
Abstract
Drawdown of Lake Mead from 1998 to 2011 reduced the lake's perimeter by more than 400 km while exposing more than 25,000 ha of formerly submerged land. To evaluate primary plant succession and soil properties in this new shoreline habitat, we sampled surfaces last submerged 3, 6, 9, or 13 years before sampling (including a never-submerged control) using 150, 100 m2 plots at 3 sites varying in topography and soil parent material. Consistent with previous research, abundance of the exotic saltcedar (Tamarix ramosissima) declined with increasing surface age. Conversely, cover of native species was greatest overall on older surfaces across sites. Early successional native perennial species colonized the 13-year-old surface. Unexpectedly, multivariate soil properties did not differ between never-submerged and formerly submerged surfaces. Based on the first 13 years of plant colonization along the Lake Mead shoreline and previous research on longer term Mojave Desert vegetation succession, managers can likely anticipate (1) continued development of an early colonizing native shrubland persistent for several decades, and (2) eventual colonization by species of the mature vegetation inhabiting never-submerged surfaces. Moreover, we observed colonization of formerly submerged land by a rare plant of special conservation designation, California bearpoppy (Arctomecon californica), suggesting that even after many years of submersion, rare plant habitat along the Lake Mead shoreline may be reclaimed. While Lake Mead's drawdown might be viewed negatively from a perspective of maintaining full-pool water storage, it has re-exposed a vast area of new terrestrial habitat increasingly colonized by native species as invasive species abundance declines through time.
Public benefits and challenges to the environmental water quality of Lake Mead in southwestern United States have been extensively discussed (Rosen et al. Citation2012); however, functional benefits and challenges to maintaining environmental quality of its shoreline habitat are not as well articulated. Changing water levels are a particularly important influence to shoreline habitat in lakes and reservoirs such as Lake Mead, characterized by large and variable fluctuations in water level (Hill et al. Citation1998, Ali Citation2006, Auble et al. Citation2007). Water levels of Lake Mead have fluctuated since its creation by the Hoover Dam in 1935, ranging from the lake's full pool elevation of 372 m in 1941 and 1983 to a low of 330 m in 2010 (Holdren and Turner Citation2010). Over the last 15 years, lake levels have declined to among their lowest for an extended period of time during the 77-year history of the lake (Holdren and Turner Citation2010). Decreasing water levels reduce the lake perimeter while increasing the amount of shoreline area relative to the unsubmerged amount when the lake is at full pool. By 2011, Lake Mead's drawdown had reduced the lake's perimeter by more than 400 km while exposing more than 25,000 ha of formerly submerged land, an area roughly equivalent to the size of the city of Sacramento, California ().
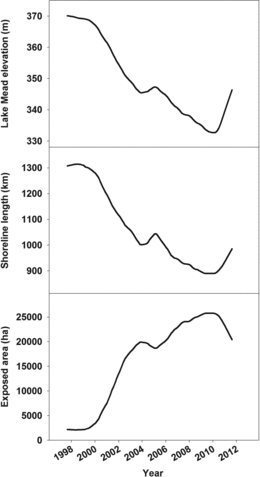
This vast area of newly exposed terrestrial habitat, together with continued fluctuations in lake levels and inherent uncertainty to projections of future lake levels, affords an interesting management dilemma. A positive aspect is that the newly exposed area offers significant potential for supporting a variety of terrestrial Mojave Desert communities, including those that contain rare plant (e.g., California bearpoppy [Arctomecon californica]) and animal (e.g., Phainopepla [Phainopepla nitens]) species that have special conservation protection. Additional lakeshore recreation opportunities for humans might also be available in this new habitat. A negative aspect is that bare land that was previously inundated might be colonized by invasive exotic plants such as saltcedar (Tamarix ramosissima) instead of native species. Another possibility is that plant colonization might slow altogether, leaving bare soil (Walker et al. Citation2006) that could result in soil erosion and fugitive dust that creates air quality problems, a human health hazard. Moreover, any attempted management activities (e.g., exotic species treatment, revegetating with native species, creation of recreation sites) near the new shoreline might be counteracted if lake levels again rise (Tallent et al. Citation2011).
The elevation of Lake Mead fluctuates seasonally, averaging about a 5 m difference between winter maximum and summer minimum depth. The vegetative community along the lakeshore varies in composition due to inherently fluctuating water levels. Riparian vegetation is historically thought to be sparse, even along much of the historical Colorado River corridor that Lake Mead submerged (Chandra et al. Citation2012), and much of the existing shoreline is sparsely vegetated. The consistent decline in water level since 1998 has prevented the establishment of a stable riparian community. Instead, xeric habitats remain in higher elevation areas that were formerly submerged and provide opportunities for colonization along the land–water interface by only the most weedy of riparian species that may quickly colonize and adapt to fluctuating water levels. Instead, xeric habitats remain in higher elevation areas that were formerly submerged and provide opportunities for colonization along the land-water interface by only the most weedy of riparian species that may quickly colonize and adapt to fluctuating water levels (Busch and Smith 1995).
The US National Park Service manages areas surrounding the lake as part of Lake Mead National Recreation Area. The National Park Service includes managing and conserving native species habitats as a key part of its mission and therefore has opportunities to manage the thousands of hectares of formerly submerged land. Lake Mead shoreline habitat is inherently an artificial habitat, yet it is a location where native species could be conserved. The shoreline area could also become a liability to protected upland habitat, however, such as if the shoreline creates source populations that spread exotic species.
Understanding vegetation colonization patterns and soil properties can inform evaluations of environmental quality and should be considered when making management decisions about recently disturbed areas. Plant colonization of the formerly submerged shoreline area would be primary succession along the primary–secondary successional continuum in terrestrial habitats (Walker and del Moral Citation2003). Studies of primary plant community succession have occurred on debris flows (new soil surfaces created by formation of alluvial fans through flooding-induced mass soil movement) in the Mojave Desert, where Lake Mead is located (Abella Citation2010). These studies reported that long time periods were required for colonization of these surfaces by species typical of nearby mature communities. In their study of debris flows along the Grand Canyon, for example, Bowers et al. (Citation1997) reported that the late-successional creosote bush (Larrea tridentata) was not present on any flows younger than 285 years; however, the studies suggested that within 10 years, early successional plants can colonize the surfaces and provide a total plant cover approaching half or more of the cover in mature communities (Abella Citation2010). It is unclear how closely successional patterns on these surfaces might match those in the shoreline environment or if properties of the formerly submerged soil are related to plant colonization. Submersion could have altered soils through processes such as sedimentation, erosion, incorporation or dissolution of elements, and production and decomposition of organic matter, all of which could affect the environment experienced by colonizing plants.
The receding level of Lake Mead might be viewed as disconcerting from a perspective of maintaining water storage and hydropower, but it has resulted in vast areas of newly exposed terrestrial habitat in regions that may have been submerged for more than 7 decades. This habitat and its potential values are poorly understood. The objective of this study was to determine plant community colonization and soil properties along a time-since-submersion, surface age gradient over the last 13 years during the present Lake Mead drawdown. We hypothesized that (1) species richness and native plant colonization increase with increasing surface age, (2) exotic species dominance declines on the oldest surfaces, and (3) soil properties differ along the surface age gradient, with higher fine fraction content, lower organic carbon (C) and total nitrogen (N), and elevated salt-related properties on submerged compared to nonsubmerged surfaces. Results can augment theory regarding vegetation change in arid lands while helping to understand quality of the new shoreline habitat to support potential shoreline management planning.
Methods
Site selection
We conducted this study at 3 sites (Boulder Beach, Overton, and Stewarts Point) representing a range of topography, soil parent material, and management considerations (). Topography across sites varies from gently sloping relatively flat and homogeneous shoreline sloping toward the lake to more rugged terrain. Boulder Beach soils are described as gravelly outwash, mixed alluvium piedmont fans, and are part of the Carrizo-Riverbend association classified as Typic Torriorthents and Typic Haplocalcids (Lato Citation2006).
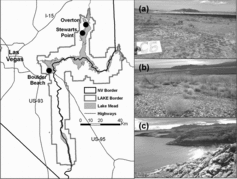
Management considerations at this site include visitor use of nearby beaches for water recreation, fire management, fugitive dust control, and weed encroachment. Stewarts Point contains soils associated with gypsum parent material, interspersed with fan alluvium derived from igneous, metamorphic, and sedimentary rock in the Cololag-badland association (Lato Citation2006). Never-submerged soils near Stewarts Point support rare plant species of management priority such as southwestern ringstem (Anulocaulis leiosolenus var. leiosolenus) and California bearpoppy. Because of the open, running Muddy River adjacent to the Overton sties, the water table remains high in this area, allowing marshy habitats and siltier soils immediately adjacent to the river. Soils at Overton also are associated with gypsum and are within the Drygyp-Bluegyp association classified as Typic Petrogypsids and Leptic Haplogypsids (Lato Citation2006). Management considerations here include trespass cattle, unauthorized off-road vehicle use, and weeds. Additionally, here, as at Stewarts Point, there is potential for creating habitat suitable for use by rare or threatened species.
Ten transects perpendicular to the Lake Mead shoreline were established along each of the 3 sites with five 10 × 10 m plots placed along each transect at elevations representing average lake levels for 1998 (the most recent time when the lake was near full pool; 370 m), 2002 (354 m), 2005 (347 m), and 2008 (338 m). Transects were assigned perpendicular to the lakeshore running through a randomly generated point, then plots were placed at each of the 4 target elevations along a transect. To be included in the study, plots must be undisturbed by roads, off-road vehicle traffic, or hiking trails, and have a slope gradient of <30%. Site elevations for each target year were defined as the mean elevation from January to December within that year, as recorded by the US Department of Interior, Bureau of Reclamation (http://www.usbr.gov/lc/region/g4000/hourly/mead-elv.html). Accurate elevations were determined by Light Detection and Ranging (LiDAR) elevation grids (captured August–September 2009, provided by National Park Service, Boulder City, NV). Additionally, transects were extended to include a control plot 50 m past the high water mark (sites that were never submerged) for each transect. Terminology in this paper will refer to sites last submerged in 2008 as the “youngest” plots, those last submerged in 1998 as the “oldest” plots, and plots that were never submerged as “control” plots. Plants examined in the 150 sample plots are referred to by scientific name hereafter to avoid confusion with similar common names.
Data collection
Plots were established along transects during December 2010. Plant and soils data were collected from February to May 2011, a period that coincides with the peak flowering period and biomass of most Mojave Desert species (particularly native annuals). Percent cover for all live species present in the 100 m2 plots was visually estimated to the nearest 1%. A category of <1% was created for species with very low coverage, which was converted to 0.1% for data analyses. Additionally, we recorded the number (density) of live Tamarix ramosissima (hereafter Tamarix) individuals within each plot. Tamarix individuals are often hard to delineate from one another, but in our plots, the youngest sites often consisted of plants <2 m tall with a single main stem. On older sites where Tamarix density was low, individuals were typically large and distinct, allowing accurate individual counts.
Soil samples (480 cm3) were collected from the top 5 cm of soil (excluding surface litter), with half the volume collected from the southwest and half from the northeast inside corner of each plot. An additional 480 cm3 was collected (total from each of 2 plot corners) to estimate bulk density and coarse fragment content. To reflect soil conditions experienced by seeds and newly establishing plants, we measured 0–5 cm soils, a depth that Guo et al. (Citation1998) reported contained 97% of seeds in Mojave Desert seed banks. Soils were air dried, passed through a 2 mm sieve, and analyzed for texture (hydrometer method); calcium carbonate (CaCO3; manometer method); total C, N, and sulfur (S; dry combustion, elemental CNS analyzer); organic C (difference between total and inorganic C), and electrical conductivity (saturated paste, presented in dS/m where 1 dS/m = 1 mS/cm = 1000 μS/cm) following Burt (Citation2004). Bulk density was estimated by sieving through a 2 mm sieve, oven drying the <2 mm fraction at 105 C for 24 h, and making calculations both with and without volume of coarse fragments >2 mm included in the total soil volume.
Data analysis
We calculated species richness and cover for each plot and grouped species into 4 groups for each metric: native annual, native perennial, exotic annual and exotic perennial, as defined by the US Department of Agriculture (USDA) PLANTS database (http://plants.usda.gov/). Washingtonia filifera, while listed as native to North America on the USDA-PLANTS website, is not considered indigenous to the area. This species is thought to have been moved to the area by humans and planted around springs, which is supported by records kept by Lake Mead National Recreation Area (Boulder City, NV). As a result, we included this species in the “exotic” category, which is consistent with other parks in the northern Mojave Desert such as Death Valley National Park, where the species can become invasive around springs and wetlands (Holmquist et al. Citation2011).
Table 1 Mean cover (% ± 1 SE) of exotic species found along the Lake Mead shoreline sampled 3, 6, 9, and 13 years since submersion, and sites that have never been submerged (control). “<0.1” indicates species that were present but had a mean cover <0.1%. Data presented are means across all plots and sites within that age class (n = 30).
We analyzed cover (total and relative), richness, and Tamarix density using a 2-factor analysis of variance with time since submersion (surface age) and site as fixed effects and transect nested within site as a repeated measure in SAS software (v.9.2, PROC MIXED, Tukey's method for post hoc comparisons; SAS Institute Citation2009). Post hoc multiple comparisons are referred to in the text as statistically different when Tukey's-derived p-values are <0.05. Tamarix density and cover, and relative exotic annual, native annual, and native perennial cover were log transformed to meet assumptions of normality.
Soil characteristics were analyzed using the same analysis of variance model as for plant richness and cover. Nitrogen, organic C, total S, and electrical conductivity were log transformed to meet assumptions of normality. We ordinated soil variables using principal components analysis (PCA; cross-products matrix derived from correlation) in PC-Ord software (McCune and Mefford Citation1999) to examine variation among sites and surface ages. Multi-response permutation procedures, performed in PC-ORD, were utilized to examine differences in multivariate soils composition among sites and among surface ages within sites.
Results
Species richness and cover
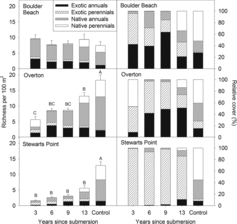
On the 150 plots we recorded 118 species consisting of 11 exotic, 67 native annual, and 40 native perennial species. Total richness increased significantly with surface age at the Overton site, where the youngest plots had the fewest species and the oldest had significantly more but not yet equivalent to control plots (site × year interaction: F8,95 = 7.53, P < 0.0001; ). There also was a trend toward increasing richness with surface age at Stewarts Point, but at Boulder Beach, species richness did not differ significantly among any surfaces.
Differences in total richness within sites were driven largely by richness of native annual and native perennial species (year effect on exotic annual: >0.05; native annual F4,95 = 18.5, P < 0.0001; native perennial F4,95 = 64.41, P < 0.0001; ). Native annual species richness did not differ between control and 13-year-old surfaces at Overton, but younger surfaces had lower richness, showing reestablishment of native annual species after 13 years (site × year interaction: F8,95 = 8.12, P < 0.0001); however, native annual richness was lower in all previously submerged plots than in the control plots at Stewarts Point and did not differ among surface ages at Boulder Beach. Richness of native perennial species increased with surface age at all sites (site × year interaction: F8,95 = 6.47, P < 0.0001). At the Boulder Beach site, richness in the oldest disturbed plots was equivalent to the control plots.
The sole exception to the pattern of increasing native perennial richness with plot age was the Overton site, where native perennial richness in the youngest plots did not differ from control plots. This was related to the proximity of the Overton site to the Muddy River riparian corridor, where 3-year-old surfaces transitioned from submerged lake to wetland habitat supporting more perennial littoral and riparian species than the other sites (Supplemental Appendix 1). While no rare species were detected within the boundaries of the study plots, at Stewarts Point we observed establishment of Arctomecon californica immediately adjacent to one of the 13-year-old plots (and in the vicinity of 2 other 13-year-old plots), indicating that it is possible for rare species to recolonize the disturbed area.
Total plant cover did not vary with surface age within Boulder Beach or Stewarts Point; however, within the Overton site, cover was greater in the youngest plots (62 ± 11%; mean ± 1 SE) than in 9-year-old (26 ± 5%), 13-year-old (21 ± 7%), or control plots (26 ± 5%; site × year interaction: F8,95 = 2.56, P = 0.01). This related to the influence of Typha domingensis, which averaged 26% ± 12% cover on the youngest surfaces, 4 of which were near the banks of the Muddy River. Typha was absent from all other plots. Relative cover of native perennial species increased with time since submersion in all sites except Stewarts Point, where the only statistical differences were between formerly submerged and control plots (site × year interaction: F8,95 = 9.19, P < 0.0001; ).
Species composition
Across sites, Tamarix was the most abundant species, particularly on the younger surfaces. Larrea tridentata, which was almost ubiquitous on undisturbed surfaces, was only rarely observed (at the Boulder Beach site) on the disturbed surfaces (Supplemental Appendix 1). Native perennial species were sparse across all 3 sites, with an increase in frequency with site age, most often with species such as Ambrosia dumosa, Bebbia juncea, and Encelia farinosa only found in the 13-year-old sites. Eriogonum inflatum and Stephanomeria pauciflora are 2 native perennial species that were found at 9-year-old or younger sites, but younger sites most often entirely lacked native perennial establishment. Native annual species established more prolifically than native perennial species, and annual community composition was more variable among sites. Eriogonum deflexum was the species most abundant and common across all 3 sites; however, Chaenactis fremontii, Cryptantha sp., Pectocarya platycarpa, and Phacelia crenulata were common within Boulder Beach and Overton sites, but lacking at Stewarts Point.
Exotic species
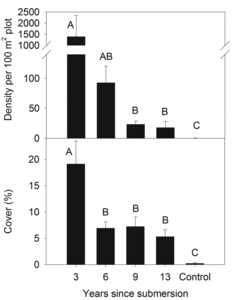
Among 11 exotic species detected on plots, Tamarix was most abundant (). As hypothesized, Tamarix density and cover decreased with surface age (year: F4,107 = 22.26, P < 0.001), with a similar pattern in cover (year: F4,95 = 11.60, P < 0.001; ).
Soil properties
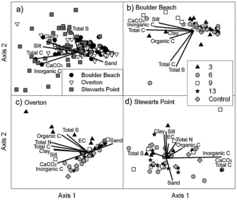
The first 2 components within the soil PCA accounted for 60% of the variance at Boulder Beach, 77% at Overton, and 57% at Stewarts Point. While there was little clear segregation among sites, soils at Boulder Beach and Overton were more similar to each other than to those at Stewarts Point, which had a larger breadth of composition and generally had greater total S, CaCO3, and inorganic C (multiple response permutation procedure [MRPP]: A = 0.09, P < 0.0001; ).
There were generally few differences in soil within sites among age groups of formerly submerged plots, and also a lack of conspicuous differences between submerged plots and control plots (–). The exception to this was that some of the younger surfaces at Overton had a stronger correlation with electrical conductivity (EC), total S, organic C, total N, total C, and a greater percentage of clay and silt. Similarly, at Boulder Beach several of the submerged surfaces across a range of ages have strong associations with inorganic C, total C, organic C, CaCO3, silt, and clay. At each of these 2 sites, variation in soil composition within control plots was lower than variation within formerly submerged plots, which had large variation in soils; however, there were no overall differences among ages at the 3 sites (MRPP: P > 0.05).
EC decreased on older surfaces at Boulder Beach, with an average of 3.85 ± 0.90 dS/m on the youngest surfaces and 1.29 ± 0.53 dS/m on 13-year-old surfaces (the oldest plots did not differ from control sites; site × year: F8,95 = 3.27, P = 0.003). EC at Overton showed the same pattern, with a mean of 11.19 ± 3.72 dS/m for the youngest surfaces and 0.96 ± 0.18 dS/m for 13-year-old surfaces (which also did not differ from control sites). EC did not differ with surface age at Stewarts point. Soil analyses at this site reflected gypsum parent material, with greater EC and 9 times greater total S concentrations than at other sites (site: F2,27 = 36.67, P < 0.0001).
Discussion
Vegetation
Two of 3 sites (Overton and Stewarts Point) exhibited the general pattern that total species richness increased with surface age, whereas the third site (Boulder Beach) showed no pattern with surface age. Native species richness generally displayed the same pattern as total richness. Other successional studies in North American deserts have collectively reported little relationship between species richness and time since disturbance or surface age; most studies have found that richness recovers to or exceeds that of undisturbed levels shortly after succession begins (Abella Citation2010). In a study of primary succession on debris flows around Grand Canyon, Arizona, for example, Bowers et al. (Citation1997) found that 80% of sites younger than the oldest site (∼3000 years) examined had greater species richness than the oldest site. The pattern was different in our study where the oldest surfaces had either similar or significantly greater richness than younger surfaces. Considering only native perennial species richness, however, we did find a trend in our study similar to Bowers et al. (Citation1997) where richness on our 13-year-old surfaces overall began to most closely resemble the control plots.
The hypothesis that native plant colonization is greatest on the oldest surfaces was supported, except that Overton had high relative cover of native plants on both the youngest and the oldest submerged surface. Aside from Overton, where the youngest site exhibited large native perennial relative cover due to an abundance of Typha domingensis adjacent to the riverbank (Supplemental Appendix 1), apparently about 13 years is required for appreciable native perennial plant colonization to occur during this primary succession. On Grand Canyon and Death Valley debris flows, significant native perennial plant cover had accrued on 5-year-old surfaces (Webb et al. Citation1987, Bowers et al. Citation1997).
Exotic plant dominance declined with increasing surface age. This pattern was largely driven by Tamarix, which sharply declined in density and cover from the youngest to the oldest surfaces. These results concurred with those of a previous study in the Boulder Basin of Lake Mead, where Tamarix dominated within 200 m of the water's edge but sharply declined at greater distances and was absent beyond 250 m (Walker et al. Citation2006). The presence of dense, dead Tamarix stems on the older, formerly submerged surfaces suggested that environmental conditions on these surfaces were no longer suitable for survival of adult plants or for recruitment. Presumably this decline was linked to depletion of subsurface water as the lake receded, making conditions too dry to support Tamarix.
Other exotic species displayed little pattern with surface age, apparently consistent with their abilities to occupy either moist or dry sites. For instance, this study's finding of Bromus rubens inhabiting recently submerged surfaces near the shoreline and never-submerged surfaces concurs with a landscape-scale distribution study of Bromus rubens within Lake Mead National Recreation Area, where the species occupied a variety of riparian and upland sites (Abella et al. Citation2012).
Soil
The sites themselves differed from each other in their soil and topographic characteristics, but the finding of no difference in soil properties among surface ages was consistent across this diversity of sites. Further, few univariate patterns were strong (Supplemental Appendix 2). EC, correlated with salt concentration, was highest on the youngest surface at Boulder Beach and Overton but displayed no consistent pattern at Stewarts Point. The high conductivity and total S concentration on the never-submerged surface at Stewarts Point relative to other sites likely relates to properties of the gypsum parent material (Sheldon Thompson and Smith Citation1997).
We had anticipated that processes such as deposition of fine soil fractions during submersion might alter texture. Unexpectedly, soil texture did not differ with time since submersion, suggesting that submersion did not alter soil texture or that the net effects of submersion or subsequent exposure resulted in no textural changes. While all tributaries to Lake Mead carry heavy sediment loads (Holdren and Turner Citation2010), we did not see lasting effects of silt deposition on younger soil surfaces. Variation among sites differing in parent materials suggests that the underlying soil type of a site might more strongly influence surface soil properties than did submersion. Sediments deposited during submersion may not be detected years later due to redistribution by shallow water turbulence or dispersal by desert winds after the waters receded.
Our results of no strong overall difference in soils among surfaces and individualistic variation of specific soil properties concurs with a previous study by Walker et al. (Citation2006) in Lake Mead's Boulder Basin and another by Tallent et al. (Citation2011) downriver around Lake Mohave's shoreline. Walker et al. (Citation2006) found that soil total N and organic matter did not differ between submerged and non-submerged surfaces, while salinity and gravimetric moisture did differ between submerged and non-submerged surfaces but not among surface ages within submerged sites.
Walker et al. (Citation2006) and Tallent et al. (Citation2011) found that salinity was positively correlated with Tamarix abundance. Our study did not find a direct relationship between salinity and Tamarix abundance, possibly related to several factors. Due to the stochastic nature of water availability in this system, the sites with greatest Tamarix abundance were also the youngest, and the plants may have established recently enough in the previously inundated land to have little effect on salts in the soil profile. Additionally, the soils are subject to submersion periodically with the intraannual lake level fluctuations, which could flush the system of additional salts. Lastly, the direct link between Tamarix and significantly increased soil salinity is not necessarily a causal relationship at all sites, but may be coupled with Tamarix's high tolerance of salinity and ability to establish in saline conditions (Ladenberger et al. 2006, Stromberg et al. 2009).
Given that overall soil was not appreciably different among surfaces, and only some individual variables displayed patterns with submersion status, an important unresolved question is whether native plant colonization is in any way limited by properties of formerly submerged soil. Most soil properties of formerly submerged surfaces were similar to never-submerged surfaces and thus were unlikely to limit native plants relative to never-submerged areas. Moreover, with an electrical conductivity of 11 dS/m, only the youngest Overton surface exceeded the 8 dS/m tolerance range of most species. According to Shafroth et al. (Citation2008), electrical conductivity ≤4 dS/m is satisfactory for plant growth, 4–8 dS/m limits some species, and 8–16 dS/m supports salt-tolerant species. It is unclear whether observed salinity levels would be sufficient to restrict native plant establishment, or whether desert plants locally adapted to gypsum parent materials (much of the Stewarts Point site falls within the drygyp series) had greater abilities to tolerate salinity. We have some evidence of gypsoclinic species (Astragalus preussii, Atriplex hymenelytra), establishing on the gypsiferous soils in the Stewarts Point site and not at other sites; however, due to general successional dynamics in the Mojave, rates of establishment were low (generally 10–20% frequency and only in the oldest sites; Supplemental Appendix 1).
Conducting experimental plantings or seedings of native species in future research along the shoreline might help directly resolve if soil limits native plant colonization relative to other limiting factors such as seed dispersal and protected microsites for establishment (James et al. Citation2005). Research on these topics would actively inform managers on management techniques necessary to restore or create habitat in exposed land post-inundation.
Plant succession principles and projections
Results from this study support some of the (relatively recently) established successional principles developed for Mojave Desert ecosystems (Abella Citation2010, Allen et al. Citation2011). First, the observed early colonization by both annual and perennial plants concurs with the general principle that both plant groups are represented in both early and late-successional desert communities. Second, many of the colonizing perennial species, such as Hymenoclea salsola, Bebbia juncea, and Stephanomeria pauciflora, also inhabit washes in nonanthropogenically disturbed ecosystems. Washes are natural environments periodically disturbed by flooding, and species inhabiting them colonize a variety of other disturbed environments, including the Lake Mead shoreline. Third, Ambrosia dumosa was both an early colonizer of the new shoreline and an inhabitant of the never-submerged community, consistent with the principle that deserts contain “versatile” species found throughout a successional sequence. Fourth, exclusive late-successional species such as Larrea tridentata remained sparse after 13 years of surface exposure, consistent with the principle that establishment of late-successional communities requires long time periods on the order of decades to centuries.
Several factors can delay recruitment of late-successional species in deserts, such as rare occurrence of conditions suitable for recruitment, seed availability (many native perennials do not form persistent soil seed banks), and absence of “nurse” plants that afford protection and provide “safe sites” for seedling establishment (McAuliffe Citation1988).
We found that total perennial plant cover on the 13-year formerly submerged plots reached an average of 43% of perennial cover in control plots at the Boulder Beach site and 56% of the perennial cover at the Overton site. This finding was similar to the reestablishment of perennial plant cover (49% after 13 years post-disturbance) reported from the average of 29 studies of secondary succession following clearing and fire disturbances in the Mojave and Sonoran Deserts (Abella Citation2010). Perennial cover in disturbed plots at the Stewart's Point site was 250% perennial cover in the control plots, however, largely due to the retention of Tamarix individuals in the older Stewarts Point sites in this site whereas the proportion of Tamarix was much lower in the Boulder Beach and Overton sites ().
If vegetation change continues along trends of the first 13 years and is consistent with longer term patterns of other studies (Abella Citation2010), then a potential scenario of change can be projected. For example, an early successional native perennial plant community dominated by Stephanomeria pauciflora, Hymenoclea salsola, and Ambrosia dumosa inhabited a 38-year-old bulldozed area of never-submerged terrestrial habitat just northwest of our Boulder Beach site (Abella et al. Citation2007). This was consistent with the persistence of these early successional communities reported in other studies (Abella Citation2010), and the community contained 4 times the perennial plant density and similar species richness as undisturbed old Larrea tridentata communities. Assuming the shoreline is not resubmerged, an early successional community similar in species composition to sites that have incurred surface disturbance from construction, fire, or other disturbance will likely continue to persist for decades before colonization by species of old surfaces, given suitable climatic conditions (Webb et al. Citation1987, Bowers et al. Citation1997). As illustrated in Abella et al. (Citation2007) and other studies (Abella Citation2010), these early successional communities can have total plant cover, density, and richness that exceeds those of old communities.
Management implications
Fluctuating lake levels complicate developing shoreline habitat management strategies. Management activities such as controlling exotic plants or actively establishing native species near the current land–lake interface can be counteracted when lake levels rise, which occurred in 2012 following this study. While fluctuations within and across years result in both increases and decreases in the elevation of Lake Mead, scientific consensus indicates that a return to full pool elevation is unlikely within future decades (Barnett and Pierce Citation2008, Barsugli et al. Citation2009). Therefore, even if some of the currently exposed shoreline becomes reinundated, there will still be thousands of hectares of newly exposed land.
A possible active management strategy would involve identifying a potential new upper lake level (Barnett and Pierce Citation2009) and prioritizing efforts at elevations above that projected lake level. Another strategy might be to target or create islands of land higher than surrounding areas that would still be above water if the lake rises (Alice Newton, Lake Mead National Recreation Area, pers. comm, 2011). This strategy might be most effective in areas of gentler topography and may provide “refuges” for terrestrial organisms and seed sources for rare or endemic species supported by soil composition and structure specific to certain sites (e.g., gypsophiles), allowing them to repeatedly colonize a fluctuating environment (Bainbridge Citation2007).
A positive aspect of the fluctuating lake level is that inundation is not optimal for Tamarix regeneration while continued reservoir recession has resulted in vast Tamarix die-off areas (Walker et al. Citation2006, this study). Further research that explores whether Tamarix or any of its possible soil legacy effects interferes with native plant colonization would be useful (Shafroth et al. Citation2008). If revegetating priority sites is a management goal, planting can be a viable and efficient option for revegetation of disturbed sites that would otherwise experience slow natural colonization rates. Previous research has shown that planting nursery-grown stock using proper practices (e.g., timing plantings properly) can achieve ≥50% survival for perennial plants in the Mojave Desert (Bainbridge Citation2007, Abella and Newton Citation2009). As a result, active revegetation can establish native perennials relatively quickly. Another unexpected benefit from dropping lake levels may be the establishment of wetlands along river corridors that have not functioned as riparian plant communities since pre-dam days. We observed this in the Overton site, where the lake receded below the elevation of the river that flows into Lake Mead, resulting in establishment of riparian communities dominated by Typha domingensis along the Muddy River floodplain where the land was formerly submerged by the lake.
The past 15 years of overall drawdown of Lake Mead has re-exposed a dynamic new terrestrial habitat. Over time, exotic plant cover has generally decreased while native perennial plant cover has increased in this habitat. Moreover, at least one rare plant species of special conservation status (Arctomecon californica) had colonized the new habitat near sampling plots. These observations suggest that while declining water levels might be viewed negatively from a perspective of maintaining full-pool water storage capacity, the decline has benefited native terrestrial habitat by exposing surfaces for native species colonization.
Acknowledgments
We thank Alice Newton and Kent Turner (LMNRA) for facilitating this study; S. Tran, K. Choy, J. Curtis, and K. Edwards for field and lab assistance; Mark Sappington (LMNRA) for calculating shoreline metrics for ; and the Associate Editor and 3 anonymous reviewers for reviewing the manuscript.
Funding
This study was funded through a cooperative agreement organized by Alice Newton (Lake Mead National Recreation Area [LMNRA]) between the National Park Service (LMNRA) and the University of Nevada Las Vegas (UNLV).
Supplemental Material
Supplemental data for this article can be accessed on publisher's website.
Supplement_Appendix_2.doc
Download MS Word (184 KB)Supplement_Appendix_1.doc
Download MS Word (466 KB)References
- Abella SR. 2010. Disturbance and plant succession in the Mojave and Sonoran Deserts of the American Southwest. Int J Environ Res Public Health 7:1248–1284.
- Abella SR, Embrey TM, Schmid SM, Prengaman KA. 2012. Biophysical correlates with the distribution of the invasive annual red brome (Bromus rubens) on a Mojave Desert landscape. Invasive Plant Sci Manage. 5:47–56.
- Abella SR, Newton AC, Bangle DN. 2007. Plant succession in the eastern Mojave Desert: an example from Lake Mead National Recreation Area, southern Nevada. Crossosoma. 33(2R):45–55.
- Abella SR, Newton AC. 2009. A systematic review of species performance and treatment effectiveness for revegetation in the Mojave Desert, USA. In: Fernandez-Bernal A, De La Rosa MA, editors. Arid environments and wind erosion. Hauppauge (NY): Nova Science Publishers. p. 45–74.
- Ali MM. 2006. Shoreline vegetation of Lake Nubia, Sudan. Hydrobiologia. 570:101–105.
- Allen EB, Steers RJ, Dickens SJ. 2011. Impacts of fire and invasive species on desert soil ecology. Rangeland Ecol Manage. 64:450–462.
- Auble GT, Shafroth PB, Scott ML, Roelle JE. 2007. Early vegetation development on an exposed reservoir: implications for dam removal. Environ Manage. 39:806–818.
- Bainbridge DA. 2007. A guide for desert and dryland restoration. Washington (DC): Island Press.
- Barnett TP, Pierce DW. 2008. When will Lake Mead go dry? Water Resour Res. 44, W03201, doi:10.1029/2007WR006704
- Barnett TP, Pierce DW. 2009. Sustainable water deliveries from the Colorado River in a changing climate. P Nat Acad Sci-USA. 106:7334–7338.
- Barsugli JJ, Nowak K, Rajagopalan B, Prarie JR, Harding B. 2009. Comment on “When will Lake Mead go dry?” by Barnett TP, Pierce DW. Water Resour Res. 45, W09601, doi:10.1029/2008WR007627
- Bowers JE, Webb RH, Pierson EA. 1997. Succession of desert plants on debris flow terraces, Grand Canyon, Arizona, USA. J Arid Environ. 36:67–86.
- Busch DE, Smith SD. 1995. Mechanisms associated with decline of woody species in riparian ecosystems of the Southwestern US. Ecol Monogr. 65:347–370.
- Burt R. 2004. Soil survey laboratory methods manual. Soil survey investigations. Washington (DC): US Department of Agriculture, Natural Resources Conservation Service. Report no. 42, version 4.0.
- Chandra S, Abella SR, Albrecht BA, Barnes JG, Engel EC, Goodbred SL, Holden PB, Kegerries RB, Jaeger JR, Orsak E, et al. 2012. Wildlife and biological resources. In: Rosen MR, Turner K, Goodbred SL, Miller JM, editors. A synthesis of aquatic science for management of Lakes Mead and Mojave. . US Geological Survey Circular 1381. p. 70–104.
- Guo Q, Rundel PW, Goodall DW. 1998. Horizontal and vertical distribution of desert seed banks: patterns, causes, and implications. J Arid Environ. 38:465–478.
- Hill NM, Keddy PA, Wisheu IC. 1998. A hydrological model for predicting the effects of dams on the shoreline vegetation of lakes and reservoirs. Environ Manage. 22:723–736.
- Holdren GC, Turner K. 2010. Characteristics of Lake Mead, Arizona–Nevada. Lake Reserv Manage. 26:230–239.
- Holmquist JG, Schmidt-Gengenbach J, Slaton MR. 2011. Influence of invasive palms on terrestrial arthropod assemblages in desert spring habitat. Biol Conserv. 144:518–525.
- James JJ, Tiller RL, Richards JH. 2005. Multiple resources limit plant growth and function in a saline-alkaline desert community. J Appl Ecol. 93:113–126.
- Ladenburger CG, Hild AL, Kazmer DJ, Munn LC. 2006. Soil salinity patterns in Tamarix invasions in the Bighorn Basin, Wyoming, USA. J Arid Environ. 65:111–128.
- Lato LJ. 2006. Soil survey of Clark County area, Nevada. Washington (DC): US Department of Agriculture, Natural Resources Conservation Service.
- McAuliffe JR. 1988. Markovian dynamics of simple and complex desert plant communities. Am Natl. 131:459–490.
- McCune B, Mefford MJ. 1999. PC-ORD: multivariate analysis of ecological data. Version 4. User's guide. Gleneden Beach (OR): MjM Software Design.
- Rosen MR, Turner K, Goodbred SL, Miller JM, editors. 2012. A synthesis of aquatic science for management of Lakes Mead and Mojave: US Geological Survey Circular . 1381, 162 p.
- SAS Institute. 2009. SAS/STAT 9.2 user's guide. Cary (NC): SAS Institute, Inc.
- Shafroth PB, Beauchamp VB, Briggs MK, Lair K, Scott ML, Sher AA. 2008. Planning riparian restoration in the context of Tamarix control in western North America. Restor Ecol. 16:97–112.
- Sheldon Thompson SK, Smith SD. 1997. Ecology of Arctomecon californica and Arctomecon merriamii (Papaveraceae) in the Mojave Desert. Madroño. 44:151–169.
- Stromberg JC, Chew MK, Nagler PL, Glenn EP. 2009. Changing perceptions of change: The role of scientists in Tamarix and river management. Restor Ecol. 17:177–186.
- Tallent N, Nash M, Cross CL, Walker LR. 2011. Patterns in shoreline vegetation and soils around Lake Mohave, Nevada and Arizona: implications for management. West N Am Natl. 71:374–387.
- Walker LR, Barnes PL, Powell EA. 2006. Tamarix aphylla: a newly invasive tree in southern Nevada. West N Am Natl. 66:191–201.
- Walker LR, del Moral R. 2003. Primary succession and ecosystem rehabilitation. New York: Cambridge University Press.
- Webb RH, Steiger JW, Turner RM. 1987. Dynamics of Mojave Desert assemblages in the Panamint Mountains, California. Ecology. 68:478–490.