
1. Introduction
For many years, bipedal locomotion has been considered to be one of the main characteristics of the evolution of the Homo genus. However, this locomotor mode is not human specific. In fact, it is certainly occasionally performed in all non-human primates. Nevertheless, in humans, the erected bipedal gait is unique and is characterized by a succession of phases of single and double support using 3 rockers. The foot is an important interface with the ground that allows this unique bipedal style. In Homo sapiens, the (bipedal) anatomy is thus very specialized and different from that of other hominids. For instance, the bonobo, Pan paniscus, is our closest relative and walk bipedally occasionally, yet it does not have an developed Achilles tendon as in Homo sapiens. The baboon, Papio anubis, is a non-hominoid primate capable of occasional bipedalism as well, yet, contrary to bonobos, it has an Achilles tendon which is anatomically similar to the one of Homo sapiens. Comparative structural anatomy is used to infer differences between species.Foot bones are commonly used, but the structural analysis does not always reflect the functional abilities. For this purpose, comparative anatomy can focus on muscles, Biomechanical modelling and in particular the rigid multi-body method makes it possible to create prototypes of the foot that can be activated in direct dynamics. Anatomical transfer algorithms enable to simulate, using bone information only, the feet of different individuals by transferring the muscles and ligaments from an Atlas model.
The objective of this preliminary study is to perform an anatomical transfer from Homo sapiens to Pan paniscus and Papio anubis in order to run simulations by rigid multi-body technics. We hypothesize that the non-functional parts of the biomechanical model will help us to determine what structural anatomical knowledge we need to acquire in order to make species specific models.
2. Methods
2.1. Data acquisition, bone segmentation and meshing
The anatomical data acquisition of primates is difficult on living animals. It can be performed in an opportunistic manner upon the death of an individual. Thus, a full-body CT scan of a 8 years old female Papio anubis was performed with a resolution of 2048 x 1024 on 1681 0.6 mm slices. A full-body CT scan of a young Pan paniscus male with a resolution of 511 x 374 on 1981 0.4 mm slices was shared by the American Museum of Natural History, New York (Doug Boyer provided access to these dataNSF BCS 1552848 and NSF DBI 1701714. The files were downloaded from www.MorphoSource.org, Duke University). A manual segmentation was performed using the Amira software (Thermofisher) then the surface mesh was optimized by the Modeleditor software (Texisense) to obtain a mesh compatible with a rigid multibody simulation.
2.2. Anatomical transfer
The implementation of the atlas to patient registration procedure is divided into three steps increasingly introducing distortion in the data: (1) a rigid registration that roughly positions the patient data set with respect to the atlas model, (2) an affine deformation that compensates for global scale discrepancies, followed by 3) an elastic registration that accurately fits the bony contours and the skin surface. Once all three deformation functions are combined, the resulting deformation is applied to the atlas dataset to transfer the atlas information (muscles, ligament insertions, plantar fascia) into the patient’s referential. The procedure producing the musculoskeletal component of the patient model is automatic and takes about two hours.
2.3. Simulations
For each subject, the following data are available: bones from the segmentation belonging to the individual, ligaments, muscles and fascias from the Homo sapiens atlas. The models constructed are thus hybrid models. The simulations are performed in the 3 D simulation platform ArtiSynth (www.artisynth.org) with the Paradiso solver and a semi-implicit method: Constrained Backward Euler. We used the surface objects of each bone and assigned them the density of 1,850 kg/m3, Souzanchi et al. (2012). 210 Homo sapiens foot ligaments were connected to primate bone models in the shape of 1 D cables. These cables constrain the movements of 3 D objects by limiting their distance in tension, in linear, with the stiffness of 395 kPa in elongation and 0 kPa in compression. This value is the equivalent in elongation coefficient of the parameters used by Gefen et al. (2003) for ligaments with a Young’s modulus of 200 MPa and a fish coefficient of 0.4. These models do not have an ideal joint, so only the ligaments and contacts created between the bones affect joint movement. The interpenetration tolerance of the surfaces is 0.1 mm and the coefficient of friction is null. We also modelled the plantar fascia. From a muscular aspect, we used the Hill model and positioned 15 intrinsic and extrinsic muscles of the foot. Their trajectories were located on the MRI of Homo sapiens before transfer. Each muscle can be activated independently of the other muscles. The objective of the simulation was to perform dorsal and plantar talo-crural flexion and simulate the successive contraction of the muscles of the anterior and posterior muscle lodges. The anatomical integrity is then analysed and joints with abnormal positions are identified.
3. Results and discussion
The anatomical transfer is only functional if the target model and the atlas have the same number of bones. Thus, it was necessary to remove, from the Papio anubis model, the sesamoid bones present on the metatarsophalangeal joints of the 2,3,4 and 5th rays as well as an accessory cuboid bone. For Pan paniscus, an accessory navicular bone was removed. The anatomical transfer did not allow stable resting models, instability of the metatarsophalangeal and metatarsocunean joints for Papio anubis and the metatarsophalangeal and metatarsocuboidian instability for Pan paniscus. Simulationsr performed without toes and metatarsals have good joint stability for both specimens. A set of polyarticular ligament structure, fascias and ligaments as well as very specific uniarticular ligament structures, particularly on metatarsal-phalangeal ligaments enable good mid and forefoot stability on the validated Homo sapiens model. The muscles of these species are well described in the literature, but not the joints or the union means of polyarticular cohesion such as aponeuroses. In addition, the muscles have different paths allowing the opposability of the first ray.
4. Conclusions
Although anatomical transfer algorithms are effective, simulations have not yet yielded fully functional models. Indeed, some joints are unstable when they integrate the anatomy of Homo sapiens. These preliminary results allow to understand the potential limitations of a functional comparative anatomy with anatomical transfer. Our next work will therefore focus on anatomical studies specific to foot cohesion structures, particularly fascia and metatarsalphalangeal structures.
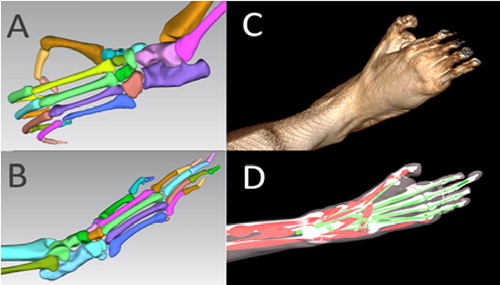
Figure 1. A: Pan paniscus bones, B: Papio anubis bones, C: Papio anubis volume rendering, D: Papio anubis multibody simulation.
Acknowledgements
CTScan of P. anubis have been collected within the frame of the CNRS-INEE International Research Network IRN-GDRI0870. Comparative analyses are conducted within the ANR-18-CE27-0010-01 HoBiS
References
- Aerts P, D’Août K, Thorpe S, Berillon G, Vereecke E. 2018. The gibbon’s Achilles tendon revisited: consequences for the evolution of the great apes? Proc Royal Soc B: Biol Sci. 285(1880):pii: 20180859.
- Bucki M, Luboz V, Perrier A, Champion E, Diot B, Vuillerme N, Payan Y. 2016. Clinical workflow for personalized foot pressure ulcer prevention. Med Eng Phys. 38(9):845–853.
- Fernández PJ, Holowka NB, Demes B, Jungers WL. 2016. Form and function of the human and chimpanzee forefoot: implications for early hominin bipedalism. Sci Rep. 6(1):30532.
- Jungers WL, Harcourt-Smith WEH, Wunderlich RE, Tocheri MW, Larson SG, Sutikna T, Due RA, Morwood MJ. 2009. The foot of Homo floresiensis. Nature. 459(7243):81–84.