
Abstract
Jacques O, Pienitz R, Ibrahim G. 2020. Paleolimnological assessment of long-term changes in a boreal recreational lake of the Fermont mining region (subarctic Quebec, Canada). Lake Reserv Manage. 36:314–334.
Lac Carheil is a boreal recreational lake located near Fermont, a remote town of subarctic Quebec built in the 1970s to support iron ore mining activities. Harmful cyanobacterial blooms were recently observed at its surface, suggesting that it suffered from eutrophication. However, long-term water quality data were lacking to properly understand its current condition. We therefore undertook a paleolimnological study to investigate past limnological changes in the lake based on the multiproxy analysis of sediment cores. The cores retrieved from Lac Carheil were marked by numerous distinct laminae representing past episodes of high-energy events in the lake catchment. Furthermore, diatom assemblages of the sediments revealed that the limnological conditions of the lake had been relatively stable for more than a thousand years before beginning to change around 1840 AD. This was mainly evidenced by the emergence of species indicative of nutrient enrichment (e.g., Aulacoseira subarctica, Fragilaria crotonensis), which suggested an early eutrophication potentially attributable to climate change. The period of modern settlement (1971–2015) was characterized by a decrease in the carbon to nitrogen ratio (C/N) of the sediments, and marked increases in their organic matter content, diatom total abundance, and diatom-inferred total phosphorus concentrations, which indicated an enhanced primary production. Nevertheless, our results suggested that the disturbance level of the lake was weak and that its prospect for recovery is good. Our study provides valuable reference data to assist the management of Lac Carheil, and reveals first insights into the sedimentological and limnological history of the Fermont mining region.
Lac Carheil is located in the Fermont mining region, a remote area of subarctic Quebec (Canada). Between 2005 and 2011, this lake experienced harmful cyanobacterial blooms that raised concerns about its health, as these often result from eutrophic waters (Paerl and Otten Citation2013). Lac Carheil has great recreational and touristic values for the town of Fermont and is also at the head of the Moisie River, a stream renowned for sports fishing and paddling activities. Efforts have thus been made to reduce nutrient inputs to the lake in order to enhance its water quality and eradicate the cyanobacterial blooms. In particular, major modifications were made to the town wastewater treatment plant in 2010–2011, including the addition of a phosphorus removal system. The point of discharge of the plant is located upstream from Lac Carheil, and the municipal wastewater has been identified in the past as the only known important anthropogenic source of nutrient inputs to this water body (OBVD Citation2010).
Until recently, Lac Carheil had only been sporadically monitored. Verette et al. (Citation1976) evaluated its water quality a few years after the construction of the town of Fermont following the opening of the Mont-Wright iron ore mining complex at the beginning of the 1970s. They determined that the lake had at the time unproductive waters. Its water quality was only reassessed many years later, in 2011. In this most recent investigation, it was confirmed that the lake was slightly enriched in phosphorus based on regional standards (OBVD Citation2011). However, because of the large gap in the data, only poor insights existed into how the lake conditions evolved over recent years in relation to human development and activities in its catchment. Moreover, nothing was known about its natural variability. It was thus difficult to determine whether modifications made to the wastewater treatment plant would be sufficient to significantly enhance the water quality of Lac Carheil or whether other actions would have to be undertaken, for example. Within this context, we initiated a paleolimnological investigation based on a multiproxy analysis of sediment cores collected from the lake. Our main objective was to fill the knowledge gap about the past evolution of this water body, to help the decision-making process of local water protection organizations in their preservation and restoration efforts. Effective management of aquatic ecosystems requires a good understanding of their evolutive pathway (i.e., long-term data), as this provides crucial insights into the causes and extent of their deterioration and helps predict future trajectories (Pienitz and Vincent Citation2003). Paleolimnology represents an efficient method to reconstruct past water quality of lake ecosystems where historical monitoring data are scarce or nonexistent (Smol Citation1992). Along with various physical and geochemical analyses, we used fossil diatoms (class Bacillariophyceae) as the main proxy to study past limnological and ecological changes in Lac Carheil. Diatoms are microscopic siliceous algae commonly used to track variations in water quality and habitat availability, and they are especially useful indicators of lake eutrophication (Hall and Smol Citation2010).
In addition to providing important background data for the management of Lac Carheil, our work contributes to the body of knowledge on the impacts of human development and sewage on boreal subarctic lake ecosystems needed to guide future sustainable and eco-responsible development of the North. We also document for the first time the long-term limnological and sedimentological history of a poorly studied region of subarctic Quebec. Such basic knowledge is particularly important in the context of the rapid transformation of northern aquatic landscapes due to climate change (Rouse et al. Citation1997, AMAP Citation2017).
Study area
Lac Carheil (52°43′40′′N; 67°06′20′′W) is located at 576 m elevation above sea level (a.s.l.), 6 km south of the town of Fermont near the border of Quebec and Labrador in eastern subarctic Quebec (). It is part of a chain of 3 lakes including also Lac Daviault and Lac Sans Nom (recently named Lac des Huards), located further upstream. Municipal wastewater is discharged into a small stream at the outflow of Lac Daviault, and then travels through Lac Sans Nom before reaching Lac Carheil. The shores of Lac Carheil are poorly developed with only a few residences present around the lake. A sandpit is also located near its northern edge. The lake is composed of 2 deep elongated basins. Our study focused on its northern part, which is ∼6.5 km long and has a maximum depth of ∼49 m. At the time of sampling, Lac Carheil had relatively clear (Secchi: 4.0 m) and well-oxygenated waters (DOsurface: 102.2%) with a circumneutral pH (7.5), and a low conductivity (38 μS/cm; ). The waters also had a light brown stain, suggesting notable concentrations of dissolved organic carbon. The lake was weakly stratified, and the mixing depth of the surficial water layers extended to ∼4 m depth (, S1).
Figure 1. Map of the study area in eastern subarctic Quebec and bathymetric contours (in meters) of the northern basin of Lac Carheil (modified from MRN Citation1977).
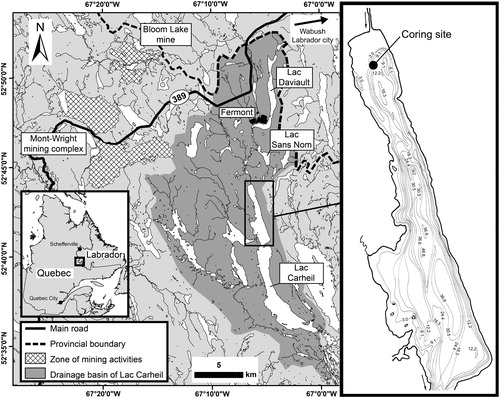
Table 1. Modern and historical limnological data from Lac Carheil’s waters.
The Fermont region is historically part of a hunting ground used by Indigenous Peoples, although their occupation of our study site has likely only been sporadic. Mining exploration in the area began around 1950. It later led to the creation of the Mont-Wright mining complex and the municipality of Fermont, which were built between 1971 and 1974 (Neal Citation2000). The mine is located 15 km west of the town, outside the watershed of Lac Carheil. Further north, another mining complex, the Bloom Lake iron ore mine, was created later in 2010 yet closed only a few years afterward in 2014. Two other mining towns built in the early 1960s, Labrador City and Wabush, are located 20 km northeast of Fermont in Newfoundland and Labrador. The population of Fermont grew quickly to a maximum of 4216 inhabitants in 1981, but has since progressively declined to 2474 in 2016 (Statistics Canada Citation1982, Citation2017). The seasonal population of the town is estimated to be very low and has remained stable over the years.
The Fermont region is part of the parautochthonous belt in the Grenville geological province of the Canadian Shield (Rivers et al. Citation1989). It is also located in the southern part of the Labrador Trough, rich in iron ore deposits (Neal Citation2000). The bedrock is mainly composed of gneisses, schists, and metasedimentary iron units, including small amounts of carbonates that are overlain by younger layers of gabbros, granites, syenites, migmatites, and amphibolites. These are covered by Pleistocene-age and recent deposits of clay, sand, gravel, and erratic boulders of the last Wisconsinan glaciation (Clarke Citation1960, Murphy Citation1960). Deglaciation of the Laurentide Ice Sheet occurred around 6.8–7.5 ka BP (Occhietti et al. Citation2011). Regional topography is characterized by a rugged (hilly) high plateau terrain situated at elevations between 570 and 700 m a.s.l.
According to climate normals compiled from the Wabush Lake A station (1981–2010; located 25 km northeast of Fermont), mean annual air temperature is –3.1 C, total precipitation is 839.5 mm (rain: 502.9 mm; snow: 336.6 mm), and the freezing period extends from mid September to early June (ECCC Citation2018). Lac Carheil is covered by ice from approximatively mid December to early June. Forest cover in the region is open spruce woodland with high abundance of lichens. Dominant plant species include black spruce (Picea mariana), dwarf birch (Betula glandulosa), planeleaf willow (Salix planifolia), northern blueberry (Vaccinium boreale), brownish sedge (Carex brunnescens), and Canadian burnet (Sanguisorba canadensis; Blondeau and Dignard Citation2001). Soils are generally poorly drained with abundant wetlands, mostly peatlands. The region is located within the sporadic permafrost zone of northern Quebec (<2% of the territory; Allard and Séguin Citation1987).
Materials and methods
Core collection and sediment subsampling
In June 2015, 2 sediment cores, 34.5 and 55.5 cm long, were extracted at approximatively the same location in Lac Carheil using an Aquatic Research percussion gravity corer equipped with ∼6.7 cm inner diameter plastic coring tubes (). The shorter core (CAR-1) had a well-preserved water-sediment interface, whereas the longer one (CAR-2) had a disturbed interface but provided a longer sedimentary sequence. Coring was done close to the site where the cyanobacterial blooms were previously observed (depth: 10.2 m). The sediment surface layers of the 2 cores were stabilized in the field using Zorbitrol (Tomkins et al. Citation2008) prior to transport.
In the laboratory, the coring tubes were cut along their longitudinal axis using a Dremel saw and sediments were split in 2 halves with a length of fishing line. One half of each core was then transversely subsampled at 0.5 cm intervals, whereas the other halves were kept intact for nondestructive geochemical X-ray microfluorescence (μ-XRF) analyses. The analyzed half-cores were then stored in a cold room (∼4 C). All sediment subsamples were freeze-dried during a 48 to 72 h time period before subsequent analyses. Water content (percent water) of the sediment was estimated by weighing the samples before and after the drying process.
Core correlation
Both cores were submitted to μ-XRF and loss-on-ignition (LOI) analyses. Results obtained were then used along with percent water data and visual clues to correlate the sedimentary sequences of the cores to create a composite core stratigraphy (i.e., the bottom section of the long core was added to the short core). All subsequent analyses, including radiometric dating as well as grain-size, diatom, and C-N analyses, were then processed on this composite core, instead of on the entire short and long cores, to reduce laboratory time and costs.
Sediment samples were generally analyzed at 0.5 cm (LOI) or 1 cm (grain-size, C-N) intervals. Diatom analyses were executed at 0.5 cm intervals for the upper 10 cm of the composite core, at 1 cm intervals between 10 and 30 cm depth, and at 2 cm intervals for the rest of the core.
Dating and chronology
Samples of 1.0 g of dry sediment from various stratigraphic levels of the upper part of the composite core were submitted to the Laboratoire de Radiochronologie of the Centre d’études nordiques (CEN) at Université Laval (Québec City, Canada), where their 210Pb (total), cesium-137 (137Cs), and radium-226 (226Ra) activities were measured using a high-purity germanium (HPGe) detector. The constant rate of supply model (CRS) was applied on the unsupported 210Pb concentrations to establish the chronology of recent sediments (<150 yr; Appleby Citation2001).
Two wood and 2 bulk sediment samples were also extracted from different stratigraphic levels and dated using carbon-14 (14C). Samples were submitted to the Laboratoire de Radiochronologie (CEN), where they were pretreated, when possible, using hydrochloric acid (HCl) and sodium hydroxide (NaOH), and then transformed into carbon dioxide (CO2) before being sent to an external laboratory (Keck Carbon Cycle AMS Facility, University of California, Irvine) for accelerator mass spectrometry (AMS) dating. All radiocarbon dates were corrected for total isotopic fraction and calibrated according to the IntCal13 dataset using the program CALIB 7.1 (Stuiver et al. Citation2017).
μ-XRF
The μ-XRF analyses of the 2 complete half-cores were conducted at the Centre Eau Terre Environnement (ETE) of Institut national de la recherche scientifique (INRS; Québec City, Canada) using an ITRAX Core Scanner system. The latter performs high resolution nondestructive analyses that provide relative variations of chemical element concentrations in sediment cores. Half-cores were scanned at a downcore resolution of 1 mm with an exposure time of 20 s per increment using a Cr-anode X-ray tube, which allows a better detection of lighter chemical elements (Croudace et al. Citation2006). Results obtained from the XRF measurements were generally expressed as ratios of the detrital element titanium (Ti) to provide a better detection of geochemical variations unrelated to simple changes in background sedimentation and diagenesis (e.g., Antoniades et al. Citation2011). Ti is mostly found in hard resistant minerals and is therefore less prone to weathering than other naturally occurring elements (Force Citation1991). Results for Ti were normalized by the total counts per second (cps) of the XRF spectrum to take into account the heterogenic nature of the sediment matrix.
Optical images and microradiographs of the analyzed half cores were also acquired using the ITRAX Core Scanner. Radiographs (pixels: 0.1 × 0.025 mm) mainly reflect variations in the sediment bulk density, with darker and lighter gray levels respectively representing higher and lower density.
Loss on ignition
Subsamples of 0.20–0.25 g of dry sediment were placed in a laboratory oven for 24 h at 105 C to eliminate residual humidity. Samples were then weighed and placed into a muffle furnace at 550 C for 4 h, following recommendations by Heiri et al. (Citation2001). The organic matter content of the sediments (% OM), which was lost during this last combustion step, was then estimated by weighing the remaining samples.
Grain size
Grain-size analyses were performed on the loss-on-ignition residues at the Laboratoire de géomorphologie et de sédimentologie of Université Laval. In order to deflocculate the sediment grains, a sodium hexametaphosphate solution (10%) was first added to the samples 24 h before proceeding to size measurements. They were then analyzed by laser diffraction using a Horiba particle-size analyzer. The mean grain size of each sample was calculated according to the arithmetic method of moments using the GRADISTAT program, version 8.0 (Blott Citation2010).
C-N
C-N analyses were performed on 0.10 g subsamples of dry sediment submitted to the Laboratoire de Radiochronologie (CEN). Carbon (% C) and nitrogen (% N) concentrations were measured using a LECO CHN628 elemental analyzer.
Diatoms
In the Laboratoire de Paléoécologie Aquatique (LPA) at Université Laval, 50 mg subsamples of dry sediment were transferred into glass vials along with a few milliliters of 10% HCl solution to remove potential traces of carbonates. Samples were rinsed using distilled water 24 h later and a mixture of sulfuric (H2SO4) and nitric (HNO3) acids was then added to the vials to eliminate organic material. The following day, the vials were placed into a hot water bath (∼60 C) for 2 h to accelerate the digestive reaction. Samples were afterward rinsed several times using distilled water until acid residues were removed. The slurries were then diluted and mixed with synthetic microspheres to assess diatom concentrations in the samples (Battarbee and Kneen Citation1982). Resulting suspensions were then dried at room temperature on ethanol-cleaned glass coverslips, which were later mounted onto microscope slides using the high-refractive-index mounting medium Naphrax.
For each sample, at least 400 diatom valves were identified and enumerated along random transects using a Leica DMRB optical microscope set at 1000× magnification. Identification was carried out at the finest taxonomic level possible, although varieties or morphotypes of some species were later grouped together for data analysis (hereafter referred to as taxonomic complexes). Diatom identification was primarily based on floras published by Krammer and Lange-Bertalot (Citation1986, Citation1988, Citation1991a,Citationb), Fallu et al. (Citation2000), and Spaulding et al. (Citation2018), but many other taxonomic references were also consulted.
The relative abundance (% of total count) of each diatom taxon (or taxonomic complex) was calculated and the most abundant taxa were plotted in a diagram using C2, version 1.7.6 (Juggins Citation2014). Biostratigraphic zones were then objectively delimited by testing several statistical splitting (binary, optimal) and agglomerative (CONISS, CONIIC) procedures with sums of squares or information content as the dissimilarity measure. Zonation calculations were based on diatom taxa presenting a relative abundance ≥1% in at least one sample to reduce noise in the dataset. Data were square-root transformed to downweigh the statistical influence of dominant taxa. Zonation computing was done using the PSIMPOLL program, version 4.27 (Bennett Citation2009). The number of significant diatom zones was validated using the broken-stick model (Bennett Citation1996). A principal component analysis (PCA) was also performed on the square-root data, and the axis 1 sample scores were then plotted into the stratigraphic diagram to illustrate the changes in the entire diatom assemblages.
In order to reconstruct changes in diatom-inferred total phosphorus (TP) concentrations, the fossil diatom assemblages from the composite sediment core were compared to modern diatom assemblages of 55 southern Quebec lakes characterized by a broad range of trophic states (Tremblay et al. Citation2014). This dataset was selected because of the geographical proximity between Fermont and southern Quebec lakes and their similar geological context. The degree of resemblance between the fossil and modern diatom assemblages was evaluated. Calculations were done with the C2 program using the Modern Analog Technique (MAT) on diatom taxa with an occurrence in ≥4 lakes in the southern Quebec dataset, and a relative abundance ≥1% in at least one sample of the composite core. The 75th and 95th percentiles of the dissimilarity coefficient distribution among southern Quebec lakes were used to define confidence intervals, and average dissimilarities between southern Quebec lakes and Lac Carheil were calculated for each analyzed stratigraphic level. According to similar criteria used in previous studies (e.g., Laing et al. Citation1999, Köster et al. Citation2005), levels with a dissimilarity average value below the 75% confidence interval may be considered good analogs of a reference dataset, whereas levels between the 75 and 95% intervals are weak analogs, and those exceeding the 95% interval express no similarity.
Historical water TP concentrations of Lac Carheil were reconstructed with the inference models developed by Tremblay et al. (Citation2014) using diatom taxa with an occurrence in ≥4 lakes in the southern Quebec dataset and based on weighted average regression analysis with classical deshrinking (classic WA) and weighted averaging partial least squares regression (WA-PLS). Transfer functions resulting from these models were applied on the fossil diatom data (≥1% in at least one sample) of Lac Carheil. Relative abundances were previously Hellinger transformed to be consistent with the procedure followed by Tremblay et al. (Citation2014) for the development of the inference models. Computing was done using R software, version 3.3.2 (R Core Team Citation2016).
Results
General stratigraphy
The core correlation revealed that the upper 5 cm of the longer core had been lost during coring. The composite core therefore had a total length of 60.5 cm, consisting of the entire 34.5 cm sequence of the shorter core and the bottom 26 cm of the longer core (). Most of its sedimentary sequence (60.5–10.5 cm) was characterized by different tones of light brown sediments repeatedly interrupted by black laminae, which had a significantly higher water content and mean grain size than the main sediment matrix (see Supplementary Material). Traces of oxidation (orange stains) were observed from 10.5 to 7.75 cm. The sediments were then more homogeneous toward the surface, with color changing to brown-olive from 7.75 to 2.5 cm and to dark brown from 2.5 to 0.0 cm.
Figure 2. Optical photographs of the sediment cores retrieved from the bottom of Lac Carheil with the resulting composite core.

Dating and chronology
The unsupported 210Pb concentrations in the upper part of the composite core followed an exponential decay curve marked by a few peaks and troughs likely related to past changes in sediment accumulation rates (). The 137C activity was very low in the core and no clear peak corresponding to the 1963 maximum global fallout from nuclear weapons testing could be identified. The CRS chronology could therefore not be validated using this isotope. Instead, its accuracy was tested by calculating the depth where the inventory of 210Pb was reduced by 90% compared to the surface value, which should be ∼74 yr old (Appleby and Oldfield Citation1990, Sorgente et al. Citation1999). In the composite core, this level approximately corresponded (by interpolation) to 7.9 cm, which had an estimated age of 76 yr, thus suggesting that the model was reliable. Moreover, the 14C dating of a wood piece retrieved from the 13.0–13.5 cm level indicated a rounded median probability age of 1840 yr cal. AD (), which is consistent with an extrapolation of the curve of the CRS chronology to this depth ().
Figure 3. (A) Unsupported 210Pb decay curve and 137Cs activity in the upper part of the composite core (CAR-1; 0–34.5 cm); (B) resulting age–depth curve of the uppermost section (0–12 cm) based on the application of the CRS model; and (C) estimated chronological curve for the entire composite core (CAR-1 + CAR-2; 0–60.5 cm) based on 210Pb and 14C dating.
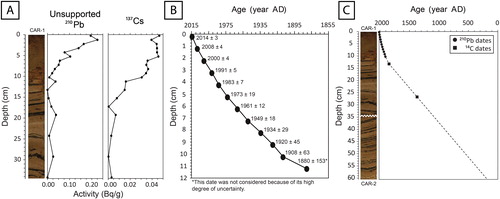
Table 2. Samples dated using 14C AMS in the composite core with their respective radiocarbon and calendar ages.
Results from 210Pb dating indicated that the period of modern settlement in the catchment (i.e., 1971–2015) was contained in the upper 5.5 cm of the core (). Another piece of wood from the 26.5–27.0 cm level was dated to a rounded median probability age of 1360 yr cal. AD. Further 14C dating of 2 bulk sediment samples yielded suspiciously old dates (in one case, older than the period of ice retreat), which were subsequently eliminated. By extrapolating the results of the 210Pb and 14C dating, we estimated that the complete composite core covered approximatively the past 1800 yr in the history of Lac Carheil ().
Microradiographs
Microradiographs obtained of the composite sediment core showed a general trend from darker to lighter tones (i.e., a decreasing density) from the bottom to the top, attributed to the sediment compaction effect (). However, many abrupt variations were observed at several levels that were associated with changes in sediment color. Although variable, the black laminae were generally represented by lighter tones on the radiographs and were thus characterized by lower bulk densities compared to the main sediment matrix.
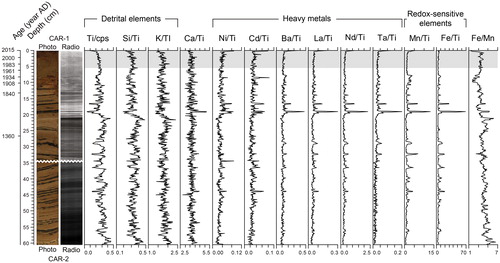
Figure 4. Microradiographs (radio) and relative concentration stratigraphic profiles of the selected elements obtained from the μ-XRF analyses. The gray shaded area corresponds to the period of modern settlement in the catchment (i.e., 1971–2015).
XRF elemental concentrations
Concentration data were acquired for 28 chemical elements as detected in the composite core using the Itrax Core Scanner (). However, many displayed noisy profiles, likely due to low concentrations near or below the detection limit, and were therefore not considered. All others were kept for discussion and included elements representative of the detrital phase (Ti; potassium, K; silicon; Si) and carbonate fraction (calcium; Ca) of the sediments, heavy toxic (nickel, Ni; cadmium, Cd) and nontoxic (barium, Ba; lanthanum, La; neodymium, Nd; tantalum, Ta) metals, and redox-sensitive elements (iron, Fe; manganese, Mn). An important proportion of Si in lake sediments may not only be derived from allochthonous sources but may also come from autochthonous contributions (Peinerud Citation2000). Here, Si and K were however strongly correlated (r = 0.79; p < 0.001), suggesting that changes in Si mainly followed variations in the detrital exports to Lac Carheil.
The variations in the concentration of the selected elements indicated a few important stratigraphic changes (). In particular, the values of the detrital element ratios Ti/cps, Si/Ti, and K/Ti were generally higher from 60.5 to 20.0 cm and then lower between 20.0 and 0 cm (∼1600–2015 AD). These differences were significant according to nonparametric Mann–Whitney (MW) U-tests (p < 0.001). The Ca/Ti ratio displayed a relatively flat profile, indicating that the calcium inputs followed background changes in sedimentation and diagenesis. The most striking results came from the black laminae, which corresponded with higher concentrations of heavy metals and, more importantly, redox-sensitive elements, as displayed by the associated increases in the values of the ratios Ni/Ti, Cd/Ti, Ba/Ti, La/Ti, Nd/Ti, Ta/Ti, Mn/Ti, and Fe/Ti. Mn and Fe concentrations were also higher in the 10.5–7.75 cm layer (∼1900–1940 AD), where traces of oxidation were observed. The black laminae were often associated with peaks in the Fe/Mn values (although not always), whereas the 10.5–7.75 cm layer was related to lower ones. Other changes in the sediment color could also be linked to small variations in the Fe/Mn ratio. No variations in elemental concentrations were associated with the period of modern settlement in the catchment.
Water and organic matter
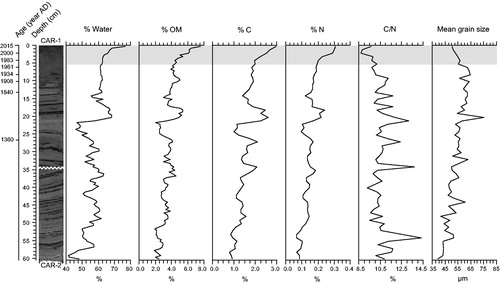
The sediments from the composite core had an average water content of 41–77% and were poor in OM (1.9–7.6%; ). Important stratigraphic variations were nonetheless observed in the values of the percent water and % OM variables. Their profiles indicated a general trend toward higher values from the bottom to the top, with a more pronounced increase in the upper 5.5 cm corresponding with the period of modern settlement in the catchment. Maximum percentage values were reached at the surface. Other notable variations included a drop in the percent water and % OM values between 26.0 and 21.0 cm (∼1390–1560 AD), followed by an important increase between 21.0 and 17.0 cm (∼1560–1710 AD).
Figure 5. Stratigraphic variation profiles of water (% water), organic matter (% OM), carbon (% C) and nitrogen (% N) concentrations, carbon to nitrogen ratio (C/N), and mean grain size within the composite core. The gray shaded area corresponds to the period of modern settlement in the catchment (i.e., 1971–2015).
Carbon, nitrogen, and C/N
Carbon and nitrogen concentrations followed the same trends in the composite sediment core as the water content and organic matter as estimated by LOI (). From the bottom to the top, values varied between 0.8 and 3.0% for C and between 0.07 and 0.31% for N. Maximum values were reached at the surface following a progressive increase in the upper 5.5 cm corresponding with the period of modern settlement in the catchment. Here again, notable variations included a drop in the % C and % N values between 26.0 and 21.0 cm (∼1390–1560 AD) and an important increase between 21.0 and 17.0 cm (∼1560–1710 AD).
The carbon to nitrogen ratio (C/N) of the sediments in the composite core varied between 8.7 and 14.3 with a mean value of 10.6 (). Important peaks were observed at various levels throughout the lower part of the core (60.5–10.5 cm). The 10.5–7.75 cm oxidized layer (∼1900–1940 AD) also corresponded with higher C/N values (10.1–11.2). The values of the ratio were characterized by a brief drop between the levels 3.0 and 0.0 cm (8.7–9.7) associated with the recent history of the lake (∼1995–2015 AD).
Grain size
The analysis of the grain size data using the GRADISTAT program revealed that the sediments from the composite core consisted of a mix of fine sand and coarse silt. Throughout the whole sedimentary sequence, the mean grain size values varied between 40 and 75 μm (). They showed a general increasing trend from 60.5 to 5.5 cm and then slightly decreased in the upper 5.5 cm, corresponding with the period of modern settlement in the catchment.
Diatom assemblages
In total, 364 diatom taxa belonging to 66 genera were identified in the sediments of Lac Carheil. The assemblages preserved in the composite core were dominated by benthic taxa of mainly small fragilarioid groups. The zonation analyses indicated 2 statistically significant biostratigraphic zones. The depth value separating the zones varied according to the method used. Agglomerative and splitting procedures with sums of squares as the dissimilarity measure all indicated the level 13.25 cm as the separation depth, whereas procedures based on information content all yielded different results (13.25, 14.25, 20.25 cm). As a greater consensus existed around the level 13.25 cm (1840 AD), this latter was retained to delimitate the zones. The zonation was in agreement with the profile of the PCA axis 1 scores (λ = 87%), which showed an increasing trend from 60.5 to 13.25 cm before displaying a directional change toward lower values from 13.25 to 0.0 cm ().
Figure 6. Stratigraphic relative abundance variation profiles of the most common diatoms (≥2.5% in at least one level, except for Stephanodiscus minutulus) preserved in the composite core along with the profile of the PCA axis 1 sample scores summarizing the stratigraphic variations in the composition of the whole diatom assemblages. The black line (1840 AD) delimits the 2 statistically distinct diatom zones, whereas the gray shaded area marks the period of modern settlement in the catchment (i.e., 1971–2015). Note. The Staurosirella pinnata complex includes S. pinnata (mainly), S. pinnata var. acuminata, and S. pinnata var. intercedens; the Pseudostaurosira brevistriata complex includes P. brevistriata (mainly), P. brevistriata var. papillosa, and P. brevistriata var. elliptica; and the Achnanthidium minutissimum complex includes capitate and noncapitate forms of A. minutissimum.
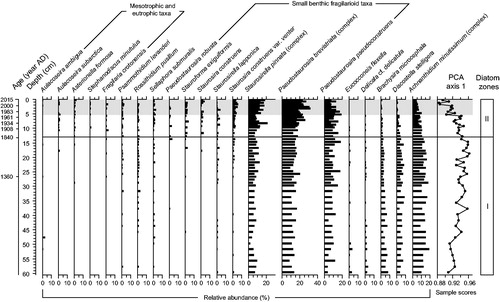
The first zone (60.5–13.25 cm) was mainly dominated by small benthic fragilarioid taxa belonging to the Staurosirella pinnata and Pseudostaurosira brevistriata complexes and to Pseudostaurosira pseudoconstruens (). Altogether, these taxa generally accounted for ∼30% of total diatom assemblages. Important taxa in zone I also included Discostella stelligera and the Achnanthidium minutissimum complex, which averaged relative abundances of 6% and 14%, respectively. The relative abundance of the species Brachysira microcephala declined progressively from the lower to the upper limits of this zone with values ranging between 9% and 2%.
The relative abundance of several small benthic fragilaroid taxa increased even more in the second zone (13.25–0.0 cm). The relative abundances of the S. pinnata and P. brevistriata complexes were higher, especially in the upper 5.5 cm, corresponding with the period of modern settlement in the catchment, where they reached maximum values of 21% and 32%, respectively. Several fragilaroid taxa that were rare in zone I were more frequent and abundant in zone II, such as Pseudostaurosira robusta, Staurosirella lapponica, Stauroforma exiguiformis, Staurosira construens, and Staurosira construens var. venter. The occurrence and relative abundance of a few taxa typical of enriched waters (Aulacoseira subarctica, Stephanodiscus minutulus, Fragilaria crotonensis, and Asterionella formosa) also increased in this zone, although they collectively never exceeded 7% of total diatom counts. In contrast, the relative abundances of the species D. stelligera and of the A. minutissimum complex progressively decreased from 13.25 to 0.0 cm. This decline was more pronounced in the upper 5.5 cm. In this depth interval, D. stelligera almost disappeared from the assemblages (average relative abundance of 1%), whereas the A. minutissimum complex reached a relative abundance minimum value of 3%. The relative abundance of the species B. microcephala continued to decline in zone II following the trend previously observed from 60.5 to 13.25 cm.
Diatom abundance and reconstructed TP concentrations
The diatom absolute abundance showed important variations in the composite core (). The valve concentration was more or less stable from 60.5 to 5.5 cm around a mean value of 6.11 × 108 valves per gram of dry matter (DM). However, it was significantly higher (MW U-test, p < 0.001) in the upper 5.5 cm, corresponding with the period of modern settlement in the catchment (mean: 11.43 × 108 valves/g DM).
Figure 7. Stratigraphic variation profiles of the diatom absolute concentration and diatom-inferred total phosphorus (DI-TP) concentrations in the composite core. The black line (1840 AD) delimits the 2 statistically distinct diatom zones, whereas the gray shaded area marks the period of modern settlement in the catchment (i.e., 1971–2015).
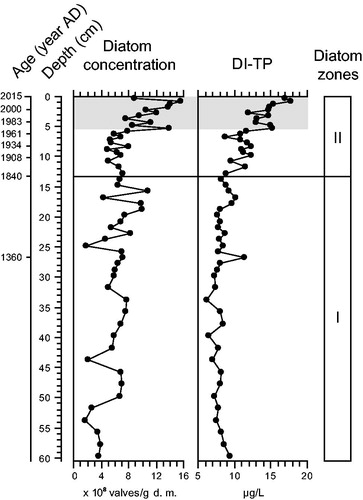
The analog technique performed on the fossil diatom assemblages of Lac Carheil and the modern diatom assemblages of southern Quebec lakes revealed that all analyzed levels of the composite core had dissimilarity average values lower than the 75% confidence interval. The 2 datasets were thus similar and comparable. The inference models developed by Tremblay et al. (Citation2014), based on classic WA and WA-PLS analyses, allowed for reconstructions of historical TP concentrations ranging between 6 and 19 μg/L in Lac Carheil. For this range of values, Tremblay et al. (Citation2014) determined that the biases of the classic WA and WA-PLS models were often opposite. Similar to the approach adopted by Tremblay (Citation2015) to reconstruct certain historical TP concentrations, averages of the values obtained from both models were therefore calculated for each analyzed level and used for discussion.
From 60.5 to 13.25 cm, the diatom-inferred total phosphorus (DI-TP) concentrations were relatively stable in the composite core with a mean value of 8.2 ± 1.9 μg/L (). The reconstructed concentrations were then higher from 13.25 to 5.5 cm (mean: 10.9 ± 1.9 μg/L) and furthermore increased in the upper 5.5 cm, corresponding with the period of modern settlement in the catchment (mean: 14.8 ± 1.9 μg/L), where a maximum value of 17.8 ± 1.9 μg/L was reached near the surface. A nonparametric Kruskal–Wallis test indicated that the means were significantly different (p < 0.001) between these 3 depth intervals.
Discussion
Past sedimentological activity
The core section comprising the black laminae that marks the stratigraphy of Lac Carheil illustrates past changes in the nature of the clastic material deposited at the bottom of the lake. The laminae had organic matter content (% OM) and C/N signatures ( S2) that were similar to the main sediment matrix and were not associated with peaks in the concentration of detrital elements. This suggests that they did not correspond to periods of increased water primary production or enhanced erosion in the catchment. However, the black laminae were characterized by higher concentrations of Fe, Mn, and heavy metals, which indicates a distinct mineralogical composition. Variations in the concentrations of Fe and Mn in lake sediments are often linked to changes in the hypolimnetic redox conditions. However, most peaks in the concentrations of these elements associated with the black laminae also corresponded with peaks in the Fe/Mn ratio, suggesting that the variations that we observed were rather due to changes in the external supply of these elements (Engstrom and Wright Citation1984). We interpret their high concentrations in the black laminae as a sign of high abundance of clastic iron and manganese oxide minerals in the sediments (e.g., magnetite, hematite, manganite). Many of these minerals have a dark (black) color and can be abundant in the Labrador Trough (Neal Citation2000). Observations of both pale and black sand deposits on the beaches of Lac Carheil and other lakes around Fermont clearly demonstrate that the erosion of the bedrock in this region provides 2 different types of naturally segregated sediments ( S3). The black sands are strongly attracted to magnets, which suggest they have a high iron content and are therefore similar and have the same origin as the black laminae in our sediment cores. However, the depositional mechanisms of this material at our coring site are unclear. The coarser grain size of the black laminae ( S2) suggests that they were formed under conditions of higher hydrological energy (e.g., Campbell et al. Citation2000). It is possible that the material from the black laminae has only been transported to and deposited at our coring site during periods of strong water flow, for example, during important spring floods or after storm events, whereas it is usually transported only over shorter distances. Alternatively, the material could originate from underwater landslides in shallower zones of the lake. The fact that we did not observe black laminae in the upper 10 cm of our composite sediment core may indicate that the frequency of the high-energy events that led to their formation was greatly reduced or even nonexistent in recent times.
The gradual increase in mean grain size from 60.5 to 5.5 cm could be attributable to changes in the hydrological conditions of the main inflow feeding Lac Carheil at its northern edge. It is possible that the morphometry of the river has progressively changed over time, resulting in higher water velocities transporting coarser material to our coring site. We could also consider that an increase in mean grain size may be indicative of enhanced ice erosion, wind activity, storminess, or precipitation, for example. However, we believe that such parameters are more likely to vary on short time scales or during temporary periods, whereas the transition toward coarser grains in our composite sediment core is gradual and continuous over a long period of time (∼1750 years). Moreover, the increase in mean grain size was not correlated to changes in our geochemical proxies, suggesting that it is not associated with an enhanced erosional activity in the catchment or higher delivery of allochthonous material.
Limnological background (preindustrial period)
Prior to the 19th century, paleolimnological evidence indicates that the limnological conditions of Lac Carheil have been relatively stable for hundreds of years. The physical and geochemical proxies detected minor changes in the sediment characteristics associated with different periods of time that could be linked to past minor variations in erosional activity in the lake catchment and/or allochthonous/autochthonous primary production. Likewise, the composition of the diatom assemblages remained fairly constant, suggesting that no important changes in the water quality or lake characteristics occurred. The assemblages were typical of cold nutrient-poor waters, and the diatom-phosphorus reconstruction models indicated natural background TP concentrations of ∼8 μg/L. However, we think that the reconstructed values are probably overestimated, as the minimum DI-TP concentration calculated in the composite core (6 μg/L) was higher than instrumental measurements completed in October 2011 and July 2012 (). This could be attributed to the dominance of small benthic fragilarioid species in the sediments of Lac Carheil which have TP optima varying around 10 μg/L in southern Quebec lakes (Tremblay Citation2015). Such taxa tend be more abundant in arctic and subarctic lakes than in temperate lakes, due to colder temperatures and more prolonged ice cover (e.g., Lotter and Bigler Citation2000). Therefore, they generally do not occur in the same proportion in the diatom assemblages of these 2 different environments, and we think this may have cause a bias in the inferred TP concentrations.
The decrease in the relative abundance of B. microcephala in the preindustrial sediments of Lac Carheil could indicate a gradual loss of plant microhabitats in the littoral zone of the lake, as this species has a documented preference for epiphytic substrates and habitats (e.g., Cellamare et al. Citation2012). Although not shown in our results section, we also noted that other epiphytic and moss-associated taxa (e.g., Eunotia spp., Pinnularia spp.) were also slightly more abundant in the bottom part of our composite core. These changes might be related to the concurrent increase in the mean grain size of the sediments. Coarser grains can constrain macrophyte growth (Madsen et al. Citation2001). Inversely, reduced macrophyte abundance in the littoral zone of a lake can also result in the transport of materials with a higher grain size to deeper parts of the water body (Carpenter and Lodge Citation1986).
First period of limnological changes (ca. 1840–1970 AD)
The sediment archives revealed that the limnological conditions of Lac Carheil first began to change significantly near the middle of the 19th century (ca. 1840 AD). This was mainly evidenced by variations in the composition of the diatom assemblages that progressively took place in the sediment layers corresponding to this time period, including (1) the decline of the species D. stelligera; (2) the rise of nutrient-loving taxa (A. subarctica, F. crotonensis, and A. formosa); and (3) the increase in the DI-TP concentrations, which may collectively be interpreted as signs of early water enrichment and eutrophication. Discostella stelligera has a low TP optimum in lakes of southern Quebec (Tremblay Citation2015), and several studies have observed a decrease in its relative abundance in a context of nutrient enrichment (e.g., Clerk et al. Citation2000, Köster et al. Citation2005). Similarly, A. minutissimum is also sometimes seen as an indicator of low nutrient levels in lakes (e.g., Kernan et al. Citation2009), but its ecological preferences remain ambiguous (Potapova and Hamilton Citation2007). Changes in the relative abundance of A. minutissimum may also depend on macrophyte richness (e.g., Garrison and Wakeman Citation2000). Inversely, some small fragilarioid taxa that increased between 1840 and 1970 AD may be favored by nutrient enrichment, such as S. exiguiformis, which can be associated with increased lake productivity (e.g., Buczkó et al. Citation2009).
Alternatively, we also considered that the variations in the diatom assemblages could indicate that the average water mixing depth at Lac Carheil has been increasing since the mid 1800s. Indeed, D. stelligera is also known to be favored by shallow mixing depths and may be indicative of changes in the thermal structure of lakes (Saros et al. Citation2012, Citation2016). Its decrease in the sediments of Lac Carheil was consistent with the concurrent increase in the relative abundance of a few Aulacoseira species, such as A. subarctica, which inversely thrives in more turbulent waters because of its heavily silicified frustule and high sinking rate (Gibson et al. Citation2003). However, the present-day mixing depth of the lake (∼4–6 m; ) closely corresponds to the optimum value of 4 ± 2.5 m (mean ± SE) calculated for D. stelligera by Saros et al. (Citation2012), which suggests that this parameter may not have been the most important factor limiting its growth since the mid 1800s. Moreover, the hypothesis of increasing mixing depths is not in agreement with the data from local meteorological stations (1961–2015; Figures S4 and S5) and climate record from Labrador (1881-2011; Way and Viau Citation2015), which indicate trends toward increasing air temperatures and lower wind activity, which would more likely result in shallower mixing depths. In the following paragraphs, we explore various scenarios that could explain the early eutrophication of Lac Carheil between 1840 and 1970 AD, including the influence of Indigenous Peoples, forest fires, long-range transport of airborne nutrients, and increased air temperature and precipitation.
In some regions of North America, it has been documented that the traditional lifestyle of Indigenous Peoples contributed to an early enrichment of lakes prior to the arrival of European settlers (e.g., Ekdahl et al. Citation2007). As mentioned earlier in the text, Indigenous Peoples have certainly wandered the region of Lac Carheil at some time in the past. However, they are not known to have established permanent camps around the lake, and no sign of human disturbances in its catchment was reported prior to the establishment of the town of Fermont in the 1970s. Therefore, we doubt that Indigenous Peoples could be responsible for the early eutrophication of the lake.
At the time the town of Fermont was constructed, evidence of recent forest fires was reported (Dufresne Citation1975). Comparison of aerial photographs from the region also demonstrates that forest cover was sparse around Lac Carheil in 1949, and that it progressively expanded in the following decades ( S7). We thus considered that a massive forest fire may have occurred in the region around 1840 AD and that this contributed to the early enrichment of the lake. We can expect that a fire could have resulted in higher inputs of nutrients and ions in Lac Carheil following the burning of organic material and the loss of vegetation cover (e.g., McEachern et al. Citation2000). However, the changes in the diatom assemblages from this time period were not accompanied by significant variations in our geochemical proxies that would indicate an increased delivery of allochthonous material to the lake. Moreover, examples from the literature demonstrate that the impacts of forest fires on lakes are often small and/or only perceptible on a short time scale (e.g., Enache and Prairie Citation2000, Paterson et al. Citation1998, Citation2002).
It has been demonstrated that long-range transport of airborne nutrients from early stages of agriculture and industrialization have resulted in the early eutrophication of many lakes across the northern hemisphere (e.g., Bergström and Jansson Citation2006). Prior to 1950 AD, however, no industrial or agriculture centers were present in a 300 km radius of Fermont. Furthermore, the earliest evidence of human-induced atmospheric nitrogen deposition in sediments of remote North American lakes has been traced by Holtgrieve et al. (Citation2011) to ∼1895 AD, many decades after the limnological conditions of Lac Carheil began to change. Studies have also shown that the impacts of these airborne nutrients are usually more significant since the 1950s following the increased emissions from fossil fuels and the more extensive use of artificial fertilizers (e.g., Wolfe et al. Citation2003, Saros et al. Citation2011).
Evidence of increased air temperatures has been reported in many regions of North America since the mid 1800s (e.g., Rühland et al. Citation2003, Citation2008). It has been suggested that this warming has resulted in the eutrophication of water bodies from these regions by inducing changes in the internal cycling of nutrients (e.g., Moser et al. Citation2002). As previously mentioned, local meteorological data (1961–2015) show increasing air temperatures in the Fermont region, especially during the ice-free season (Figure S4). Although no data prior to 1961 exist at our study site, the adjacent region of Labrador is known to have experienced a warming trend during the past century with an overall increase of ∼1.5 C from 1881 to 2011 (Way and Viau Citation2015). It is possible that this temperature rise began near 1840 AD concurrently with the changes that we observed in the diatom assemblages. It has been demonstrated that climate warming may lead to an increase in phosphorus concentrations in the water column of lakes, especially in the case of those with long water retention time (Malmaeus et al. Citation2006). The water retention time at Lac Carheil is not precisely known but is estimated to be long (a few years). In such lakes, processes of diffusion from the sediments and mineralization may be expected to have a more important influence on the water chemistry. Phosphorus release from aerobic sediment surfaces is dependent on temperature and may increase significantly as a result of warming (Jensen and Andersen Citation1992). We can also expect that a higher summer temperature would lead to a stronger/longer stratification (i.e., higher stability of the water column) and could enhance sedimentation (i.e., deposition of nutrients) and reduce phosphorus export from lakes (via the outflow; Moser et al. Citation2002). Therefore, the increased temperature during the ice-free season in the Fermont region may have resulted in a higher availability of phosphorus for algal communities, especially during fall turnover following the transfer of nutrients from the hypolimnion to the surficial water layers of the lake. Another contributing factor may be that a warming of air temperatures could have contributed to denser vegetation and more developed soils in the catchment and thus increased nutrient fluxes to the lake. The recent emergence of the small fragilarioid taxa P. robusta, S. exiguiformis, S. construens, and S. construens var. venter may also have been directly favored by warmer waters (e.g., Joynt and Wolfe Citation2001, Solovieva et al. Citation2005, Finkelstein and Gajewski Citation2008).
Finally, local meteorological data (1961–2012) also show a trend toward increasing rain precipitation in the Fermont region that could have hypothetically originated near 1840 AD ( S6). We may expect that increased rainfall could have resulted in a greater in-wash of nutrients from the catchment and contributed to the early eutrophication of Lac Carheil. However, as previously discussed, the changes in the diatom assemblages from 1840 to 1970 AD were not accompanied by significant variations in our geochemical proxies that would indicate an enhanced erosion or leaching of soils caused by wetter conditions that could have resulted in the enrichment of the lake.
Among all the hypotheses that we explored in the preceding paragraphs, the increase in the regional air temperatures appears to be the most probable explanation for Lac Carheil’s early eutrophication. It could also have been the result of several of the possibilities enumerated in the preceding (e.g., combined effects of increased air temperatures and enrichment from airborne nutrients). Our current data and the lack of studies in this region of Quebec–Labrador prevent us from pinpointing the exact causes of the changes in the limnological conditions of the lake between 1840 and 1970 AD.
Second period of limnological changes (ca. 1971–2015 AD)
Although the natural condition of Lac Carheil had already been altered, the paleolimnological evidence demonstrated that the construction of the town of Fermont and the municipal wastewater discharges further modified its limnological state. Indeed, we consider (1) the increase in the C, N, and organic matter content of the sediments; (2) the decrease in the C/N ratio of the sediments (Meyers Citation2009); (3) the almost full disappearance of D. stelligera in the diatom assemblages; (4) the increase in DI-TP water concentrations; and (5) the increase in the diatom absolute abundance, which all corresponded with the period of modern settlement in the catchment, to be clear signs of an increased water primary production and of the accelerated eutrophication of Lac Carheil. The further increase in the relative abundance of small fragilarioid taxa may also be indicative of poorer water clarity caused by the higher abundance of phytoplankton in the water column, as such taxa have low light requirements (Lotter and Bigler Citation2000). Based on the OECD (Citation1982) classification of lakes, the DI-TP water values suggest that the lake has evolved toward a mesotrophic state in the last decades. As previously discussed, however, we think that the reconstructed concentrations are slightly overestimated as compared to the measured values. Considering that the mesotrophic or eutrophic diatom taxa preserved in the sediments of Lac Carheil never constituted a dominant part of the biological assemblages, we are convinced that lake waters never reached a mesotrophic state in the past. Over the last few decades, the lake likely evolved from an ultra-oligotrophic to an oligotrophic or oligo-mesotrophic state.
Although our paleolimnological proxies indicated that Lac Carheil suffers from cultural eutrophication, its disturbance level is weak. In comparison, Laperrière et al. (Citation2008) reconstructed DI-TP concentrations >100 μg/L (hypertrophic state) in Lac Dauriat at the culmination of mining activities and population density in Schefferville, close to our study site. The young age of the town of Fermont and the fact that a wastewater treatment plant was built early in its history probably limited water quality deterioration of Lac Carheil. Moreover, Lac Sans Nom, which is the first water body to receive the municipal wastewater before it drains downstream, certainly also acts as a buffer and reduces the amount of nutrients that reach our study lake. Because of its important size and depth and the resulting high water volume, the Lac Carheil basin also has an important dilution capacity.
Considering the low level of nutrient enrichment of Lac Carheil, the occurrence over recent years of harmful cyanobacterial blooms at its surface is surprising. We suggest that the blooms observed in the lake may have been washed in from upstream Lac Sans Nom, where they previously developed in its more enriched waters (OBVD Citation2011). Alternatively, the blooms may have grown in overflows either from the wastewater treatment plant or from Lac Sans Nom once having reached Lac Carheil. Municipal wastewater has a particularly low N/P ratio (e.g., Vallentyne Citation1974), which may promote the dominance of cyanobacterial communities in lake ecosystems (Smith Citation1983). In 1998, many new buildings were connected to the sewer system of Fermont. Although our study was not designed to infer past trends in cyanobacteria, we hypothesize that this event has resulted in increased wastewater discharges toward Lac Carheil, which favored the emergence of blooms a few years later (2005 to 2011).
Implications for lake management and recommendations
Given that the disturbance level of Lac Carheil is weak, its prospect for recovery is good. In this context and considering that the municipal wastewater discharges are the main source of nutrient enrichment to the lake, the recent improvements made to the treatment plant in 2010–2011 should be sufficient to significantly enhance the water quality of the lake. Phosphorus release from anoxic surficial sediments can pose a major obstacle to the recovery of eutrophic lakes (e.g., Jeppesen et al. Citation2005). Yet considering the low level of enrichment of Lac Carheil, this should not be a serious problem as its internal phosphorus load is likely small. Moreover, we also measured high dissolved oxygen levels (94.8%) in its hypolimnion (10.2 m) during our fieldwork ( S1). In fact, based on the TP measurements of 2011 and 2012, the health of Lac Carheil already seems to have improved following the reduction of nutrient inputs from municipal wastewater (). No cyanobacterial bloom has since been reported at the surface of the lake. Because the changes made to the wastewater treatment plant are recent and the sedimentation rate in Lac Carheil is low, we could not adequately evaluate whether its water quality did indeed improve over the last few years. We therefore recommend that a new limnological assessment be undertaken soon to better evaluate its present-day condition. In addition, an annual monitoring program should be established to allow a quick detection and response to water quality problems that could reappear in the future. Considering that Lac Carheil is fed by Lac Sans Nom, particular attention should also be paid to the health of this upstream lake.
In addition to direct anthropogenic disturbances, the limnological conditions of Lac Carheil have also been modified by uncontrollable environmental factors (i.e., potentially, increased air temperatures), which have initiated its early eutrophication near the middle of the 19th century. In this context, a complete return of the lake to its natural pristine state seems unrealistic. It is likely that these environmental changes made it more vulnerable to further nutrient enrichment and, eventually, harmful cyanobacterial blooms. Lake management should be done with the consciousness that the limnological conditions of Lac Carheil may further uncontrollably evolve in the context of global warming. Modern climate change can exacerbate the symptoms of eutrophication and the problems it causes (Jeppesen et al. Citation2010). In the future, it is expected to promote a global expansion of harmful cyanobacteria in aquatic ecosystems (O’Neil et al. Citation2012, Paerl and Paul Citation2012).
It is worth mentioning that the results that we obtained from our coring site in Lac Carheil may not be representative of overall changes in the entire lake basin, given its high surface area. The cores that we collected were taken near the inflow carrying wastewater from the town of Fermont and at a relatively shallow depth. The biological communities at this site are certainly more sensitive to the upstream anthropogenic disturbances and could also be more sensitive to climate change and its associated effects (e.g., variations in the mixing depth of the lake and the length/strength of stratification).
Conclusion
Our paleolimnological investigation confirmed that Lac Carheil has undergone some eutrophication. However, our results showed that its disturbance level was weak, which should facilitate the management efforts targeting its restoration and preservation. We also unexpectedly found that the limnological conditions of the lake began to change significantly many decades prior to modern settlement in its catchment. Further research is needed in this region of eastern subarctic Quebec to better evaluate the nature and the extent of the environmental and limnological changes that are currently occurring. As the arctic and subarctic regions are increasingly subject to human stress and cultural eutrophication, climate change poses a real challenge for lake management and should be taken into account in future action plans.
Supplemental Material
Download PDF (749.9 KB)Acknowledgements
We are grateful to the municipality of Fermont and the Centre d’études Nordiques (CEN) for their logistical support. Thanks also to Roxane Tremblay for sharing the data and scripts necessary to apply the diatom–phosphorus inference models, and to Dermot Antoniades and the current and past members of the Laboratoire de paléoécologie aquatique at Université Laval for their help in the laboratory and for sharing their opinions at all stages of this research project. Finally, we thank 2 anonymous reviewers and LRM’s associate editor A. Paterson for their comments and suggestions that helped to enhance the quality of this article.
Additional information
Funding
References
- Allard M, Séguin MK. 1987. Le pergélisol au Québec nordique: bilan et perspectives. GPQ. 411:141–152. doi:10.7202/032671ar.
- Antoniades D, Michelutti N, Quinlan R, Blais JM, Bonilla S, Douglas MSV, Pienitz R, Smol JP, Vincent WF. 2011. Cultural eutrophication, anoxia, and ecosystem recovery in Meretta Lake, High Arctic Canada. Limnol Oceanogr. 56(2):639–650. doi:10.4319/lo.2011.56.2.0639.
- Appleby PG. 2001. Chronostratigraphic techniques in recent sediments In: Last WM, Smol JP, editors. Tracking environmental change using lake sediments. Volume I: basin analysis, coring and chronological techniques. Dordrecht (Netherlands): Kluwer Academic Publishers. p. 171–203.
- Appleby PG, Oldfield F. 1990. Radioisotope studies of recent lake and reservoir sedimentation In: Crickmore MJ, Tazioli GS, Appleby PG, Oldfield F, editors. The use of nuclear techniques in sediment transport and sedimentation. Paris (France): UNESCO. p. 659–662.
- Arctic Monitoring and Assessment Programme (AMAP). 2017. Snow, Water, Ice and Permafrost in the Arctic (SWIPA) 2017. Oslo: AMAP.
- Battarbee RW, Kneen MJ. 1982. The use of electronically counted microspheres in absolute diatom analysis. Limnol Oceanogr. 27(1):184–188. doi:10.4319/lo.1982.27.1.0184.
- Bennett KD. 1996. Determination of the number of zones in a biostratigraphical sequence. New Phytol. 132(1):155–170. doi:10.1111/j.1469-8137.1996.tb04521.x.
- Bennett KD. 2009. PSIMPOLL version 4.27: C program for plotting pollen diagrams and analyzing pollen data. Belfast (Ireland): Queen’s University of Belfast. http://www.chrono.qub.ac.uk/psimpoll/psimpoll.html.
- Bergström AK, Jansson M. 2006. Atmospheric nitrogen deposition has caused nitrogen enrichment and eutrophication of lakes in the northern hemisphere. Global Change Biol. 12:635–643. doi:10.1111/j.1365-2486.2006.01129.x.
- Blondeau M, Dignard N. 2001. Rapport d’herborisation à Fermont, Québec: liste des espèces vasculaires et analyse sommaire de la flore [incluant quelques extensions d’aire dans la MRC de Caniapiscau]. Charlesbourg (QC): Ministère des Ressources Naturelles. Notices floristiques no. 4.
- Blott S. 2010. GRADISTAT version 8.0: A grain size distribution and statistics package for the analysis of unconsolidated sediments by sieving or laser granulometer. Crowthorne: Kenneth Pye Associates Ltd. http://www.kpal.co.uk/gradistat.html.
- Buczkó K, Magyari EK, Soróczki-Pintér É, Hubay K, Braun M, Bálint M. 2009. Diatom-based evidence for abrupt climate changes during the Late Glacial in the Southern Carpathian Mountains. Cent Eur Geol. 52(3–4):249–268. doi:10.1556/CEuGeol.52.2009.3-4.3.
- Campbell ID, Last WM, Campbell C, Clare S, McAndrews JH. 2000. The late-Holocene paleohydrology of Pine Lake, Alberta. J Paleolimnol. 24(4):427–441. doi:10.1023/A:1008155622656.
- Carpenter SR, Lodge DM. 1986. Effects of submersed macrophytes on ecosystem processes. Aquat Bot. 26:341–370. doi:10.1016/0304-3770(86)90031-8.
- Cellamare M, Morin S, Coste M, Haury J. 2012. Ecological assessment of French Atlantic lakes based on phytoplankton, phytobenthos and macrophytes. Environ Monit Assess. 184(8):4685–4708. doi:10.1007/s10661-011-2295-0.
- Clarke PJ. 1960. Rapport préliminaire sur la région de Normanville. Québec: Ministère des Mines R. P. 413.
- Clerk S, Hall R, Quinlan R, Smol JP. 2000. Quantitative inferences of past hypolimnetic anoxia and nutrient levels from a Canadian Precambrian Shield lake. J Paleolimnol. 23(3):319–336. doi:10.1023/A:1008147127606.
- Croudace IW, Rindby A, Rothwell G. 2006. ITRAX: description and evaluation of a new multi-function X-ray core scanner In: Rothwell RG, editor. New techniques in sediment core analysis. Vol. 267. London (UK): Geol Soc London Spec Publ; p. 51–63. doi:10.1144/GSL.SP.2006.267.01.04.
- Dufresne G. 1975. Problèmes d’aménagement nordiques et l’exemple de Fermont (N. Q.). [Bachelor’s Thesis]. Québec (QC): Université Laval.
- Ekdahl EJ, Teranes JL, Wittkop CA, Stoermer EF, Reavie ED, Smol JP. 2007. Diatom assemblage response to Iroquoian and Euro-Canadian eutrophication of Crawford Lake, Ontario, Canada. J Paleolimnol. 37(2):233–246. doi:10.1007/s10933-006-9016-7.
- Enache M, Prairie YT. 2000. Paleolimnological reconstruction of forest fire induced changes in lake biogeochemistry (Lac Francis, Abitibi, Quebec, Canada). Can J Fish Aquat Sci. 57(S2):146–154. doi:10.1139/f00-114.
- Engstrom DR, Wright HE. 1984. Chemical stratigraphy of lake sediments as a record of environmental change In: Haworth EY, Lund JWG, editors. Lake sediments and environmental history. Leicester (UK): Leicester University Press. p. 11–68.
- Environment and Climate Change Canada (ECCC). 2018. Canadian climate normals. http://climate.weather.gc.ca/climate_normals/index_e.html.
- Fallu M-A, Allaire N, Pienitz R. 2000. Freshwater diatoms from northern Quebec and Labrador (Canada): species-environment relationships in lakes of boreal forest, forest tundra and tundra regions. Bibliotheca Diatomologica 45. Berlin: J. Cramer.
- Finkelstein SA, Gajewski K. 2008. Responses of Fragilarioid-dominated diatom assemblages in a small Arctic lake to Holocene climatic changes, Russell Island, Nunavut, Canada. J Paleolimnol. 40(4):1079–1095. doi:10.1007/s10933-008-9215-5.
- Force ER. 1991. Geology of titanium-mineral deposits. Geol S Am S. 259:1–112.
- Garrison PJ, Wakeman RS. 2000. Use of paleolimnology to document the effect of lake shoreland development on water quality. J Paleolimnol. 24(4):369–393.
- Gibson CE, Anderson NJ, Haworth EY. 2003. Aulacoseira subarctica: taxonomy, physiology, ecology and palaeoecology. Eur J Phycol. 38:83–101.
- Hall RI, Smol JP. 2010. Diatoms as indicators of lake eutrophication In: Smol JP, Stoermer EF, editors. The diatoms: applications for the environmental and earth sciences. 3rd ed. Cambridge (UK): Cambridge University Press. p. 128–168.
- Heiri O, Lotter AF, Lemcke G. 2001. Loss on ignition as a method for estimating organic and carbonate content in sediments: reproducibility and comparability of results. J Paleolimnol. 25(1):101–110. doi:10.1023/A:1008119611481.
- Holtgrieve GW, Schindler DE, Hobbs WO, Leavitt PR, Ward EJ, Bunting L, Chen G, Finney BP, Gregory-Eaves I, Holmgren S, et al. 2011. A coherent signature of anthropogenic nitrogen deposition to remote watersheds of the Northern Hemisphere. Science. 334(6062):1545–1548. doi:10.1126/science.1212267.
- Jensen HS, Andersen FØ. 1992. Importance of temperature, nitrate, and pH for phosphate release from aerobic sediments of four shallow, eutrophic lakes. Limnol Oceanogr. 37(3):577–589. doi:10.4319/lo.1992.37.3.0577.
- Jeppesen E, Moss B, Bennion H, Carvalho L, DeMeester L, Feuchtmayr H, Friberg N, Gessner MO, Hefting M, Lauridsen TL, Liboriussen L, et al. 2010. Interaction of climate change and eutrophication In: Kernan M, Battarbee R, Moss B, editors. Climate change impacts on freshwater ecosystems. Chichester (UK): Blackwell Publishing Ltd. p. 119–151.
- Jeppesen E, Søndergaard M, Jensen JP, Havens KE, Anneville O, Carvalho L, Coveney MF, Deneke R, Dokulil MT, Foy B, et al. 2005. Lake response to reduced nutrient loading – an analysis of contemporary long-term data from 35 case studies. Freshwater Biol. 50(10):1747–1771. doi:10.1111/j.1365-2427.2005.01415.x.
- Joynt EH, Wolfe AP. 2001. Paleoenvironmental inference models from sediment diatom assemblages in Baffin Island lakes (Nunavut, Canada) and reconstruction of summer water temperature. Can J Fish Aquat Sci. 58(6):1222–1243. doi:10.1139/f01-071.
- Juggins S. 2014. C2 version 1.7.6: Software for ecological and palaeoecological data analysis and visualisation. Newcastle upon Tyne (UK): University of Newcastle. https://www.staff.ncl.ac.uk/stephen.juggins/index.html.
- Kernan M, Ventura M, Bitušík P, Brancelj A, Clarke G, Velle G, Raddum GG, Stuchlík E, Catalan J. 2009. Regionalisation of remote European mountain lake ecosystems according to their biota: environmental versus geographical patterns. Freshwater Biol. 54(12):2470–2493. doi:10.1111/j.1365-2427.2009.02284.x.
- Köster D, Pienitz R, Wolfe BB, Barry S, Foster DR, Dixit SS. 2005. Paleolimnological assessment of human-induced impacts on Walden Pond (Massachusetts, USA) using diatoms and stable isotopes. Aquat Ecosyst Health. 8(2):117–131. doi:10.1080/14634980590953743.
- Krammer K, Lange-Bertalot H. 1986. Bacillariophyceae 1. Teil: Naviculaceae In: Ettl H, Gerloff J, Heynig H, Mollenhauer D, editors. Süßwasserflora von Mitteleuropa. Stuttgart (Germany)/New York (NY): Gustav Fischer Verlag.
- Krammer K, Lange-Bertalot H. 1988. Bacillariophyceae 2. Teil: Bacillariaceae, Epithemiaceae, Surirellaceae In: Ettl H, Gerloff J, Heynig H, Mollenhauer D, editors. Süßwasserflora von Mitteleuropa. Stuttgart (Germany)/New York (NY): Gustav Fischer Verlag.
- Krammer K, Lange-Bertalot H. 1991a. Bacillariophyceae 3. Teil: Centrales, Fragilariaceae, Eunotiaceae In: Ettl H, Gerloff J, Heynig H, Mollenhauer D, editors. Süßwasserflora von Mitteleuropa. Stuttgart/Jena (Germany): Gustav Fischer Verlag.
- Krammer K, Lange-Bertalot H. 1991b. Bacillariophyceae 4. Teil: Achnanthaceae, Kritische Ergänzungen zu Navicula (Lineolatae) und Gomphonema In: Ettl H, Gerloff J, Heynig H, Mollenhauer D, editors. Süßwasserflora von Mitteleuropa. Stuttgart/Jena (Germany): Gustav Fischer Verlag.
- Laing TE, Rühland K, Smol JP. 1999. Past environmental and climatic changes related to tree-line shifts inferred from fossil diatoms from a lake near the Lena River Delta, Siberia. Holocene. 9(5):547–557. doi:10.1191/095968399675614733.
- Laperrière L, Fallu M-A, Hausmann S, Pienitz R, Muir D. 2008. Paleolimnological evidence of mining and demographic impacts on Lac Dauriat, Schefferville (subarctic Québec, Canada). J Paleolimnol. 40(1):309–324. doi:10.1007/s10933-007-9162-6.
- Lotter AF, Bigler C. 2000. Do diatoms in the Swiss Alps reflect the length of ice-cover? Aquat Sci. 62(2):125–141. doi:10.1007/s000270050002.
- Madsen JD, Chambers PA, James WF, Koch EW, Westlake DF. 2001. The interaction between water movement, sediment dynamics and submersed macrophytes. Hydrobiologia. 444(1/3):71–84. doi:10.1023/A:1017520800568.
- Malmaeus JM, Blenckner T, Markensten H, Persson I. 2006. Lake phosphorus dynamics and climate warming: a mechanistic model approach. Ecol Model. 190(1–2):1–14. doi:10.1016/j.ecolmodel.2005.03.017.
- McEachern P, Prepas EE, Gibson JJ, Dinsmore WP. 2000. Forest fire induced impacts on phosphorus, nitrogen, and chlorophyll a concentrations in boreal subarctic lakes of northern Alberta. Can J Fish Aquat Sci. 57(Suppl. 2):73–81. doi:10.1139/f00-124.
- Meyers PA. 2009. Organic geochemical proxies In: Gornitz G, editor. Encyclopedia of paleoclimatology and ancient environments. Dordrecht (Netherlands): Springer-Verlag; p. 659–662.
- Ministère des Richesses naturelles (MRN). 1977. Courbes bathymétriques du lac Carheil. Map C-8412, 1/42 353. Québec (Qc).
- Moser KA, Smol JP, MacDonald GM, Larsen C. 2002. 19th century eutrophication of a remote boreal lake: a consequence of climate warming? J Paleolimnol. 28(2):269–281. doi:10.1023/A:1021635024757.
- Murphy DL. 1960. Rapport préliminaire sur la région des Lacs Carheil et le Gentilhomme, district électoral de Saguenay. Québec: Ministère des Mines R. P. 412.
- Neal HE. 2000. Iron deposits of the Labrador Trough. Explor Mining Geol. 9:113–121. doi:10.2113/0090113.
- O’Neil JM, Davis TW, Burford MA, Cobler CJ. 2012. The rise of harmful cyanobacteria blooms: The potential roles of eutrophication and climate change. Harmful Algae. 14:313–334. doi:10.1016/j.hal.2011.10.027.
- Occhietti S, Parent M, Lajeunesse P, Robert F, Govare E. 2011. Late Pleistocene-Early Holocene decay of the Laurentide Ice Sheet in Québec-Labrador In: Ehlers J, Gibbard PL, Hughes PD, editors. Developments in Quaternary Science 15. Amsterdam (Netherlands): Elsevier. p. 601–630.
- Organisme de bassins versants Duplessis (OBVD). 2010. Portrait diagnostique de la problématique des cyanobactéries – bassin versant du lac Carheil. Sept-Îles (QC).
- Organisme de bassins versants Duplessis (OBVD). 2011. Cyanobactéries aux lacs Daviault, Sans Nom et Carheil. Volet I: hydrodynamique et physico-chimie. Étude partielle. Sept-Îles (QC).
- Organization for Economic Co-Operation and Development (OECD). 1982. Eutrophication of waters. Monitoring, assessment and control. Paris: OECD.
- Paerl HW, Otten TG. 2013. Harmful cyanobacterial blooms: causes, consequences, and controls. Microb Ecol. 65(4):995–1010. doi:10.1007/s00248-012-0159-y.
- Paerl HW, Paul VJ. 2012. Climate change: links to global expansion of harmful cyanobacteria. Water Res. 46(5):1349–1363. doi:10.1016/j.watres.2011.08.002.
- Paterson AM, Cumming BF, Smol JP, Blais JM, France RL. 1998. Assessment of the effects of logging, forest fires and drought on lakes in northwestern Ontario: a 30-year paleolimnological perspective. Can J Res. 28(10):1546–1556. doi:10.1139/x98-138.
- Paterson AM, Morimoto DS, Cumming BF, Smol JP, Szeicz JM. 2002. A paleolimnological investigation of the effects of forest fire on lake water quality in northwestern Ontario over the past ca. 150 years. Can J Bot. 80(12):1329–1336. doi:10.1139/b02-117.
- Peinerud EK. 2000. Interpretation of Si concentrations in lake sediments: three case studies. Environ Geol. 40(1–2):64–72. doi:10.1007/PL00013330.
- Pienitz R, Vincent WF. 2003. Generic approaches towards water quality monitoring based on paleolimnology In: Kumagai M, Vincent WF, editors. Freshwater management: global versus local perspectives. Tokyo (Japan): Springer-Verlag; p. 61–82.
- Potapova M, Hamilton PB. 2007. Morphological and ecological variation within the Achnanthidium minutissimum (Bacillariophyceae) species complex. J Phycol. 43(3):561–575. doi:10.1111/j.1529-8817.2007.00332.x.
- R Core Team. 2016. R: a language and environment for statistical computing. Vienna: R Foundation for Statistical Computing. https://www.r-project.org/.
- Rivers T, Martignole J, Gower CF, Davidson A. 1989. New tectonic divisions of the Grenville Province, Southeast Canadian Shield. Tectonics. 8(1):63–84. doi:10.1029/TC008i001p00063.
- Rouse WR, Douglas MSV, Hecky RE, Hershey AE, Kling GW, Lesack L, Marsh P, McDonald M, Nicholson BJ, Roulet NT, et al. 1997. Effects of climate change on the freshwaters of Arctic and Subarctic North America. Hydrol Process. 11(8):873–902. doi:10.1002/(SICI)1099-1085(19970630)11:8<873::AID-HYP510>3.0.CO;2-6.
- Rühland K, Paterson AM, Smol JP. 2008. Hemispheric-scale patterns of climate-related shifts in planktonic diatoms from North American and European lakes. Glob Change Biol. 14:2740–2754. doi:10.1111/j.1365-2486.2008.01670.x.
- Rühland K, Priesnitz A, Smol JP. 2003. Paleolimnological evidence from diatoms for recent environmental changes in 50 lakes across Canadian Arctic treeline. Arct Antarct Alp Res. 35(1):110–123. doi:10.1657/1523-0430(2003)035[0110:pefdfr]2.0.co;2.
- Saros JE, Clow DW, Blett T, Wolfe AP. 2011. Critical nitrogen deposition loads in high-elevation lakes of the Western US inferred from paleolimnological records. Water Air Soil Pollut. 216(1–4):193–202. doi:10.1007/s11270-010-0526-6.
- Saros JE, Northington RM, Anderson DS, Anderson NJ. 2016. A whole-lake experiment confirms a small centric diatom species as an indicator of changing lake thermal structure. Limnol Oceanogr. 1(1):27–35. doi:10.1002/lol2.10024.
- Saros JE, Stone JR, Pederson GT, Slemmons KEH, Spanbauer T, Schliep A, Cahl D, Williamson CE, Engstrom DR. 2012. Climate-induced changes in lake ecosystem structure inferred from coupled neo- and paleoecological approaches. Ecology. 93(10):2155–2164. doi:10.1890/11-2218.1.
- Smith VH. 1983. Low nitrogen to phosphorus ratios favor dominance by blue-green algae in lake phytoplankton. Science. 221(4611):669–671. doi:10.1126/science.221.4611.669.
- Smol JP. 1992. Paleolimnology: an important tool for effective ecosystem management. J Aquat Ecosyst Health. 1(1):49–58. doi:10.1126/science.221.4611.669.
- Solovieva N, Jones VJ, Nazarova L, Brooks SJ, Birks HJB, Grytnes JA, Appleby PG, Kauppila T, Kondratenok B, Renberg I, et al. 2005. Palaeolimnological evidence for recent climatic change in lakes from the northern Urals, Arctic Russia. J Paleolimnol. 33(4):463–482. doi:10.1007/s10933-005-0811-3.
- Sorgente D, Frignani M, Langone L, Ravaioli M. 1999. Chronology of marine sediments: interpretation of activity-depth profiles of 210Pb and other radioactive tracers – part I. Bologna (Italy): Consiglio Nazionale delle Ricerche Istituto per la Geologia Marina Bologna Technical Report n. 54.
- Spaulding SA, Bishop IW, Edlund MB, Lee S, Potapova M. 2018. Diatoms of North America. https://diatoms.org/.
- Statistics Canada. 1982. Census tracts: population, occupied private dwellings, private households, census families in private households – selected characteristics: Québec. Statistics Canada catalogue no. E-563. Ottawa (ON): Ministry of Supply and Services Canada.
- Statistics Canada. 2017. Fermont, V [Census subdivision], Quebec and Sept-Rivières–Caniapiscau, CDR [Census division], Quebec (table). Census Profile. 2016 Census. Statistics Canada Catalogue no. 98-316-X2016001. https://www12.statcan.gc.ca/census-recensement/2016/dp-pd/prof/index.cfm?Lang=E.
- Stuiver M, Reimer PJ, Reimer RW. 2017. CALIB 7.1 [WWW program]. http://calib.org.
- Tomkins JD, Antoniades D, Lamoureux SF, Vincent WF. 2008. A simple and effective method for preserving the sediment–water interface of sediment cores during transport. J Paleolimnol. 40(1):577–582. doi:10.1007/s10933-007-9175-1.
- Tremblay R, Pienitz R, Legendre P. 2014. Reconstructing phosphorus levels using models based on the modern diatom assemblages of 55 lakes in southern Quebec. Can J Fish Aquat Sci. 71(6):887–914. doi:10.1139/cjfas-2013-0469.
- Tremblay R. 2015. Élaboration d’une méthode basée sur les diatomées pour évaluer l’intégrité écologique des lacs tempérés du Québec (MILQ) [dissertation]. Québec (QC): Université Laval.
- Vallentyne JR. 1974. The algal bowl: lakes and man. Ottawa (ON): Department of the Environment, Fisheries and Marine Service Miscellaneous Special Publication 22.
- Verrette JL, Barbeau C, Moreau G. 1976. Ecological study of Carheil River – 1976. Québec (QC): Centre de recherches sur l'eau, Université Laval.
- Way RG, Viau AR. 2015. Natural and forced air temperature variability in the Labrador region of Canada during the past century. Theor Appl Climatol. 121(3–4):413–424. doi:10.1007/s00704-014-1248-2.
- Wolfe AP, Van Gorp AC, Baron JS. 2003. Recent ecological and biogeochemical changes in alpine lakes of Rocky Mountain National Park (Colorado, USA): a response to anthropogenic nitrogen deposition. Geobiology. 1(2):153–168. doi:10.1046/j.1472-4669.2003.00012.x.