Abstract
This essay describes the evolution of stress as a medical scientific idea. Claude Bernard, Walter B. Cannon and Hans Selye provided key founding concepts for the current view. Bernard introduced the idea of the internal environment bathing cells—the milieu intérieur—maintained by continual compensatory changes of bodily functions. Cannon coined the word, “homeostasis,” referring to a set of acceptable ranges of values for internal variables. Cannon taught that threats to homeostasis evoke activation of the sympathoadrenal system as a functional unit. Selye defined stress as a state characterized by a uniform response pattern, regardless of the particular stressor, that could lead to long-term pathologic changes. “Allostasis” was introduced as a concept in recognition that there is no single ideal set of steady-state conditions in life; instead, setpoints and other response criteria change continuously. Stress is now viewed neither as a perturbation nor a stereotyped response pattern but as a condition characterized by a perceived discrepancy between information about a monitored variable and criteria for eliciting patterned effector responses. Different stressors elicit different patterns of activation of the sympathetic nervous, adrenomedullary hormonal, hypothalamic-pituitary-adrenocortical and other effectors, closing negative feedback loops. This systems concept of stress yields predictions that observation or experimentation can test and that are applicable to normal physiology and to a variety of acute and chronic disorders.
Historical overview of stress concepts
Cannon, homeostasis and the sympathoadrenal system
In 1865, in his masterpiece, “Introduction à la médecine expérimentale,” Claude Bernard described his earlier observations on the role of the liver in secreting glucose formed from glycogen stores and how his studies of heat regulation led to the discovery of vascular blood flow regulation by sympathetic nerves. By a tremendously insightful generalization from such specific observations he developed the concept that the ability of an organism to maintain a constant fluid environment bathing cells of the body—the “milieu intérieur”—is essential for life independent of the external environment.
Cannon (Citation1929a,Citationb, Citation1939) subsequently coined the term “homeostasis” to describe the maintenance within acceptable ranges of several physiological variables, such as blood glucose, oxygen tension and core temperature. Mechanisms for maintaining this stability require sensors to recognize discrepancies between the sensed and set acceptable values and require effectors that reduce those discrepancies—i.e., negative feedback systems. For instance, when the core temperature of a mammal rises, the thermoregulatory system evokes sweating and diversion of blood flow from the viscera to the skin, which enhances heat loss; and when core temperature falls, shivering increases heat production, while cutaneous vasoconstriction diminishes heat loss by diversion of blood to internal organs. Analogous negative feedback systems regulate other physiological variables. Cannon extended this concept to include psychosocial threats to homeostasis. In the early 1900s he described for the first time the acute changes in adrenal gland secretion associated with what he called “fight or flight” responses.
According to Cannon, a wide variety of threats to homeostasis, such as exposure to cold, hypotensive hemorrhage, traumatic pain, insulin-induced hypoglycemia, or emotional distress, elicit activation of the adrenal medulla and sympathetic nervous system. These two effectors were thought to function as a unit—termed the “sympathoadrenal” or “sympathico-adrenal” system—to restore homeostasis. Rapid activation of the sympathoadrenal system then preserves the internal environment by producing compensatory and anticipatory adjustments that enhance the likelihood of survival. In 1939, Cannon formally proposed adrenaline (called epinephrine in the USA) as the active principle of the adrenal gland and also as the neurotransmitter of the sympathetic nervous system (Cannon and Lissak Citation1939), consistent with the functional unity of the sympathoadrenal system. The identity of the substance released at sympathetic nerve terminals remained controversial until 1946, when von Euler (Citation1946) correctly identified noradrenaline as the major sympathetic neurotransmitter in mammals.
Cannon (Citation1929a,Citationb, Citation1939) emphasized that disparate threats to homeostasis would incite the same sympathoadrenal response. References to the sympathoadrenal or “adrenergic” system persist (Cryer Citation1980; Shah et al. Citation1984; Sofuoglu et al. Citation2001; Dronjak et al. Citation2004; Vogel et al. Citation2005), although researchers have come to recognize that sympathetic nervous and adrenomedullary hormonal system activities can change differentially in different forms of stress, as discussed below.
In the sheltered confines of a laboratory, with controlled temperature and ad libitum water, nutrients and calories, mammals do not seem to require an intact sympathetic nervous system (Cannon Citation1931). It has become clear, however, that even under resting conditions, pulse-synchronous bursts of skeletal muscle sympathetic nerve activity and plasma levels of noradrenaline are detectable, and noradrenaline continuously enters the venous drainage of most organs. We also now recognize that activities of daily life, such as meal ingestion (Patel et al. Citation2002), public speaking (Gerra et al. Citation2001), changing posture (Lake et al. Citation1976), and locomotion—i.e. not only emergencies—are associated with rapid adjustments in sympathetic nervous system outflows. Each of these activities is associated with a somewhat different set of apparent steady-states, directed by the brain and determined by coordinated actions of a variety of effector systems. These observations contributed to the development of the concept of “allostasis,” discussed below.
Selye, the doctrine of nonspecificity, and the hypothalamic-pituitary-adrenocortical system
We are indebted to Selye (Citation1956) for popularizing the concept of stress. Selye redefined the word, stress, from its meaning in the physical world as a force that results in a deformity and results in strain, the opposing force tending to restore the unstressed state. His definition of stress as being (or a state resulting in) the “the nonspecific response of the body to any demand upon it” (Selye Citation1974) was so persuasive that it persisted and remains widely used today. By “nonspecific” Selye meant a set of shared elements of responses—regardless of the nature of the causative agent, or stressor.
Selye proposed three universal stages of coping with a stressor—the “General Adaptation Syndrome”—an initial “alarm reaction,” analogous to Cannon's “fight or flight” response, a stage of adaptation, associated with resistance to the stressor, and eventually a stage of exhaustion and organismic death. In Selye's early experiments, after injection of any of a variety of tissue extracts or of formalin into rats, the animals developed a pathological triad of enlargement of the adrenal glands, atrophy of lymphoid tissue in the thymus, spleen, and lymph nodes and bleeding gastrointestinal ulcers. It was later demonstrated that these changes are associated with, and to at least some extent result from, activation of the hypothalamic-pituitary-adrenocortical (HPA) axis. Steroids released into the circulation from the adrenal cortex contribute to resistance but may also be responsible for pathological changes. Selye's concept that prolonged stress can produce physical disease and mental disorders is now widely accepted.
Selye acknowledged that responses to stressors have specific components that tend to reverse the effects of the stressor; however, in addition to the specific responses, there is a nonspecific stress syndrome. Chrousos and Gold (Citation1992) modified Selye's doctrine of nonspecificity, by proposing that above a threshold intensity, any stressor elicits the nonspecific stress syndrome.
More than a half century elapsed before Selye's doctrine of nonspecificity underwent experimental testing, which failed to confirm it (Pacak et al. Citation1998). Nevertheless, modern lay and even scientific literature continues to accept the notion of a unitary stress response. For instance, a Google search yielded about 582,000 hits for “the stress responses”. According to WebMD, “The stress response is the set of physical and emotional changes the human body makes in response to a threat or stress. It sometimes is called the ‘fight-or-flight’ response” (as indicated above, it was Cannon who introduced the latter phrase).
Just as Cannon emphasized activation of the sympathoadrenal system to maintain homeostasis, so did Selye and his students emphasize activation of the HPA axis in the General Adaptation Syndrome. Selye (Citation1956) also introduced the term “heterostasis” (from the Greek heteros = other) to describe the establishment of a new steady-state by changing the “setpoint” to resist unusually high demands. This new steady-state, however, is attained by treatment with remedies that have no direct curative action but enhance the body's natural defenses, e.g. immunization to combat infection (as in treatment of rabies). The concept of changes in the homeostatic setpoint as a natural adaptive mechanism awaited the introduction of the notion of allostasis.
Modern concepts of stress
More modern concepts view stress as a consciously or unconsciously sensed threat to homeostasis (McEwen et al. 1993; Goldstein and McEwen Citation2002), in which the response has a degree of specificity, depending, among other things, on the particular challenge to homeostasis, the organism's perception of the stressor and the perceived ability to cope with it (Goldstein Citation2001).
Although homeostasis suggests constancy of values for variables, ranges of acceptable values are now recognized to be decidedly inconstant. There are diurnal variations in body temperature, heart rate and blood pressure. In addition, adaptations to different stressors include alterations in acceptable levels for monitored variables. Sterling and Eyer introduced the term, “allostasis,” to describe the attainment of stability by natural alterations in acceptable ranges of variables attending the adjustments of the cardiovascular system during rest and activity (Sterling et al. Citation1988). Such alterations are distinct from treatment-induced alterations (Selye's “heterostasis” discussed above). The brain is the site at which effects of stressors are sensed and appropriate coordinated behavioral and neuroendocrine responses initiated. Adaptations involving allostasis to cope with real, simulated, or imagined challenges are determined by genetic, developmental and previous experiential factors. While they may be effective for a short interval, over time the alterations may have cumulative adverse effects. For instance, chronic elevation of blood pressure to ensure adequate blood flow to the brain might eventually lead to atherosclerosis and stroke or coronary occlusion. Risk of such adverse effects is termed “allostatic load.”
Principles of operation of homeostatic systems
Central to a systems concept of stress is that the body possesses numerous homeostatic comparators, which have been called “homeostats” (Goldstein Citation1995a,Citationb). Each homeostat compares information supplied by a sensor with a setpoint for responding, determined by a regulator or set of regulator mechanisms (). A sufficiently large sensed discrepancy between afferent information about the level of the monitored variable and the setpoint for responding elicits altered activities of effectors, the actions of which decrease the discrepancy in the monitored variable.
Figure 1 A systems definition of stress.
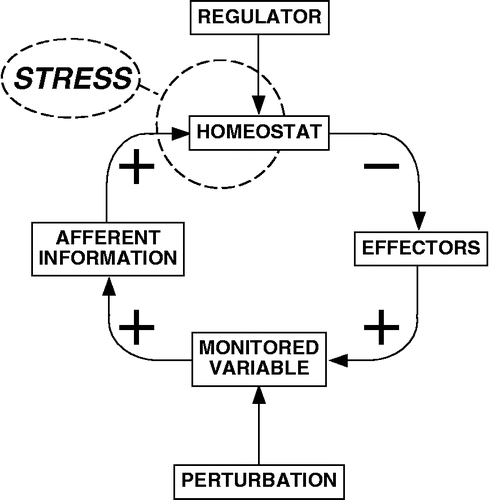
Homeostatic systems operate according to a few principles that despite their simplicity can explain some complex physiological phenomena and might help resolve controversial issues in the area of stress and disease.
Negative feedback regulation
In a home temperature control system, the thermostat plays a central role, by sensing discrepancy between the setpoint, determined by a regulator (the home owner) and the temperature, which produces differential bending of metal bands in the thermostat (the sensor). This type of system is a classical example of regulation by negative feedback. Relationships among components of homeostatic systems can be positive or negative; in a negative feedback loop, there is an odd number of negative relationships in the loop (note the odd number of ( − ) signs in ). It can be shown mathematically that in response to a constant perturbing influence (e.g. cold external temperature), a system regulated by negative feedback yields a stable level of the monitored variable somewhere between the sensed and set values.
Physiological homeostatic systems also include regulation by negative feedback. Increases in values of the monitored variable result in changes in effector activity that oppose and thereby “buffer” changes in that variable. This feedback regulation can be modulated at several levels and therefore can be quite complex.
A large array of homeostatic systems detect perturbations of monitored variables. In line with the home heating analogy, this includes afferent information to the brain about cutaneous and blood temperature, leading to altered activities of cholinergic and noradrenergic nerve fibers in the skin that regulate sweating and vasomotor tone (Frank et al. Citation1997).
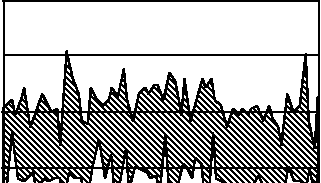
Disruption of a negative feedback system, such as by blockade of afferent information or by dysfunction of a homeostat, increases the variability of levels of the monitored variable. Thus, baroreceptor deafferentiation increases the variability of blood pressure, as does bilateral destruction of the nucleus of the solitary tract, the likely brainstem site of the arterial barostat (Nathan and Reis Citation1977). As shown in , in humans, treatment of head and neck cancer by local irradiation accelerates carotid arteriosclerosis, and encasement of baroreceptors in rigidified carotid sinuses can help explain the development of labile blood pressure years later (Sharabi et al. Citation2003).
Figure 2 Twenty-four hour ambulatory blood pressure monitoring records for a control subject and three patients (Pt) with baroreflex failure as a late sequela of neck irradiation.
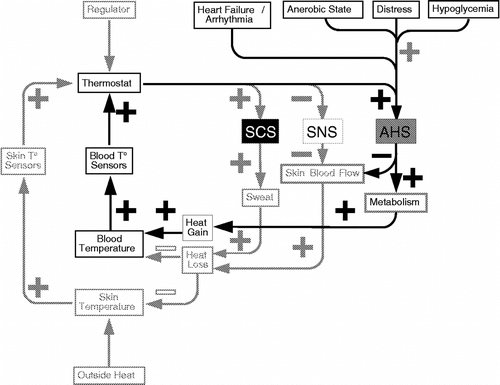
Induction of a positive feedback loop threatens homeostasis, by accelerating changes in levels of the monitored variable. Examples of positive feedback loops include tilt-induced neurally mediated syncope and heat shock. A pattern of adrenomedullary hormonal system stimulation and sympathetic noradrenergic system inhibition often precedes fainting (Goldstein et al. Citation2003), as indicated by an accelerating increase in the plasma adrenaline level and concurrent decline in forearm vascular resistance (). Ordinarily, humans can tolerate even extremely hot external temperature, by evaporative heat loss, promoted by thermoregulatory sweating mediated by the sympathetic cholinergic system and cutaneous vasodilation mediated by local sympathetic noradrenergic inhibition (). In this setting plasma noradrenaline levels increase, probably reflecting increased sympathetic outflows to visceral organs and skeletal muscle, promoting shunting of blood from the core to the periphery, while plasma adrenaline levels change relatively little (Kukkonen-Harjula et al. Citation1989; Gisolfi et al. Citation1991; Brenner et al. Citation1997; Niimi et al. Citation1997); however, activation of the adrenomedullary hormonal system, such as by exceeding an anerobic threshold during exercise, dehydration, hypoglycemia, emotional distress and myocardial dysfunction, might interfere with evaporative heat loss and also induce a positive feedback loop, with rapid precipitation of heat shock and even death ().
Figure 3 Values for arterial plasma adrenaline (EPI), noradrenaline (NE), forearm vascular resistance (FVR) and mean arterial pressure (MAP) in a patient with tilt-induced syncope. Grey arrows highlight changes in EPI and FVR preceding hypotension and syncope.

Figure 4 Overview of thermoregulatory mechanisms. SCS = sympathetic cholinergic system; SNS = sympathetic noradrenergic system.
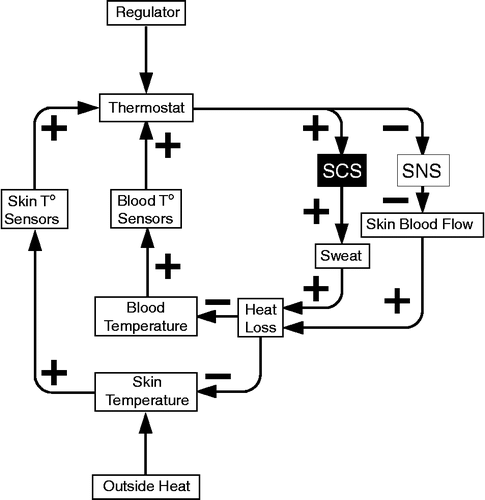
Figure 5 Hypothetical changes associated with heat shock. AHS = adrenomedullary hormonal system. Note positive feedback loop, indicated by a circuit with all (+) signs, as well as interference with a negative feedback loop, indicated by an even number of ( − ) signs.
There are some examples of beneficial effects of a positive feedback loop. For instance, if bleeding is to be terminated by blood clotting, acceleration of clot formation by its own initiation is a useful positive feedback process. While distress can be a protective, such as by initiating flight, distress can also be involved in positive feedback. If cognitive recognition of the effects of released adrenaline enhances anxiety and promotes further adrenaline release, the results could be disastrous.
Multiple effectors
Home temperature control systems usually include multiple effectors. The redundancy comes at relatively little cost, compared with three advantages. The multiplicity extends the range of external temperatures that can be tolerated to maintain constant the internal temperature; when a single effector fails to function, others are activated to compensate, helping to maintain the temperature at about the set level; and one can pattern the use of the effectors as appropriate to maximize economy and efficiency (Goldstein Citation2001; Goldstein 2006).
Analogously, physiological homeostatic systems frequently use multiple effectors to govern values for the monitored variable. An example would be insulin, glucagon, adrenaline, cortisol and growth hormone in regulation of blood glucose levels. Effector redundancy extends the ranges of control of monitored variables.
Compensatory activation
Having multiple effectors enables compensatory activation of alternative effectors, assuming no change in homeostat settings (). Examples of compensatory activation in physiology include augmentation of sympathoneural responsiveness by adrenalectomy, hypophysectomy, or thyroidectomy (Udelsman et al. Citation1987; Goldstein et al. Citation1993; Fukuhara et al. Citation1996).
Figure 6 Model of compensatory activation of the sympathetic noradrenergic system (SNS) by thyroidectomy. THY: thyroid; TEMP: temperature.
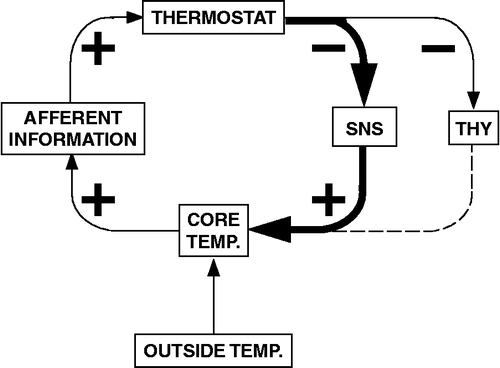
Compensatory activation of alternative vasoactive systems after destruction of the sympathetic noradrenergic system (Gauthier et al. Citation1972; Julien et al. Citation1990) helps to explain why many workers, including Cannon, erroneously concluded that the sympathetic nervous system acts only as an “emergency” system.
Stressor-specificity
After adequately sensitive assay methods of plasma levels of noradrenaline and adrenaline became available, evidence rapidly accumulated for different noradrenergic vs. adrenergic responses in different situations (Robertson et al. Citation1979; Young and Landsberg Citation1979; Cryer Citation1980; Young et al. Citation1984). A new concept began to emerge, in which noradrenaline levels, and thereby overall sympathetic nervous “activity,” would play key roles in appropriate distribution of blood volume and homeostasis of blood pressure (or blood delivery to the brain), such as during orthostasis, cold exposure, mild blood loss, locomotion, exercise, altered salt intake and water immersion. Adrenaline levels, reflecting the adrenomedullary hormonal system “activity,” respond to global or metabolic threats, such as hypoglycemia, hemorrhagic hypotension, exercise beyond an anerobic threshold, asphyxiation, emotional distress and shock. Evidence also accumulated for an association between noradrenaline and active escape, avoidance, or attack, and an association between adrenaline and passive, immobile fear.
Thus, in contrast with Selye's doctrine of nonspecificity, according to the allostatic concept of stress responses, acceptable activities of effector systems are coordinated in relatively specific patterns, including neuroendocrine patterns. These patterns serve different needs, and the sympathetic nervous and adrenomedullary hormonal systems play important roles in many of them. For instance, the level of sympathetic nervous system activation predominates in response to orthostasis, moderate exercise and exposure to cold, whereas the level of adrenomedullary hormonal system activation predominates in response to glucoprivation and emotional distress.
In terms of the body's thermostat, studies of humans exposed to cold or with mild core hypothermia have provided support for the notion of primitive specificity of neuroendocrine stress responses. Cold exposure increases plasma noradrenaline levels, with little if any increases in plasma adrenaline levels, consistent with sympathetic neuronal activation and relatively less adrenomedullary hormonal activation. Mild core hypothermia also increases antecubital venous levels of noradrenaline but not adrenaline (Frank et al. Citation2002). Both noradrenaline and adrenaline levels in arterial plasma increase in this setting, but with larger noradrenaline responses (Goldstein and Frank Citation2001). These findings make sense, in that one can maintain body temperature effectively by sympathetically-mediated cutaneous vasoconstriction, piloerection and shivering. When these mechanisms give way, and core temperature falls, then high circulating adrenaline levels increase generation of calories (Staten et al. Citation1987), associated with the experience of distress, which motivates escape and avoidance, and augments noradrenaline release from sympathetic nerve terminals for a given amount of nerve traffic (Chang et al. Citation1994).
Effector redundancy introduces the potential for patterned effector responses. Patterning of neuroendocrine, physiological and behavioral effectors increases the likelihood of adaptation to the particular challenge to homeostasis, providing another basis for natural selection to favor the evolution of systems with multiple effectors. As summarized in , across a range of stressors, adrenomedullary hormonal system activation seems to correlate better with HPA axis activation than with sympathetic nervous system activation, in contrast with Cannon's concept of a unitary sympathoadrenal system. Differential changes in adrenomedullary and sympathoneural outflows are particularly clear in the setting of cold exposure, which features sympathetic noradrenergic activation, and fainting or hypoglycemia, which feature adrenomedullary hormonal system activation. Thus, “sympathoadrenal imbalance,” with high adrenaline levels and smaller or no increases in noradrenaline levels, often precedes fainting (Goldstein et al. Citation2003); and in response to hypoglycemia, increases in plasma noradrenaline levels result entirely from adrenomedullary stimulation, since elevated plasma noradrenaline does not occur in adrenalectomized subjects (DeRosa et al. Citation2004).
Figure 7 Relative intensities of activation of the HPA axis, adrenomedullary hormonal system (AHS) and sympathetic noradrenergic system (SNS) during exposure to different stressors, based on literature review.

For each stressor, neuroendocrine and physiological changes are coupled with behavioral changes. For instance, the regulation of total body water in humans depends on an interplay between behavior (the search for water and drinking), an internal experience or feeling (thirst), and the elicitation of a neurohumoral response pattern (in this case dominated by vasopressin, the antidiuretic hormone; and to a lesser extent angiotensin, a potent stimulator of drinking). Evoked changes in homeostat function often produce not only neuroendocrine and physiological effects but also behavioral responses; however, because of traditional boundaries among physiology, endocrinology and psychology, interactions producing integrated patterns of response remain incompletely understood.
Selye recognized that during the stage of resistance, exposure to a novel stressor can produce an exaggerated response. This observation presaged the concept of “dishabituation,” in which neuroendocrine responses to stressors depend on the prior experience of the organism with respect to qualitatively different stressors (Graessler et al. Citation1989; Nisenbaum et al. Citation1991; Abercrombie Citation1992; Rusnak et al. Citation2001; Kvetnansky et al. Citation2003; Dronjak et al. Citation2004; Kvetnansky Citation2004; Radikova et al. Citation2004; Liu et al. Citation2005).
Effector sharing
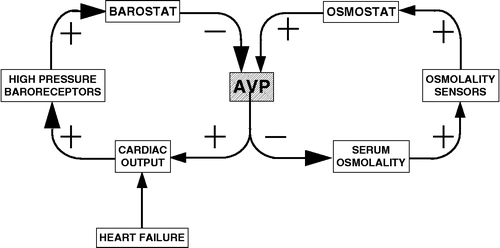
Different homeostats can regulate the activity of the same effector system. For instance, the osmostat and barostat share the arginine vasopressin effector (Quillen and Cowley Citation1983), illustrated in .
Figure 8 Model of effector sharing. The “barostat” and “osmostat” share the arginine vasopressin (AVP) effector.
Systems definition of stress
Even a simple homeostatic reflex reflects stress, when a perceived discrepancy between a setpoint for a monitored variable and information about the actual level of that variable elicits compensatory responses to decrease the discrepancy. Thus, one way of looking at stress is as a condition where expectations, whether genetically programmed, established by prior learning, or deduced from circumstances, do not match the current or anticipated perceptions of the internal or external environment, and this discrepancy between what is observed or sensed and what is expected or programmed elicits patterned, compensatory responses ().
Distress
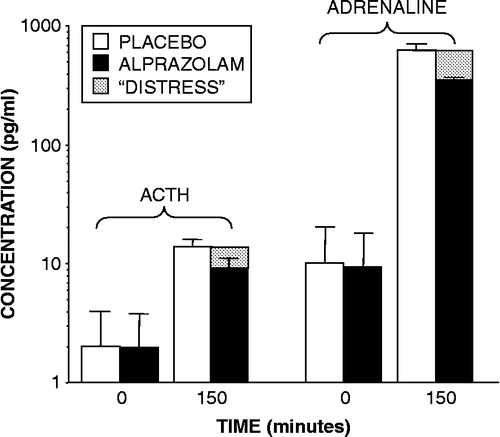
Distress is cognitive recognition of a condition that is aversive to the organism, as evidenced by motivation for learning to escape or avoid the stressor. Distress is a subclass of stress that depends on the character, intensity, and meaning of the stressor as perceived by the organism and on the organism's perceived ability to cope with it. According to this concept, for an organism to experience distress requires consciousness, to interpret the situation in terms of the ability to cope. This notion can help explain the finding () that sedation with the benzodiazepine, alprazolam, attenuates the ACTH and adrenaline responses to glucoprivation (Breier et al. Citation1992).
Figure 9 Plasma concentrations of corticotropin (ACTH) and adrenaline before and at 150 minutes after injection of 2-deoxyglucose into healthy volunteers, in the context of pretreatment with placebo or alprazolam. Differences between placebo and alprazolam conditions might reflect distress (gray rectangles).
Selye (Citation1974) characterized distress as unpleasant or harmful. The systems view of stress does not assume an equivalence of noxiousness (i.e. negatively reinforcing properties) with production of pathological changes; that is, it does not assume that distress causes disease.
Homeostatic responses do not necessarily cause distress, but distress can be part of a positive feedback loop. This applies not only to neuroendocrine aspects of those responses (such as the glucose counter-regulatory actions of pituitary-adrenocortical and adrenomedullary stimulation during insulin-induced hypoglycemia) but also to psychological aspects (such as conditioned aversive and instrumental avoidance learning). Distress responses evolved and probably continue to be expressed even in higher organisms, including humans who actually are only rarely exposed to truly “fight-or-flight” agonistic encounters, because of the importance of those responses in instinctive communication.
Allostasis and allostatic load
Levels of physiological activity required to re-establish or maintain homeostasis differ, depending on continually changing conditions in which the organism finds itself—e.g., running vs. standing vs. lying down. “Allostasis,” a term used by Sterling and Eyer in 1988 (McEwen Citation1998), refers to levels of activity required for the individual to “maintain stability through change”—i.e., to adapt (McEwen Citation1998; Schulkin et al. Citation1998; McEwen Citation2000).
In terms of the present theory, “allostasis” refers to the set of apparent steady-states maintained by multiple effectors. In the analogy of the home temperature control system, one can regulate temperature at different levels, by appropriate use of effectors. Among individuals, levels of glucose, blood pressure, body temperature, metabolism, and so forth are normally held stable at different levels, with different patterns of effector activation.
Homeostat resetting redefines the conditions required to maintain homeostasis. Regulation around an altered apparent steady-state is the essence of allostasis. This would be analogous to a different thermostatic setting in the home in winter compared to the summer. Resetting alters activities of multiple effector systems required to maintain allostasis, at least for short durations. Allostatic settings can change dynamically. An example is the regulation of blood glucose in the context of exercise. Even in anticipation of the need for metabolic fuel, by activation of “central command” the blood glucose level tends to increase; during light to moderate exercise, increased glucose production typically balances increased glucose utilization, so that blood glucose does not change; and intense exercise is often associated with hyperglycemia.
During stress, short-term changes in homeostatic settings generally enhance the long-term well-being and survival of the organism. Responses during exercise provide an obvious example. When superimposed on a substrate of pathology, however, homeostatic resetting can cause harm. For instance, in the setting of ischemic heart disease, global or patterned increases in sympathetic outflows from homeostat resetting would increase cardiac work, the resulting imbalance between oxygen supply and demand precipitating angina pectoris, myocardial infarction, or sudden death.
Allostatic load
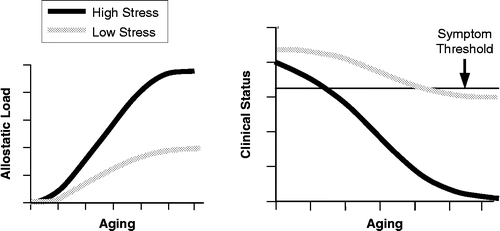
“Allostatic load” (McEwen and Stellar Citation1993) refers to effects of prolonged continuous or intermittent activation of effectors involved in allostasis. In the analogy of the home temperature control system, allostatic load would increase if a window or door were left open. In this situation, one or more effectors might be activated frequently or even continuously. An even more extreme example would be having the air conditioner and the furnace on at the same time, as is in an apartment in the spring when there is a warm day before the boilers have been shut down. Continued use of the furnace and air conditioner in opposition to one another, an example of an inefficient “allostatic state,” consumes fuel and contributes to wear-and-tear on both pieces of equipment. Long-term allostatic load—the wear-and-tear cost of adaptation—provides a conceptual basis for studying long-term health consequences of stress ().
Figure 10 Predicted effects of stress and aging on allostatic load and clinical status, based on a kinetic model.
Medical and psychological consequences of stress
As noted above, induction of a positive feedback loop in a homeostatic system evokes a type of instability in which levels of the monitored variable accelerate. An example would be renin-angiotensin-aldosterone system activation in congestive heart failure. Activation of this system increases sodium retention and vascular tone, leading to increased cardiac preload and afterload that worsen the congestive heart failure. Therefore, treatment with an angiotensin-converting-enzyme inhibitor or angiotensin II receptor blocker can successfully treat congestive heart failure (Kluger et al. Citation1982).
Another example may be fainting reactions (). Fainting is preceded by high circulating adrenaline levels and yet withdrawal of sympathetic vasoconstrictor tone (Mosqueda-Garcia et al. Citation1997; Goldstein et al. Citation2003). This elicits skeletal muscle vasodilation, and total peripheral resistance to blood flow falls. If there were enough “shunting” of blood to the skeletal muscle, then blood flow to the brainstem might fall. The person would not feel “right.” This could evoke more adrenomedullary secretion of adrenaline, and the consequent neurocirculatory positive feedback loop would lead to critical brainstem hypoperfusion and loss of consciousness within seconds to minutes.
In people who faint repeatedly, it is often the case that between episodes they do not feel completely normal. Patients who are susceptible to neurocardiogenic syncope may complain of chronic fatigue, headache, chest pain, orthostatic intolerance, difficulty concentrating and heat intolerance, which can be debilitating. The patients also have tonic suppression of noradrenaline spillover from the heart (Goldstein et al. Citation2002). In essence this may reflect consequences of long-term allostatic load, as discussed below.
Induction of a positive feedback loop “nested” in a larger system that includes negative feedback can lead to a new steady-state group of settings and values for monitored variables, rather than “explosion” of the system. For example, a distressing situation might elicit fear, resulting in release of noradrenaline in the brain and adrenaline in the periphery, both of which could augment vigilance behavior and heighten the experience of distress, resulting in greater fear (Aston-Jones et al. Citation1994). The organism could enter an “escape mode,” with a different set of homeostatic regulatory settings; however, there is a risk of the positive feedback loop leading to a behavioral “explosion,” panic, or a pathophysiologic “explosion,” pulmonary edema. The notion of induction of a nested positive feedback loop can also provide a model for developmental changes in adolescence, where stability would actually be abnormal, but there is a greater chance for both psychological and physiological disorders to emerge.
The systems view of stress and the concept of allostasis can help understanding of chronic as well as acute medical consequences of stress. Chronic activation of allostatic effectors in allostatic states promotes allostatic load. For instance, chronic elevations in adrenomedullary and HPA outflows might worsen insulin resistance, and chronic cardiac sympathetic activation might accelerate cardiovascular hypertrophy and development of heart failure (Rundqvist et al. Citation1997).
Another application of the homeostatic idea to medical consequences of stress is in terms of the perceived ability to cope. As noted above, an organism experiences distress upon sensing that the effector responses will not be sufficient to restore or maintain allostasis. In contrast with distress, stress does not imply a conscious experience. For instance, even heavily sedated humans have substantial adrenomedullary stimulation in response to acute glucoprivation. Indeed, the greater extent of the adrenomedullary response to the same stressor in alert than in sedated humans might provide a measure of the distress. Distress instinctively elicits observable signs and pituitary-adrenocortical and adrenomedullary activation (Goldstein Citation1995a,Citationb; Goldstein Citation2001). Via these neuroendocrine changes, distress could worsen pathophysiologic processes. For instance, because of adrenomedullary activation, in a patient with coronary artery stenosis distress could elicit cardiovascular stimulation and produce an excess of myocardial oxygen consumption over supply, precipitating myocardial infarction or lethal ventricular arrhythmias. Moreover, long-term distress could augment both the risk of a mood disorder and the risk of worsening coronary artery disease.
Long-term physical or mental consequences of stress would depend on long-term effects of allostatic load. Prolonged, intensive activation of effector systems could exaggerate effects of intrinsic defects in any of them, just as increased air pressure in a tyre could expand and eventually “blow out” a weakened area. It is not difficult to imagine that repeated or long-term stress or distress could lead to a medical or psychiatric “blowout.”
Maintenance of allostatic states requires energy. This requirement is perhaps clearest in allostasis of core temperature. In mammals, maintenance of a constant core temperature accounts for a substantial proportion of total body energy expenditure at rest. One may hypothesize that reducing allostatic load exerts beneficial health effects, just as one may hypothesize that excessive allostatic load exerts deleterious health effects. In the analogy of the home temperature control system, maintaining a temperature of 66 degrees Fahrenheit in the summer would require a great expenditure of energy and involve cooling systems being on continuously, whereas in the winter, maintaining the same temperature would be energy-efficient. One can imagine that the likelihood of system breakdown would depend on the extent of long-term energy use by the effector systems.
Chronic effector system activation might alter the efficiency of the homeostatic system itself. For instance, chronic sympathetic nervous stimulation of the cardiovascular system could promote cardiovascular hypertrophy, “splinting” arterial baroreceptors in stiff blood vessel walls, in turn contributing to systolic hypertension and the risk of heart failure, kidney failure and stroke.
Moreover, an inappropriately large adrenomedullary response to a stressor might exaggerate the experience of emotional distress (Schachter and Singer Citation1962). Exaggerated distress responses might increase the risk of worsening an independent pathologic process, such as in panic-induced angina pectoris (Mansour et al. Citation1998; Wilkinson et al. Citation1998).
In summary, this essay describes the evolution from Bernard's concept of stability of the internal environment to present concepts of stress. Cannon described homeostasis through feedback regulation of physiological processes. Selye popularized the notion of stress and emphasized the nonspecific responses to stressors. He described pathological changes with severe or prolonged stress. Subsequent studies revealed arrays of specific mechanisms enlisted to respond to perceived threats to stability and safety. The importance of changes in homeostatic requirements as a means of adaptation to stressors emerged in the concept of allostasis, and potentially deleterious effects of allostasis as allostatic load. Multiple systems are required to meet the needs for allostasis. Genetic, environmental and experiential factors are major determinants of mechanisms and outcomes of allostatic load. These more recent concepts provide a basis for explaining and predicting physical and psychiatric effects of acute and chronic stress.
Stress is an interdisciplinary topic, and understanding health consequences of stress requires an integrative approach. Research and ideas about stress must move beyond considering only one effector system, such as the “sympathoadrenal system”, and only one monitored variable, such as serum glucose levels, to incorporate multiple effectors and multiple homeostatic systems that are regulated in parallel. They must also move beyond the notion of a single set of ideal values for monitored variables—homeostasis—to incorporate dynamic changes in homeostatic settings—allostasis. It has become evident that allostasis usually involves multiple systems, and more complete understanding of stress requires a full appreciation of these systems and their interactions. Merging of the homeostatic definitions of stress and distress with the concept of allostasis should provide a better understanding of the roles of stress and distress in chronic diseases and also provide a conceptual basis for the further development of scientific integrative medicine.
References
- Abercrombie ED. Enhanced tyrosine hydroxylation in hippocampus of chronically stressed rats upon exposure to a novel stressor. J Neurochem 1992; 58: 276–281
- Aston-Jones G, Rajkowski J, Kubiak P, Alexinsky T. Locus coeruleus neurons in monkey are selectively activated by attended cues in a vigilance task. J Neurosci 1994; 14: 4467–4480
- Breier A, Davis O, Buchanan R, Listwak S, Holmes C, Pickard D, Goldstein D. Effects of alprazolam on pituitary-adrenal and catecholaminergic responses to metabolic stress in humans. Biol Psychiatry 1992; 32: 880–890
- Brenner IK, Zamecnik J, Shek PN, Shephard RJ. The impact of heat exposure and repeated exercise on circulating stress hormones. Eur J Appl Physiol Occup Physiol 1997; 76: 445–454
- Cannon WB. Bodily changes in pain, hunger, fear and rage. D. Appleton & Co, New York 1929a
- Cannon WB. Organization for physiological homeostasis. Physiol Rev 1929b; 9: 399–431
- Cannon WB. The effects of progressive sympathectomy on blood pressure. Am J Physiol 1931; 97: 592–595
- Cannon WB. The wisdom of the body. W.W. Norton, New York 1939
- Cannon WB, Lissak K. Evidence for adrenaline in adrenergic neurones. Am J Physiol 1939; 125: 765–777
- Chang PC, Grossman E, Kopin IJ, Goldstein DS. On the existence of functional beta-adrenoceptors on vascular sympathetic nerve endings in the human forearm. J Hypertens 1994; 12: 681–690
- Chrousos GP, Gold PW. The concepts of stress and stress system disorders. Overview of physical and behavioral homeostasis. J Am Med Assoc 1992; 267: 1244–1252
- Cryer PE. Physiology and pathophysiology of the human sympathoadrenal neuroendocrine system. N Engl J Med 1980; 303: 436–444
- DeRosa MA, Cryer PE. Hypoglycemia and the sympathoadrenal system: Neurogenic symptoms are largely the result of sympathetic neural, rather than adrenomedullary, activation. Am J Physiol Endocrinol Metab 2004; 287: E32–41
- Dronjak S, Jezova D, Kvetnansky R. Different effects of novel stressors on sympathoadrenal system activation in rats exposed to long-term immobilization. Ann N Y Acad Sci 2004; 1018: 113–123
- Frank SM, Higgins MS, Fleisher LA, Sitzmann JV, Raff H, Breslow MJ. Adrenergic, respiratory, and cardiovascular effects of core cooling in humans. Am J Physiol 1997; 272: R557–R562
- Frank SM, Cattaneo CG, Wieneke-Brady MB, El-Rahmany H, Gupta N, Lima JA, Goldstein DS. Threshold for adrenomedullary activation and increased cardiac work during mild core hypothermia. Clin Sci 2002; 102: 119–125
- Fukuhara K, Kvetnansky R, Cizza G, Pacak K, Ohara H, Goldstein DS, Kopin IJ. Interrelations between sympathoadrenal system and hypothalamo-pituitary-adrenocortical/thyroid systems in rats exposed to cold stress. J Neuroendocrinol 1996; 8: 533–541
- Gauthier P, Nadeau R, De Champlain J. Acute and chronic cardiovascular effects of 6-hydroxydopamine in dogs. Circ Res 1972; 31: 207–217
- Gerra G, Zaimovic A, Mascetti GG, Gardini S, Zambelli U, Timpano M, Raggi MA, Brambilla F. Neuroendocrine responses to experimentally-induced psychological stress in healthy humans. Psychoneuroendocrinology 2001; 26: 91–107
- Gisolfi CV, Matthes RD, Kregel KC, Oppliger R. Splanchnic sympathetic nerve activity and circulating catecholamines in the hyperthermic rat. J Appl Physiol 1991; 70: 1821–1826
- Goldstein DS. Stress as a scientific idea: A homeostatic theory of stress and distress. Homeostasis 1995a; 4: 177–215
- Goldstein DS. Stress, catecholamines, and cardiovascular disease. Oxford University Press, New York 1995b
- Goldstein DS. The autonomic nervous system in health and disease. Marcel Dekker, Inc, New York, NY 2001
- Goldstein DS, Frank SM. The wisdom of the body revisited: The adrenomedullary response to mild core hypothermia in humans. Endocrine Regul 2001; 35: 3–7
- Goldstein DS, Garty M, Bagdy G, Szemeredi K, Sternberg EM, Listwak S, Deka-Starosta A, Hoffman A, Chang PC, Stull R, Gold PW, Kopin IJ. Role of CRH in glucopenia-induced adrenomedullary activation in rats. J Neuroendocrinol 1993; 5: 475–486
- Goldstein DS, Holmes C, Frank SM, Dendi R, Cannon RO, III, Sharabi Y, Esler MD, Eisenhofer G. Cardiac sympathetic dysautonomia in chronic orthostatic intolerance syndromes. Circulation 2002; 106: 2358–2365
- Goldstein DS, Holmes C, Frank SM, Naqibuddin M, Dendi R, Snader S, Calkins H. Sympathoadrenal imbalance before neurocardiogenic syncope. Am J Cardiol 2003; 91: 53–58
- Goldstein DS, McEwen B. Allostasis, homeostats, and the nature of stress. Stress 2002; 5: 55–58
- Goldstein DS. Adrenaline and the Inner World: An Introduction to Scientific Integrative Medicine. The Johns Hopkins University Press, Baltimore, MD 2006
- Graessler J, Kvetnansky R, Jezova D, Dobrakovova M, Van Loon GR. Prior immobilization stress alters adrenal hormone responses to hemorrhage in rats. Am J Physiol 1989; 257: R661–R667
- Julien C, Kandza P, Barres C, Lo M, Cerutti C, Sassard J. Effects of sympathectomy on blood pressure and its variability in conscious rats. Am J Physiol 1990; 259: H1337–H1342
- Kluger J, Cody RJ, Laragh JH. The contributions of sympathetic tone and the renin-angiotensin system to severe chronic congestive heart failure: Response to specific inhibitors (prazosin and captopril). Am J Cardiol 1982; 49: 1667–1674
- Kukkonen-Harjula K, Oja P, Laustiola K, Vuori I, Jolkkonen J, Siitonen S, Vapaatalo H. Haemodynamic and hormonal responses to heat exposure in a Finnish sauna bath. Eur J Appl Physiol Occup Physiol 1989; 58: 543–550
- Kvetnansky R. Stressor specificity and effect of prior experience on catecholamine biosynthetic enzyme phenylethanolamine N-methyltransferase. Ann N Y Acad Sci 2004; 1032: 117–129
- Kvetnansky R, Rusnak M, Dronjak S, Krizanova O, Sabban EL. Effect of novel stressors on tyrosine hydroxylase gene expression in the adrenal medulla of repeatedly immobilized rats. Neurochem Res 2003; 28: 625–630
- Lake CR, Ziegler MG, Kopin IJ. Use of plasma norepinephrine for evaluation of sympathetic neuronal function in man. Life Sci 1976; 18: 1315–1325
- Liu X, Kvetnansky R, Serova L, Sollas A, Sabban EL. Increased susceptibility to transcriptional changes with novel stressor in adrenal medulla of rats exposed to prolonged cold stress. Brain Res Mol Brain Res 2005; 141: 19–29
- Mansour VM, Wilkinson DJ, Jennings GL, Schwarz RG, Thompson JM, Esler MD. Panic disorder: Coronary spasm as a basis for cardiac risk?. Med J Australia 1998; 168: 390–392
- McEwen BS. Stress, adaptation, and disease. Allostasis and allostatic load. Ann NY Acad Sci 1998; 840: 33–44
- McEwen B, Stellar E. Stress and the individual. Mechanisms leading to disease. Arch Int Med 1993; 153: 2093–2101
- McEwen BS. Allostasis and allostatic load: Implications for neuropsychopharmacology. Neuropsychopharmacology 2000; 22: 108–124
- Mosqueda-Garcia R, Furlan R, Fernandez-Violante R, Desai T, Snell M, Jarai Z, Ananthram V, Robertson RM, Robertson D. Sympathetic and baroreceptor reflex function in neurally mediated syncope evoked by tilt. J Clin Invest 1997; 99: 2736–2744
- Nathan MA, Reis DJ. Chronic labile hypertension produced by lesions of the nucleus tractus solitarii in the cat. Circ Res 1977; 40: 72–81
- Niimi Y, Matsukawa T, Sugiyama Y, Shamsuzzaman AS, Ito H, Sobue G, Mano T. Effect of heat stress on muscle sympathetic nerve activity in humans. J Auton Nerv Syst 1997; 63: 61–67
- Nisenbaum LK, Zigmond MJ, Sved AF, Abercrombie ED. Prior exposure to chronic stress results in enhanced synthesis and release of hippocampal norepinephrine in response to a novel stressor. J Neurosci 1991; 11: 1478–1484
- Pacak K, Palkovits M, Yadid G, Kvetnansky R, Kopin IJ, Goldstein DS. Heterogeneous neurochemical responses to different stressors: A test of Selye's doctrine of nonspecificity. Am J Physiol 1998; 275: R1247–R1255
- Patel JN, Coppack SW, Goldstein DS, Miles JM, Eisenhofer G. Norepinephrine spillover from human adipose tissue before and after a 72-hour fast. J Clin Endocrinol Metab 2002; 87: 3373–3377
- Quillen EW, Jr, Cowley AW, Jr. Influence of volume changes on osmolality-vasopressin relationships in conscious dogs. Am J Physiol 1983; 244: H73–H79
- Radikova Z, Penesova A, Koska J, Kvetnansky R, Jezova D, Huckova M, Vigas M, Macho L. Does orthostatic stress influence the neuroendocrine response to subsequent hypoglycemia in humans?. Ann N Y Acad Sci 2004; 1018: 576–581
- Robertson DA, Johnson GA, Robertson RM, Nies AS, Shand DG, Oates JA. Comparative assessment of stimuli that release neuronal and adrenomedullary catecholamines in man. Circulation 1979; 59: 637–643
- Rundqvist B, Elam M, Bergmann-Sverrisdottir Y, Eisenhofer G, Friberg P. Increased cardiac adrenergic drive precedes generalized sympathetic activation in human heart failure. Circulation 1997; 95: 169–175
- Rusnak M, Kvetnansky R, Jelokova J, Palkovits M. Effect of novel stressors on gene expression of tyrosine hydroxylase and monoamine transporters in brainstem noradrenergic neurons of long-term repeatedly immobilized rats. Brain Res 2001; 899: 20–35
- Schachter S, Singer J. Cognitive, social, and physiological determinants of emotional state. Psychol Rev 1962; 69: 379–399
- Schulkin J, Gold PW, McEwen BS. Induction of corticotropin-releasing hormone gene expression by glucocorticoids: Implication for understanding the states of fear and anxiety and allostatic load. Psychoneuroendocrinology 1998; 23: 219–243
- Selye H. The stress of life. McGraw-Hill, New York 1956
- Selye H. Stress without distress. New American Library, New York 1974
- Shah SD, Tse TF, Clutter WE, Cryer PE. The human sympathochromaffin system. Am J Physiol 1984; 247: E380–E384
- Sharabi Y, Dendi R, Holmes C, Goldstein DS. Baroreflex failure as a late sequela of neck irradiation. Hypertension 2003; 42: 110–116
- Sofuoglu M, Nelson D, Babb DA, Hatsukami DK. Intravenous cocaine increases plasma epinephrine and norepinephrine in humans. Pharmacol Biochem Behav 2001; 68: 455–459
- Staten MA, Matthews DE, Cryer PE, Bier DM. Physiological increments in epinephrine stimulate metabolic rate in humans. Am J Physiol 1987; 253: E322–E330
- Sterling P, Eyer J. Allostasis: A new paradigm to explain arousal pathology. Handbook of Life Stress, Cognition, and Health, J Fisher, J Reason. Johns Wiley & Sons Inc., New York 1988; 629–649
- Udelsman R, Goldstein DS, Loriaux DL, Chrousos GP. Catecholamine-glucocorticoid interactions during surgical stress. J Surg Res 1987; 43: 539–545
- von Euler US. A specific sympathomimetic ergone in adrenergic nerve fibres (sympathin) and its relations to adrenaline and nor-adrenaline. Acta Physiol Scand 1946; 12: 73–96
- Vogel ER, Sandroni P, Low PA. Blood pressure recovery from Valsalva maneuver in patients with autonomic failure. Neurology 2005; 65: 1533–1537
- Wilkinson DJC, Thompson JM, Lambert GW, Jennings GL, Schwarz RG, Jeffreys D, Turner AG, Esler MD. Sympathetic activity in patients with panic disorder at rest, under laboratory mental stress, and during panic attacks. Arch Gen Psychiatry 1998; 55: 511–520
- Young JB, Landsberg L. Sympathoadrenal activity in fasting pregnant rats: Dissociation of adrenal medullary and sympathetic nervous system responses. J Clin Invest 1979; 64: 109–116
- Young JB, Rosa RM, Landsberg L. Dissociation of sympathetic nervous system and adrenal medullary responses. Am J Physiol 1984; 247: E35–E40