Abstract
The presence of behavioral control over a stressor can blunt many of the effects of the stressor. We have recently reported that uncontrollable stress (inescapable electric tailshock, IS) reduces later social exploration of a juvenile whereas controllable stress (escapable shock, ES) does not. Activation of the ventral medial prefrontal cortex (vmPFC) is crucial to blunting the effects of IS on later escape behavior (learned helplessness). The goal of the current study was to test the role of the vmPFC in modulating the effects of stressor controllability on anxiety in the social exploration test. Thus, adult male rats were implanted with cannula guides for drug microinjection into the vmPFC. In Experiment 1, temporary inactivation of the vmPFC with the GABAA agonist muscimol before exposure to ES prevented the protective effects of stress control, leading to reduced social exploration. In Experiment 2, excitation of the vmPFC prior to IS with the GABA-activated Cl( − ) channel antagonist picrotoxin mimicked the stress resistance produced by control and prevented IS-induced reduction in social exploration. These results are consistent with prior work and identify the vmPFC as a critical component of the neural circuitry mediating the effects of stressor control on later behaviors. The relationship between the vmPFC, dorsal raphé nucleus, and other structures mediating stress-induced anxiety are discussed.
Introduction
The degree of behavioral control that an individual has over a stressor often determines the consequence of that stressor (Maier and Watkins Citation2005) and may play a critical role in the development of pathological behaviors after a traumatic event (Foa et al. Citation1992). Indeed, numerous behavioral outcomes have been reported to follow uncontrollable stress but not an equal exposure where the stressor is controllable (Shors Citation2004; Maier and Watkins Citation2005). For example, exposure to a series of inescapable electrical tailshocks (IS) produces behavioral changes that include exaggerated fear conditioning, poor shuttle escape learning, and reduced social interaction, while exposure to tailshock that is escapable (ES) does not (Short and Maier Citation1993; Amat et al. Citation2005; Christianson et al. Citation2008).
Our laboratory has determined that one mechanism by which behavioral control operates is to prevent stress-induced activation and sensitization of the serotonergic dorsal raphé nucleus (DRN), the mechanism by which uncontrollable stress leads to the behavioral consequences cited above. Using in vivo microdialysis we have found that extracellular serotonin (5-HT) concentrations in the DRN nearly triple at the beginning of the shock session, whether the tailshocks are uncontrollable or controllable. However, when behavioral control is present 5-HT levels quickly return to baseline as the subject learns the escape response, even though tailshocks continue (Maswood et al. Citation1998; Amat et al. Citation2005). Extracellular 5-HT within the DRN reflects DRN 5-HT activation because activated DRN 5-HT neurons release 5-HT within the DRN from axon collaterals, as well as at axon terminals in projection regions (Matos et al. Citation1996). Extracellular DRN 5-HT may also reflect the activity of serotonergic inputs to the DRN, such as the median raphé (MR) but we have demonstrated that MR is activated equally by ES and IS (Takase et al. Citation2004). Interestingly, both IS and ES activate brain regions that send excitatory input to the DRN (Amat et al. Citation2001) but as noted, only IS results in a sustained activation of 5-HT neurons. Thus, the differential net impact of IS and ES would seem to require ES-induced inhibitory inputs to the DRN.
In the rat, the infralimbic/prelimbic region of the ventromedial prefrontal cortex (vmPFC) sends glutamatergic efferents to the DRN (Gabbott et al. Citation2005) and this vmPFC region has been theorized to contribute to resilience after stress (Jordan et al. Citation1994). Efferent axons from the vmPFC to the DRN synapse preferentially on GABAergic interneurons that inhibit 5-HT cells (Jankowski and Sesack Citation2004). Thus, activation of descending vmPFC pyramidal neurons enhances inhibition within the DRN and decreases 5-HT neuronal activity (Celada et al. Citation2001). The foregoing suggested that perhaps the vmPFC detects behavioral control when it is present, leading to output to the DRN and inhibition of DRN 5-HT neurons. This hypothesis has been supported in a number of ways. First, inhibition of the vmPFC via microinjection of the GABAA agonist muscimol prior to stressor exposure had no effect on the DRN 5-HT neuronal response to IS, but led ES to produce DRN activation, later escape failure, and exaggerated post-shock freezing similar to IS, despite successful shock escape during the initial ES (Amat et al. Citation2005). Thus, preventing vmPFC activation during ES made the subject appear as if it had experienced IS. Conversely, pharmacological activation of the vmPFC by microinjection of the GABA-activated Cl( − ) channel antagonist picrotoxin during IS reduced the stimulation of DRN 5-HT and prevented escape deficits after IS (Amat et al. Citation2008). That is, pharmacological activation of the vmPFC appeared to give rats the “illusion of control”. In sum, the evidence suggests that stress exposure per se activates the DRN, which may be a final common pathway whose projections lead to anxiety-like behaviors. However, when stress is controllable the vmPFC inhibits the DRN and prevents many of the sequelae of uncontrollable stress.
The data implicating the critical role of the vmPFC, however, all depend on post-shock freezing and shuttlebox shock escape failure as the behavioral endpoints. Thus, the generality of the vmPFC effects described above is unknown. We have recently reported that social exploration of a juvenile conspecific, a putative measure of anxiety (Christianson et al. Citation2008), is reduced by IS, but not ES. Furthermore, this effect of IS was dependent on activation of DRN 5-HT (Christianson et al. Citation2008). The goal of the present study was to determine whether the stress-protective effect of behavioral control on juvenile social exploration is the result of vmPFC activation. In Experiment 1, muscimol was microinjected into the vmPFC before ES or home-cage control (HC) treatment to determine whether activation of this region by ES is necessary for the protective effects of behavioral control on stressor-induced reductions in social exploration. Inactivation of vmPFC by muscimol should release the DRN from inhibition by stressor control and generate a behavior profile similar to that produced by IS. In Experiment 2, picrotoxin was microinjected into the vmPFC before IS or HC treatment to determine whether activation of the region during stress is sufficient to prevent the usual reduction in social exploration produced by IS.
Materials and methods
Subjects
Adult (60–70 days old and weighing 275–350 g at the time of testing) and juvenile (28–32 days old and weighing 90–100 g at the time of testing) male Sprague-Dawley (Harlan, Indianapolis, IN, USA) rats were used. Rats were housed in plastic tub cages, 4 rats/cage with free access to food and water. The vivarium maintained a 12 h light/dark cycle with lights on at 7:00 am. All behavioral procedures were conducted in the first 5 h of the light cycle. The experimental protocols were reviewed and approved by the University of Colorado Institutional Animal Care and Use Committee and were in accordance with NIH guidelines.
Escapable and inescapable tailshock procedures
Escapable tail shocks were administered in 14 × 11 × 17 cm acrylic wheel turn boxes enclosed in sound-attenuating chambers. Electric shock was delivered through copper electrodes augmented by electrode paste attached 2 and 4 cm from the base of the tail by a Precision Regulated Animal Shocker (Coulbourn Instruments, Allentown, PA, USA). One hundred tailshocks were presented on a variable interval 60-s schedule (VI-60). For ES subjects, turning a wheel at the front of the chamber terminated each tailshock according to a protocol previously described (Amat et al. Citation2005). Shock intensity was 1.0 mA for the first 33 trials, 1.3 mA for the following 33 trials, and 1.6 mA for the remaining 34 trials. These parameters were used to maintain escape behavior in the ES subjects. For IS, 100, 5 s 1.6 mA inescapable tailshocks were administered in clear plastic restraining tubes on a VI-60-s schedule. This method has been reported previously, approximates the amount of shock received in yoked-IS, and elicits behavior that is indiscriminable from 100 trials of yoked-IS treatment on social exploration (Christianson et al. Citation2008). Naive, HC rats remained in their cages. A restraint stress-only control group was not included as this treatment was indescriminable from HC controls in a prior experiment (Christianson et al. Citation2008).
Juvenile social exploration test
Each experimental adult rat was allocated a separate transparent plastic tubcage with shaved wood bedding and a wire lid located in a brightly lit testing room; food and water were not available in the testing cages. Twenty-four hours before stress exposure rats were removed from the vivarium and placed into the single cage. After 60 min a juvenile stimulus rat was added to the cage. Investigative behaviors, including sniffing, pinning, and allogrooming, initiated by the adult rat were timed by an observer who was blind to group membership. After 3 min, the juvenile was removed and the adult was returned to the homecage. Juvenile stimulus rats were used for multiple tests but were never exposed to the same adult more than once. This social exploration pretest was used to habituate the subjects to the procedure and to screen for rats with unusual baseline social exploration. In our experience, healthy naïve rats typically explore for approximately 80 s; thus, rats with pretrest social exploration times less than 50 s were excluded as outliers. Sickness, induced by lipopolysaccharide for example, reduces social exploration (Bluthe et al. Citation1992). Twenty-four hours after stress, social exploration tests were conducted exactly as on the day before stress.
Cannula placement and microinjection procedures
All surgery was conducted under inhalational anesthesia (2–3% isoflurane in oxygen). Each rat was implanted with a single dual guide cannula (26 g, 1 mm center-to-center distance; Plastics One, Roanoke, VA, USA) so that bilateral injector tips would reach the border between the infralimbic and prelimbic cortices (AP +2.9, LM ± 0.5, DV − 2.9). Coordinates were taken from bregma and dura according to Paxinos and Watson (Citation1998). Cannulae were fixed to the skull with three screws and dental cement. A stylet was placed in the cannula extending 1 mm below the tip of the guide. After surgery each rat received prophylactic antibiotic, s.c. 0.25 ml Twin-Pen (AgriLabs, St. Joseph, MO, USA) per kg body weight. Intracerebral microinjections were made by gently restraining the rat in a towel and replacing the stylet with a microinjector that extended 1 mm beyond the cannula tip (33 g; Plastics One) joined to a cannula connector. In Experiment 1, muscimol, 500 ng/side, in 0.5 μl 0.9% saline or saline alone was injected at a rate of 1 μl/min through PE-50 tubing by a 25 μl Hamilton syringe and a Kopf micromanipulator. Injectors remained in place for 2 min to permit diffusion. In Experiment 2, 0.5 μl of picrotoxin, 100 ng/side, in 0.9% saline or saline alone was injected in the same way. Muscimol and picrotoxin doses were determined in pilot studies. At the end of each experiment rats were overdosed with sodium pentobarbital (60 mg/kg i.p.) and brains removed and processed for cresyl violet verification of cannula placement using standard procedures.
Experimental procedures
Experiment 1
After at least 7 days of acclimation to the vivarium, bilateral guide cannulae were implanted. After 7–10 recovery days, all rats were given a social exploration pretest and then randomly assigned to one of four groups in a (Stress: ES or HC) by (Drug: Muscimol or Saline) between subjects factorial design (n = 10/group). IS rats were not included because the question posed was whether inactivation of the vmPFC during ES would now lead ES to reduce social exploration. IS subjects were not needed to answer this question, while intra-vmPFC muscimol had no effect on IS subjects in prior studies (Amat et al. Citation2005, Citation2006; Baratta et al. Citation2007). On the following day, the rats received microinjection of either muscimol or saline 60 min before ES. Rats in the HC group were returned to the vivarium after injection. Twenty-four hours after the onset of stress all rats were given the social exploration test.
Experiment 2
As in Experiment 1, rats were implanted with cannulae, allowed 7–10 days to recover and given a social exploration pretest. Rats were randomly assigned to one of four groups in a (Stress: IS or HC) by (Drug: Picrotoxin or Saline) between-subjects factorial design (n = 10/group). On the following day, rats received intracrebral microinjection of either picrotoxin or saline 60 min before IS exposure. Rats in the HC group were returned to the vivarium after injection. Twenty-four hours after the onset of stress all rats were given the social exploration test. ES subjects were not included because the question posed was whether intra-vmPFC picrotoxin would block the reduction in social exploration produced by IS. Picrotoxin had no effect in ES subjects in prior studies (Amat et al. Citation2008).
Results
Experiment 1
Exclusion of subjects with social exploration times less than 50 s yielded the following group sizes: ES-Saline, n = 8, ES-Muscimol, n = 10, HC-Saline, n = 8, HC-Muscimol, n = 9. Pretreatment with muscimol did not interfere with acquisition of the wheel-turn escape response and all ES rats quickly reached the maximum escape requirement (four full turns) with escape latencies under 5 s for the majority of trials; these data are comparable to those previously reported (Amat et al. Citation2005). Location of the cannula tips and mean social exploration time are shown in and , respectively. In intracerebral saline-treated rats, ES had no effect on social exploration relative to unstressed HC controls. However, inhibition of the vmPFC by muscimol led to a significant reduction in social exploration after ES but had no effect on the HC group. A two-way ANOVA identified a main effect of Stress, F(1, 30) = 5.754, p = 0.023, and a significant Stress by Drug interaction, F(1, 30) = 14.65, p < 0.001. The main effect of Drug did not reach significance, p = 0.14. Fisher's protected least significant difference (PLSD) post-hoc comparisons indicated that mean social exploration in the ES-Muscimol group was significantly less than ES-Saline and both HC groups, ps < 0.01. No other comparisons reached significance.
Figure 1 Location of cannula injector tips in the ventromedial prefrontal cortex. Anatomical illustrations adapted from The Rat Brain in Stereotaxic Coordinates (Paxinos and Watson Citation1998).
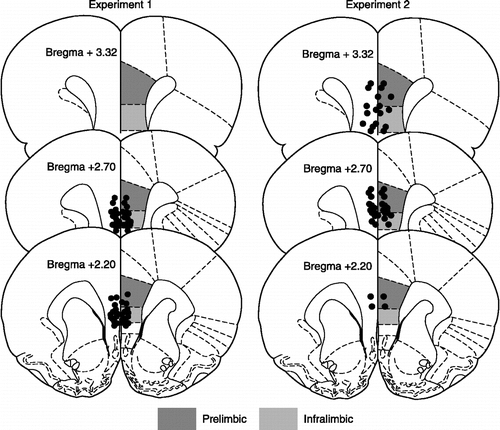
Figure 2 Escapable stress and social exploration. Mean ( ± SEM) time spent in social exploration of a juvenile during a 3-min test. Escapable stress had no effect on social exploration (ES-Saline group) but inactivation with muscimol microinjection of vmPFC during stress interfered and significantly reduced social exploration in the ES-Muscimol group. HC: home cage controls. *ES-Muscimol group significantly less than all other groups, ps < 0.01. n = 8–10 rats per group.
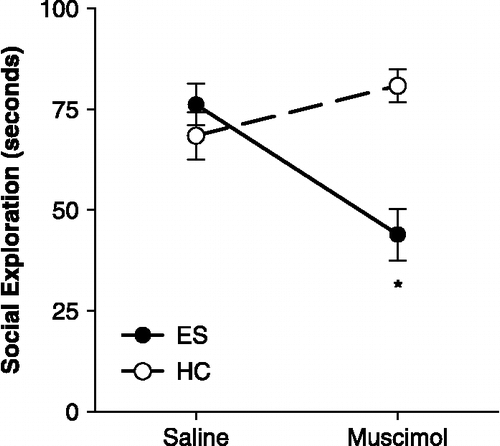
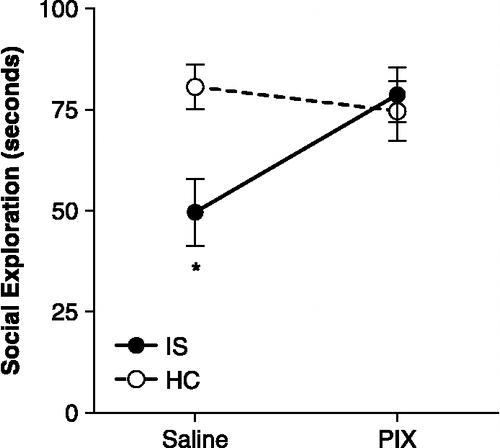
Figure 3 Inescapable stress and social exploration. Mean ( ± SEM) time spent in social exploration of a juvenile during a 3-min test. Inescapable stress (IS-Saline group) reduced social exploration compared to the HC. However, activation of the vmPFC at the time of stress with picrotoxin microinjection (IS-PIX group) completely prevented the effect of stress and social exploration time was similar to controls (HC). *IS-Saline group significantly less than HC-Saline and IS-Picrotoxin groups, ps < 0.01. n = 10 rats per group.
Experiment 2
No rats had social exploration times less than 50 s in the social exploration pretest. Location of cannula tips and mean social exploration time are shown in and , respectively. IS reduced social exploration but activation of vmPFC before IS completely prevented this effect. A two-way ANOVA identified a marginally significant main-effect of Stress, F(1, 36) = 3.634, p = 0.065 and a significant Stress by Drug interaction, F(1, 36) = 6.16, p = 0.018. The main effect of Drug did not reach significance, p = 0.11. Fisher's PLSD post-hoc comparisons revealed that mean social exploration time was significantly reduced in the IS-Saline group compared to HC-Saline and IS-Picrotoxin groups, ps < 0.01. No other comparisons reached significance.
Discussion
The data from the present experiments further implicate the vmPFC in the neural circuitry that mediates the resistance to the behavioral effects of a stressor that occurs when behavioral control over the stressor is present. Rats were exposed to escapable or inescapable tailshocks, with concomitant pharmacological inactivation or activation of the vmPFC during the stressor, respectively. Prior experiments have suggested that the control over stress activates vmPFC regions (Amat et al. Citation2005, Citation2006; Baratta et al. Citation2007). Thus, rats with an active (ES-Saline and IS-Picrotoxin) vmPFC during the stressor showed no effect of the stressor session in the social exploration test 1 day later. Based on previous observations in our laboratory, with respect to social exploration, inactivation of the vmPFC during IS (IS-Muscimol) has no effect on IS subjects (Amat et al. Citation2005); likewise, activation of the mPFC (ES-picrotoxin) has no effect on ES subjects (Amat et al. Citation2008). Therefore, these experimental groups were unnecessary in the present study. The present results complement prior work that utilized post-shock freezing and shuttle escape measures and illuminate the way by which control over stress relates to anxiety-like endpoints.
As noted, IS results in prolonged activation of the DRN, leading to downregulation of somatodendritic 5-HT1A autoreceptor mRNA (Greenwood et al. Citation2003), resulting in sensitization of 5-HT neurons to subsequent aversive stimuli (Amat et al. Citation1998). Inhibition of the DRN during IS prevents the subsequent reduction in social exploration (Christianson et al. Citation2008). Taken with the anatomical relationship between the vmPFC and the DRN described above, the present data further support the argument that control over stress regulates the activity of the DRN by a circuit involving the vmPFC.
The vmPFC receives a convergence of thalamic and subcortical inputs and is considered to be an executive center suited to regulate a range of neural processes (Dalley et al. Citation2004). It is likely, therefore, that when the vmPFC receives sensory information that contains a contingent relationship between motor behaviors and shock termination (i.e., the escape response) then it is able to inhibit stress responsive nuclei such as the DRN. In contrast, when there is no apparent relationship between behavior and shock termination the vmPFC does not send inputs to stress-responsive regions, and so they remain unregulated. If this is the case, then the critical variable determining the outcomes of stressor controllability experiments is not whether the subject detects the non-contingency of behavior and shock as was originally proposed (Maier and Seligman Citation1976), but rather that behavioral control over stress activates vmPFC output.
These experiments implicate the vmPFC in the regulation of anxiety-like behavior. Social exploration is a widely used measure of anxiety-like behavior (File and Seth Citation2003) and DRN activity is correlated with reduced exploration (Higgins et al. Citation1992; File et al. Citation1996; Overstreet et al. Citation2006). The idea here is that activity within the DRN modulates anxiety behavior via projections to limbic regions such as the basolateral amygdala (Lowry et al. Citation2005). Therefore, social exploration may be reduced in IS-treated rats because the DRN is sensitized and is hyper-responsive in the mildly-anxiogenic context of a social encounter. However, the vmPFC might instead, or in addition, directly modulate amygdala activity (Quirk et al. Citation2003). The role of the DRN and vmPFC in anxiety are reinforced by two human studies. Lanzenberger et al. (Citation2007) reported a reduction in DRN 5-HT1A receptors in social anxiety disorder patients and Monk et al. (Citation2008) found an inverse relationship between prefrontal cortex and amygdala activity when patients with generalized anxiety were presented with anxiogenic stimuli. Because the present data do not directly address the relationship between the vmPFC and the amygdala, the possibility of an important action of the vmPFC by this route remains open. Nonetheless, the role of the vmPFC in anxiety-like behavior may be simply to prevent sensitization of the DRN.
Acknowledgements
Funding for these experiments was provided by National Institutes of Health Grant MH050479 to S.F.M. and MH082453 to J.P.C.
Declaration of interest: The authors report no conflicts of interest. The authors alone are responsible for the content and writing of the paper.
References
- Amat J, Matus-Amat P, Watkins LR, Maier SF. Escapable and inescapable stress differentially alter extracellular levels of 5-HT in the basolateral amygdala of the rat. Brain Res 1998; 812: 113–120
- Amat J, Sparks PD, Matus-Amat P, Griggs J, Watkins LR, Maier SF. The role of the habenular complex in the elevation of dorsal raphe nucleus serotonin and the changes in the behavioral responses produced by uncontrollable stress. Brain Res 2001; 917: 118–126
- Amat J, Baratta MV, Paul E, Bland ST, Watkins LR, Maier SF. Medial prefrontal cortex determines how stressor controllability affects behavior and dorsal raphe nucleus. Nat Neurosci 2005; 8: 365–371
- Amat J, Paul E, Zarza C, Watkins LR, Maier SF. Previous experience with behavioral control over stress blocks the behavioral and dorsal raphe nucleus activating effects of later uncontrollable stress: Role of the ventral medial prefrontal cortex. J Neurosci 2006; 26: 13264–13272
- Amat J, Paul E, Watkins LR, Maier SF. Activation of the ventral medial prefrontal cortex during an uncontrollable stressor reproduces both the immediate and long-term protective effects of behavioral control. Neuroscience 2008; 154: 1178–1186
- Baratta MV, Christianson JP, Gomez DM, Zarza CM, Amat J, Masini CV, Watkins LR, Maier SF. Controllable versus uncontrollable stressors bi-directionally modulate conditioned but not innate fear. Neuroscience 2007; 146: 1495–1503
- Bluthe RM, Dantzer R, Kelley KW. Effects of interleukin-1 receptor antagonist on the behavioral effects of lipopolysaccharide in rat. Brain Res 1992; 573: 318–320
- Celada P, Puig MV, Casanovas JM, Guillazo G, Artigas F. Control of dorsal raphe serotonergic neurons by the medial prefrontal cortex: Involvement of serotonin-1A, GABA(A), and glutamate receptors. J Neurosci 2001; 21: 9917–9929
- Christianson JP, Paul ED, Irani M, Thompson BM, Kubala KH, Yirmiya R, Watkins LR, Maier SF. The role of prior stressor controllability and the dorsal raphe nucleus in sucrose preference and social exploration. Behav Brain Res 2008; 193: 87–93
- Dalley JW, Cardinal RN, Robbins TW. Prefrontal executive and cognitive functions in rodents: Neural and neurochemical substrates. Neurosci Biobehav Rev 2004; 28: 771–784
- File SE, Seth P. A review of 25 years of the social interaction test. Eur J Pharmacol 2003; 463: 35–53
- File SE, Gonzalez LE, Andrews N. Comparative study of pre- and postsynaptic 5-HT1A receptor modulation of anxiety in two ethological animal tests. J Neurosci 1996; 16: 4810–4815
- Foa EB, Zinbarg R, Rothbaum BO. Uncontrollability and unpredictability in post-traumatic stress disorder: An animal model. Psychol Bull 1992; 112: 218–238
- Gabbott PL, Warner TA, Jays PR, Salway P, Busby SJ. Prefrontal cortex in the rat: Projections to subcortical autonomic, motor, and limbic centers. J Comp Neurol 2005; 492: 145–177
- Greenwood BN, Foley TE, Day HE, Campisi J, Hammack SH, Campeau S, Maier SF, Fleshner M. Freewheel running prevents learned helplessness/behavioral depression: Role of dorsal raphe serotonergic neurons. J Neurosci 2003; 23: 2889–2898
- Higgins GA, Jones BJ, Oakley NR. Effect of 5-HT1A receptor agonists in two models of anxiety after dorsal raphe injection. Psychopharmacology (Berl) 1992; 106: 261–267
- Jankowski MP, Sesack SR. Prefrontal cortical projections to the rat dorsal raphe nucleus: Ultrastructural features and associations with serotonin and gamma-aminobutyric acid neurons. J Comp Neurol 2004; 468: 518–529
- Jordan S, Kramer GL, Zukas PK, Moeller M, Petty F. In vivo biogenic amine efflux in medial prefrontal cortex with imipramine, fluoxetine, and fluvoxamine. Synapse 1994; 18: 294–297
- Lanzenberger RR, Mitterhauser M, Spindelegger C, Wadsak W, Klein N, Mien LK, Holik A, Attarbaschi T, Mossaheb N, Sacher J, Geiss-Granadia T, Kletter K, Kasper S, Tauscher J. Reduced serotonin-1A receptor binding in social anxiety disorder. Biol Psychiatry 2007; 61: 1081–1089
- Lowry CA, Johnson PL, Hay-Schmidt A, Mikkelsen J, Shekhar A. Modulation of anxiety circuits by serotonergic systems. Stress 2005; 8: 233–246
- Maier SF, Seligman ME. Learned helplessness: Theory and evidence. J Exp Psychol Gen 1976; 105: 3–46
- Maier SF, Watkins LR. Stressor controllability and learned helplessness: The roles of the dorsal raphe nucleus, serotonin, and corticotropin-releasing factor. Neurosci Biobehav Rev 2005; 29: 829–841
- Maswood S, Barter JE, Watkins LR, Maier SF. Exposure to inescapable but not escapable shock increases extracellular levels of 5-HT in the dorsal raphe nucleus of the rat. Brain Res 1998; 783: 115–120
- Matos FF, Urban C, Yocca FD. Serotonin (5-HT) release in the dorsal raphe and ventral hippocampus: Raphe control of somatodendritic and terminal 5-HT release. J Neural Transm 1996; 103: 173–190
- Monk CS, Telzer EH, Mogg K, Bradley BP, Mai X, Louro HM, Chen G, McClure-Tone EB, Ernst M, Pine DS. Amygdala and ventrolateral prefrontal cortex activation to masked angry faces in children and adolescents with generalized anxiety disorder. Arch Gen Psychiatry 2008; 65: 568–576
- Overstreet DH, Knapp DJ, Angel RA, Navarro M, Breese GR. Reduction in repeated ethanol-withdrawal-induced anxiety-like behavior by site-selective injections of 5-HT1A and 5-HT2C ligands. Psychopharmacology (Berl) 2006; 187: 1–12
- Paxinos G, Watson C. The rat brain in stereotaxic coordinates4th ed. Academic Press, New York 1998
- Quirk GJ, Likhtik E, Pelletier JG, Pare D. Stimulation of medial prefrontal cortex decreases the responsiveness of central amygdala output neurons. J Neurosci 2003; 23: 8800–8807
- Shors TJ. Learning during stressful times. Learn Mem 2004; 11: 137–144
- Short KR, Maier SF. Stressor controllability, social interaction, and benzodiazepine systems. Pharmacol Biochem Behav 1993; 45: 827–835
- Takase LF, Nogueira MI, Baratta M, Bland ST, Watkins LR, Maier SF, Fornal CA, Jacobs BL. Inescapable shock activates serotonergic neurons in all raphe nuclei of rat. Behav Brain Res 2004; 153: 233–239