participants we were able to recruit for this study over a two-semester period within the context of data collection for other projects; we did not conduct a power analysis prior to collecting data, nor did we have a specific sample size goal for this study, given that no prior work had examined stress mismatch in relation to inhibitory control. This sample size achieved 80% power to detect small-moderate correlations (r = .20). When identifying their ‘primary’ race or ethnicity, 79.2% of participants identified as White, 11.7% as Hispanic or Latino/a/e, 3.9% as Black or African American, 3.9% as Asian or Asian American, and 1.3% as Native Hawaiian or Pacific Islander.
Materials
Stress exposure
Early and recent life stress were assessed using a modified version of the Childhood Trauma Questionnaire-Short Form. The CTQ-SF is valid and reliable (Bernstein et al., Citation2003; Slavich & Shields, Citation2018); for example, greater childhood adversity as quantified by the CTQ-SF predicts the occurrence of doctor-diagnosed autoimmune disorders (Slavich & Shields, Citation2018). The CTQ-SF contains 28 questions, 25 of which retrospectively assess frequencies of specific forms of stress exposure, and three of which are validity questions that quantify a lack of consideration in responses (e.g. “I had the perfect family” should not receive a maximum frequency response). The CTQ-SF was modified to retrospectively assess the occurrence of each stressor in both childhood and adulthood. In particular, each CTQ-SF item contained two sets of response options directly below it. The first of these asked participants how frequently true a given statement was “as a child (<13 years old),” and the second asked participants how frequently true a given statement was “over the last year.” Additionally, questions relating to parental and family support in childhood (e.g. “people in my family said hurtful or insulting things to me”) were reframed appropriately for adulthood (e.g. “people I cared about said hurtful or insulting things to me”). Participants indicated how often given statements were true using a Likert scale ranging from 1 (Never true) to 5 (Very often true). Internal consistency was very good for early life stress (α = .85) and excellent for recent life stress (α = .92). Stress mismatch was calculated by taking the absolute difference of each question between the early and recent timeframes, then summing those absolute differences.
Inhibitory control
A modified hybrid stop-signal/flanker task was used to assess inhibitory control (see Supplemental Material).
Response inhibition is indexable in this task via stop-signal reaction time (SSRT), calculated using the recommended integration method (Verbruggen et al., Citation2013). SSRT represents the time required for a participant to inhibit an activated response; lower values thus indicate better response inhibition.
Cognitive inhibition is indexable in this task via flanker interference, calculated using correct trials only as mean(RTincongruent) – mean(RTcongruent). Larger interference effects indicate worse cognitive inhibition.
A priori exclusion criteria for this task were making five or more responses under 150 ms (indicating intentional premature responding), having ten or more failures to respond on “go” trials (indicating waiting, violating SSRT model assumptions), less than 80% target accuracy on “go” trials, and less than 35% or greater than 65% accuracy on stop-signal trials (indicating responses violating SSRT model assumptions).
Procedure
Participants came to the lab for one-hour timeslots and were each seated in individual cubicles. Participants then provided informed consent, completed a demographics questionnaire, and completed the modified CTQ-SF among three filler questionnaires before finally completing the inhibitory control task. Participants were then thanked, debriefed, and dismissed. This protocol was approved by the University of Arkansas IRB.
Results
Descriptive statistics and correlations between raw stressor variables and inhibition measures are presented in . Early and recent stress were log transformed to correct for skew (i.e. skewness > 1; both skewness statistics were > 1.67) for Gaussian models. No other variable analyzed in Gaussian models required transformation (|skewness| < 0.59). Using nontransformed variables only strengthened what follows.
Table 1. Descriptive statistics for and correlations between raw stress and inhibition variables.
In a moderated regression analysis, we found that early and recent life stress significantly interacted to predict response inhibition (i.e. stop-signal reaction time), β = .174, p = .032, but not cognitive inhibition (i.e. flanker interference), β = .046, p = .581. Excluding two mismatch outliers (response inhibition: p = .020; cognitive inhibition: p = .645) or the participants who failed validity checks in the stress assessment (response inhibition: p = .035; cognitive inhibition: p = .337) only strengthened this interaction.Footnote1 Simple slopes analyses, described below, showed that a mismatch in either direction (i.e. high-early-but-low-recent stress or low-early-but-high-recent stress) was associated with better response inhibition.
We conducted simple slopes analyses sequentially, first holding recent life stress constant at various fixed points (e.g. low, high), then holding early life stress constant at various fixed points. When holding recent life stress constant at different points, higher early life stress was only associated with lower stop-signal reaction time (i.e. better response inhibition) when recent life stress was low: high (mean + 1SD) recent life stress, βearly-life-stress-to-SSRT = .076, p = .545; mean recent life stress, βearly-life-stress-to-SSRT = −.110, p = .217; low (mean − 1SD) recent life stress, βearly-life-stress-to-SSRT = −.297, p = .016. Similarly, when holding early life stress constant at different points, higher recent life stress was only associated with lower stop-signal reaction time (i.e. better response inhibition) when early life stress was low or average: high (mean + 1SD) early life stress, βrecent-life-stress-to-SSRT = .008, p = .946; mean early life stress, βrecent-life-stress-to-SSRT = −.179, p = .050; low (mean − 1SD) early life stress, βrecent-life-stress-to-SSRT = −.366, p = .009.
An inverted-U relation also showed that a stress mismatch in either direction was associated with better response inhibition (see Supplemental Material).
Stressor-specific mismatch analyses
The above analyses operate on interactions between sum scores for early life and recent life stress (i.e. at the level of summary scores). However, as described in Materials, the novel stress assessment permits quantification of mismatch at the level of individual stressors, which allows participants to simultaneously quantify discrepancies in the frequency of each stressor between timepoints (i.e. early life and recently), providing a more sensitive estimate of mismatch in each stressor. In what follows, we analyze this mismatch score, which cannot be examined as an interaction (i.e. it is the sum of absolute difference between each stressor).
Because stress mismatch, as a count variable (i.e. stress frequency absolute differences), could be examined in relation to indices of inhibitory control using generalized linear models, zero-inflated models, or hurdle models with either a Poisson or negative binomial distribution, we examined each of these models and present the results of these analyses in and .
Table 2. Models examining relations between response inhibition and early/recent life stress mismatch.
Table 3. Models examining relations between cognitive inhibition and early/recent life stress mismatch.
With respect to response inhibition (i.e. stop-signal reaction time) in relation to stress mismatch, we found that the best-fitting model was a negative binomial generalized linear model (ΔBICs > 4.9). Importantly, though, the results of every model converged on the same significant result obtained in the best-fitting model: Better response inhibition (i.e. lower stop-signal reaction time) was associated with a greater degree of stress mismatch B = −0.01, p < .001, R2pseudo = .046 (see ). That is, a greater degree of mismatch between early and recent life stress was associated with better response inhibition.Footnote2 Estimated marginal means indicated that participants who showed better response inhibition than mean performance by one SD had nearly double the mismatch (1.91-fold higher) between early and recent life stress (mismatch M = 13.61, SE = 1.61) than the mismatch had by participants who showed worse response inhibition than the mean by one SD (mismatch M = 7.11, SE = 0.87), z = 3.79, p < .001.
Figure 1. Stressor mismatch in relation to inhibitory control outcomes.
Note. A greater mismatch between early and recent life stress was associated with better response inhibition (i.e. less stop-signal reaction time, which indicates the time needed to stop an ongoing response) (a). However, mismatch between early and recent life stress was unassociated with cognitive inhibition (i.e. flanker interference) (b). Note that these figures omit two outliers in stress mismatch for ease of visualization; analyses excluding those outliers did not differ from those in the main text and are presented in Supplemental Material.
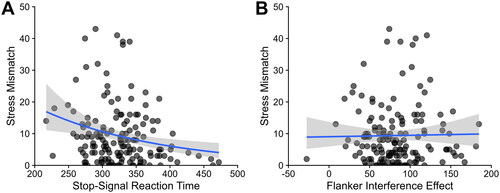
With respect to our measure of cognitive inhibition (i.e. flanker interference effects) in relation to stressor mismatch, we found that the best-fitting model was a negative binomial generalized linear model (ΔBICs > 4.9), with no association between flanker interference and stress mismatch, p = .610. Importantly, the results of every model converged on the same result as obtained in the best-fitting model: Cognitive inhibition (i.e. flanker interference) was not associated with stressor mismatch, ps > .110 ().
Discussion
In this study, we examined the associations between different forms of stressor exposure at different points in life and their associations with inhibitory control processes. Contrary to predictions, we found that, in bivariate associations, greater early and recent life stress were each associated with better response inhibition. However, both of these associations were qualified by an interaction between the two of them: Consistent with the mismatch hypothesis, we found that a mismatch between early and recent life stress was uniquely associated with response inhibition, such that only when there was a degree of mismatch between early and recent life stress was stress associated with better response inhibition. In contrast, there were no associations between stress and cognitive inhibition, either mismatch or cumulative. These results suggest that a mismatch between recent and early stressor exposure relates to response inhibition in a similar way to the causal effect of acute stress on response inhibition, but no similar pattern is evident in cognitive inhibition.
Our finding that a greater degree of mismatch between early and recent life stress exposure was associated with better response inhibition was unexpected, as it is inconsistent with some (e.g. Möschl et al., Citation2022; Shields et al., Citation2019; Weckesser et al., Citation2021), though not all (e.g. Gao et al., Citation2022; see also Grillon et al., Citation2017) prior work on chronic stress and response inhibition. Importantly, though, this association is consistent with work on acute stress. In particular, acute stress improves response inhibition (Chang et al., Citation2020; Dierolf et al., Citation2018; Shields et al., Citation2016), and this finding has been interpreted to reflect a reallocation of cognitive resources that enhances control over actions ideal for fighting or fleeing from the current stressor (Shields et al., Citation2016). Individuals with mismatched early and recent life stress may show improved response inhibition in general because such a mismatch may indicate a generally uncertain environment and thus that actions should be driven less by habit than in more stable and predictable environments. This mismatch, which itself could represent uncertainty and unpredictability (and thus contain characteristics of a stressor), then, may produce states similar to those induced by acute stress, which could explain the observed mismatch association with response inhibition.
Although early adversity can be associated with better cognitive performance (Frankenhuis & de Weerth, Citation2013), to our knowledge, this study is the first to document an improvement in functioning from a stress mismatch. To speculate about mechanism, the mismatch hypothesis does not operate on the level of subjective states, such as hypervigilance or anxiety, but on the level of stress system (mis-)calibration. In line with this, Santarelli et al. (Citation2017) found that a mismatch between early and recent stress affected expression of both mineralocorticoid and glucocorticoid receptors in adulthood. Importantly, antagonism of mineralocorticoid receptors abolishes the beneficial effects of acute stress on stop-signal reaction time (Schwabe et al., Citation2013). It is thus possible that a mismatch between early and recent life stress improves stop-signal reaction time by altering the functioning of the mineralocorticoid receptor system. Future research could examine this possibility.
Although this study has a number of strengths, including simultaneous assessment of multiple inhibitory control processes, a multiverse analytic approach examining primary associations with multiple model types, and a novel stress assessment that permitted assessment of mismatch at the level of individual stressors, it also has limitations that should be noted. First, response and cognitive inhibition can influence each other when they are assessed within the same task (Verbruggen et al., Citation2004), and it is possible that these results would have come out differently if the inhibition task used required only one type of inhibitory control. Second, although we achieved 80% power to detect a small-moderate correlation of r = .20, we only achieved 34% power to detect a correlation of r = .10, entailing that we would have likely failed to detect true associations between stress and cognitive inhibition if they exist but are small in magnitude. Third, our participants’ mean age was approximately 19 years. Because of this, participants’ early stress (< 13 years) was relatively close in temporal proximity to their recent stress (preceding year). It is possible that an early-recent stress mismatch could exhibit different associations in older participants. Fourth, although we observed no moderations by sex, we may have observed such moderations if our sample had been larger. Our data cannot address these possibilities, so we do not claim that the associations that we observed would not differ by age, sex, race, or other factors in other samples. Fifth, we did not assess physiological stress indices, which may have helped to elucidate mechanisms or provided other insights into the patterns that we observed. Finally, as this study collected data from a college student sample, this study included a Western, Educated, Industrialized, Rich, and Democratic (WEIRD) sample, and future research should determine whether these results replicate in a non-WEIRD sample. Nonetheless, participants’ race/ethnicity were similar to other studies using the University’s psychology participant pool (e.g. Shields et al., Citation2024), and to the Fayetteville, Arkansas data currently on census.gov.
Conclusion
Although cumulative stress exposure is detrimental in many ways, recent work has highlighted the importance of a mismatch between early and recent stress exposure in predicting specific outcomes. Previously, however, no work had examined this mismatch in relation to cognitive function. In this study, we found that a greater degree of mismatch between early and recent stress exposure predicted better response inhibition, but this mismatch was not associated with cognitive inhibition. These findings suggest that stressor mismatch may be associated with generally improved motor-related executive control, which may help to navigate generally unpredictable environments by reducing the extent to which habitual actions govern behavior.
Supplemental Material
Download MS Word (117.6 KB)Disclosure statement
No potential conflict of interest was reported by the author(s).
Data availability statement
Data and syntax for analyses are available at https://osf.io/qau6y/?view_only=384bd223ff5e494cb9e67e68b6a12670.
Additional information
Funding
Notes on contributors
Grant S. Shields
Grant S. Shields, Ph.D., is an assistant professor in the Department of Psychological Science at the University of Arkansas. He studies cognition in context, with the majority of his work aiming to understand cognitive processes under stress.
Colton L. Hunter
Colton L. Hunter, M.A., is a Ph.D. student at the University of Arkansas. Colton’s research focuses on distortions in memory and cognition, particularly under conditions of stress and heightened emotions.
Notes
1 See Supplemental Material for all of the following analyses with these two sets of exclusion criteria; all results were consistent with the following results.
2 This association did not differ by sex, p = .070, and held when examining female participants alone, B = -.01, p < .001. Similarly, age did not moderate this association, p = .791.
References
- Bernstein, D. P., Stein, J. A., Newcomb, M. D., Walker, E., Pogge, D., Ahluvalia, T., Stokes, J., Handelsman, L., Medrano, M., Desmond, D., & Zule, W. (2003). Development and validation of a brief screening version of the Childhood Trauma Questionnaire. Child Abuse & Neglect, 27(2), 1–7. https://doi.org/10.1016/S0145-2134(02)00541-0
- Chang, J., Hu, J., Li, C. S. R., & Yu, R. (2020). Neural correlates of enhanced response inhibition in the aftermath of stress. NeuroImage, 204, 116212. https://doi.org/10.1016/j.neuroimage.2019.116212
- Del Giudice, M., Ellis, B. J., & Shirtcliff, E. A. (2011). The adaptive calibration model of stress responsivity. Neuroscience and Biobehavioral Reviews, 35(7), 1562–1592. https://doi.org/10.1016/j.neubiorev.2010.11.007
- Dierolf, A. M., Schoofs, D., Hessas, E.-M., Falkenstein, M., Otto, T., Paul, M., Suchan, B., & Wolf, O. T. (2018). Good to be stressed? Improved response inhibition and error processing after acute stress in young and older men. Neuropsychologia, 119, 434–447. https://doi.org/10.1016/J.NEUROPSYCHOLOGIA.2018.08.020
- Frankenhuis, W. E., & de Weerth, C. (2013). Does early-life exposure to stress shape or impair cognition? Current Directions in Psychological Science, 22(5), 407–412. https://doi.org/10.1177/0963721413484324
- Gao, H., Wang, X., Huang, M., & Qi, M. (2022). Chronic academic stress facilitates response inhibition: Behavioral and electrophysiological evidence. Cognitive, Affective & Behavioral Neuroscience, 22(3), 533–541. https://doi.org/10.3758/s13415-021-00974-x
- Grillon, C., Robinson, O. J., O’Connell, K., Davis, A., Alvarez, G., Pine, D. S., & Ernst, M. (2017). Clinical anxiety promotes excessive response inhibition. Psychological Medicine, 47(3), 484–494. https://doi.org/10.1017/S0033291716002555
- McEwen, B. S. (1998). Stress, adaptation, and disease: Allostasis and allostatic load. Annals of the New York Academy of Sciences, 840(1), 33–44. https://doi.org/10.1111/j.1749-6632.1998.tb09546.x
- McMullin, S. D., Shields, G. S., Slavich, G. M., & Buchanan, T. W. (2021). Cumulative lifetime stress exposure predicts greater impulsivity and addictive behaviors. Journal of Health Psychology, 26(14), 2921–2936. https://doi.org/10.1177/1359105320937055
- Möschl, M., Schmidt, K., Enge, S., Weckesser, L. J., & Miller, R. (2022). Chronic stress and executive functioning: A specification-curve analysis. Physiology & Behavior, 243, 113639. https://doi.org/10.1016/j.physbeh.2021.113639
- Nederhof, E., & Schmidt, M. V. (2012). Mismatch or cumulative stress: Toward an integrated hypothesis of programming effects. Physiology & Behavior, 106(5), 691–700. https://doi.org/10.1016/j.physbeh.2011.12.008
- Santarelli, S., Lesuis, S. L., Wang, X.-D., Wagner, K. V., Hartmann, J., Labermaier, C., Scharf, S. H., Müller, M. B., Holsboer, F., & Schmidt, M. V. (2014). Evidence supporting the match/mismatch hypothesis of psychiatric disorders. European Neuropsychopharmacology: The Journal of the European College of Neuropsychopharmacology, 24(6), 907–918. https://doi.org/10.1016/j.euroneuro.2014.02.002
- Santarelli, S., Zimmermann, C., Kalideris, G., Lesuis, S. L., Arloth, J., Uribe, A., Dournes, C., Balsevich, G., Hartmann, J., Masana, M., Binder, E. B., Spengler, D., & Schmidt, M. V. (2017). An adverse early life environment can enhance stress resilience in adulthood. Psychoneuroendocrinology, 78, 213–221. https://doi.org/10.1016/j.psyneuen.2017.01.021
- Schwabe, L., Höffken, O., Tegenthoff, M., & Wolf, O. T. (2013). Stress-induced enhancement of response inhibition depends on mineralocorticoid receptor activation. Psychoneuroendocrinology, 38(10), 2319–2326. https://doi.org/10.1016/j.psyneuen.2013.05.001
- Shields, G. S., Hunter, C. L., Trudell, E. V., Gray, Z. J., Perkins, B. C., Patterson, E. G., & Zalenski, P. K. (2024). Acute stress influences the emotional foundations of executive control: Distinct effects on control-related affective and cognitive processes. Psychoneuroendocrinology, 162, 106942. https://doi.org/10.1016/j.psyneuen.2023.106942
- Shields, G. S., Ivory, S. L., & Telzer, E. H. (2019). Three-month cumulative exposure to testosterone and cortisol predicts distinct effects on response inhibition and risky decision-making in adolescents. Psychoneuroendocrinology, 110, 104412. https://doi.org/10.1016/j.psyneuen.2019.104412
- Shields, G. S., Sazma, M. A., & Yonelinas, A. P. (2016). The effects of acute stress on core executive functions: A meta-analysis and comparison with effects of cortisol. Neuroscience and Biobehavioral Reviews, 68, 651–668. https://doi.org/10.1016/j.neubiorev.2016.06.038
- Slavich, G. M., & Shields, G. S. (2018). Assessing lifetime stress exposure using the stress and adversity inventory for adults (adult STRAIN): An overview and initial validation. Psychosomatic Medicine, 80(1), 17–27. https://doi.org/10.1097/PSY.0000000000000534
- Verbruggen, F., Liefooghe, B., & Vandierendonck, A. (2004). The interaction between stop signal inhibition and distractor interference in the flanker and Stroop task. Acta Psychologica, 116(1), 21–37. https://doi.org/10.1016/j.actpsy.2003.12.011
- Verbruggen, F., Chambers, C. D., & Logan, G. D. (2013). Fictitious inhibitory differences: How skewness and slowing distort the estimation of stopping latencies. Psychological Science, 24(3), 352–362. https://doi.org/10.1177/0956797612457390
- Weckesser, L. J., Schmidt, K., Möschl, M., Kirschbaum, C., Enge, S., & Miller, R. (2021). Temporal stability and effect dynamics between executive functions, perceived chronic stress, and hair cortisol concentrations. Developmental Psychology, 57(7), 1149–1162. https://doi.org/10.1037/dev0001193