
ABSTRACT
Paterson AM, Rühland KM, Anstey CV, Smol JP. 2017. Climate as a driver of increasing algal production in Lake of the Woods, Ontario, Canada. Lake Reserv Manag. 33:403–414.
Lake of the Woods (LOW) is a large, transboundary lake that straddles the provinces of Ontario and Manitoba, and the state of Minnesota. Although algal blooms have been reported in the lake since the early 1800s, monitoring data and anecdotal evidence suggest that toxic, cyanobacterial blooms have increased in frequency and intensity in recent years. However, total phosphorus inputs from the lake's primary tributary, the Rainy River, have declined significantly since the late 1960s. We explore this disconnect by examining spectrally-inferred determinations of chlorophyll a (Chl-a) in lake sediment cores, as a measure of past changes in aquatic primary production. Beginning in the late 1970s to early 1980s, inferred Chl-a increased at 5 impact sites in the north end of LOW that currently experience cyanobacterial blooms in late summer and autumn. In contrast, no change in Chl-a was observed at an oligotrophic reference site with much lower cyanobacteria biomass. At the impact sites, Chl-a generally showed no significant relationship to long-term trends in diatom-inferred total phosphorus concentrations, but was significantly and positively correlated to climatic variables, including mean annual air temperature at all sites and total annual precipitation at 4 sites. These data suggest that climate change may exacerbate algal blooms in this moderately-enriched lake. The results also show that the effects of climate change on aquatic production may be enhanced at sites with higher nutrient concentrations, likely because of positive feedbacks between cyanobacteria biomass, water temperature and nutrient availability. The impact of climate change should be considered carefully in future management initiatives.
Several recent studies and reviews have reported that the frequency, extent, and reporting of harmful algal blooms (HABs) have increased significantly over the past few decades in Canada (Winter et al. Citation2011, Pick Citation2016) and globally (O'Neil et al. Citation2012, Taranu et al. Citation2015). While nutrient enrichment, or elevated nutrient concentrations, is a key component of many cyanobacterial blooms (Watson et al. Citation1997, Heisler et al. Citation2008), emerging evidence suggests that climate change may be an aggravating factor (Paerl and Huisman Citation2008, LeBlanc et al. Citation2008, Weyhenmeyer and Broberg Citation2014). In Ontario, Canada, for example, HABs are now being reported in oligotrophic lakes within the Precambrian Shield, and these reports are occurring later in the ice-free season (e.g., November), coincident with warmer fall temperatures over the past 2 decades (Winter et al. Citation2011).
One lake that has experienced severe HABs in recent years is the Lake of the Woods (LOW), a large transboundary lake that straddles the borders of Minnesota and the Canadian provinces of Ontario and Manitoba. Its primary tributary, the Rainy River, contributes over 70 percent of the inflow and approximately 75 percent of the annual TP load to LOW (Hargan et al. Citation2011). Despite significant, long-term reductions in nutrient loading to LOW from the Rainy River since the 1970s (Hargan et al. Citation2011), the lake continues to experience severe, toxic, cyanobacterial blooms on an annual basis (Chen et al. Citation2007, Citation2009, DeSellas et al. Citation2009, Clark and Sellers Citation2014), and there is a perception that conditions have worsened over the past few decades. This apparent disconnect between historical nutrient loading and algal bloom intensity has created a conundrum for lake managers, triggering further research on the influence of multiple stressors on algal communities, including climate change (Rühland et al. Citation2008, Citation2010, Hyatt et al. Citation2011).
Cyanobacterial blooms in LOW are dominated by filamentous, nitrogen-fixing taxa (Dolichospermum, Aphanizomenon), with seasonal blooms common in mid-summer to mid-fall (Chen et al. Citation2009, Watson and Kling Citation2017). Remote sensing imagery of LOW from 2003 to 2011 has shown that bloom intensity, timing, and aerial extent may vary inter-annually in relation to local climate variables (Binding et al. Citation2011). For example, bloom intensity (defined as the peak chlorophyll concentration), and the aerial extent of blooms in LOW among years, were strongly associated with the sum of monthly mean temperatures from January to August, and negatively related to total precipitation from April to July. This suggests that bloom intensity was highest in warm, dry years (Binding et al. Citation2011).
Despite the public perception that blooms have increased in intensity and extent in recent years (Clark and Sellers Citation2014), Binding et al. (Citation2011) reported no significant trend in bloom intensity from 2003 to 2011. However, environmental changes within LOW and its catchment have occurred over decades, including the construction of hydrological controls (i.e., dams; Rühland et al. Citation2010, Serieyssol et al. Citation2009), increased nutrient loading associated with industrial, agricultural, and settlement activities (Anderson et al. Citation2017), changes in lake ice phenology (Rühland et al. Citation2008), and significant increases in mean annual and seasonal air temperatures, particularly since the early 1980s (Rühland et al. Citation2008, Citation2010). Thus, while monitoring efforts have increased markedly in the Rainy-LOW basin over the past decade (Anderson et al. Citation2017), these data are still too short in duration to fully assess the response of LOW to multiple stressors.
In the absence of long-term monitoring data, paleolimnological records can be used to assess long-term changes in physical, chemical, and biological conditions (Smol Citation2008). In large, complex lake basins such as LOW, an assessment of sediment cores from multiple sites can provide information on both spatial and temporal variation in environmental and ecological variables (Hyatt et al. Citation2011). Here, we assess long-term changes in spectrally-inferred chlorophyll a (Michelutti et al. Citation2010, Michelutti and Smol Citation2016) at 5 sites in the north end of LOW that currently experience severe, annual cyanobacterial blooms (“impact” sites), and at 1, low nutrient site (“reference site”) with markedly lower cyanobacteria biomass (Watson and Kling Citation2017). From these data, we assess: 1) whether or not the perceived increase in the intensification of algal blooms over the past 2 decades is recorded in lake sediment cores; and 2) if so, if this intensification is related to changes in climate (air temperature or precipitation), or inferred total phosphorus concentrations over the past century. The management implications of these findings are also discussed.
Materials and methods
Study sites and core collection
Lake of the Woods () (49°18'N, 94°36'W) is a large (∼3850 km2), international lake that is comprised of hundreds of interconnected bays, more than 14,000 islands, and over 10,000 km of shoreline (Heiman and Smith Citation1991, Pla et al. Citation2005). The north end of LOW is underlain by granitic Precambrian bedrock, and is deeper (∼15 to > 60 m maximum depth) than the south end, which is characterized by generally flatter terrain. The Rainy River-LOW basin is dominated by forests, wetlands, and lakes (∼96%), with developed areas accounting for ∼1.3% of the total basin area (Olmanson and Bauer Citation2017). Updated Landsat imagery for the basin shows a slight increase in forest harvesting between 1990 and 2010 (4.6% to 5.6% of the watershed area; Olmanson and Bauer Citation2017), but overall there have been minimal changes in land-use over the past 2 to 3 decades.
Figure 1. Map showing the location of the Lake of the Woods catchment within the Provinces of Ontario and Manitoba, and the State of Minnesota. The dashed oval shows the position of Lake of the Woods in the catchment, and the arrow shows the direction of its outflow into the Winnipeg River. The inset map shows the location of the 5 impact sites (circles), the reference site (square), and the Kenora airport climate station (star).
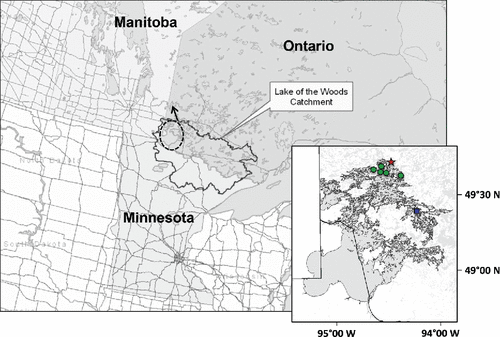
The lake sediments available for analyses for this study were from 6 sites in LOW, previously cored to examine long-term responses of algal communities (Rühland et al. Citation2008; Hyatt Citation2010) and chironomid assemblages (Summers et al. Citation2012) to multiple environmental stressors. This included 5 nutrient-rich sites with strong connections to the lake's main south-to-north channel, and a low nutrient site (“reference site”, ) in a deep bay that is hydrologically removed from the main inflow tributary, the Rainy River. Herein, the nutrient-rich locations are referred to as “impact sites” because of their higher nutrient concentrations, and because they experience severe, annual cyanobacterial blooms in late summer and/or fall (Chen et al. Citation2007, Citation2009). The 5 “impact sites” are located in the north sector of the lake (Bigstone Bay, Site PP1 (unofficial name), Forrest Island, Poplar Bay, White Partridge Bay), and the low-nutrient “reference” site is located in the east sector of the lake (Whitefish Bay) (, ). With the aid of hydrographic maps, sediment cores were collected in 2006 from the deepest locations of Bigstone Bay, Site PP1 (located between Thompson and Town islands), and at a location near Forrest Island, as part of a detailed study examining the response of algal communities to turn-of-the-century damming and recent warming (Rühland et al. Citation2010). In August 2008, sediment cores were collected from the central, deep water regions of Poplar and White Partridge bays, respectively (). At all locations, cores were obtained using a Glew gravity corer, and extruded on-site into 0.5 to 1.0 cm intervals using a Glew extruder (Glew et al. Citation2001). Samples were then kept cool until they could be stored in a cold room at the Paleoecological Environmental Assessment and Research Laboratory (PEARL) at Queen's University in Kingston, Ontario.
Table 1. a) Location, mixing regime (dimictic or polymictic), water column depth and year in which sediment cores were collected, spring total phosphorus (TP) concentrations, and the years averaged to determine the measured spring TP concentration for 5 impact sites and 1 reference site (in italics) in Lake of the Woods, Ontario. b) Min and max values, and 1971–2000 mean values, for mean annual temperature and total annual precipitation from the Kenora, ON climate station. Climate data are from the Historical Adjusted Climate Database for Canada, Environment Canada, for the City of Kenora, Ontario (49.78°N, 94.37°W; http://www.cccma.ec.gc.ca/hccd/).
A 55-cm core from the north end of Whitefish Bay was collected in March, 2002 by the Department of Fisheries and Oceans Canada (Rühland et al. Citation2008, Citation2010). Whitefish Bay is considered a reference site in our study as it is an oligotrophic (), deep bay that is hydrologically isolated from the main inflow tributary, and shows lower annual concentrations of cyanobacteria (Chen et al. Citation2009, Watson and Kling Citation2017).
210Pb dating
Approximately 20 sediment intervals per core from the impact sites were analyzed for 210Pb, 137Cs, and 214Bi activity using gamma spectroscopy at PEARL, following the procedures outlined by Schelske et al. (Citation1994). Briefly, sediment samples were freeze-dried and ∼0.5 g of dried sediment was placed into plastic test tubes and sealed with 2-Ton Epoxy® to ensure equilibrium between 226Ra and 214Bi prior to gamma counting. Chronologies for each core were calculated from estimates of 210Pb activities and cumulative dry mass using the constant rate of supply (CRS) model (Appleby Citation2001). All sediment intervals from Whitefish Bay were analyzed for 210Pb activity using gamma spectroscopy at the Freshwater Institute, Department of Fisheries and Oceans Canada in Winnipeg, Manitoba.
Climate data
Instrumental temperature and precipitation data used in this study are from the Historical Adjusted Climate Database for Canada, Environment Canada, for the City of Kenora, Ontario (49.78°N, 94.37°W; http://www.cccma.ec.gc.ca/hccd/) (). Temperature data from this station have been recorded since 1899, and show significant increases annually and in all seasons except autumn over the > 100-year record, with the highest rate of increase found in the winter and spring seasons (Rühland et al. Citation2010). Total annual precipitation also shows a significant positive linear trend over time, but is more variable than temperature with decadal-scale variations, and higher than average precipitation recorded in the late 1940s, the late 1960s, and since ∼1990. A comparison with other nearby stations shows that temperature trends are highly consistent, and that the Kenora data are representative of regional trends (Table 3.2 in DeSellas et al. Citation2009, in Rühland et al. Citation2010). Total annual precipitation is also significantly correlated across meteorological sites in the region (e.g., Kenora, ON, Winnipeg, MB, Lake 239 at the Experimental Lakes Area, ON, and International Falls, MN), but with generally weaker coherence among sites than observed for temperature (DeSellas et al. Citation2009). LOW currently experiences a continental climate, with mean July and January temperatures of ∼19.5 and −16.8 degrees Celsius respectively from 1971–2000. Total annual precipitation measured from 1971–2000 averaged 748.6 mm/year, but has been higher in recent years (∼864 mm/year, on average, from 2000 to 2007).
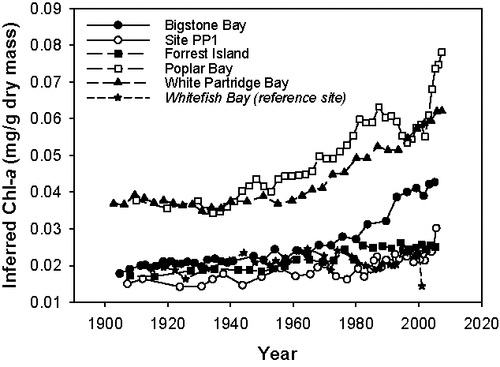
Figure 2. Line graphs of chlorophyll a (Chl-a in mg/g dry mass) changes over time, inferred using visible range spectroscopy. Data are shown for 5 impact sites and 1 reference site (in italics) in Lake of the Woods, Ontario. The time period represented by each core is: Bigstone Bay (∼1905 – 2006); Site PP1 (∼1907 – 2006); Forrest Island (∼1908 – 2006); Poplar Bay (∼1910 – 2008); White Partridge Bay (∼1903 – 2008); and Whitefish Bay (∼1915 – 2002).
Diatom-inferred total phosphorus (TP) concentrations
For comparison with long-term inferences of Chl-a, diatom-inferred TP concentrations were generated for each core using diatom species assemblage data and an inference model published by Hyatt et al. (Citation2011). The training set consists of 123 surficial diatom taxa and 71 lakes or sites from the Northern Lakes and Forest (NLF; 55 lakes, Reavie et al. Citation2005) ecoregion of northern Minnesota, and across LOW (16 sites). A weighted-averaging partial-least-squares model was developed for log TP using the software program C2 (bootstrapped r2 = 0.57, root mean square error of prediction = 0.150). Further information on data handling and model development can be found in Hyatt et al. (Citation2011).
Spectrally-inferred Chl-a
Chlorophyll a (Chl-a) was measured for selected sediment intervals using visible range spectroscopy (VRS), a rapid, non-destructive method for reconstructing aquatic production over time (i.e., paleo-production; Michelutti et al. Citation2010, Michelutti and Smol Citation2016). Briefly, sediments were freeze-dried and sieved (125 μm mesh) to remove the influence of water content and particle size on the spectral signal. Sediment spectra were then obtained using a FOSS NIRSystems Model 6500 series Rapid Content Analyzer (Tidestone Technologies Inc.), operating over the range of 400 to 2500 nm. The sediments were analyzed directly through the base of glass vials, with each sample representing an average of 32 scans. The algorithm developed by Michelutti et al. (Citation2005, Citation2010) was used to infer total Chl-a from sediment spectral data. As detailed in a recent review by Michelutti and Smol (Citation2016), the VRS technique captures both primary and degraded chlorophyll a because their visible spectroscopic properties are broadly similar. Thus, the VRS method tracks trends in whole lake production, and not the amount of degradation over time. This is important when reconstructing past aquatic production because Chl-a may undergo photo-degradation in the water column (Carpenter et al. Citation1986), or diagenetic modification once incorporated into lake sediments (Bianchi et al. Citation1991). The technique is sensitive to changes in sedimentary Chl-a on the order of 0.01 mg/g dry mass (Michelutti et al. Citation2005).
Statistical analyses
Long-term changes in spectrally-inferred Chl-a were plotted for all cores as both absolute (mg/g dry mass) and relative values (as Z-scores, standardized within cores to a mean of 0 and a standard deviation of 1). Chl-a values were also plotted versus diatom-inferred TP concentrations, and continuous air temperature and precipitation data from the Kenora climate station (from the early 1900s to the dates of core collections). Within cores, Pearson Product Moment correlations were run between spectrally-inferred Chl-a and diatom-inferred TP concentrations (comparing intervals for which both variables had been assessed). Direct comparisons between instrumental climate records and lake sediment data can be challenging, as the time period represented by each sediment interval may vary with core compression and varying sedimentation rates. Because the sediment intervals downcore may fluctuate from a few to several years, it is difficult to compare the sediment record with the annually resolved climate data. To address this issue, every available sedimentary sample was analyzed for Chl-a, and the instrumental climate data were averaged within cores to the duration of each sediment interval (with durations determined by the 210Pb chronologies). The interval-specific durations varied from 1 to 5 or 6 years within cores. Pearson Product Moment correlations were then run between the climate data and the spectrally-inferred Chl-a records. Performing multiple correlation tests increases the likelihood of erroneously reporting significant relationships. To correct for this, we applied Benjamini and Hochberg's (Citation1995) correction procedure for the possibility of a false discovery rate. Separate corrections were applied to the temperature and precipitation correlation tests. For graphing purposes only, mean annual air temperature and total annual precipitation are shown as annual data and as LOESS-smoothed values (smoothing factor = 0.3).
Results
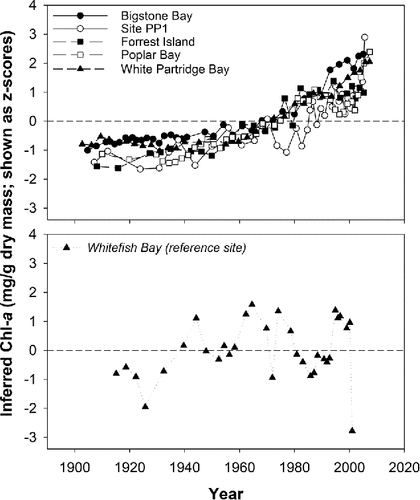
Spectrally-inferred estimates of Chl-a in surface sediments indicated that the reference site (Whitefish Bay) had the lowest concentrations, closely followed by sites PP1, Forrest Island, and Bigstone Bay (). Higher concentrations were found in White Partridge and Poplar bays, 2 sites that are more isolated from the main channel in Lake of the Woods (). At the deeper, larger reference site, where TP concentrations are much lower, and where annual cyanobacterial biomass is much less (Watson and Kling Citation2017), no clear trend over time was observed in the Chl-a data (, bottom panel). Furthermore, inferred Chl-a of the uppermost sample at the reference site was low, likely because sedimentation rates at the top of the core were high enough to resolve seasonal patterns, and the core was collected in the winter when chlorophyll levels were likely very low because of ice cover. Conversely, at the impact sites where measured TP is notably higher and where cyanobacterial blooms occur annually, distinct, temporally coherent increases in Chl-a were recorded over the past few decades, with concentrations consistently above long-term mean values since the late 1970s or early 1980s (, top panel).
Figure 3. Line graphs of chlorophyll a (Chl-a in mg/g dry mass) changes over time, inferred using visible range spectroscopy. Data are plotted as z-scores (standardized within cores to a mean of 0 and a standard deviation of 1), and are shown for 5 impact sites (top panel) and 1 reference site (bottom panel, in italics) in Lake of the Woods, Ontario. The time period represented by each core is: Bigstone Bay (∼1905 – 2006); Site PP1 (∼1907 – 2006); Forrest Island (∼1908 – 2006); Poplar Bay (∼1910 – 2008); White Partridge Bay (∼1903 – 2008); and Whitefish Bay (∼1915 – 2002).
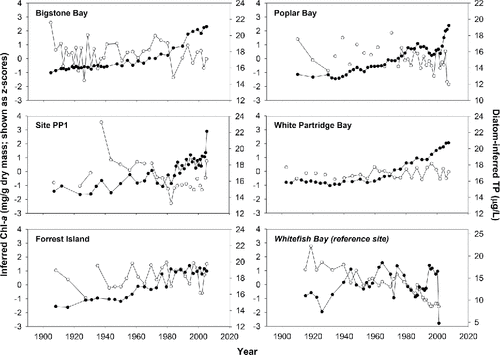
As noted by Hyatt et al. (Citation2011), and by comparing results from and in our study, the diatom-inferred TP model produced reliable predictions of modern-day TP concentrations at all sites (with a slight under-prediction in Bigstone Bay). At 4 of the impact sites, and at the reference site, there was no significant relationship between spectrally-inferred Chl-a and diatom-inferred TP concentrations (, ). In Poplar Bay, where there was a significant correlation between these variables, the relationship was negative, with Chl-a increasing over time as inferred TP concentrations declined gradually from ∼17 µg/L in the mid-twentieth century to ∼14 µg/L in recent decades.
Table 2. Performance results for Pearson Product Moment correlations of spectrally-inferred chlorophyll a (Chl-a) versus diatom-inferred total phosphorus (TP) concentrations (back-transformed from log values). The following metrics are shown: the correlation coefficient (r), p-value, and the number of intervals (n) compared within each core. Bold values are significant after correction for the False Discovery Rate (following Benjamini and Hochberg Citation1995). Results are shown for 5 impact sites and 1 reference site (in italics) in Lake of the Woods, Ontario.
Figure 4. Comparative line plots showing the relationship between spectrally-inferred chlorophyll a (Chl-a, solid points) and diatom-inferred total phosphorus (TP) concentrations (open points, back-transformed from log values) for 5 impacts sites and 1 reference site (in italics) in Lake of the Woods, Ontario. For some sites, diatom-inferred TP concentrations were not computed for every downcore sample. In those cases, the TP plots are not represented by continuous lines. Inferred Chl-a values are standardized as z-scores.
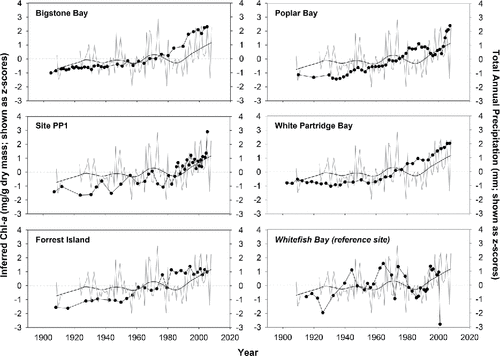
There was no significant relationship between spectrally-inferred Chl-a trends in the reference lake and either mean annual temperature or total annual precipitation data from the Kenora climate station (). In contrast, inferred Chl-a estimates from the 5 impact sites were highly and positively correlated to Kenora mean annual air temperature trends (, ). Based on Z-scores, both mean annual air temperature and spectrally-inferred Chl-a showed values above the long-term mean beginning in the late 1970s or early 1980s (). The relationship between inferred Chl-a and total annual precipitation was also significant at 4 of the 5 impact sites, although the strength of the relationship was generally weaker than with temperature (, ).
Table 3. Performance results for Pearson Product Moment correlations of spectrally-inferred chlorophyll a (Chl-a) versus mean annual temperature (MAT) and total annual precipitation at Kenora airport. The following metrics are shown: the correlation coefficient (r), p-value, and the number of intervals (n) compared within each core. Bold values are significant after correction for the False Discovery Rate (following Benjamini and Hochberg Citation1995). Results are shown for 5 impact sites and 1 reference site (in italics) in Lake of the Woods, Ontario.
Figure 5. Comparative line plots showing the relationship between spectrally-inferred chlorophyll a (Chl-a, solid points) and mean annual temperature (MAT) at Kenora airport. MAT is plotted as both annual means (light grey line), and as LOESS-smoothed values (black line), calculated from the annual mean data (smoothing factor = 0.3). Inferred Chl-a and MAT values are standardized as z-scores.
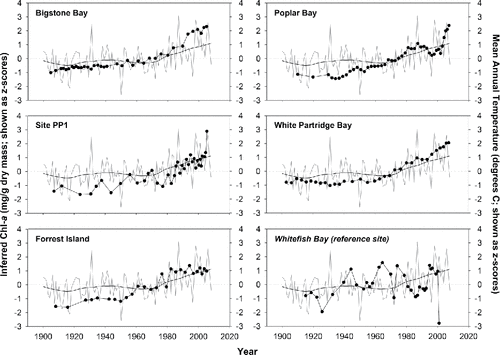
Figure 6. Comparative line plots showing the relationship between spectrally-inferred chlorophyll a (Chl-a, solid points) and total annual precipitation at Kenora airport. Precipitation is plotted as both annual means (light grey line), and as LOESS-smoothed values (black line), calculated from the annual mean data (smoothing factor = 0.3). Inferred Chl-a and precipitation values are standardized as z-scores.
Discussion
The varying degrees of change in historical Chl-a concentrations at the study sites are broadly consistent with differences in trophic status between the reference and impact sites. Increases in Chl-a at the nutrient-rich impact sites that experience late summer cyanobacterial blooms, coupled with survey data showing that cyanobacteria dominate the phytoplankton biomass at these sites at an annual (ice-free season) scale (Chen et al. Citation2007, Watson and Kling Citation2017), suggest that Chl-a trends are tracking long-term changes in cyanobacteria production. This is in contrast with the oligotrophic reference site (Whitefish Bay) where cyanobacteria occur at lower abundance, and where a directional trend in Chl-a was not observed over the past century.
Differences in morphometry between the impact and reference sites may contribute to site-specific differences in inferred aquatic production over time. For example, the reference site is deeper and larger than the impact sites, and a dilution effect may partly explain its lower nutrient levels and muted response to recent climate change. However, despite its larger size and volume, previous work has shown that the taxonomic composition of algal communities in Whitefish Bay is indeed sensitive to recent climate change. Using the same sediment core examined in our study, Rühland et al. (Citation2008) noted significant relationships between changes in sedimentary diatom composition and long-term changes in air temperature and ice-records.
Thus, while morphometry may be an indirect cause for the lack of trend in inferred Chl-a at the reference site, the variation in response across sites is likely driven by site-specific differences in trophic status, and higher cyanobacteria biomass (Watson and Kling Citation2017) at the impact sites. As noted by others (Taranu et al Citation2012), climate (temperature) may become an important driver of cyanobacterial abundance once critical nutrient thresholds are crossed, and the effects of temperature on algal biomass are often clearer at sites with elevated nutrient concentrations. This may be because positive feedback loops develop between cyanobacteria, water temperature and nutrient availability, as algal biomass increases (Taranu et al. Citation2012). As blooms develop, they absorb sunlight, further increasing surface water temperatures (Kahru et al. Citation1993). In turn, this may enhance thermal stability, increase the length of the growing season, and increase the period of thermal stratification, all favouring buoyant cyanobacteria taxa that can regulate their position in the water column (Paerl et al. Citation2011). Blooms may also increase nutrient availability through nitrogen fixation (Schindler et al. Citation2008), and via the recycling of phosphorus from lake sediments (James Citation2017). Finally, at sites where hypolimnetic anoxia occurs in late summer (e.g., Poplar Bay; Summers et al. Citation2012), a lengthening of the ice-free season may worsen seasonal oxygen depletion, thus increasing internal loading of phosphorus from lake sediments and further supporting blooms (Vahtera et al. Citation2007).
Warmer temperatures may also provide a direct competitive advantage to cyanobacteria, even in the absence of long-term increases in nutrient levels (McQuatters-Gollop et al. Citation2007, Weyhenmeyer and Broberg Citation2014). Some species grow preferentially at higher temperatures (Jöhnk et al. Citation2008) and through buoyancy regulation may optimize their position in the water column. As noted above, this allows cyanobacteria to dominate during periods of enhanced thermal stability of the water column (Paerl and Huisman Citation2009), while also gaining access to nutrient-rich waters at depth in the water column (Kromkamp and Walsby Citation1990).
Increased precipitation since ∼Citation1990 may also have contributed to the recent rise in inferred aquatic production. However, the relationship between Chl-a and precipitation is generally weaker than its relationship with temperature, and higher than average precipitation over the past 2 decades shows no relationship with diatom-inferred TP concentrations. Thus, while increases in the amount and intensity of precipitation can affect algal production through enhanced nutrient loading (Paerl and Huisman Citation2009), this mechanism was not obvious in our study. In part, this may be due to our analysis of total annual precipitation, where extreme events may be lost in the annual number. Further work should be done to assess the importance of seasonal and finer-scale climate signals on algal production in LOW, as unusual, short-lived precipitation events are expected to become more frequent with climate change (Chiotti and Lavender Citation2008).
As observed by Hyatt et al. (Citation2011) in a comparison of present-day and pre-industrial sediment samples from 17 sites within the Ontario portion of LOW, diatom-inferred TP concentrations showed no change or a slight decline over the past few decades at all of our sites. This may seem counterintuitive given the rise in primary production over the same time period. However, similarities in the timing of change in aquatic production among the impact sites, and significant correlations between Chl-a trends and Kenora mean annual temperature records, suggest that pronounced warming over the past few decades has triggered an increase in the intensity of nuisance blooms at these already nutrient-rich sites. While spring TP concentrations at these sites suggest only moderate nutrient levels, seasonal data show that the highest nutrient concentrations commonly occur in the mid to late summer, coinciding with peak cyanobacterial abundance (DeSellas et al. Citation2009).
Notably, we observed similar timing and rates of increase in Chl-a across all of the impact sites, regardless of the mixing regime (i.e., dimictic versus polymictic sites, , ). In LOW, as has been observed elsewhere (Mitchel and Prepas 1990 in Taranu et al. Citation2012), stratification may develop at polymictic sites over a period of several calm days, and satellite imagery from the south end of LOW suggests that short-term fluctuations in wind-induced mixing can have a profound effect on algal biomass detection from remote sensing (Binding et al. Citation2010). Over decades, as regional wind speeds have declined (Herb et al. Citation2005, Rühland et al. Citation2015) and water temperatures have increased, the number and duration of calm periods may have increased, favoring buoyant cyanobacteria that dominate the LOW phytoplankton assemblage (e.g., Aphanizomenon and Dolichospermum). The deployment of monitoring buoys in large, shallow Big Traverse Bay (south end of LOW) suggests that calm periods may develop over hours to days, resulting in rapid depletion of oxygen near the sediment water interface (St. Croix Watershed Research Station, Science Museum of Minnesota, unpublished data). These extended calm periods may enhance nutrient availability through internal loading, but further work is needed to confirm this phenomenon. Thus, both direct (increases in temperature, enhanced thermal stratification) and indirect effect (increases in nutrient availability) from climate change are likely at play.
Increases in spectrally-inferred Chl-a concentrations may also reflect an increase in the length of the open-water season (Michelutti et al. Citation2010), as a longer growing season may allow more time for the incorporation of algae and Chl-a into the sediments (Nelligan et al. Citation2016). In Whitefish Bay, for example, the length of the ice-free season increased by ∼ 28 days from 1964 to 2006 (Rühland et al. Citation2008). However, this cannot be the sole explanation for the observed increases in inferred Chl-a over time, as Chl-a did not increase in the relatively nutrient-poor Whitefish Bay over time, despite a large increase in growing season length. Also, other limnological indicators have changed at these sites, consistent with climate-induced changes to water column properties and habitat availability. This includes long-term changes in diatom species composition (Rühland et al. Citation2010) and declines in chironomid-inferred deep-water oxygen concentrations (Summers et al. Citation2012) that are related to shifts in ice cover, and changes in thermal stability and stratification (Rühland et al. Citation2015).
In summary, our results support the public perception that algal blooms have increased in intensity in recent decades, with notable changes occurring since the late 1970s or early 1980s. However, as noted above, the observed increases in algal production occurred during a period when diatom-inferred and measured (Clark and Sellers Citation2014) TP concentrations were stable or in decline. Furthermore, increases in Chl-a have occurred despite significant reductions in external nutrient loading from the Rainy River since the 1970s (Hargan et al. Citation2011), suggesting that improvements within the lake may lag management initiatives, or require further investigation. As management efforts continue, including an on-going Total Maximum Daily Load study in the Minnesota waters of the south end of LOW, our results highlight the need for careful consideration of the effects of climate change in future scenario modeling. Climate-induced changes in lake physical properties, and enhanced internal loading of nutrients, may partly counteract the benefits of implementing Best Management Practices and watershed management plans in the basin.
Acknowledgments
We thank Tom Mosindy (retired) and Gavin Olson from the Ministry of Natural Resources and Forestry Fisheries Assessment Unit in Kenora, Ontario, for guidance and logistical support when collecting cores. We also thank Mike Stainton (Department of Fisheries and Ocean Canada) for providing raw sedimentary material and 210Pb dates for the Whitefish Bay core (reference site). This project was supported by a Natural Sciences and Engineering Research Council of Canada grant and an Ontario Ministry of the Environment Best in Science grant to JPS. Fieldwork costs were also supported by the Ontario Ministry of the Environment and Climate Change.
References
- Appleby PG. 2001. Chronostratigraphic techniques in recent sediments. In: Last WM, Smol JP, editors. Tracking Environmental Change Using Lake Sediments. Volume 1: Basin Analysis, Coring, and Chronological Techniques. Dordrecht: Kluwer Academic Publishers. p. 171–203.
- Anderson JP, Paterson AM, Reavie ED, Edlund MB, Rühland KM. 2017. An introduction to Lake of the Woods – from science to governance in an international waterbody. Lake Reserv Manage. 33:325–334.
- Benjamini Y, Hochberg Y. 1995. Controlling the false discovery rate: a practical and powerful approach to multiple testing. J R Stat Soc B Methodol. 57:289–300.
- Bianchi TS, Findlay S, Fontvieille D. 1991. Experimental degradation of plant materials in Hudson River sediments. Biogeochem. 12:171–187.
- Binding CE, Greenberg TA, Jerome JH, Bukata RP, Letourneau G. 2010. An assessment of MERIS algal products during an intense bloom in Lake of the Woods. J Plank Res. 33:793–806.
- Binding CE, Greenberg TA, Bukata RP. 2011. Time series analysis of algal blooms in Lake of the Woods using the MERIS maximum chlorophyll index. J Plank Res. 33:1847–1852.
- Carpenter SR, Elser MM, Elser J. 1986. Chlorophyll production, degradation, and sedimentation: implications for paleolimnology. Limnol Oceanogr. 31:112–124.
- Chen H, Burke JM, Dinsmore WP, Prepas EE, Fedorak PM. 2007. First assessment of cyanobacterial blooms and microcystin-LR in the Canadian portion of Lake of the Woods. Lake Reserv Manage. 23:169–178.
- Chen H, Burke JM, Mosindy T, Fedorak PM, Prepas EE. 2009. Cyanobacteria and microcystin-LR in a complex lake system representing a range in trophic status: Lake of the Woods, Ontario, Canada. J Plank Res. 31:993–1008.
- Chiotti Q, Lavender B. 2008. Ontario. In: Lemmen S, Warren FJ, Lacroix J, Bush E (Eds). From impacts to adaptation: Canada in a changing climate 2007. Government of Canada. P. 227–274.
- Clark BJ, Sellers T. 2014. Rainy-Lake of the Woods State of the Basin Report. 2nd Edition. Multi-Agency Technical Report. http://www.lowwsf.com/progress-we-are-making/docman/state-of-the-basin-report.html. 225 p.
- DeSellas AM, Paterson AM, Clark BJ, Baratono NG, Sellers TJ. 2009. State of the Basin Report for the Lake of the Woods and Rainy River basin. Multi-agency Technical Report. http://www.lowwsf.com/progress-we-are-making/docman/state-of-the-basin-report.html. 133 p.
- Glew JR, Smol JP, Last WM. 2001. Sediment core collection and extrusion. In: Last WM, Smol JP, editors. Tracking Environmental Change Using Lake Sediments. Volume 1: Basin Analysis, Coring, and Chronological Techniques. Dordrecht: Kluwer Academic Publishers. p. 73–105.
- Hargan KE, Paterson AM, Dillon PJ. 2011. A total phosphorus budget for the Lake of the Woods and the Rainy River catchment. J Great Lakes Res. 37:753–763.
- Heiman C, Smith K. 1991. Clearwater Bay Cottage Pollution Control Program, 1989–1990. Ontario Ministry of the Environment, Kenora, ON. Data Report.
- Heisler J, Gilbert PM, Burkholder JM, Anderson DM, Cochlan W, Dennison WC, Dortch Q, Gobler CJ, Heil CA, Humphries E, Lewitus A, Magnien R, Marshall HG, Sellner K, Stockwell DA, Stoecker DK, Suddleson M. 2008. Eutrophication and harmful algal blooms: a scientific consensus. Harmful Algae. 8:3–13.
- Herb W, Mohseni O, Stefan H. 2005. Lake of the Woods shoreline erosion: analysis of historical shorelines, climate and lake level. University of Minnesota, Project Report No. 466, prepared for the Minnesota Pollution Control Agency. St. Paul, MN.
- Hyatt CV. 2010. A diatom-based paleolimnological investigation of historical water-quality and ecological changes in the Lake of the Woods, Ontario. [master's thesis]. [Kingston (ON)]: Queen's University.
- Hyatt CV, Paterson AM, Rühland KM, Smol JP. 2011. Examining 20th century water quality and ecological changes in the Lake of the Woods, Ontario, Canada: A paleolimnological investigation. J Great Lakes Res. 37:456–469.
- James W. 2017. Diffusive phosphorus fluxes in relation to the sediment phosphorus profile in Big Traverse Bay, Lake of the Woods. 33:360–368.
- Jöhnk KD, Huisman J, Sharples J, Sommeijer B, Visser PM, Strooms J. 2008. Summer heatwaves promote blooms of cyanobacteria. Glob Change Biol. 14:495–512.
- Kahru M, Leppänen JM, Rud O. 1993. Cyanobacterial blooms cause heating of the sea surface. Mar Ecol Prog Ser. 101:1–7.
- Kromkamp J, Walsby AE. 1990. A computer model of buoyancy and vertical migration in cyanobacteria. J Plankton Res. 12:161–183.
- LeBlanc S, Pick FR, Hamilton PB. 2008. Fall cyanobacterial blooms in oligotrophic-to-mesotrophic temperate lakes and the role of climate change. Verh Int Verein Limnol. 30:90–94.
- McQuatters-Gollop A, Raitsos DE, Edwards M, Pradhan Y, Mee LD, Lavender SJ, Attrill MJ. 2007. A long-term chlorophyll data set reveals regime shift in North Sea phytoplankton biomass unconnected to nutrient trends. Limnol Oceanogr. 52:635–648.
- Michelutti N, Blais JM, Cumming BF, Paterson AM, Rühland KM, Wolfe AP, Smol JP. 2010. Do spectrally inferred determinations of chlorophyll a reflect trends in lake trophic status? J Paleolimnol. 43:205–217.
- Michelutti N, Smol JP. 2016. Visible spectroscopy reliably tracks trends in paleo-production. J Paleolimnol. 56:253–265.
- Michelutti N., Wolfe AP, Vinebrooke RD, Rivard B, Briner JP. 2005. Recent primary production increases in arctic lakes. Geophys Res Lett. 32:L19715. DOI: 10.1029/2005GL023693.
- Nelligan C, Jeziorski A, Rühland KM, Paterson AM, Smol JP. 2016. Managing lake trout lakes in a warming world: a paleolimnological assessment of nutrients and lake production at three Ontario sites. Lake Reserv Manage. 32:315–328.
- Olmanson L, Bauer M. 2017. Land cover classification of the Lake of the Woods/Rainy River Basin by object-based image analysis of Landsat and lidar data. Lake Reserv Manage. 33:335–346.
- O'Neil JM, Davis TW, Burford MA, Gobler CJ. 2012. The rise of harmful cyanobacterial blooms: The potential roles of eutrophication and climate change. Harmful Algae. 14:313–334.
- Paerl HW, Huisman J. 2008. Blooms like it hot. Science. 320:57–58.
- Paerl HW, Huisman J. 2009. Climate change: a catalyst for global expansion of harmful cyanobacterial blooms. Env Microbiol Rep. 1:27–37.
- Paerl HW, Hall NS, Calandrino ES. 2011. Controlling harmful cyanobacterial blooms in a world experiencing anthropogenic and climatic-induced change. Sci Tot Environ. 409:1739–1745.
- Pick FR. 2016. Blooming algae: a Canadian perspective on the rise of toxic cyanobacteria. Can J Fish Aquat Sci. 73:1149–1158.
- Pla S, Paterson AM, Smol JP, Clark BJ, Ingram R. 2005. Spatial variability in water quality and surface sediment diatom assemblages in a complex lake basin: Lake of the Woods, Ontario, Canada. J Great Lakes Res. 31:253–266.
- Reavie E, Kingston J, Peterson M, Edlund M. 2005. Monitoring diatom algae in northeastern Minnesota. Final report on sediment diatom reconstruction model for Minnesota lakes. NRRI Technical Report NRRI/TR-2005/18.
- Rühland KM, Paterson AM, Smol JP. 2008. Hemispheric-scale patterns of climate-related shifts in planktonic diatoms from North American and European lakes. Glob Change Biol. 14:2740–2754.
- Rühland KM, Paterson AM, Hargan K, Jenkin A, Clark BJ, Smol JP. 2010. Reorganization of algal communities in the Lake of the Woods (Ontario, Canada) in response to turn-of-the-century damming and recent warming. Limnol Oceanogr. 55:2433–2451.
- Rühland KM, Paterson AM, Smol JP. 2015. Lake diatom responses to warming: reviewing the evidence. J Paleolimnol. 54:1–35.
- Schelske CL, Peplow A, Brenner M, Spencer CN. 1994. Low-background gamma counting: applications for 210 Pb dating of sediments. J Paleolimnol. 10:115–128.
- Schindler DW, Hecky RE, Findlay DL, Stainton MP, Parker BR, Paterson MJ, Beaty KG, Lyng M, Kasian SEM. 2008. Eutrophication of lakes cannot be controlled by reducing nitrogen input: Results of a 37-year whole-ecosystem experiment. PNAS. 105:11254–11258.
- Serieyssol CA, Edlund MB, Kallemeyn LW. 2009. Impacts of settlement, damming, and hydromanagement in 2 boreal lakes: a comparative paleolimnological study. J Paleolimnol. 42:497–513.
- Smol JP. 2008. Pollution of lakes and rivers: a paleoenvironmental perspective. Second edition. Oxford: Blackwell Publishing. 383 p.
- Summers JC, Rühland KM, Kurek J, Quinlan R, Paterson AM, Smol JP. 2012. Multiple stressor effects on water quality in Poplar Bay, Lake of the Woods, Ontario, Canada: a midge-based assessment of hypolimnetic oxygen conditions over the last two centuries. J Limnol. 71:34–44.
- Taranu ZE, Gregory-Eaves I, Leavitt PR, Bunting L, Buchaca T, Catalan J, Domaizon I, Guilizzoni P, Lami A, McGowan S, Moorhouse H, Morabito G, Pick FR, Stevenson MA, Thompson PL, Vinebrooke RD. 2015. Acceleration of cyanobacterial dominance in north temperate-subarctic lakes during the Anthropocene. Ecol Lett. 18:375–384.
- Taranu ZE, Zurawell RW, Pick F, Gregory-Eaves I. 2012. Predicting cyanobacterial dynamics in the face of global change: the importance of scale and environmental context. Glob Change Biol. 18:3477–3490.
- Vahtera E, Conley DJ, Gustafsson BG, Kuosa H, Pitkänen H, Savchuk OP, Tamminen T, Viitasalo M, Voss M, Wasmund N, Wulff F. 2007. Internal Ecosystem Feedbacks Enhance Nitrogen-fixing Cyanobacteria Blooms and Complicate Management in the Baltic Sea. Ambio. 36:186–194.
- Watson SB, Kling H. 2017. Lake of the Woods phyto- and picoplankton: spatiotemporal patterns in blooms, community composition, and nutrient status. Lake Reserv Manage. 33:415–432.
- Watson SB, McCauley E, Downing JA. 1997. Patterns in phytoplankton taxonomic composition across temperate lakes of differing nutrient status. Limnol Oceanogr. 42:487–495.
- Weyhenmeyer GA, Broberg N. 2014. Increasing algal biomass in Lake Vänern despite decreasing phosphorus concentrations: a lake-specific phenomenon? Aquat Ecosys Health Manage. 17:341–348.
- Winter JG, DeSellas AM, Fletcher R, Heintsch L, Morley A, Nakamoto L, Utsumi K. 2011. Algal blooms in Ontario, Canada: Increases in reports since 1994. Lake Reserv Manage. 27:107–114.
