
ABSTRACT
Watson SB and Kling H. 2017. Lake of the Woods phyto- and picoplankton: spatiotemporal patterns in blooms, community composition, and nutrient status. Lake Reserve Manage. 33:415–432.
This study evaluated the phytoplankton and trophic status in Lake of the Woods (LOW; Ontario, Canada), a large, multi-basin waterbody with marked gradients in water quality and annual cyanobacterial blooms. We combined a broad comparison of average total phosphorus (TP)–biomass relationships in LOW and other north temperate lakes with a detailed spatiotemporal analysis of net-, nanno-, and picoplankton, total bacterioplankton, nutrient status, and water quality across six hydrological sectors of the lake between 2008 and 2010. Average total phytoplankton biomass (TB) varied considerably among zones and years but, similar to TP, was highest in the shallower, more eutrophic southern zones with a generally lower than average TB/TP yield within the among-lakes dataset. Summer–fall blooms of N2-fixing cyanobacteria were dominated by Aphanizomenon across most sectors except in the north, where Dolichospermum predominated. Unlike the larger size fractions, patterns in picoplankton suggested non-nutrient constraints: phycoerythrin-rich picocyanobacteria (<2 µm) showed a distinct south–north increase correlated with water transparency, while the more abundant phycocyanin-rich fraction was related to dissolved organic material with no distinct spatial patterns. Bacterial numbers were higher in surface layers and significantly related to temperature but not nutrients; spatial distribution indicated they were not directly introduced from riverine inputs. Overall, planktonic P deficiency was higher in the north and increased in late summer concurrent with N co-limitation and increases in N2-fixers. The combined data suggest that P ultimately limits LOW plankton, but short-term N and Si deficiencies influence the plankton community composition.
Recent studies of Lake of the Woods (LOW) phytoplankton have focused on cyanobacteria and the annual appearance of dense, mid–late summer blooms of these taxa (Chen et al. Citation2007, Chen et al. Citation2009, DeSellas et al. Citation2009). Harmful cyanobacterial blooms (cHABs) can produce toxins that seriously affect humans and other organisms, and have multiple other adverse impacts on drinking water supplies, beaches and shorelines, recreational activities, property value, fisheries, and tourist, hospitality, and other industries (e.g., Watson and Molot Citation2013, Clapper and Caudill Citation2014). The ecological impacts of cHABs are pervasive, and affect water quality, transparency, and dissolved oxygen, and may result in harmful or lethal effects on fish, waterfowl and other food web levels.
Public awareness and concern with cHABs in LOW has increased over the past decade, although it is not clear whether these events have in fact become more frequent or severe. Sporadic reports of algal blooms date back to the 1800s, when the water was described as “shoal and foul, frequently with a green scum” (Joseph Delafield, 30 Jul 1823), and in summer 1857 by S.J. Dawson as “covered with a sort of green scum or vegetable matter which thickens as we proceed” (crossing Grand Traverse Bay from the Rainy River; Canada Provincial Secretary's Office Citation1859). On the other hand, the first published report on the LOW algal community described the water as “unusually clear and free from contamination by drainage or sewage,” with an algal community typical of large oligotrophic Shield lakes (Lowe Citation1924). Nevertheless, by the early 1990s, the southern portion of the lake was characterized as “eutrophic,” based on Carlson indices of chlorophyll a (Chl-a), total phosphorus (TP), and Secchi disc depth (MPCA Citation2001). Modeling and remote sensing suggest that the blooms have not increased over the past 10 years (Binding et al. Citation2011a, Zhang et al. Citation2013) and, furthermore, that they are controlled by a number of factors in addition to nutrients. In fact, the total phosphorus (TP) load from the Rainy River decreased by ∼66% since 1970 as a result of reduced inputs from domestic and pulp mill wastewater (Hutchinson Environmental Sciences Citation2011). In-lake TP concentrations also declined, but dense and often spatially extensive blooms continue to occur across the lake, attributed by some authors to the increased length of the ice-out period and associated changes in mixing and circulation (DeSellas et al. Citation2009). Paleolimnological analyses have related long-term changes in diatoms, pigments, and other fossils from sediment core profiles to climate change and hydrological engineering (damming; Rühland et al. Citation2010, Hyatt et al. Citation2011, Reavie et al. Citation2017). Overall, however, the sources, bioavailability and in-lake transport/sequestration of nutrients—and their influence on the plankton communities—are still not well understood.
Initially, LOW studies focused on the blooms in the southern basins, where high densities of cyanobacteria were attributed to nutrient inputs from the Rainy River. Anderson et al. (Citation2006) reported a mixed community of diatoms and cyanobacteria in the earlier part of 2005, which shifted to a predominance of cyanobacteria (numerically) dominated by “Anacystis” and Aphanizomenon. Chen et al. (Citation2007, Citation2009) conducted broader surveys of the dominant cyanobacteria and water quality, and described an assemblage which varied in composition and biomass across basins and seasons. It has been generally held that mid–late summer cyanobacterial blooms develop in the south and move northward, ostensibly via wind, currents, and circulation patterns (e.g., DeSellas et al. Citation2009) although there is no direct evidence that the northern blooms are not locally initiated. Furthermore, there is little information on the total phytoplankton community composition and nutrient status.
To address these gaps we undertook a detailed study of the plankton biomass and species composition across LOW, including the smallest (<2 µm) components of the planktonic food web (bacteria, autotrophic picoplankton). At the same time, we measured planktonic nutrient deficiency (stoichiometry of particulate organic C, N, and P [POC, PON, and PP]; N-debt) and a suite of water quality parameters (Pascoe et al. Citation2014). We also addressed the issue of sampling strategy: over the past decade, monitoring in LOW has been carried out by the Ontario Ministry of the Environment and Climate Change and the Minnesota Pollution Control Agency using different collection protocols, mixed layer integrated and discrete (1 m) depth samples. The compatibility of these data has not been considered, and to examine this issue, we evaluated differences between surface and integrated samples for water quality parameters including nutrients and Chl-a. A comparison was also made between surface and integrated samples from a late-summer bloom for biomass and species composition.
Study site
LOW is a large, multi-basin waterbody spanning a mixed geology of northern Canadian Shield igneous bedrock and southern glacio-lacustrine sediment. The lake is spatially complex, with numerous embayments, inlets, and islands, an extensive shoreline (105,000 km) and a high catchment-to-lake-area ratio (8.2:1; DeSellas et al. Citation2009). Based on patterns in water exchange and nutrients, Zhang et al. (Citation2013) divided LOW into six segments to develop a linked hydrodynamic/nutrient/biomass model, and this same zonation was applied in the current study (). The large, shallower, southern segments (zones 1, 2) are well-mixed, with a high degree of wind-induced turbulence, while the smaller, deeper, northern segments (zones 3–6) are thermally stratified during the summer.
Figure 1. Map of Lake of the Woods showing major hydrologic zones (1–6) and sampling sites, 2008–2010.
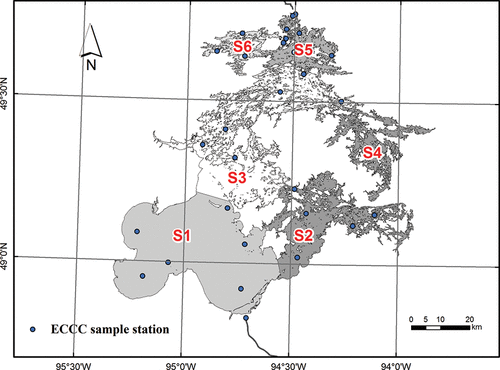
A south–north gradient in water chemistry reflects differences in the geology, vegetation, and development of the drainage basin and limited exchange among basins (Pla et al. Citation2005). Water flows in a general south-to-north direction from the south basin (zone 1; mean, maximum depth [Zm, Zmax] of 7.2 m, 11.6 m, respectively), which receives inflow via the Rainy River from a glacial till catchment, into the deeper interconnected northern basins with soft-water boreal Shield drainage. The northern segments vary considerably in size, shape, and depth (Zm 3.7–8.2 m; Zmax 9.6–40.4 m; Zhang et al. Citation2013; ). The outflow at Kenora is regulated by two dams, which influence water levels, flushing, and mixing patterns. Inputs from the Rainy River account for ∼75% of the estimated external TP loading (Hargan et al. Citation2011), over ∼50% of which is in the dissolved fraction and thus readily available for algal growth. Nutrients show a strong south–north gradient but significant inter-annual and spatial variance (Rühland et al. Citation2010, Zhang et al. Citation2013). Up to 50% of P inputs may be retained in the sediments as a potential source of internal loading, particularly in deep basins that develop hypolimnetic anoxia (e.g., Hargan Citation2010, Pascoe et al. Citation2014). Shoreline wastewater inputs (e.g., septic systems, gray-water pits) are as yet unquantified, but may account for up to ∼30–40% of the P loading to some of the deeper embayments (Hargan et al. Citation2011).
Methods
LOW phytoplankton trophic status and temporal/spatial patterns were evaluated using archived and newly collected samples from multiple years and sites from past and present studies carried out by different agencies. We evaluated the TP–biomass relationship in LOW by comparing it to summer mean euphotic zone concentrations of TP, Chl-a, and biomass from a previously published dataset of over 90 North temperate lakes (Watson et al. Citation1997, Citation2008; Downing et al. Citation2001). Data for LOW was combined from this study, unpublished data from Hedy Kling, and published LOW data from other studies (Chen et al. Citation2007, Citation2009).
In 2008–2010, open-water samples were collected from multiple sites 2–3 times per year (spring, summer, fall) as part of a larger LOW science initiative (e.g., Pascoe et al. Citation2014). At each site, depth profiles of temperature, dissolved oxygen (DO), pH, and conductivity were measured using a YSI profiler. Water samples were taken using a Van Dorn or Niskin water sampler at subsurface (1 m) and deep (2 m above the bottom) strata, along with an integrated mixed-layer sample collected from the surface to the top of the thermocline (where the site was stratified), or to just above the sediments (where the water column was isothermal). Samples were collected into acid-washed 4 L polyethylene bottles and kept cool and dark until processed later that day onshore for shipping and analysis for major nutrients (C, N, P), major ions, and Chl-a at the National Laboratory for Environmental Testing (NLET) using standard methods (Environment Canada Citation1997).
Aliquots of 100 mL from the 1-m samples were preserved with Lugol's iodine solution for later microscope analysis of algal biomass and taxonomic identification using the Utermöhl technique (Findlay and Kling Citation1998). Lugol's samples were collected from each field survey during the study (with the exception of June 2008); not all archived samples have been analyzed and here we report the data from 2010. As the primary focus has been on the late-summer blooms, our analyses concentrated on this time period, with a smaller number of samples enumerated from the spring period. Samples were pooled by zone and analyzed for major taxonomic groups and species. In addition, a comparison was made between surface and mixed-depth integrated samples collected from station 710 (zone 1) in late summer 2010.
For picoplankton, 1-mL subsamples from 1 m, depth-integrated, and bottom samples were added to paraformaldehyde-spiked 1.5-mL cryovials (final concentration of 4%), dark fixed for 8 h at 4 C and then flash frozen at −80 C until analysis. Blanks (membrane-filtered distilled water) and duplicates were collected for quality control. Samples were analyzed by flow cytometry at the J.J. MacIsaac Aquatic Cytometry Facility at the Bigelow Laboratory for Ocean Sciences using an adapted method from Vaulot et al. (Citation1989). Cells were enumerated based on size and fluorescence and grouped according to PE- and PC-rich Synechococcus (PE, PC) and pico-eukaryotes (Pk). Bacteria were enumerated from subsamples stained with the DNA stain PicoGreen (Invitrogen). Abundances of the different fractions were analyzed against the major physical, chemical, and nutrient parameters, together with the nutrient deficiency metrics. To accommodate unequal numbers, a general linear models procedure was used (SAS) together with multivariate regression analyses with backwards selection (α = 0.05). Data were log transformed to normalize the variance for statistical analysis.
Planktonic nutrient deficiency was evaluated on samples collected between 2008 and 2010 using sestonic stoichiometric ratios (C:N:P) and N-debt assays measuring NH4 uptake over 24 h dark-incubation (Healey and Hendzel Citation1980). Molar ratios of C:P, N:P, and C:N were compared against levels indicative of moderate deficiency as defined by Healey and Hendzel (Citation1980). POC/Chl-a and Chl-a/PP were also evaluated as indices of “general deficiency,” which may reflect, for example, silica, light iron, or other limiting resources. Data were analyzed using SAS multivariate analyses, using log transformation to normalize the variance.
Results and discussion
Among-lakes comparison
Chl-a shows a strong positive relationship to TP in most P-limited lakes (Watson et al. Citation1997 and many others) and has been used as a relatively simple measure of the phytoplankton community standing stock in LOW over the past decade. Among-lakes comparisons can provide initial insight into the general status of individual waterbodies along a trophic gradient, and a comparison of TP–Chl-a data from all sectors in LOW with those from a range of temperate zone lakes across the Northern Hemisphere () highlights the wide spatiotemporal variance in Chl-a and TP levels in LOW, and the significant range of Chl-a over almost 2 orders of magnitude within a relatively small range of TP. Multivariate analysis of the 2008–2010 data collected during this study (1-m samples) showed that Chl-a can be predicted by a nonlinear second-order model, both for LOW as a whole and for each of the zones with the exception of zone 6, which followed a linear response (). Although a practical and widely used metric, Chl-a is a community-level measure that provides no insight into the underlying taxa, and in fact may not correlate well with biomass, depending on the taxa and environmental conditions (e.g., LaBaugh Citation1995, Felip and Catalan Citation2000). For example, Chl-a shows a reasonable correlation with algal biomass when cyanobacteria and Chlorophyta (green algae) are dominant, less so with other algal groups, diatoms, and chrysophytes or under nutrient and light stress (EI-Shaarawi et al. Citation1978, Kasprzak et al. Citation2008, Garmendia et al. Citation2013).
Figure 2. Top left: Seasonal (Jun–Sep) average total phosphorus (TP) vs. chlorophyll (chl) plotted for north temperate lakes dataset (NTLk) and collective data from Lake of the Woods (LOW); top right: LOW data plotted on a smaller scale to show relationship within this lake. Bottom left: Seasonal (Jun–Sep) average TP vs. total algal biomass and (bottom right) total cyanobacteria biomass, plotted for large among-lakes dataset (Watson et al. Citation1997, light blue symbols) and LOW, using collective data from this and other LOW studies (see text).
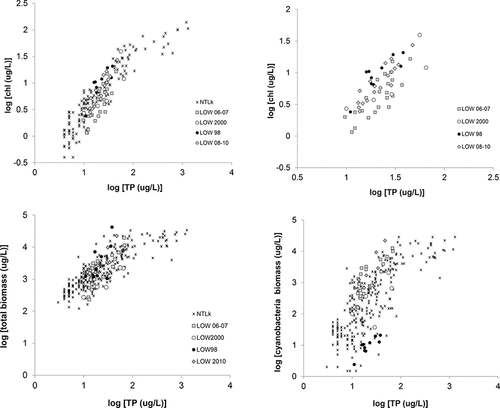
Table 1. Summary results of multivariate analysis for nonlinear models of (log) Chl-a vs. log total phosphorus (LTP) across each LOW zone using backward selection (α = 0.05); only significant relationships reported.
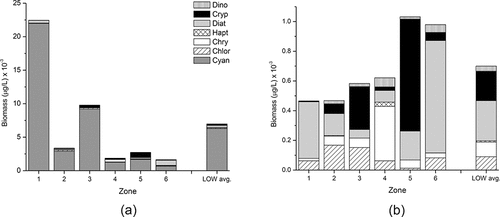
Similarly, over a broad range of lakes, total algal biomass (TB) also follows a nonlinear relationship with TP but shows considerably more scatter, some of which is derived from the error typically associated with enumeration, differences in methods, and computation of biovolume (Watson et al. Citation1997, Citation2008). LOW phytoplankton biomass data from the present study were combined with those from previous studies for the years 1998, 2001, 2004, 2006, and 2007. Seasonal (Jun–Sep) average TB fell within the mid–low range of the broader among-lakes empirical dataset from Watson et al. (Citation1997), suggesting a generally lower average biomass/TP yield which, similar to Chl-a, varied considerably among years (). Although the LOW data cover a smaller TP range than the larger lakes dataset, TB was significantly correlated with Chl-a and TP both on linear and log-transformed scales (P < 0.0001). However, there was no correlation between TB and total dissolved P (TDP), dissolved inorganic N (DIN; i.e. NH4 + NO3), or surface temperature within this dataset (P > 0.05; data not shown). We note that although the LOW data were represented by seasonal average values (by zones) they included both discrete (1-m; this study) and euphotic zone integrated samples (Chen et al. Citation2009). This may have contributed to the apparent lack of a systematic trend in average TB over the period examined (1998–2010; ), although it is consistent with the absence of any sustained patterns in long term Chl-a data and satellite imagery (DeSellas et al. Citation2009, Binding Citation2011a). The among-lakes data also incorporated a variety of sampling methods, and such issues are often unavoidable in broad empirical studies. Grouping the data collected during 1998–2010 into zones showed that, as might be expected, average TB was higher and more variable in the southern sectors (; ) and decreased in a northern direction, although zones 5 and 6 showed significant outliers in 1998 in Big Stone and LilyPad Bays, with total biomass reaching 19,120 µg/L and 19,880 µg/L, respectively.
Figure 3. Box and whisker plot of total phytoplankton biomass in LOW from subsurface samples (1 m) at the six in-lake EC zones. Data from 1998 to 2010 (see text).
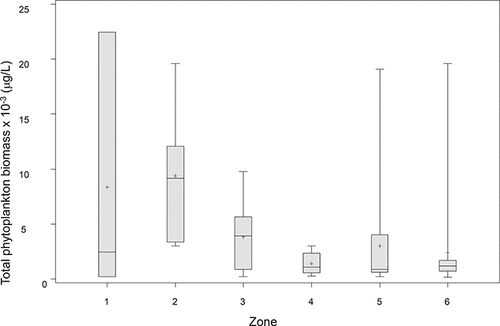
Table 2. Summary statistics of biomass for total phytoplankton (TB), cyanobacteria (CYAN), N-fixers (Nfix), Dolichospermum spp. (Dol), Aphanizomenon spp. (Aph), and levels of total microcystin toxins (MC) in LOW, 1998–2010; all units in µg/L.
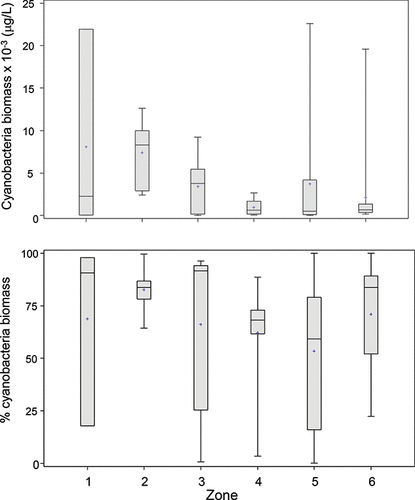
The relative contribution of the major taxa to the total community biomass has implications for both ecosystem health and water quality impairment. Empirical among-lake relationships have been described between seasonal average biomass of some of the major taxonomic groups and nutrients (e.g., Watson et al. Citation1997, Citation2008; Downing et al. Citation2001); of these, cyanobacteria show the strongest increase with TP. Within the TP range typically seen in LOW surface waters (e.g. ∼7–100 µg/L, measured in our surveys between 2008 and 2010), the larger among-lakes dataset varied in average cyanobacteria biomass from <200 to 10,000 µg/L, and LOW cyanobacteria biomass generally fell within the upper range of this variance (). This suggests a higher than average vulnerability to cyanobacterial dominance, although it should be noted that the majority of the data are from summer–fall period (Aug–Sep), when cyanobacteria comprised >50% of the total biomass in most sectors. Similar to total biomass, cyanobacteria biomass showed high variance in zones 1 and 2, and also in zones 5 and 6 ().
Figure 4. Cyanobacteria biomass (top) and percent contribution to total phytoplankton biomass (bottom) from subsurface samples (1 m) averaged across sites within in each of the six LOW hydrologic zones, 1998–2010.
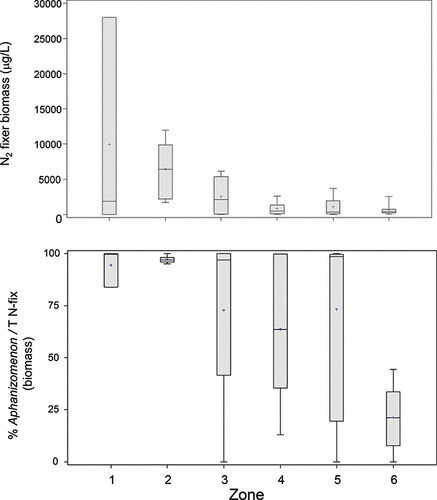
Earlier studies have described cyanobacterial assemblages in LOW dominated by N2 fixers, notably species of Aphanizomenon and Dolichospermum, with non-fixers playing a minor role in most sectors of the lake (e.g., Chen et al. Citation2007, Citation2009; Kling unpubl.). Consistent with this, broad patterns in N2-fixing cyanobacteria biomass across the zones mirror those of total cyanobacteria (e.g., Chen et al. Citation2009, this study; ). N2-fixing communities in the southern sectors (zones 1, 2, and 3) are overwhelmingly dominated by Aphanizomenon; for example, in samples collected during the present study, these taxa accounted for an average of 70–90% of the total N2-fixing biomass (). Raft-forming Aphanizomenon are common in blooms in other shallow eutrophic lakes, notably Lake Winnipeg and many prairie lakes (McLachlan et al. Citation1963, Mitchell and Prepas Citation1990, Kling et al. Citation2011). The biomass and relative importance of Aphanizomenon in the summer cyanobacteria assemblages across LOW is reduced in some of the northern basins, where Dolichospermum spp. can account for an equivalent, or at times greater, proportion of the N2-fixers, as also observed by Chen et al. (Citation2009).
Figure 5. Left: biomass of the two dominant N2-fixing cyanobacteria taxa (Dolichospermum and Aphanizomenon) and percent contribution of Aphanizomenon to N2-fixing biomass averaged from sites within the six zones in LOW.
Spatial and seasonal differences in major phytoplankton species, 2010
In 2010, late summer (Sep) samples from all zones were predominated by cyanobacteria, except in the most remote northern zone 6 (). As seen by earlier studies (Anderson et al. 2006; Chen et al. Citation2007, Citation2009), southern sectors were overwhelmingly dominated by the Aphanizomenon flos-aquae species complex. These species of large, buoyancy-regulating cyanobacteria are not reported to produce any of the known toxins, and form large rafts of stacked filaments, which can accumulate in thick surface scums during calm conditions visible by remote sensing (e.g., Binding et al. Citation2011a, Citationb). As a result, surface samples can show significantly higher biomass than depth-integrated samples (below). Aphanizomenon accounted for 97% of the total biomass at zone 1 (Big Traverse, Muskeg, and Buffalo Bays) where cyanobacteria were the overwhelmingly dominant group (98% of the total biomass). In contrast, Dolichospermum (D. flos-aquae and D. spiroides) accounted for less than 1% of the total cyanobacteria biomass in this sector. Other cyanobacteria were also present but at very low abundance (Pseudanabaena sp., Coelosphaerium kuetzingianum, Microcystis ichthyoblabe, Woronichinia eloranta, Romeria sp., and Synechococcus spp.).
Figure 6. Late-summer biomass of major taxonomic groups in samples from LOW at all LOW zones, Sep 2010. Top: all taxa; bottom: same plot with cyanobacteria removed.
Zone 2 samples (Hay and Bigsby Islands, Basil Point) showed a much lower total biomass, which was dominated by a more diverse cyanobacteria assemblage (86% of the total phytoplankton biomass). The dominant taxa again included members of the Aphanizomenon flos-aquae complex (50% TB) and a few filaments of Cuspidothrix issatschenkoi. Strains of C. issatschenkoi have been reported to produce the neurotoxin anatoxin-a (Wood et al. Citation2007, Ballot et al. Citation2010), but this species was not observed to exceed 1% of the total biomass in any LOW samples. Dolichospermum was not observed, but several buoyancy-regulating colonial chlorococcalean cyanobacteria were important, notably species of Microcystis (650 µg/L or 19% TB), which included M. flos-aquae, M. ichthyoblabe, and M aeruginosa, all of which are widely known as potential sources of microcystin (MC), and M. wesenbergii, which to date is not a reported toxin producer. Woronichinia species (W. naeglianum, W. cf. eloranta) were also relatively abundant (∼200 µg/L or 7% total phytoplankton biomass).
Compared to zone 2, both total and cyanobacteria biomass in zone 3 (Mica Point, Monkey Rocks, and sites near East Allie and Donald Duck Islands) were significantly higher (9205 µg/L, 9788 µg/L, respectively). The community, again largely dominated by cyanobacteria (94% TB), showed a higher relative contribution of potentially toxic species and smaller proportion of Aphanizomenon (62% of the cyanobacteria biomass). Cuspidothrix issatschenkoi was not observed, but there was a higher abundance of Microcystis (∼3200 µg/L), representing 38% of the total cyanobacteria biomass and largely dominated by M. aeruginosa (3110 µg/L). Woronichinia and Dolichospermum were present at low levels (12 µg/L, 3 µg/L, respectively) together with other cyanobacteria (Pseudanabaena rutilis-viridis, Radiocystis geminata, Cyanodictyon reticulatum, and Synechococcus sp.).
Water transparency was significantly higher in the northern embayments (Pascoe et al. Citation2014) but there were distinct differences among these sectors in total phytoplankton biomass and composition. Zone 4 (Yellow Girl and Whitefish Bays) showed a markedly lower and more diverse cyanobacteria biomass (1245 µg/L or 67% TB) than southern sectors, with much less Aphanizomenon (230 µg/L or 18% TB) and more Dolichospermum, largely present as D. planctonicum (420 µg/L or 23% TB). D. planctonicum filaments do not form colonies, and this species has been reported elsewhere as a potential source of anatoxin-a (Dadheech et al. Citation2014). Microcystis was not observed but there was a measureable abundance of other non-fixers, notably small cyanobacteria filaments (e.g., Limnothrix and Pseudanabaena) and unicellular and colonial picocyanobacteria (e.g., Aphanothece sp., Aphanocapsa sp., Cyanodictyon planktonica, Lemmermaniella pallida, and unidentified unicells).
Total algal biomass in zone 5 (Bigstone, Rat Portage, Kenora, and Safety Bays) was similar in magnitude and composition to zone 2 (2720 µg/L and 3368 µg/L, respectively) and overwhelmingly dominated by the Aphanizomenon flos-aquae complex (1400 µg/L or 83% total cyanobacteria biomass). Dolichospermum species (D. flos-aquae, D. mendotae, and D. planctonicum) were present as minor constituents, accounting for less than 1% of the cyanobacteria biomass. Microcystis was similarly low in abundance, but present as more compact forms (M. novacekii, M. botrys, M. viridis) compared to the large colonies of M. flos-aquae or diffuse and/or clathrate colonies of M. aeruginosa seen in the south (all of which species are reported elsewhere as potentially toxic; Watanabe et al. Citation1995, Falconer Citation2005, Sant'anna et al. Citation2008). The most northwestern sector, Clearwater Bay (zone 6), showed the lowest total biomass of all sectors (1650 µg/L; ) and a lower absolute and relative biomass of cyanobacteria (670 µg/L or 40% of the total phytoplankton biomass). The cyanobacteria assemblage more closely resembled that observed at zone 4—also a more isolated sector—with a higher representation of Dolichospermum, notably individual trichomes of D. macrospora, which, to date is not a reported toxin producer (e.g., Gugger et al. Citation2002). Aphanizomenon (mainly Aphanizomenon flos-aquae but also trichomes of A. gracile, which has been reported elsewhere to produce paralytic shellfish toxins; Ballot et al. Citation2010) and Microcystis (M. ichthyoblabe, M. wesenbergii) were relatively low in biomass (24% and 6% of the cyanobacteria biomass, respectively). Other chroococcales, such as Woronichinia, and small non-fixing filaments (Pseudanabaena, Limnothrix) were of minor importance. Nevertheless, combined 1998–2010 data show considerable variance in biomass in this zone, with median values of both total and cyanobacteria biomass higher in this sector than zone 5 ().
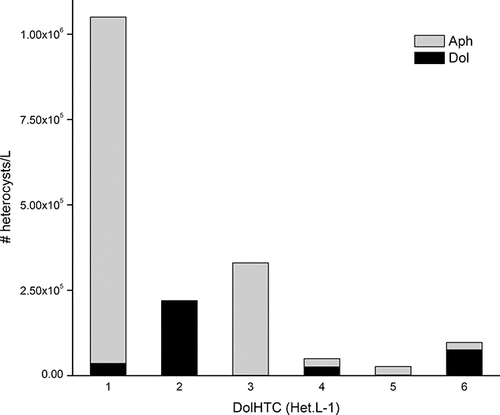
The combined evidence from TP–biomass analyses and nutrient deficiency measures (below) from this and other studies suggests that while the LOW algal biomass is primarily limited by P, N has a significant influence on community composition. Most notably (as discussed above), N2-fixing cyanobacteria predominate summer and fall cyanobacteria blooms, and in this study, heterocysts (HTCs) were mostly associated with Aphanizomenon across all sectors except zone 4, where HTC numbers were approximately equivalent for Aphanizomenon and Dolichospermum (). Heterocyst frequency (i.e., the proportion of heterocysts vs. vegetative cells in a population) was also higher in these two zones, and generally far greater in Aphanizomenon than Dolichospermum (). HTC was strongly correlated with both cyanobacteria biomass and PON, and showed a generally negative correlation with N-deficiency as measured by N-debt (Fig. S-1; supplementary data).
Figure 7. Abundance of heterocysts associated with the two major N2-fixing cyanobacteria taxa in LOW, Dolichospermum (gray series) and Aphanizomenon (black series), across the six hydrologic zones, Sep 2010.
Cyanobacteria toxins in LOW are reported in detail in Zastepa et al. (Citation2017) and discussed only briefly here for the samples collected in September 2010. As in other years, whole-water total microcystin (MC) toxins were consistently low across the lake, ranging from below detection to a maximum of 0.7 µg MC/L measured from a bloom at station 760 (zone 1) predominated by Aphanizomenon and trace amounts of Microcystis ichthyoblabe. This same sample contained Chl-a levels of 126 µg/L showing that, although dense, the bloom was extremely low in toxicity. Average toxin levels were slightly higher in this zone and, to a lesser extent, zone 5, where low numbers of Microcystis novacekii/M. botrys and M. viridis were also present. It is likely that there are several sources of toxins in LOW, which are present at low abundance and vary among seasons and basins. Toxigenic species may include the more likely candidates such as Microcystis and Dolichospermum, and less obvious sources such as Pseudanabaena rutilus-viridis, which we recently described from LOW and several other lakes (including Lake Winnipeg; Kling et al. Citation2012). This small filamentous toxin producer grows both free and within the colonial mucilage of nontoxic and toxic bloom species, which may lead to spurious correlations between toxin levels and non-producers. Some of the cyanobacteria identified in LOW have been reported as producers of other cyanobacteria toxins (anatoxin-a, saxitoxin), but have not been evaluated for this lake.
Among the eukaryotic phytoplankton groups, diatoms represented only a minor component of the late-summer algal biomass throughout the lake and showed no clear south–north patterns in diatom abundance, which was slightly higher in zones 1 and 6 (2–7% TB, ). At both sites, the diatom assemblage was predominated by meso-eutrophic centric taxa, notably the filamentous species Aulacoseira granulata (57% and 74% of the diatom biomass at zones 1 and 6, respectively) and the large-celled centric Stephanodiscus niagarae (74% and 10% of the diatom biomass, respectively). Cryptophytes were relatively more important in zone 5, largely present as Cryptomonas reflexa and the nannoflagellate Plagioselmis nannoplanctica (syn. Rhodomonas minuta var. nannoplanctica). In zone 4 (Whitefish Bay), bactivorous chrysophytes were relatively more important (20% TB), notably large ochromonads and Pseudopedinella cf. elastica. These taxa were similarly significant in the spring assemblage in this more isolated region of the lake where average bacterial levels were low (), highlighting the likely importance of the microbial loop in zone 4, which has high water transparency and the lowest average total and cyanobacterial biomass across the six zones ().
Table 3. Average abundances of picoplankton (<2 µm) fractions across LOW zones, 2008–2010 (1-m samples) measured using flow cytometry. See text for abbreviations; APP/HPP = [PC + PE + Pk]/BAC.
Samples collected in the spring show a very different and often more diverse community, which is underrepresented by the current effort largely focused on the summer/fall blooms, yet these earlier assemblages are important, both in terms of their higher nutritional value to grazers and because they have a profound influence on spring nutrient inputs and their recycling and/or loss to sediments. Spring under-ice diatom blooms (mainly Aulacoseira islandica during low-snow years) have been observed in some northern embayments (H. Kling, pers. obs.), but rarely sampled. Anderson et al. (Citation2006) reported a mixed spring community of diatoms and cyanobacteria in the southern sectors in 2005, which shifted to an increasing predominance of cyanobacteria later in the season, but provided little taxonomic breakdown of these communities. In a 2004 study of the northern sectors, Chen et al. (Citation2007) reported a spring biomass dominated by a mix of small centric and pennate diatoms and cryptoflagellates, which shifted toward cyanobacterial dominance over the summer and early fall. In the current study, we examined spring samples collected in June 2010 in zones 1, 4, and 5, and found that in contrast to summer communities, the spring assemblage was dominated by diatoms and flagellates and significantly higher in the northern zones compared to the southern zone where biomass was largely dominated by green algae (). Cyanobacteria were far less important in the spring samples, accounting for 13–18% of the total phytoplankton biomass, mostly present as small (∼1–2 µm) celled (unidentified) taxa with small numbers of Limnothrix sp. and the benthic cyanobacterium Tychonema sp. present in the zone 1 sample, the latter likely indicative of resuspension. Unlike Anderson et al. (Citation2006), we saw very few Aphanizomenon in the spring sample from this southern sector (<1 µg/L). N2-fixing cyanobacteria were relatively more abundant in the northern sectors: filaments and akinetes of Dolichospermum (D. fuscum and to a lesser extent D. lemmermannii) represented 40% of the cyanobacteria biomass at zones 4 and 5 with small surface blooms of these taxa starting as early as mid-June in some years. The presence of Dolichospermum akinetes in these spring surface water samples suggests that these taxa may be seeded locally, and not largely derived from other basins. Aphanizomenon was present but at very low biomass in the two northern sectors (5–6 µg./L).
Figure 8. Spring biomass of major taxonomic groups in LOW from EC zones 1, 4, and 5, Jun 2010.
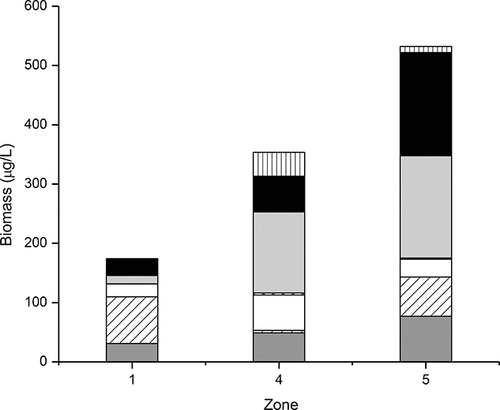
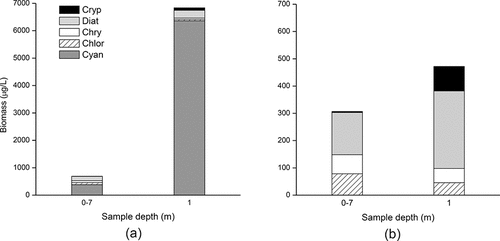
Figure 9. Phytoplankton biomass in surface (1 m) and mixed-depth integrated (0–7 m) samples at collected at station 710 (southern basin), Sep 2010. Left: all major taxa present in the samples; right: plot of all major groups with cyanobacteria removed to illustrate the changes in these less abundant taxa.
Diatoms comprised 40% of the total spring biomass at zone 4, as a mixture of pennate (Tabellaria fenestrata; 59% of the diatom biomass), Asterionella formosa (12%), Fragilaria (F. crotonensis, Fragilaria sp.; 12%), and centric diatoms (Lindavia bodanica complex [syn. Puncticulata bodanica or Cyclotella bodanica]; 16%), similar to the communities reported by Chen et al. (Citation2007). A comparable but slightly more diverse diatom community was observed in zone 5 (33% total biomass), with small numbers of the meso-eutrophic taxa (Stephanodiscus niagarae, S. agassizensis, Aulacoseira ambigua).
Chlorophyta were relatively more important in spring samples from zones 1 and 5 (). In the southern sector (zone 1), 57% of the chlorophyte biomass was present as small green picoeukaryotes, as also seen in the independently enumerated picoplankton samples ( and below); other green algae present in the sample included a mix of chloroccocales (Oocystis borgei, O. solitaria, Monoraphidium arcuatum, Ankyra lanceolata, Coelastrum pseudomicroporum, Pediastrum boryanum), volvocales (Chlamydomonas sp.), and desmids (Closterium acutum). In addition to the mix of chloroccocales, we observed small numbers of the colonial green alga Botryococcus braunii, known as a rich source of triterpene oils (botryococcenes). This species was present at slightly higher abundances in samples collected from August and September, particularly in northern zones.
Figure 10. Phycocyanin-rich picoplankton (PC; top left), phycoerythrin-rich picoplankton (PE; top right), eukaryotic picoplankton (Pk; bottom left), and bacteria (BAC; bottom right) across zones 1–6, 2008–2010 (1-m samples).
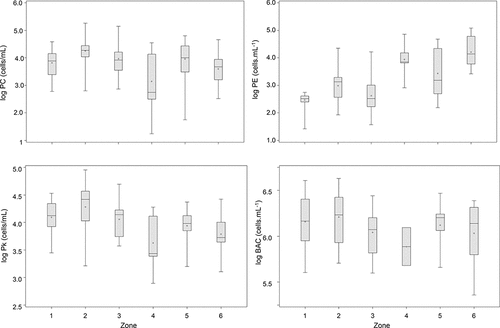
Figure 11. Phycoerythrin-rich picoplankton (PE; cells < 2 µm) vs. total dissolved P (top) and secchi depth (bottom); black symbols: southern zones 1–3; gray symbols: northern zones 4–6.
Much of the spring biomass at zones 4 and 5 was composed of flagellates, notably chrysophytes, cryptophytes, and dinoflagellates. These flagellates, many of which are mixotrophic, are typical spring taxa seen across a wide range of oligo-mesotrophic lakes. With the exception of the few scaled chrysophytes present in the sample (Mallomonas elongata, M. tonsurata), most of the chrysoflagellates at zone 4 were photophagotrophs (Dinobryon bavaricum, D. divergens, Pseudopedinella cf. elastica, small ochromonads). Cryptophytes were also important in zones 4 and 5, accounting for 17% total biomass in zone 4 (Cryptomonas reflexa, C. marssonii, Plagioselmis nannoplanctica) and 33% of the total biomass in zone 5 where there was a more diverse mix of these species (Cryptomonas reflexa, C. marssonii, C. pusilla, C. rostratiformis, Plagioselmis nannoplanctica, P. lacustris, Rhodomonas lens). Haptophytes (e.g., Chrysochromulina parva) were present but never very abundant in the samples collected during the surveys.
We note that the taxa reported here reflect our focus on spring and summer samples, which likely underrepresents assemblages present at other times of the year. This is apparent in the diatom assemblages described above, which show marked differences to those reported from the multi-season paleorecord, where there was a predominance of small centric unicells and multiple species of Aulacoseira (Reavie et al. Citation2017). The importance of these centric unicellular and filamentous diatoms in many under-ice winter and early spring assemblages has been demonstrated in other studies, notably from Lakes Winnipeg, Erie, and Baikal (e.g., Kling et al. Citation2012, Twiss et al. Citation2012, Katz et al. Citation2015).
Our sample analysis also included the major heterotrophic algal taxa, which are often not resolved from autotrophic phytoplankton biomass in most studies. These are primarily flagellates, which in LOW include the large heterotrophic taxa Gymnodinium helveticum, Collodictyon triciliatum, Gyromitus cordiformis, and smaller heterotrophs (Katablepharis ovalis, Cyathomonas truncata, Paraphysomonas vistita, Aulomonas perdyi). Spring samples contained a low abundance of these taxa, which showed a slightly higher biomass at zone 4 due to the presence of the large colorless dinoflagellate Gymnodinium helveticum.
Surface vs. integrated samples
U.S. and Canadian agencies involved in long-term LOW monitoring collect either discrete depth (1 m) or mixed-layer integrated samples, raising questions about the compatibility of these data, for example when examining spatiotemporal trends in water quality and algal biomass. In most waterbodies, seasonal changes in mixing and species dominance can produce significant differences in biomass and composition at different depths, particularly when communities are composed of buoyancy-regulating or large motile and non-motile taxa. This is of particular relevance in a spatially complex system such as LOW where basin morphometry and circulation differ considerably between basins. Comparisons using the 2008 summer data, for example, showed that on average, Chl-a was 34% higher in surface than mixed layer samples, and where there was a thick surface bloom this difference ranged up to 200–300% (Pascoe et al. Citation2014). As a community-based metric, however, Chl-a provides no insight into bias introduced by species segregation at different depths. For example, phytoplankton samples from zone 1 (station 710) collected during a significant surface bloom in September 2010 showed ∼10-fold variance in total biomass between the 1-m and integrated samples (6840 µg/L vs. 690 µg/L, respectively), and dramatic differences in the absolute and relative biomass of the dominant taxa (). The surface sample was composed almost entirely of cyanobacteria belonging to the Aphanizomenon flos-aquae complex (6270 µg/L or 98% of the total biomass). Other cyanobacteria (e.g., Microcystis sp., Pseudanabaena limnetica, Dolichospermum planctonicum, D. mendotae, Woronichinia eloranta, Aphanocapsa holsatica) were present at very low abundance. In comparison, the integrated (0–7 m) sample had a markedly lower cyanobacteria biomass and a shift in dominant species; Aphanizomenon flos-aquae was reduced to 105 µg/L (15% TB) while Microcystis (M. botrys) was more abundant (260 µg/L or 38% TB). Other cyanobacteria (Pseudanabaena spp.) were present but Dolichospermum and Woronichinia were not detected. Total diatom biomass and diversity was also significantly reduced in the integrated sample, for example with ∼50% lower abundance of the large centric diatom Stephanodiscus niagare (90 µg/L vs. 200 µg/L). Similarly, cryptophyte biomass was reduced considerably in the integrated sample from 90 µg/L to <4 µg/L, largely due to a lower abundance of Cryptomonas reflexa.
This issue has been examined by a number of studies, yet in many cases, differences in sampling procedures persist among agencies. Due to the current global concern with cHABs, much focus has been on methods used to monitor these events (e.g., Srivastava et al. Citation2013, Meriluoto et al. Citation2016). For example, recent studies evaluated differences in sample collection and analyses in long-term data for cyanobacteria blooms and water quality in the west basin of Lake Erie (Ho and Mikalak Citation2015, Bertani et al. Citation2016, Golnick et al. Citation2016), a large, shallow, wind-swept basin analogous to the southern end of LOW, and pointed to the variance introduced by surface scums. They did not, however, examine other important taxa such as diatoms (and account for a large proportion of the total biomass in Lake Erie; e.g., Allinger and Reavie Citation2013), taxonomic detail which can be key to understanding species dominance and succession. Overall, there is a need for both standardized sampling strategies among agencies and, where required, more tailored approaches to address specific questions (Srivastava et al. Citation2013). For example, surface samples are relevant to remote sensing and the potential risk of windblown scums to shorelines, while depth-integrated samples may underrepresent these surface aggregates. Where there is high water transparency, deep-layer maxima of motile or meroplanktic algal populations can account for a high proportion of algal biomass over the water column and drive successional changes in the surface plankton (e.g., Konopka Citation1989, Camacho Citation2006). In Lake 277 (Experimental Lakes Area, Ontario, Canada), for example, annual blooms of Aphanizomenon schindlerii develop in the metalimnion where nutrient levels are elevated, and manifest as surface blooms later in the season (D. Findlay et al. unpubl. data). Other taxa, such as bactivorous flagellates (e.g., chrysophytes) form deep-layer maxima in strata that are rich in bacteria and organic materials (e.g., Bird and Kalff Citation1989). Field protocols can be adapted to identify and sample these deep maxima, for example, using fluorescence profiles.
Picoplankton
Over the three-year sampling period, there were significant differences in abundance across the zones for some of the picoplankton groups, but considerable overlap within some of the north and south sectors (). Among the autotrophic picoplankton, the picocyanobacteria fractions (Synechococcus; may include Cyanobium) showed a marked difference in spatial patterns of abundance in surface samples (, ). There was a strong south–north increase in phycoerythrin-rich Synechococcus (PE), which varied in mean abundance from ∼1.2 × 102 cells/mL (zone 1) to ∼4.3 × 104 cells/mL (zone 6). In contrast, phycocyanin-rich picocyanobacteria (PC) were significantly more abundant throughout the lake (2.6 × 104 − 6.7 × 104 cells/mL) but showed no distinct south–north spatial patterns. Average PC abundance was highest in zone 2 and (to a lesser extent) zones 3 and 5, and lowest in zone 4.
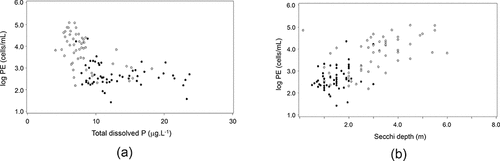
PE-rich Synechococcus (in 1-m samples) showed a highly significant negative relationship with total dissolved P (P < 0.001), which corresponded with the general gradient in TDP between the northern and southern sectors (). This picoplankton fraction was also negatively related to dissolved and particulate N (P < 0.0001), with significant interactive effect between N and P (). Their distribution and abundance showed highly significant relationships with water transparency, with a positive relationship with Secchi depth (P < 0.0001), and a negative relationship with ashed dry material (ADM, or suspended inorganic material; P < 0.0001; ; ). There was also a strong negative relationship between the proportion (%) of PE-rich picocyanobacterial and both total Chl-a and TP (data not shown; P < 0.0001), as observed in other studies (e.g., Voros et al. Citation1998). This suggests that the abundance of PE in LOW is affected by non-nutrient constraints, consistent with patterns seen in other lakes (e.g., Callieri Citation2008 and references therein) and our data indicate that light plays an important role. In many oligotrophic systems, most of the attenuation is caused by water molecules and occurs in the red spectral range, and the effective underwater PAR is predominated by the blue end of the spectrum (Kishino et al. Citation1984). In a study of the abundance of picoplankton groups in lakes spanning a wide trophic gradient, Voros et al. (Citation1998) concluded that an observed increase in the dominance of PC over PE picocyanobacteria in more eutrophic systems was related to greater light attenuation and a shift in underwater PAR from blue and UV to red wavelengths from an increased amount of adsorption and scattering by Chl-a and autochthonous DOC. To examine if this relationship holds in LOW, where high background DOC strongly attenuates shorter wavelengths (Binding et al. Citation2011b), picocyanobacterial abundances from selected sites (110, 130, 210, 230, 310, 410, and 750) were compared to changes in the wavelengths of light corresponding to the maximum absorbance of the two phycobilins (PE and PC) and Chl-a (565 nm, 620 nm, and 430 nm, respectively) using spectral attenuation data measured at each site during September 2010 (Binding, unpubl. data). The abundance of PE-rich cells (maximum at site 230) was consistent with changes in both phycobilin absorption minima across sites (Fig. S-2). In contrast, PC-rich cells (which peaked in abundance at site 410) showed no clear relationship with the spectral data but showed a highly significant relationship with DOC (), suggesting that their abundance is influenced by other factors, as also concluded by other studies (e.g., Stockner Citation1991, Voros et al. Citation1998, Callieri and Stockner Citation2000).
Table 4. Summary results of multivariate analysis for picoplankton groups (cells/mL) against major physico-chemical parameters (all P, N, and C fractions; Zmax, Zmix, Secchi, ashed dry mass [ADM], ash-free dry mass [AFDM}), DO at [bottom-2 m] depth, temperature of the mixed layer, Tzm) using backward selection (α = 0.05); only significant relationships reported.
Compared to picocyanobacteria, the eukaryotic picoplankton (Pk) fraction was less abundant and showed a general, albeit small, decrease from south to north (; ). Similar to PC, Pk had the highest average abundance in zone 2 (2.8 × 104 cells/mL) and lowest in zones 4 and 6 (7.2 × 103 cells/mL) and in fact there was a highly significant correlation between abundances of PC and Pk in all sample types (1 m, euphotic zone integrated, and 2 m above bottom; P < 0.0001). Average Pk abundances also showed a negative correlation with Secchi and a positive correlation with DOC (P < 0.0001, P = 0.0002, respectively) but no significant relationships with major nutrient fractions ().
Heterotrophic bacteria were the most abundant picoplankton fraction across all zones, and showed similar spatial patterns to Pk and to a lesser extent, PC (, ), although the wide variance about mean values translated to non-significant differences among zones. Bacteria were significantly correlated with PC and Pk, with a generally higher average abundance in the south basin, notably in zone 2, suggesting their abundance in the southern sectors is not directly derived from the Rainy River input. In fact, although there were too few data to test for significant differences among sites, there was a general decrease in surface water bacteria with distance from the Rainy River inflow, and in zone 2, a general increase in their numbers in a northward direction (data not shown; see Pascoe et al. Citation2014). Similar to the picocyanobacterial fractions (PE and PC), bacteria abundances tended to increase over the season. Average bacterial numbers were generally lower toward the bottom of the water column () although this trend was not statistically significant due to the high variance among sites and seasons (P > 0.05). This pattern may have been largely related to temperature: multivariate analysis indicated strong correlations between water temperature and bacterial abundance in grouped samples from surface, depth-integrated, and bottom-2 (P < 0.0001; ). There was less evidence that nutrients played a significant role in bacterial numbers, which showed no detectable relationship with DOC or major nutrients, except in bottom samples, where they were weakly correlated with NO3 (P < 0.05). There was very little systematic variance in the ratio of autotrophic:heterotrophic picoplankton (APP / HPP; ). Average values suggested higher relative autotrophic microbial processes in zones 2 and 6, but the interpretation of these data is limited by the significant variance about the mean values and unbalanced sample sizes. We note, however, that this analysis has only considered bottom-up constraints to picoplankton biomass, and given the abundance of mixotrophic and heterotrophic flagellates in this lake, it is likely that top-down interactions are important, particularly in the less eutrophic basins of LOW (e.g., Stocker and Shortreed 1994, Sommaruga and Robarts Citation1997). Currently we lack the data to evaluate the role of APP and HPP in energy and nutrient transfer and the overall productivity of the LOW food web and how this may be affected by changes in external loading and climate.
Table 5. Summary statistics, bacterial abundance (cells/mL), all sites, 2008–2010.
Nutrient deficiency
Spatiotemporal patterns in nutrients and deficiency indicators were used to evaluate whether LOW plankton are primarily P-limited or if N also plays a role as suggested by the late-season increases in N2-fixing cyanobacteria. Deficiency was examined on a seasonal basis at each zone and compared to ambient nutrient levels. There were some marked differences across the lake in seasonal patterns of the dissolved and particulate nutrient fractions (Pascoe et al. Citation2014) which generally corresponded with deficiency measures. Zones 1 and 6 showed a seasonal decrease in the proportion of TP present in the dissolved form (P < 0.007, P < 0.05, respectively), while the opposite trend occurred in zone 3 where %TDP increased from ∼40% to >60% (P < 0.007). Sestonic stochiometric ratios followed similar patterns, with higher overall P deficiency indicated in the northern segments, and a general increase in P deficiency in mid–late summer.
Nitrogen also demonstrated strong seasonality, with most zones showing increases in TN and total organic N (particulate + dissolved) in late summer. N-debt generally increased in late season samples and showed a good correspondence with C:N (Figs. S1, S3), although in most cases, sestonic ratios were indicative of greater deficiency than N-debt (). It is likely that both P and N were co-limiting during this period, as is similarly reported from many other lakes (e.g., North et al. Citation2007, Müller and Mitrovic Citation2015, O'Donnell et al. Citation2017). Average sestonic carbon: Chl-a indicated some degree of deficiency, especially in zones 1 and 5. This may reflect differences in the living organic C content of the seston and water transparency, with the southern shallower zones (zones 1–3) most influenced by riverine inputs and resuspended material, as reflected in Secchi and AFDM (Pascoe et al. Citation2014).
Table 6. Summary statistics for nutrient deficiency indicators, all data (1 m); bold font indicative of deficiency according to criterion levels (shown in brackets) from Healey and Hendzel (Citation1980). Stoichiometric ratios measured as moles, N-debt as µmN/Chl-a/day.
Least squares multivariate analyses showed no relationship between deficiency indicators and most major physico-chemical variables with the exception of Si, which was significantly related to both C: Chl-a and Chl-a/PP (; P < 0.01, P < 0.0002, respectively). This suggests that silica plays an important role in the planktonic community in which diatoms are important, particularly in spring and low snow cover years, when under-ice blooms can become Si limited (Hecky et al. Citation1986). Overall, none of the nutrient deficiency metrics can provide a conclusive measure of plankton nutrient status, but each can allow some insight into contemporaneous physiological processes. Short-term measures of uptake and cell quotas vary with growth and ambient supplies and other factors, such as Si and light (e.g., Sterner Citation2008). These measures are made at the community level and can be confounded by differences among co-occurring taxa in nutrient requirements, cell storage capacity, and deficiency. Many cyanobacteria, for example, are capable of luxury nutrient update and storage, and undergo vertical migration or passive transport away from the source of the supply by wind and currents. Sestonic ratios reflect precedent conditions, include all living and dead material and are often unrelated to short-term changes in nutrient supplies (e.g., Agren Citation2004) and there remains significant dispute about the value of these ratios (e.g., Hecky et al. Citation1993, Reynolds Citation1999).
Table 7. Results of multivariate analysis for general deficiency indicators against major physico-chemical and class variables (TDP, DIN, Si, Zmix/Zmax, Secchi) using backward selection (α = 0.05). Only significant relationships shown.
Summary and conclusions
LOW fits within the mid- (meso-eutrophic) range of north temperate lakes in terms of TP–algal biomass relationships, although there is significant variance in biomass for any given level of TP. In the southern sectors, the major nutrient source is the Rainy River, while northern, stratified basins show evidence of internal loading from hypoxic sediments. This requires further, more targeted investigation since it may play a key role in the timing and severity of the blooms. As seen in previous studies, the dominant species causing the widespread blooms are N2-fixing cyanobacteria, most of which (Aphanizomenon flos-aquae complex) are not known to be toxic. In general, spring samples are dominated by diatoms or other eukaryotic taxa, but at some sites, our study showed an early increase in some of the bloom-causing cyanobacteria (Dolichospermum species). Samples showed marked differences in taxa and biomass among sites and seasons, and no indication that bloom assemblages in the northern zones are directly derived from southern basins. Significant differences between surface and depth-integrated samples in Chl-a and algal biomass/composition highlight the need for a sampling strategy designed to address the issues being investigated. Sources of toxins in LOW are likely multiple and could be resolved using more focused physiological and molecular approaches. Some of the cyanobacteria identified in LOW have been reported elsewhere to produce neurotoxins that have yet to be evaluated in this waterbody (e.g., anatoxin-a, saxitoxin), and this should also be addressed in future studies.
Picoplankton abundance varied significantly across the lake but there was considerable overlap across north and south sectors. PE-rich picocyanobacteria showed a strong south–north increase but were generally an order of magnitude less abundant than PC-rich picocyanobacteria, which showed no distinct south–north spatial patterns. PE abundance appeared to be most strongly affected by non-nutrient constraints such as transparency and spectral attenuation, while PC and eukaryotic picoplankton densities were related to both light and nutrients. Overall, heterotrophic bacteria were more abundant than autotrophic picoplankton, with higher numbers in the southern sectors, notably zone 2. Our data, while limited, suggest that bacteria numbers are not directly derived from the Rainy River inflow, but show a strong seasonality in abundance, which peaks mid- or late summer. Overall, this study represents the first evaluation of picoplankton in LOW, and more frequent and focused work is required to evaluate the relative importance of bottom-up and top-down processes in regulating both these and larger plankton fractions. While the body of evidence from this and other studies suggests that P is ultimately limiting the LOW plankton, our data also show short-term deficiencies in N. Overall, there was higher P deficiency in the northern segments, and a general across-lake increase in P-deficiency in mid–late summer with evidence of N and P co-limitation in late summer and fall, concurrent with increases in N2-fixers. A strong link between low silica levels and indications of general plankton deficiency highlights the potential importance of this element in seasonal plankton dynamics in which diatoms and scaled flagellates play an important role, and which can be significantly influenced by winter (under-ice) conditions in large lakes (e.g., Kling et al. Citation2012, Beall et al. Citation2016). The influence of climate change on all of these factors is an important question to be addressed in future studies.
ULRM_1331282_Supplementary_Files.zip
Download Zip (105.7 KB)Acknowledgments
We would like to thank B. Lalonde, T. McDaniel, T. Pascoe, J. Guo, J. Copeland, L. Gagnon, T. Harvey, and C. Skrypka for assistance in this project, which was funded under the ECCC Lake Winnipeg Basin Initiative. We also thank two anonymous reviewers and the editors for their constructive comments, which helped us to greatly improve the article.
Related Research Data
References
- Agren GI. 2004. The C:N:P stoichiometry of autotrophs: theory and observations. Ecol Lett. 7(3):185–191.
- Allinger L, Reavie E. 2013. The ecological history of Lake Erie as recorded by the phytoplankton community. J Great Lakes Res. 39(3):365–382.
- Anderson J, Heiskary S, Hirst M. 2006. Lake of the Woods trophic status report. 2005 update. Minnesota Pollution Control Agency and Lake of the Woods Soil and Water Conservation District. Report No. 39-0002.
- Ballot A, Fastner J, Lentz M, Wiedner C. 2010. First report of anatoxin-a-producing cyanobacterium Aphanizomenon issatschenkoi in northeastern Germany. Toxicon. 56:964–971.
- Beall BFN, Twiss MR, Smith DE, Oyserman BO, Rozmarynowycz MJ, Binding CE, Bourbonniere RA, Bullerjahn GS, Palmer ME, Reavie ED, Waters LMK, Woityra LWC, McKay RML. 2016. Ice cover extent drives phytoplankton and bacterial community structure in a large north-temperate lake: implications for a warming climate. Environ Microbiol. 18:1704–1719.
- Bertani I, Steger CE, Obenour DR, Fahnenstiel GL, Bridgeman T, Johengen TH, Sayers MJ, Shuchman RA, Scavia D. 2016 Tracking cyanobacteria blooms: do different monitoring approaches tell the same story? Sci Total Environ. 575:294–308.
- Binding CE, Greenberg TA, Bukata R. 2011a. Time series analysis of algal blooms in Lake of the Woods using the MERIS maximum chlorophyll index. J Plankton Res. 33:1847–1852.
- Binding CE, Greenberg TA, Jerome JH. 2011b. An assessment of MERIS algal products during an intense bloom in Lake of the Woods. J Plankton Res. 33:793–806.
- Bird D, Kalff J. 1989. Phagotrophic sustenance of a metalimnetic phytoplankton peak. Limnol Oceanogr. 34:155–162.
- Callieri C. 2008. Picophytoplankton in freshwater ecosystems: the importance of small-sized phototrophs. Freshwat Rev. 1(1):1–28.
- Callieri C, Stockner J. 2000. Picocyanobacteria success in oligotrophic lakes: fact or fiction? J Limnol. 59:72–76.
- Camacho A. 2006. On the occurrence and ecological features of deep chlorophyll maxima (DCM) in Spanish stratified lakes. Limnetica 25(1–2):453–478.
- Canada Provincial Secretary's Office. 1859. Papers relative to the exploration of the country between Lake Superior and the Red River Settlement. https://archive.org/details/canadiana_org.
- Chen H, Burke JM, Dinsmore WP, Prepas EE, Fedorak PM. 2007. First assessment of cyanobacterial blooms and microcystin-LR in the Canadian portion of Lake of the Woods. Lake Reserv Manage. 23:169–178.
- Chen H, Burke JM, Mosindy T, Fedorak PM, Prepas EE. 2009. Cyanobacteria and microcystin-LR in a complex lake system representing a range in trophic status: Lake of the Woods, Ontario, Canada. J Plankton Res. 31:993–1008.
- Clapper J, Caudill SB. 2014. Water quality and cottage prices in Ontario. Appl Econ. 46:1122–1126.
- Dadheech PK, Selmeczy GB, Vasas G, Padisák J, Arp W, Tapolczai K, Casper P, Krienitz L. 2014. Presence of potential toxin-producing cyanobacteria in an oligo-mesotrophic lake in Baltic Lake District, Germany: an ecological, genetic and toxicological survey. Toxins. 6:2912–2931.
- DeSellas A, Paterson A, Clarke B, Baratono N, Sellers T. 2009 State of the Basin Report for the Lake of the Woods and Rainy River Basin. Lake of the Woods Water Sustainability Foundation, Kenora, Ontario.
- Downing JA, Watson SB, McCauley E. 2001. Predicting cyanobacteria dominance in lakes. Can J Fish Aquat Sci. 58:1905–1908.
- EI-Shaarawi A, Munawar M. 1978. Statistical evaluation of the relationships between phytoplankton biomass, chlorophyll a, and primary production in Lake Superior. J Great Lakes Res. 4:443–455.
- Environment Canada. 1997. Manual of Analytical Methods, Major Ions and Nutrients, Volume 1. National Laboratory for Environmental Testing (NLET), Burlington, Ontario.
- Falconer I. 2005. Cyanobacterial toxins of drinking water supplies. CRC Press, New York.
- Felip M, Catalan J. 2000. The relationship between phytoplankton biovolume and chlorophyll in a deep oligotrophic lake: decoupling in their spatial and temporal maxima. J Plankton Res. 22(1):91–105.
- Findlay DL, Kling HJ. 1998. Protocols for measuring biodiversity: phytoplankton in freshwater. www.eman-rese.ca/eman/ecotools/protocols/freshwater/phytoplankton/intro.html.
- Garmendia M, Borja A, France J, Revilla M. 2013. Phytoplankton composition indicators for the assessment of eutrophication in marine waters: present state and challenges within the European directives. Mar Pollut Bull. 66:7–16.
- Golnick PC, Chaffin JD, Bridgeman T, Zellner BC, Simons VE. 2016. A comparison of water sampling and analytical methods in western Lake Erie. J Great Lakes Res. 42:965–971.
- Gugger M, Lyra C, Henriksen P, Couté A, Humbert J-F, Sivonen K. 2002. Phylogenetic comparison of the cyanobacterial genera Anabaena and Aphanizomenon. Internat J Syst Evol Microbiol. 52:1–14.
- Hargan KE. 2010. A total phosphorus budget for the Lake of the Woods. MS thesis, Trent University, Peterborough, ON, Canada.
- Hargan, KE, Paterson AM, Dillon PJ. 2011. A total phosphorus budget for the Lake of the Woods and the Rainy River catchment. J Great Lakes Res. 37:753–763.
- Healey FP, Hendzel LL. 1980. Physiological indicators of nutrient deficiency in lake phytoplankton. Can J Fish Aquat Sci. 37:442–453.
- Hecky RE, Campbell P, Hendzel LL. 1993. The stochiometry of carbon, nitrogen, and phosphorus in particulate matter of lakes and oceans. Limnol Oceanogr. 38:709–724.
- Hecky RE, Kling HJ, Brunskill GJ. 1986. Seasonality of phytoplankton in relation to silicon cycling and interstitial water circulation in large, shallow lakes of central Canada. Hydrobiol. 138:117–126.
- Ho JC, Michalak AM. 2015. Challenges in tracking harmful algal blooms: a synthesis of evidence from Lake Erie. J Great Lakes Res. 41:317–325.
- Hutchinson Environmental Sciences. 2011. Phosphorus budget studies in the Lake of the Woods Watershed. Unpublished report; The Lake of the Woods Water Sustainability Foundation, Kenora (ON).
- Hyatt CV, Paterson AP, Rühland KM, Smol JP. 2011. Examining 20th century water quality and ecological changes in the Lake of the Woods, Ontario, Canada: a paleolimnological investigation. J Great Lakes Res. 37:456–459.
- Kasprzak P, Padisák J, Koschel R, Krienitz L, Gervais F. 2008. Chlorophyll a concentration across a trophic gradient of lakes: an estimator of phytoplankton biomass? Limnologica. 38:327–338.
- Katz SL, Izmest'eva LR, Hampton SE, Ozersky T, Shchapov K, Moore MV, Shimaraeva SV, Silow EA. 2015. The “Melosira years” of Lake Baikal: winter environmental conditions at ice onset predict under-ice algal blooms in spring. Limnol Oceanogr. 60:1950–1964.
- Kishino M, Booth CR, Okami N. 1984. Underwater radiant energy absorbed by phytoplankton, detritus, dissolved organic matter and pure water. Limnol Oceanogr. 29:340–349.
- Kling HJ, Laughinghouse D III, Smarda J, Komárek J, Acreman J, Bruun K, Watson SB. 2012. A new red colonial forming Pseudanabaena (Cyanoprokaryota, Oscillatoriales) from North American large lakes. Fottea. 12:327–339.
- Kling HJ, Watson SB, McCullough GK, Stainton MP. 2011. Bloom development and phytoplankton succession in Lake Winnipeg: a comparison of historical records with recent data. Aquat Ecosyst Health Mgmt. 14(2):219–224.
- Konopka A. 1989. Metalimnetic cyanobacteria in hard-water lakes: buoyancy regulation and physiological state. Limnol Oceanogr. 34:1174–1184.
- LaBaugh JW. 1995. Relation of algal biovolume to chlorophyll a in selected lakes and wetlands in the north-central United States. Can. J. Fish. Aquat. Sci. 52(2):416–424.
- Lowe CW. 1924. The freshwater algae of Canada. T Roy Soc Can XVIII, Ottawa.
- McLachlan J, Hammer U, Gorham P. 1963 Observations on the growth and colony habits of ten strains of Aphanizomenon flos-aquae. Phycologia. 2:157–168.
- Meriluoto J, Spoof L, Codd GA. 2016. Handbook of cyanobacterial monitoring and cyanotoxin analysis. John Wiley and Sons, Chichester, West Sussex, UK.
- [MPCA] Minnesota Pollution Control Agency. 2001. Rainy River basin overview. https://www.pca.state.mn.us/water/water-publications.
- Mitchell P, Prepas E. 1990. Atlas of Alberta lakes. University of Alberta Press, Edmonton.
- Müller S, Mitrovic SM. 2015. Phytoplankton co-limitation by nitrogen and phosphorus in a shallow reservoir: progressing from the phosphorus limitation paradigm. Hydrobiol. 744:255.
- North RL, Guildford SJ, Smith REH, Havens SM, Twiss MR. 2007. Evidence for phosphorus, nitrogen, and iron co-limitation of phytoplankton communities in Lake Erie. Limnol Oceanogr. 52:315–328.
- O'Donnell DR, Wilburn P, Silow EA, Yampolsky LY, Litchman E. 2017. Nitrogen and phosphorus colimitation of phytoplankton in Lake Baikal: insights from a spatial survey and nutrient enrichment experiments. Limnol Oceanogr. doi:10.1002/lno.10505.
- Pascoe T, McDaniel T, Watson SB, Yerubandi R, Binding C, Zhang W, Greenberg T. 2014. Environment Canada's Lake of the Woods Science Initiative 2008–2011. Environment and Climate Change Canada Report. ISBN 978-1-100-23605-6.
- Pla S, Paterson AM, Smol JP, Clark BJ, Ingram R. 2005. Spatial variability in water quality and surface sediment diatom assemblages in a complex lake basin: Lake of the Woods, Ontario, Canada. J Great Lakes Res. 31:253–266.
- Reavie ED, Edlund MB, Andresen NA, Engstrom DR, Leavitt PR, Schottler S, Cai M. 2017. Paleolimnology of the Lake of the Woods southern basin: continued water quality degradation despite lower nutrient influx. Lake Reserv Manage. 33:369–385.
- Reynolds CS. 1999. Non-determinism to probability, or N:P in the community ecology of phytoplankton. Archiv Hydrobiol. 146(1):23–35.
- Rühland K, Paterson AM, Hargan K, Jenkin A, Clark B, Smol JP. 2010. Reorganization of algal communities in the Lake of the Woods (Ontario, Canada) in response to turn of the century damming and recent warming. Limnol Oceanogr. 55:2433–2451.
- Sant'anna CL, De P, Azevedo MT, Werner VR, Dogo CR, Rios FR, De Carvalho LR. 2008. Review of toxic species of Cyanobacteria in Brazil. Algol Stud. 126:251–265.
- Sommaruga R, Robarts RD. 1997. The significance of autotrophic and heterotrophic picoplankton in hypertrophic ecosystems. FEMS Microb Ecol. 24:187–200.
- Srivastava A, Singh S, Ahn C-Y, Oh H-M, Asthana RK. 2013 Monitoring approaches for a toxic cyanobacterial bloom. Environ Sci Technol. 47(16):8999–9013.
- Sterner R. 2008. On the phosphorus limitation paradigm for lakes. Internat Rev Hydrobiol. 93:433–445.
- Stockner JG. 1991. Autotrophic picoplankton in freshwater ecosystems: the view from the summit. Int Revue ges Hydrobiol. 76:483–492.
- Stockner JG, Shortreed KS. 1994. Autotrophic picoplankton community dynamics in a pre-alpine lake in British Columbia, Canada. Hydrobiologia. 274(1–3):133–142.
- Twiss MR, McKay RML, Bourbonniere RA, Bullerjahn GS, Carrick HJ, Smith REH, Winter JG, D'Souza NA, Furey PC, Lashaway AR, et al. 2012. Diatoms abound in ice-covered Lake Erie: an investigation of offshore winter limnology in Lake Erie over the period 2007 to 2010. J Great Lakes Res. 38:18–30.
- Vaulot D, Courties C, Partensky F. 1989. A simple method to preserve oceanic phytoplankton for flow cytometric analyses. Cytometry. 10:629–635.
- Voros L, Callieri C, Balogh K, Bertoni R. 1998. Freshwater picocyanobacteria along a trophic gradient and light quality range. Hydrobiologia. 369/370:117–125.
- Watanabe MF, Harada K-I, Carmichael W, Fujiki H (eds). 1995. Toxic Microcystis. CRC Press, Boca Raton (FL).
- Watson S, McCauley E, Downing JA. 1997. Patterns in phytoplankton taxonomic composition across temperate lakes of differing nutrient status. Limnol Oceanogr. 42:487–495.
- Watson SB, Molot L. 2013. Harmful algal blooms. In Férard JF, Blaise C. (eds). Encyclopedia of Aquatic Ecotoxicology. Springer, The Netherlands.
- Watson SB, Ridal J, Boyer GL. 2008. Taste and odour and cyanobacterial toxins: impairment, prediction, and management in the Great Lakes. Can J Fish Aquat Sci. 65:1779–1796.
- Wood SA, Rasmussen JP, Holland PT, Campbell R, Crowe ALM. 2007. First report of the cyanotoxin Anatoxin-A from Aphanizomenon issatschenkoi (Cyanobacteria). J Phycol. 43:356–365.
- Zastepa A, Watson SB, Kling H, Kotak B. 2017. Spatial and temporal patterns in microcystin toxins in Lake of the Woods surface waters. Lake Reserv Manage. 33:433–443.
- Zhang W, Watson SB, Rao YR, Kling H. 2013. A linked hydrodynamic, water quality and algal biomass model for a large, multi-basin lake: a working management tool. Ecol Model. 269:37–50.
