
ABSTRACT
James WF. 2017. Internal phosphorus loading contributions from deposited and resuspended sediment to the Lake of the Woods. Lake Reserv Manage. 33:347–359.
The Lake of the Woods exhibits cyanobacterial blooms despite substantial declines in watershed phosphorus (P) loading since the 1970s, suggesting that internal P loads are contributing to the P budget. A better understanding of internal P loading contributions and dynamics is needed to refine management strategies and water quality goals. Sediment cores were collected from several stations within the US portion to examine potential internal P contributions from deposited and resuspended sediment. Although laboratory-derived diffusive P fluxes were highest under anaerobic conditions (8−12 mg/m2 d), aerobic conditions at the sediment–water interface may regulate in situ diffusive P fluxes due to frequent polymixis. Although much lower under aerobic conditions, modest fluxes of 0.2−0.6 mg/m2 d could play an important role in the P budget. Simulated resuspension indicated that the critical shear stress of deposited sediment was relatively low at 1.3−2.3 dynes/cm2. The wind-exposed long fetches and shallow morphometry of regions of Lake of the Woods could result in frequent resuspension of P-enriched clays and silts that contribute inorganic P to the water column. Resuspended sediments exhibited a low equilibrium P concentration (near zero) and high linear adsorption coefficient (700−3200 L/kg), suggesting they may act more as a sink rather than a source for soluble P. Resuspension could compete with cyanobacterial uptake by sequestering soluble reactive P. Resuspended inorganic P could also constitute a significant portion of the water column total P composition in addition to organic P, incorporated as cyanobacterial biomass, and needs to be considered in the P budget and chlorophyll prediction.
The Lake of the Woods (LOW), located on the border of Ontario and Manitoba, Canada, and Minnesota, USA, is a large (3850 km2), morphologically complex system that drains north into the Winnipeg River (). LOW receives up to 75% of its external phosphorus (P) loading (754 T; i.e., all point and nonpoint sources plus atmospheric deposition) from the Rainy River (568 T) and ∼54% is retained in the system (Hargan et al. Citation2011). Cyanobacterial blooms have commonly occurred in the southern portion of LOW for more than a century, which may be associated, in part, with glacial deposits naturally high in P (Pla et al. Citation2005). Although Rainy River total P (TP) loading has declined substantially from peaks exceeding 1500 to 2500 T/y in the 1970s, observations of continued cyanobacterial bloom and toxin development (Chen et al. Citation2009), particularly in the southern US portion of LOW, suggest that internally derived sources of P may be playing an increasingly important role in subsidizing algal growth (Pla et al. Citation2005, Zhang et al. Citation2013). Recent empirical P modeling analysis of LOW has found that up to 36% of the P budget may be derived from unmeasured or internal P loading sources (LOW Citation2014). These unidentified sources likely include diffusive P fluxes from deposited sediment historically retained in the lake and equilibrium P fluxes from resuspended sediment.
Figure 1. Sediment sampling station locations in the Lake of the Woods.
Big Traverse, Muskeg, and Four-Mile bays, located to the south in the US portion of LOW, are relatively shallow and expansive with a low morphometric index, suggesting frequent water column turbulence and mixing (). Internal P loading is often wrongly assumed to be negligible in large, shallow polymictic systems and not considered in P budget analysis (Nürnberg Citation1998, Citation2005, Orihel et al. Citation2015). Although frequent dissolved oxygen mixing to the sediment surface occurs in these systems, internal P loading via anaerobic diffusive P flux can still constitute a significant portion of the P budget and drive cyanobacteria blooms (Nürnberg et al. Citation2013a, Citation2013b, Orihel et al. Citation2015, Nürnberg and LaZerte Citation2016). Although difficult to detect, hypoxia and anoxia can develop rapidly at the sediment–water interface during periods of low wind activity (Nürnberg and LaZerte Citation2016). P flux from deposited sediment can occur under anaerobic conditions via reduction of iron (Fe3+) oxyhydroxides (Fe-(OOH)) to Fe2+, subsequent desorption of PO4, and diffusion into the overlying water column (Mortimer Citation1971, Nürnberg Citation1988, Loh et al. Citation2013). While anaerobic iron sulfide formation can also play a role in P release under anoxic conditions (Caraco et al. Citation1989, Roden and Edmonds Citation1997, Kleeberg Citation1998, Orihel et al. Citation2015), this mechanism is probably minor in iron-rich sediments of LOW.
Table 1. Morphometric and summer trophic state characteristics of the 3 southern bays in the Lake of the Woods.
Under oxic conditions at the sediment–water interface, as is likely the case for most regions of southern LOW bays, diffusive P flux is much lower and largely controlled by adsorption of phosphate (PO4) onto Fe-(OOH) at the thin oxic microzone (Nürnberg Citation1984). Aerobic diffusive P flux can nevertheless be detectable and represent an important source to the P budget of large, shallow, polymictic systems. For example, Doig et al. (Citation2017) measured relatively high aerobic soluble P release rates ranging between 0.17 and 3.35 mg/m2 d in laboratory sediment core systems collected from Lake Diefenbaker (Canada) and incubated at 15 C. These rates were comparable to anaerobic soluble P release rates from other mesotrophic to eutrophic systems (Nürnberg Citation1988, Carter and Dzialowski Citation2012). P release from oxic sediment can also be achieved via mineralization of recently deposited organic P (Reitzel et al. Citation2007), microbially mediated polyphosphate accumulation and release as PO4 (Gächter et al. Citation1988, Gächter and Meyer Citation1993, Hupfer et al. Citation2007, Citation2008), diffusion through a, Fe-(OOH)∼P saturated oxic microzone, and extrusion of porewater during resuspension events (Søndergaard et al. Citation2001, Cyr et al. Citation2009).
Frequent resuspension of particulate P can also represent a significant internal P flux to the water column of large, shallow, polymictic lakes, often comparable to tributary P loading (Kelderman et al. Citation2012, Matisoff and Carson Citation2014, Matisoff et al. Citation2017). Release of soluble P and bioavailability of resuspended particulate P may be low, however, depending on equilibrium conditions between solid and aqueous phases (Søndergaard et al. Citation1992, de Vicente et al. Citation2010) and seasonal variations in temperature and pH (Holmroos et al. Citation2009). P flux from resuspended sediment can occur via equilibrium reactions between an exchangeable particulate P pool and aqueous soluble P (Froelich Citation1988, Søndergaard et al. Citation2003). Resuspended sediments can either desorb or adsorb PO4 under conditions of aqueous soluble P disequilibrium, depending on the extent of P saturation of binding sites on the exchangeable particulate P pool and the aqueous equilibrium P concentration (EPC; Cyr et al. Citation2009). For instance, P desorption from resuspended sediment can occur if aqueous soluble P declines below the EPC due to algal uptake. Conversely, P adsorption onto the exchangeable P pool can occur if aqueous soluble P increases above the EPC due to a watershed loading event (James and Larson Citation2008). Thus, resuspended sediment could act as a source or sink for soluble P, depending on equilibrium relationships between the particulate exchangeable and aqueous P pool. Frequently resuspended inorganic P during the summer could represent a significant portion of the water column TP not incorporated into algal biomass (Effler and O'Donnell Citation2010, Effler et al. Citation2014, Gelda et al Citation2016). If so, empirical TP to chlorophyll a (TP:Chl-a) models would tend to overpredict summer chlorophyll.
The objectives of this research were to examine P flux potential from deposited and resuspended sediment to provide further insight into processes that may be responsible for the unaccounted for P mass in LOW. Intact sediment cores collected from south bays of LOW were incubated under aerobic and anaerobic conditions to estimate diffusive P fluxes from deposited sediments. Additional cores were subjected to shear stress-induced resuspension to quantify soluble P exchanges in relation to the EPC. The outcome of these investigations will be important in better understanding and managing cyanobacterial bloom development in LOW.
Methods
Sediment sampling procedures and locations
Intact sediment cores (∼30 cm in length) were collected from stations located in Muskeg Bay, Big Traverse Bay East, Big Traverse Bay West, and Four-Mile Bay in 2011 for determination of sediment textural and chemical characteristics, diffusive P flux, and P equilibrium characteristics during simulated sediment resuspension (). A gravity coring device was used to collect the cores (Aquatic Research Instruments, Hope, ID). The core liners, containing both sediment and overlying water, were immediately sealed with stoppers and stored in protective carrying cases cooled during transport to the laboratory.
Sediment characteristics and phosphorus fractions
The upper 10 cm was sectioned from a core collected at each station for analysis of moisture content, density, organic matter content, particle size distribution, and P fractions. A known volume of sediment was dried at 105 C for determination of moisture content and density and burned at 550 C for determination of loss-on-ignition organic matter content (Håkanson and Jansson Citation2002). The percentages of sand (>63 μ), silt (<63 μ and >2 μ), and clay (<2 μ) were estimated using settling columns and pipette techniques (Plumb Citation1981).
Sequential P fractionation was conducted according to Nürnberg (Citation1988) and Psenner and Puckso (Citation1988) to estimate loosely bound P (1 M ammonium chloride), Fe-bound P (0.11 M bicarbonate-dithionite), aluminum-bound P (0.1 N sodium hydroxide), and calcium-bound P (0.5 N hydrochloric acid). A subsample of the sodium hydroxide extract was digested with potassium persulfate to measure nonreactive sodium hydroxide-extractable P. Labile organic P was calculated as the difference between reactive and nonreactive sodium hydroxide-extractable P. Residual P was estimated as the difference between sediment total P and the sum of the other measured fractions. Each extraction was centrifuged, filtered through a 0.45 µm membrane syringe filter, and analyzed colorimetrically for soluble reactive P (SRP) on a UV-VIS spectrophotometer (Perkin-Elmer Lambda 11) according to the American Public Health Association (APHA Citation2005) using a 1 cm path length cuvette. The detection limit was 0.005 mg/L. Sediment subsamples were dried and ground for determination of total P and total Fe (expressed as mg/g dry mass). Samples for TP were digested with red mercuric oxide and potassium persulfate under acidic conditions (modification of Jirka et al. Citation1976, Bremner and Mulvaney Citation1982) prior to colorimetric analysis. For metals, sediment was digested with nitric acid, hydrochloric acid, and hydrogen peroxide (30%) and analyzed using atomic absorption spectroscopy (EPA Method 3050B).
Gross diffusive phosphorus flux from deposited sediment
Gross and net internal P loading are distinguished by the proportion of the load that settles back to the sediment annually (Nürnberg Citation2009, Nürnberg et al. Citation2012). For instance, in situ summer P mass increases, and P mass balance approaches include some deposition of internal P loads and therefore reflect a net flux (i.e., diffusive P flux input minus P deposition). Direct measurement of diffusive P flux from intact sediment cores and extrapolation to active sediment areas (Nürnberg Citation1995, Citation2009) necessarily includes past deposition of both external and internal P loads but approximates a current gross internal P load to the water column that does not include current deposition. Six replicate intact cores were collected at each station in LOW to measure gross diffusive P flux under aerobic (3 replicates) and anaerobic (3 replicate) conditions. Additional lake water was collected for later incubation with the sediment. In the laboratory, sediment cores were drained of overlying water, and the upper 10 cm of sediment was extruded intact to a smaller acrylic core incubation tube (6.5 cm diameter and 20 cm height). Filtered lake water (Gelman A-E,2 µm nominal pore size) was siphoned (300 mL) onto the sediment to serve as overlying water. The sediment incubation systems were placed in a darkened environmental chamber and incubated at 20 C to simulate peak summer bottom temperatures (M. Edlund, St. Croix Watershed Research Station, Science Museum of Minnesota, pers. comm.). The oxidation–reduction environment in the overlying water of each system was controlled by gently bubbling either air (oxic or aerobic) or nitrogen (anoxic or anaerobic) through an air stone placed just above the sediment surface (James Citation2010). Bubbling action insured complete mixing of the water column but did not disrupt the sediment. Anoxic conditions were verified using a dissolved oxygen electrode. Overlying water column pH was ∼8.3 under aerobic and anaerobic conditions, respectively. In situ summer pH can range between 7.5 and >8.0 (i.e., Jun and Aug 2016; M. Edlund and A. Heathcote, St. Croix Watershed Research Station, Science Museum of Minnesota, unpubl data). Water samples (10 mL) were collected from the center of each sediment incubation system, filtered through a 0.45 µm membrane syringe filter, and analyzed for SRP. Incubation systems were sampled daily over the first 5 days and every 2 days thereafter for up to 18 days. Rates of diffusive P flux from deposited sediment (mg/m2 d) were calculated as the linear change in concentration in the overlying water (n = 4 to 8) divided by time and the area of the incubation core liner. Regression analysis was used to estimate the rate over the linear portion of the data.
Equilibrium phosphorus characteristics
Additional wet sediment subsamples (500 mg/L dry weight equivalent) from the upper 10 cm sections collected at each station were subjected to soluble P (as KH2PO4) standards ranging from near 0 to 1.0 mg/L for examination of P adsorption and desorption after 24 h of exposure. Filtered (0.45 µm) lake water was used to prepare the P standards to maintain ionic composition. Centrifuge tubes (glass conical 50 mL capacity) containing sediment and P-amended water were shaken uniformly in a darkened environment at a temperature of 20 C, then filtered and analyzed for SRP (James and Larson Citation2008).
Linear regression relationships were developed between the mass of SRP desorbed or adsorbed (S; mg P/kg sediment) versus final equilibrium SRP (i.e., the SRP concentration after 24 h of exposure) to estimate the linear adsorption coefficient (kd; L/kg) and the EPC (in P, mg/L, Froelich Citation1988). Thus, the EPC reflected the outcome of equilibrium fluxes between suspended sediment and lake water SRP after a resuspension event. Equilibrium data were also fitted to Langmuir sorption models (Bolster and Hornberger Citation2007) to estimate the P sorption capacity for sediment (Smax; mg/kg, the maximum amount of P that can be adsorbed to binding sites on sediment surfaces). The degree of P saturation (DSP; %) for LOW sediments was calculated as the redox-P fraction (i.e., the sum of loosely bound and Fe-bound P, a surrogate measure of the sediment native exchangeable P pool) divided by Smax.
P equilibrium concentrations were also determined independently using a water extraction procedure. Water-extractable P was determined by exposing 1 g dry mass equivalent of wet sediment to 10 mL of local P-free tap water (to simulate ionic composition) for 1 h (modification of SERA-IEG 2000). Extractions were centrifuged and filtered through a 0.45 μm filter for analysis of SRP.
Equilibrium phosphorus fluxes during simulated sediment resuspension
Additional intact sediment cores (10 cm in depth) were collected using a 15 × 15 cm box corer (Wildco Wildlife Supply Co., Saginaw, MI) for laboratory simulation of sediment resuspension and P equilibrium over a range of shear stresses. Water was carefully removed from the intact cores, filtered through a glass fiber filter (Gelman A/E,), and slowly siphoned back on to the cores to serve as overlying water. A vertically oscillating particle entrainment simulator (PES) developed by Tsai and Lick (Citation1986) was used to subject intact cores to various shear stresses. The PES was programmed to oscillate an acrylic perforated disk above the sediment interface in a stepwise manner from 0 to 4 dynes/cm2 at 10 min intervals. At 8 min into each cycle, a 50 mL sample from the overlying water column was collected using a peristaltic pump. Water removed as a result of sampling was simultaneously replaced with filtered lake water using the pump. Samples were analyzed for total suspended sediment (TSS) and SRP using standard analytical procedures (APHA Citation2005). Changes in SRP as a function of shear stress were evaluated to estimate soluble P mass transfer and equilibrium SRP due to simulated sediment resuspension. The critical bottom shear stress (τc; dynes/cm2) of sediments was estimated as the inflection point where TSS increased in the water column above background conditions. The inflection point was interpreted using a segmented linear-linear regression analysis (NLIN; SAS Citation1994). Natural forces such as wave activity and flow velocity causing a shear stress (τ) at the sediment interface greater than τc will result in sediment resuspension (τ > τc).
Results
Sediment characteristics
Physical-textural characteristics of the upper 10 cm section suggested sediments were moderately compacted with higher wet and dry bulk densities and lower moisture content, porosity, and organic matter content (). Moisture content was <84% at stations in Big Traverse and Muskeg Bay and much lower, ∼60%, in Four-Mile Bay. Organic matter content was moderately low, ranging between ∼11% and 16%. The particle size distribution was co-dominated by silt (∼48%) and clay (∼50%) in Big Traverse Bay, whereas silt represented 70% of the particle size range compared to 25% clay in Muskeg Bay. The silt fraction of Four-Mile Bay sediments accounted for ∼80% and the clay fraction represented ∼14% of the sediment particle size distribution. Close proximity to Warroad and Rainy River inputs and deposition of larger, denser particles in Muskeg and Four-Mile bays likely explain the higher silt percentages at these stations. Conversely, transport and deposition of finer clay loads impacted the particle size distribution of Big Traverse Bay sediment. The sand fraction constituted <8% of the particle size distribution at all stations. Overall, correlations were weak between sediment moisture content and density versus the percent particle size distribution. Sand and silt percentages decreased while the clay percentage increased as a function of higher moisture content and low density (P < 0.10; SAS Citation1994).
Table 2. Sediment physical and chemical characteristics of the upper 10-cm sediment layer at various stations in the Lake of the Woods.
Sediment TP concentrations in the upper 10 cm layer were moderate at the 4 stations (). Big Traverse Bay sediment TP concentrations were ∼0.7 mg/g, whereas Muskeg Bay and Four-Mile Bay sediment TP ranged between 0.64 and 0.69 mg/g, respectively. Biologically labile P (i.e., the sum of the loosely bound, Fe-bound, and labile organic P fractions) accounted for 27−49% of the total sediment P (). Muskeg Bay sediments exhibited the lowest percentage of biologically labile P while nearly half of the P composition of Big Traverse Bay sediments was biologically labile (). Redox-sensitive P (redox-P, the sum of the loosely bound and Fe-bound P fractions) accounted for >50% of the biologically labile P. By contrast, labile organic P represented ∼32−49% of this fraction. The Fe-bound P fraction dominated redox-P (), and sediments from Big Traverse Bay generally exhibited highest concentrations of this fraction at ∼0.22 mg/g versus lower concentrations of ∼0.11 mg/g for sediments in Muskeg and Four-Mile bays (). Overall, correlations were positive between the clay percentage versus TP (r = 0.95; P < 0.05) and Fe-bound P (r = 0.98; P < 0.05), suggesting sediment P enrichment as a result of particle sorting during transport and deposition to Big Traverse Bay.
Figure 2. The composition of biologically labile (i.e., subject to recycling and biological uptake; loosely bound, Fe-bound, labile organic P) and biologically refractory P (i.e., more inert to recycling and uptake and subject to burial; aluminum-bound, calcium-bound, refractory organic P) fractions in the upper 10 cm sediment layer for various stations in the Lake of the Woods system.
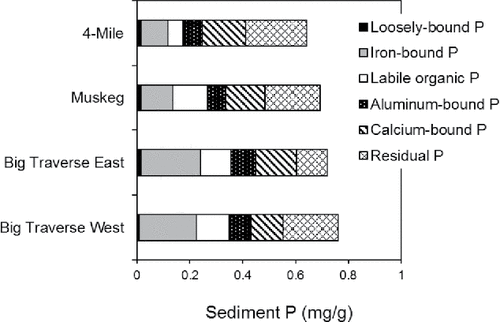
Biologically refractory sediment P represented >50% of the sediment TP composition at all stations. Residual P accounted for the much of this fraction, followed by calcium-bound and aluminum-bound P. The percentage of biologically refractory P was greatest in Muskeg and Four-Mile bays compared to Big Traverse Bay sediments.
Sediment total Fe concentrations were relatively high and greatest in Big Traverse versus Muskeg and Four Mile bays (). Sediment total Fe was also high relative to TP, resulting in Fe:P ratios that ranged between ∼26 and 43 (). The sediment Fe:P ratio was greatest in Big Traverse Bay and ∼40% lower in Muskeg and Four-Mile bay sediments.
Gross diffusive phosphorus fluxes
Mean rates of summer diffusive P flux under anaerobic conditions were high, ranging between 8.3 and 12.5 mg/m2 d (). Big Traverse West exhibited the highest mean anaerobic summer diffusive P flux. Rates were lower and similar at ∼8.5 to 9.0 mg/m2 d for Big Traverse East, Muskeg Bay, and Four-Mile Bay sediments. Anaerobic diffusive P fluxes estimated for southern bays of LOW fell within the upper end of ranges reported for other large, shallow polymictic systems in Canada and the northern United States (). Mean summer rates of diffusive P flux were much lower under aerobic conditions, ranging between ∼0.2 and 0.6 mg/m2 d (). Mean summer aerobic diffusive P fluxes were greatest for Big Traverse Bay sediment at 0.40 (0.12 SE) and 0.56 (0.26 SE) mg/m2 d for the East and West station, respectively. By comparison, mean summer aerobic diffusive P fluxes were only ∼0.2 mg/m2 d for Muskeg Bay and Four-Mile Bay sediments.
Figure 3. Mean (1 standard error, n = 3) rates of diffusive P flux from intact sediment cores under anaerobic and aerobic conditions for various stations in the Lake of the Woods.
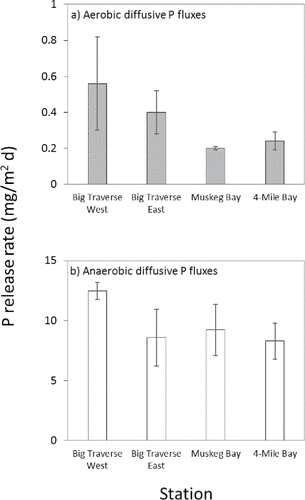
Table 3. A comparison of rates of aerobic and anaerobic rates of phosphorus (P) release from large, shallow polymictic systems in Canada and northern USA determined from laboratory incubations.
Sediment phosphorus equilibrium characteristics
Overall, sediments from LOW exhibited undetectable P desorption when subjected to low aqueous P concentrations (). As aqueous P concentration increased, sediments adsorbed P from solution. This pattern suggested that resuspended sediments were most likely a sink for PO4 and sequestered P rather than desorbing it under conditions of P disequilibrium. The EPC was near detection at ∼0.01 mg/L for Big Traverse Bay sediment and below detection limits for Muskeg and Four-Mile bay sediments (). Similarly, water-extractable P concentrations were relatively low at ∼0.010 mg/L or less (). The linear adsorption coefficient reflected the SRP concentration required to change the equilibrium P concentration by 1 mol/L (Froelich Citation1988). It was near 1000 L/kg or higher for all stations, suggesting that sediment can rapidly adsorb aqueous soluble P in the water column with little change in the EPC (i.e., well buffered with respect to equilibrium P).
Figure 4. An example of changes in the sediment exchangeable P (S) concentration as a function of the final (i.e., after 24 h of exposure) equilibrium soluble reactive phosphorus (SRP) concentration for sediment collected at Big Traverse West in the Lake of the Woods. Regression analysis between S and equilibrium SRP near the point of no net change in S was used to estimate the equilibrium P concentration (EPC).
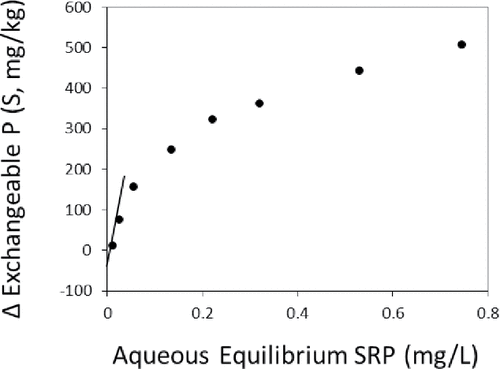
Table 4. Equilibrium phosphorus (P) characteristics, estimated P sorption capacity, and percent P saturation characteristics for surface sediment in the Lake of the Woods. EPC = equilibrium P concentration.
Patterns of P sorption tended to follow a Langmuir 2-surface layer model in which binding is regulated by P concentration in solution and by diffusion of P to interior sites on particles and polymers (). Assuming that the redox-P fraction approximated the sediment exchangeable P pool, the DPS ranged from ∼16% to 26% for the 2-surface layer model (i.e., 224 mg/kg ÷ 857 mg/kg for Big Traverse West). These ranges suggested that P saturation of binding sites on the sediments of the Lake of the Woods was currently modest ().
Phosphorus dynamics during simulated sediment resuspension
TSS concentrations were relatively low at shear stresses less than ∼1.5 dynes/cm2 and increased substantially above this value for all sediments (). Big Traverse East sediment exhibited the lowest critical shear stress at 1.3 dynes/cm2 while the other stations ranged from ∼ 2.1 to 2.3 dynes/cm2 (). Variations in sediment textural characteristics and particle size distribution did not explain the differences in critical shear stress. Perhaps differences in critical shear stresses were due to variations in particle sorting and fine-scale (mm scale) gradients in sediment properties at the sediment–water interface not captured from the composite 10 cm sediment section used for analysis. Critical shear stresses for LOW sediments also fell within ranges measured for some other lakes and impoundments that exhibit frequent sediment resuspension (). Critical shear stresses measured in this study were much higher than the literature values of ∼0.005 N/m2 for Big Traverse Bay used in Zhang et al. (Citation2013).
Figure 5. Changes in total suspended sediment (TSS, upper panels), soluble reactive phosphorus concentration (SRP, middle panels), and the net change in SRP from the initial concentration as a function of applied shear stress in the overlying water column. Arrows denote the estimated critical shear stress.
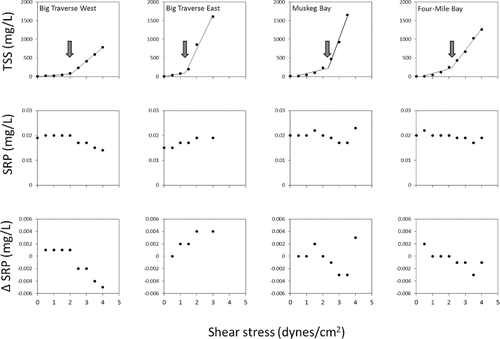
Table 5. Sediment critical shear stress of deposited sediment collected from various stations in the Lake of the Woods.
SRP concentrations in the overlying water column of sediment systems ranged between ∼0.015 and 0.020 mg/L before initiation of shear stress turbulence (). These initial concentrations were high relative to the potential for algal uptake and may be attributable to some aerobic diffusive P flux into the overlying water column during transport to the laboratory after collection. In addition, some of the SRP could have been derived from colloidal Fe3+ oxyhydroxide∼PO4 in the water column that passed through the 2 µm filter. If so, SRP would represent an overestimate of PO4 (Nürnberg Citation1984). The concentration generally declined as a function of increasing sediment entrainment for the Big Traverse West, Muskeg, and Four-Mile sediment systems, resulting in net declines in SRP concentration over initial values (). Notably, SRP concentrations increased slightly during resuspension in the Big Traverse East sediment system, and the net increase in SRP concentration was ∼0.004 mg/L at shear stresses of 2−3 dynes/cm2. SRP concentrations also increased at the greatest shear stress in the Muskeg and Four-Mile bay sediment systems. Overall, mean SRP concentrations during simulated resuspension, estimated over all shear stresses, were 0.017−0.020 mg/L with negligible net change in concentration from the initial 0.019 mg/L (). These general patterns corroborated findings from the P equilibrium experiments and water-extractable P determinations; namely, that the EPC achieved between resuspended sediment and the surrounding water was relatively low.
Table 6. Mean (1 standard error; n = 5 to 8) soluble reactive phosphorus (SRP) concentration and mean change in SRP during resuspension for various sediments. The mean initial SRP concentration (i.e., prior to sediment resuspension) was 0.019 mg/L). Plus and minus signs indicate a net increase (+) or decrease (−) in SRP versus the initial concentration.
Discussion
Summer diffusive P fluxes were relatively high under anaerobic conditions and at maximal simulated bottom temperatures, coinciding with moderately high concentrations of redox-P. In particular, the Big Traverse West mean anaerobic diffusive P flux of 12.5 mg/m2 y (0.26 SE) was comparable to those of highly eutrophic Lake Winnipeg (Loh et al. Citation2013; ) and nearly 2 times higher than anaerobic P release rates measured in hypereutrophic Nakamun Lake (Orihel et al. Citation2015). Thus, temporary stratification and bottom anoxia in bays of LOW could lead to considerable sediment diffusive P flux under anaerobic conditions.
By contrast, summer LOW aerobic diffusive P fluxes were more than 20 times lower. Sediment total Fe concentrations were high, falling within the upper range of concentrations reported in Barko and Smart (Citation1986), and the sediment Fe:P ratio exceeded 25:1. These patterns suggested that summer diffusive P flux from sediments in LOW may be coupled with Fe cycling. Under oxygenated conditions, Fe is in an oxidized state as an Fe-(OOH) and strongly adsorbs PO4, resulting in low summer diffusive P flux from sediments (Mortimer Citation1971). Fe+3 oxyhydroxide is reduced to Fe+2 in conjunction with anaerobic bacterial metabolism, resulting in desorption of PO4 and much higher summer rates of diffusive P flux, as observed for LOW sediments incubated under anaerobic conditions.
Hypothesized Fe–P dynamics in LOW sediments contrasted with other shallow polymictic lakes that exhibited low porewater Fe due to reaction with H2S to insoluble pyrite (Orihel et al. Citation2015). In Nakamun Lake (Alberta, Canada) sediments, for instance, porewater Fe was negligible and largely sequestered as FeSx, coinciding with relatively high anaerobic diffusive P flux into the bottom waters () and vertical entrainment into the euphotic zone for assimilation by cyanobacteria during periods of destratification. Orihel et al. (Citation2015) suggested that organic P breakdown, bacterial polyphosphate recycling, and lysis of aerobic bacteria under anoxic conditions may be involved in anaerobic diffusive P flux versus reduction of Fe-(OOH) because Fe-bound P species were below detection limits. High anaerobic diffusive P flux from LOW sediment needs to be linked with more information on sediment Fe2+ flux under anaerobic conditions, accumulation in the bottom waters during periods of temporary stratification and anoxia, and the hypolimnetic Fe:P ratio to better understand availability to cyanobacteria during mixing and entrainment events (James et al. Citation2015).
Because the LOW system is large and shallow, frequent wind-generated mixing may result in oxygenated conditions in the bottom waters and maintenance of an oxidized microzone at the sediment interface throughout much of the summer, driving diffusive P flux primarily under aerobic conditions. Because summer bottom temperatures can exceed 20 C for at least one month (i.e., during July; M. Edlund, pers. comm.) throughout LOW, an area-weighted July aerobic diffusive P flux of 0.35 mg/m2 d () would represent an input of ∼40 T in 1 month, or ∼15% of the annual internal P load mass (i.e., 271 T). Temporary bottom hypoxia and anoxia can potentially develop quickly during calm conditions in Big Traverse Bay (M. Edlund, pers. comm.), however, which could result in intermittent summer anaerobic diffusive P flux that would account for a greater portion of the internal P load.
An additional unknown is the impact of diffusive boundary layer thickness on aerobic and anaerobic P release from sediments in LOW. In particular, water movements generated by winds over the relatively expansive fetch of Big Traverse Bay could result in decreased diffusive boundary layer thickness at the sediment–water interface as bottom shear velocity increases (Nakamura and Stefan Citation1994, Hondzo et al. Citation2005). Diffusive efflux of P at the sediment–water interface would increase as the diffusive boundary layer thickness declines. Thus, laboratory-derived summer aerobic diffusive P fluxes determined in this study were probably conservative and could be higher in situ.
LOW surficial sediments were composed primarily of silts and clays and exhibited a moderately low critical shear stress, suggesting strong susceptibility to wind-generated resuspension. Indeed, Big Traverse, Muskeg, and Four-Mile bays were highly turbid during the September sampling period in conjunction with high wind conditions and wave activity. Equilibrium determinations indicated that sediments probably acted as a sink for PO4 and sequestered P during resuspension events when aqueous concentrations exceeded ∼0.01 mg/L as SRP. This finding was supported by several factors: the Fe concentration and the Fe:P ratio of the sediment were high, implying binding of PO4 onto Fe+3 oxyhydroxide complexes; P binding sites were undersaturated with respect to an estimated Smax; the kd was high, suggesting high buffering P capacity by resuspended sediment; the equilibrium P concentration was low; and SRP tended to decline or exhibit minor net change as a function of resuspended sediment concentrations and applied shear stress in laboratory sediment systems. These patterns suggested that periods of sediment resuspension in the LOW could control a relatively low aqueous soluble P concentration and limit soluble P availability for uptake by cyanobacteria. Sediment resuspension could also sequester and bind P previously released from sediment during periods when bottom shear velocities were below the critical shear stress.
The possible occurrence of colloidal Fe-(OOH)∼PO4 complexes during periods of sediment resuspension need to be explored to better quantify PO4 availability to cyanobacteria. Unexpectedly, background SRP in this study was detectable, suggesting that P may not limit cyanobacterial growth in LOW (); however, the actual bioavailability of SRP may be much lower if some of the molybdate-reactive P is associated with colloidal Fe-(OOH) (Haygarth et al. Citation1997, Filella et al. Citation2006, Effler and O'Donnell Citation2010, Baken et al. Citation2016a, Citation2016b), which may be the case for LOW. SRP is operationally defined as molybdate-reactive P passing through a 0.45 µm filter; however, colloidal Fe-(OOH)∼P can range between 1 and 1000 nm, pass through a 0.45 µm filter, and also react with molybdate, resulting in overestimation of ortho-P and leading to misinterpretation of SRP bioavailability. Because colloidal P can be less available for algal uptake (Baken et al. Citation2014), caution is needed when interpreting the bioavailability of SRP concentrations during resuspension events until research is conducted on colloidal Fe-(OOH)∼P in LOW.
Preliminary TP budgets for LOW have suggested that tributary P loadings do not entirely account for mean summer TP concentration, implying that additional internal P loading sources to the system may be important in sustaining cyanobacteria productivity. The present research provides findings and insights on internally derived P sources to the system via diffusive fluxes from bottom sediments and equilibrium fluxes from resuspended sediment. Although summer aerobic diffusive P flux from sediment is currently modest, P inputs from this source could be important in partially explaining the summer missing component of the P mass balance. Intermittent periods of anaerobic diffusive P flux during periods of temporary stratification may also contribute to the P budget (Zhang et al. Citation2013). In addition, climate change could exacerbate future diffusive P flux loading to LOW via extended and warmer ice-free seasons, elevated water temperature, and increased periods of temporary stratification and bottom water anoxia (Rühland et al. Citation2008, Citation2010).
A portion of the unaccounted for P, particularly in Big Traverse Bay, may also be due to frequent resuspension that contributes to high water column TP (Zhang et al. Citation2013) dominated by inorganic sediment P largely unavailable for uptake rather than P incorporated into algal cells. Resuspension events could compete with cyanobacterial uptake by sequestering available soluble P derived from diffusive P fluxes and watershed P loads. If so, predicted summer chlorophyll estimated from empirical models (i.e., Jones and Bachmann Citation1976, Nürnberg Citation1996) might overestimate observed summer means if a significant portion of the TP is resuspended inorganic P (Zhang et al. Citation2013). Pla et al. (Citation2005) found that September spatial trends in chlorophyll concentration were not correlated with TP and suggested that other factors such as nitrogen, light, mixed depth, or temperature might be controlling algal growth. Additionally, variation in the bioavailability of TP and the fraction that is resuspended inorganic versus incorporated into biomass in September need to be considered (Effler et al. Citation2014, Gelda et al. Citation2016). Zhang et al. (Citation2013) suggested that basin morphology, hydrology, circulation patterns, and turbulent mixing can also impact TP mass and algal biomass distribution and growth. Future modeling analysis and research is needed to quantify annual internal P loading that includes the entire LOW system and considers the biological availability of both tributary loads and internally derived P sources for algal uptake (i.e., soluble versus TP flux to the water column) as well other factors controlling cyanobacterial growth and distribution in LOW.
Acknowledgments
I gratefully acknowledge N. Baratono and C. Hernandez of the Minnesota Pollution Control Agency for providing field support for sediment core collection on very windy days in September. I also thank A. Carlson (Engineer Research and Development Center - Eau Galle Aquatic Ecology Laboratory) for analytical support and G. Nürnberg and other anonymous reviewers for comments that greatly improved this manuscript.
Funding
Funding was provided by the Minnesota Pollution Control Agency.
References
- [APHA] American Public Health Association. 2005. Standard methods for the examination of water and wastewater. 21st ed. Washington (DC).
- Baken S, Moens C, van der Grift B, Smolders E. 2016a. Phosphate binding by natural iron-rich colloids in streams. Water Res. 98:326–333.
- Baken S, Nawara S, Van Moorleghem C, Smolders E. 2014. Iron colloids reduce bioavailability of phosphorus in the green alga Raphidocelis subcapitata. Water Res. 59:198–206.
- Baken S, Regenlink IC, Comans RNJ, Smolders E, Koopmans GF. 2016b. Iron-rich colloids as carriers of phosphorus in streams: a field-flow fractionation study. Water Res. 99:83–90.
- Barko JW, Smart RM. 1986. Sediment-related mechanisms of growth limitation in submersed macrophytes. Ecology. 67:1328–1340.
- Bolster CH, Hornberger GM. 2007. On the use of linearized Langmuir equations. Soil Sci Soc Am J. 71:1796–1806.
- Bremner JM, Mulvaney CS. 1982. Nitrogen – total. Methods of soil analysis, part 2. Chemical and microbiological properties. Agronomy Monograph no 9, 2nd ed. Madison (WI): American Society of Agronomy, Soil Science Society of America.
- Caraco NF, Cole JJ, Likens GE. 1989. Evidence for sulphate-controlled phosphorus release from sediments of aquatic systems. Nature. 341:316–318.
- Carter LD, Dzialowski AR. 2012. Predicting sediment phosphorus release rates using landuse and water quality data. Freshwater Sci. 31:1214–1222.
- Chen H, Burke JM, Mosindy T, Fedorak PM, Prepas EE. 2009. Cyanobacteria and microcystin-LR in a complex lake system representing a range in trophic status: Lake of the Woods, Ontario, Canada. J Plankton Res. 31:993–1008.
- Cyr H, McCabe SK, Nürnberg GK. 2009. Phosphorus sorption experiments and the potential for internal phosphorus loading in littoral areas of a stratified lake. Water Res. 43:1654–1666.
- de Vicente I, Cruz-Pizarro L, Rueda FJ. 2010. Sediment resuspension in two adjacent coastal lakes: controlling factors and consequences on phosphate dynamics. Aquat Sci. 72:21–31.
- Doig LE, North RL, Hudson JJ, Hewlett C, Lindenschmidt KE, Liber K. 2017. Phosphorus release from sediments in a river-valley reservoir in the norther Great Plains of North America. Hydrobiologia. 787:323–339.
- Effler SW, O'Donnell SM. 2010. A long-term record of epilimnetic phosphorus patterns in recovering Onondaga Lake, New York. Fund Appl Limnol. 177:1–18.
- Effler SW, Prestigiacomo AR, Gelda R, Matthews DA. 2014. Partitioning the contributions of minerogenic particles and bioseston to particulate phosphorus and turbidity. Inland Waters. 4:179–192.
- Filella M, Deville C, Chanudet V, Vignati D. 2006. Variability of the colloidal molybdate reactive phosphorus concentrations in freshwaters. Water Res. 40:3185–3192.
- Froelich PN. 1988. Kinetic control of dissolved phosphate in natural rivers and estuaries: a primer on the phosphate buffer mechanism. Limnol Oceanogr. 33:649–668.
- Gächter R, Meyer JS, Mares A. 1988. Contribution of bacteria to release and fixation of phosphorus in lake sediments. Limnol Oceanogr. 33:1542–1558.
- Gächter R, Meyer JS. 1993. The role of microorganisms in mobilization and fixation of phosphorus in sediments. Hydrobiologia. 253:103–121.
- Gelda RK, Effler SW, Prestigiacomo AR, Peng F, Auer MT, Kuczynski A, Chapra SC. 2016. Simulation of the contribution of phosphorus-containing minerogenic particles to particulate phosphorus concentration in Cayuga Lake, New York. Water Air Soil Pollut. 227:365.
- Håkanson L, Jansson M. 2002. Principles of lake sedimentology. Caldwell (NJ): Blackburn Press.
- Hargan KE, Paterson AM, Dillon PJ. 2011. A total phosphorus budget for the Lake of the Woods and Rainy River catchment. J Great Lakes Res. 37:753–763.
- Haygarth PM, Warwick MS, House WA. 1997. Size distribution of colloidal molybdate reactive phosphorus in river waters and soil solution. Water Res. 31:439–448.
- Holmroos H, Niemiströ J, Weckström K, Horppila J. 2009. Seasonal variation of resuspension-mediated aerobic release of phosphorus. Boreal Environ Res. 14:937–946.
- Hondzo M, Feyaerts T, Donovan R, O'Connor BL. 2005. Universal scaling of dissolved oxygen distribution at the sediment-water interface: a power law. Limnol Oceanogr. 50:1667–1676.
- Hupfer M, Gloess S, Grossart HP. 2007. Polyphosphate-accumulating microorganisms in aquatic sediments. Aquat Microb Ecol. 47:299–311.
- Hupfer M, Glöss S, Schmieder P, Grossart HP. 2008. Methods for detection and quantification of polyphosphate and polyphosphate accumulating microorganisms in aquatic sediments. Int Rev Hydrobiol. 93:1–30.
- James WF. 2010. Nitrogen retention in a floodplain backwater of the upper Mississippi River (USA). Aquat Sci. 72:61–69.
- James WF, Barko JW. 1994. Macrophyte influences on sediment resuspension and export in a shallow impoundment. Lake Reserv Manage. 10:95–102.
- James WF, Barko JW, Butler MG. 2004a. Shear stress and sediment resuspension in relation to submersed macrophyte growth. Hydrobiologia. 515:181–191.
- James WF, Best EP, Barko JW. 2004b. Sediment resuspension and light attenuation in Peoria Lake: Can macrophytes improve water quality in this shallow system? Hydrobiologia. 515:193–201.
- James WF, Larson CE. 2008. Phosphorus dynamics and loading in the turbid Minnesota River (USA): controls and recycling potential. Biogeochemistry. 90:75–92.
- James WF, Sorge PW, Garrison PJ. 2015. Managing internal phosphorus loading and vertical entrainment in a weakly stratified eutrophic lake. Lake Reserv Manage. 31:292–305.
- Jirka AM, Carter MJ, May D, Fuller FD. 1976. Ultramicro semiautomated method for simultaneous determination of total phosphorus and total Kjeldahl nitrogen in waste waters. Environ Sci Technol. 10:1038–1044.
- Jones JR, Bachmann RW. 1976. Prediction of phosphorus and chlorophyll levels in lakes. J Water Pollut Cont Fed. 48:2176–2182.
- Kelderman P, Ang'weya RO, De Rozari P. 2012. Sediment characteristics and wind-induced sediment dynamics in shallow Lake Markermeer, the Netherlands. Aquat Sci. 74:301–313.
- Kleeberg A. 1998. The quantification of sulfate reduction in sulfate-rich freshwater lakes – a means for predicting the eutrophication process of acidic mining lakes? Water Air Soil Pollut. 108:365–374.
- Loh PS, Molot LA, Nürnberg GK, Watson SB, Ginn B. 2013. Evaluating relationships between sediment chemistry and anoxic phosphorus and iron release across three different water bodies. Inland Waters. 3:105–118.
- [LOW] Lake of the Woods, Water Sustainability Foundation. 2014. Overview. Rainy-Lake of the Woods state of the basin report; [cited 1 Jul 2014]. Available from: www.lowwsf.com/sobr-download.
- Matisoff G, Carson M. 2014. Sediment resuspension in the Lake Erie nearshore. J Great Lakes Res. 40:532–540.
- Matisoff G, Kaltenberg EM, Steely RL, Hummel SK, Seo J, Gibbons K, Bridgeman TB, Seo Y, Behbahana M, James WF, et al. 2016. Internal loading of phosphorus in western Lake Erie. J Great Lakes Res. 42:775–788.
- Matisoff G, Watson SB, Guo J, Duewiger A, Steely R. 2017. Sediment and nutrient distribution and resuspension in Lake Winnipeg. Sci Tot Environ. 575:173–186.
- Mortimer CH. 1971. Chemical exchanges between sediments and water in the Great Lakes – speculations on probable regulatory mechanisms. Limnol Oceanogr. 16:387–404.
- Nakamura Y, Stefan HG. 1994. Effect of flow velocity on sediment oxygen demand: theory. J Env Eng. 120:996–1016.
- Nürnberg GK. 1984. Iron and hydrogen sulfide interference in the analysis of soluble reactive phosphorus in anoxic waters. Water Res. 18:369–377.
- Nürnberg GK. 1988. Prediction of phosphorus release rates from total and reductant-soluble phosphorus in anoxic lake sediments. Can J Fish Aquat Sci. 45:453–462.
- Nürnberg GK. 1995. Quantifying anoxia in lakes. Limnol Oceanogr. 40:1100–1111
- Nürnberg GK. 1996. Trophic state of clear and colored, soft- and hardwater lakes with special consideration of nutrients, anoxia, phytoplankton and fish. Lake Reserv Manage. 12:432–447.
- Nürnberg GK. 1998. Prediction of annual and seasonal phosphorus concentrations in stratified and polymictic lakes. Limnol Oceanogr. 43:1544–1552.
- Nürnberg GK. 2005. Quantification of internal phosphorus loading in polymictic lakes. Verh Int Ver Limnol 29:623–626.
- Nürnberg GK. 2009. Assessing internal phosphorus load – problems to be solved. Lake Reserv Manage. 25:419–432.
- Nürnberg GK, LaZerte BD. 2016. More than 20 years of estimated internal phosphorus loading in polymictic, eutrophic Lake Winnipeg, Manitoba. J Great Lakes Res. 42:18–27.
- Nürnberg GK, LaZerte BD, Loh PS, Molot LA. 2013a. Quantification of internal phosphorus load in large, partially polymictic and mesotrophic Lake Simcoe, Ontario. J Great Lakes Res. 39:271–279.
- Nürnberg GK, Molot LA, O'Connor E, Jarjanazi H, Winter JG, Young JD. 2013b. Evidence for internal phosphorus loading, hypoxia and effects on phytoplankton in partially polymictic Lake Simcoe, Ontario. J Great Lakes Res. 39:259–270.
- Nürnberg GK, Tarvainen M, Ventelä A-M, Sarvala J. 2012. Internal phosphorus load estimation during biomanipulation in a large polymictic and mesotrophic lake. Inland Waters. 2:147–162.
- Orihel DM, Schindler DW, Ballard NC, Graham MD, O'Connell DW, Wilson LR, Vinebrooke RD. 2015. The ‘nutrient pump:’ iron-poor sediments fuel low nitrogen-to-phosphorus ratios and cyanobacterial blooms in polymictic lakes. Limnol Oceanogr. 60:856–871.
- Pla S, Paterson AM, Smol JP, Clark BJ, Ingram R. 2005. Spatial variability in water quality and surface sediment diatom assemblages in a complex lake basin: Lake of the Woods, Ontario, Canada. J Great Lakes Res. 31:253–266.
- Plumb RH. 1981. Procedures for handling and chemical analysis of sediment and water samples. Vicksburg (MS): US Army Engineer Waterways Experiment Station, Technical Report EPA/CE-81-1.
- Psenner R, Puckso R. 1988. Phosphorus fractionation: advantages and limits of the method for the study of sediment P origins and interactions. Arch Hydrobiol Biel Erg Limnol. 30:43–59.
- Reitzel K, Ahlgren J, DeBrabandere H, Waldenbäck M, Gogoll A, Tranvik L, Rydin E. 2007. Degradation rates of organic phosphorus in lake sediment. Biogeochemistry. 82:15–28.
- Roden EE, Edmonds JW. 1997. Phosphate mobilization in iron-rich anaerobic sediments: microbial Fe(III) oxide reduction versus iron sulfide formation. Arch Hydrobiol. 139:347–378.
- Rühland K, Paterson AM, Smol JP. 2008. Hemispheric-scale patterns of climate-related shifts in planktonic diatoms from North American and European Lakes. Glob Change Biol. 14:2740–2754.
- Rühland KM, Paterson AM, Hargan K, Jenkin A, Clark BJ, Smol JP. 2010. Reorganization of algal communities in the Lake of the Woods (Ontario, Canada) in response to turn-of-the-century damming and recent warming. Limnol Oceanogr. 55:2433–2451.
- Søndergaard M, Jensen JP, Jeppesen E. 2001. Retention and internal loading of phosphorus in shallow, eutrophic lakes. Sci World. 1:427–442.
- Søndergaard M, Jensen JP, Jeppsen E. 2003. Role of sediment and internal loading of phosphorus in shallow lakes. Hydrobiologia. 506–509:135–145.
- Søndergaard M, Kristensen P, Jeppesen E. 1992. Phosphorus release from resuspended sediment in the shallow and wind-exposed Lake Arresø, Denmark. Hydrobiologia. 228:91–99.
- [SERA-IEG] Southern Extension/Research Activity – Information Exchange Group 17. 2000. Methods of phosphorus analysis for soils, sediments, residuals, and waters. Southern Cooperative Series Bulletin No. 396. Available from: http://www.soil.ncsu.edu/sera17/publications/sera17-2/pn_cover.htm.
- [SAS] Statistical Analysis System. 1994. SAS/STAT Users Guide. Version 6. 4th ed. Cary (NC): SAS institute.
- Tsai CH, Lick W. 1986. A portable device for measuring sediment resuspension. J Great Lakes Res. 12:314–321.
- Zhang W, Watson SB, Rao YR, Kling HJ. 2013. A linked hydrodynamic, water quality and algal biomass model for a large, multi-basin lake: A working management tool. Ecol Model. 269:37–50.
