
Abstract
Vitamin D has a well-known role in the calcium homeostasis associated with the maintenance of healthy bones. It increases the efficiency of the intestinal absorption of dietary calcium, reduces calcium losses in urine, and mobilizes calcium stored in the skeleton. However, vitamin D receptors are present ubiquitously in the human body and indeed, vitamin D has a plethora of non-calcemic functions. In contrast to most vitamins, sufficient vitamin D can be synthesized in human skin. However, its production can be markedly decreased due to factors such as clothing, sunscreens, intentional avoidance of the direct sunlight, or the high latitude of the residence. Indeed, more than one billion people worldwide are vitamin D deficient, and the deficiency is frequently undiagnosed. The chronic deficiency is not only associated with rickets/osteomalacia/osteoporosis but it is also linked to a higher risk of hypertension, type 1 diabetes, multiple sclerosis, or cancer. Supplementation of vitamin D may be hence beneficial, but the intake of vitamin D should be under the supervision of health professionals because overdosing leads to intoxication with severe health consequences. For monitoring vitamin D, several analytical methods are employed, and their advantages and disadvantages are discussed in detail in this review.
Introduction
At the end of the nineteenth century, it was known the bones of rachitic children had a low content of calcium and phosphate. However, supplementation with these elements did not lead to the prevention or cure of the disease. Rickets occurred mostly in cities in the northern latitudes and this suggested the lack of sun exposure as one of the possible explanations [Citation1,Citation2]. In some areas, there has also been a long-standing folk tradition of using cod liver oil as a powerful preventive agent [Citation3]. In the early twentieth century, the burst in experimentation and controlled studies confirmed the curative effect of both direct sunlight and cod liver oil, and “calcium-depositing vitamin” was discovered as the factor that cured rickets [Citation4–7]. This “calcium-depositing vitamin” later became known as vitamin D.
The name vitamin D covers a group of liposoluble steroid compounds of different origins with similar chemical structures and the same biological effects. Two main forms of vitamin D are ergocalciferol (vitamin D2) and cholecalciferol (vitamin D3) (). Vitamin D1 is a historical term for a mixture of vitamin D2 with lumisterol. Vitamin D2 is synthesized by the irradiation of ergosterol in yeast, while vitamin D3 is generated from 7-dehydrocholesterol after ultraviolet (UV)-B irradiation in the human skin—this being a unique property among vitamins (). Although vitamin D2 and D3 were considered equally active for many years, current knowledge indicates that the potency of vitamin D2 is less than one-third of that of vitamin D3 [Citation8–11]. The potential responsible factors are different metabolic pathways and/or different affinity of the active metabolites of vitamins D2 and D3 toward vitamin D receptor (VDR). Vitamin D3 is the main form of vitamin D in humans, and indeed, it is estimated that about 80–90% of the vitamin D requirements are covered by the endogenous synthesis in the skin. The extent of the skin vitamin D synthesis is dependent on the length of sun exposure, the season of the year, and latitude [Citation12]. A 20-min long whole-body exposure to the summer sun is able to produce up to 250 μg of vitamin D3 [Citation13,Citation14], which yields the recommended serum level (>30 ng/mL) of its metabolite and systemic indicator, 25-hydroxyvitamin D [25(OH)D], which is also known as calcifediol or calcidiol [Citation15]. Vitamin D3 is also present in small amounts in the diet of animal origin (e.g. fatty fish and fish liver oil, egg yolk, or dairy products). For the purposes of this review, the term 'vitamin D', unless otherwise stated, means vitamin D2 and/or vitamin D3. Over the past decade, another form of vitamin D was discovered in mushrooms, particularly those exposed to UV light. This so-called vitamin D4 (22-dihydroergocalciferol) is derived from its precursor, 22,23-dihydroergosterol [Citation16].
Figure 1. Chemical structure of vitamin D2 (A), vitamin D3 (B), and vitamin D4 (C). In online version, differences are shown in blue.
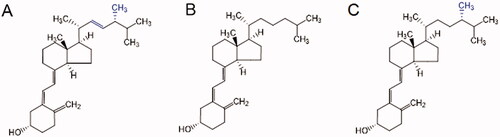
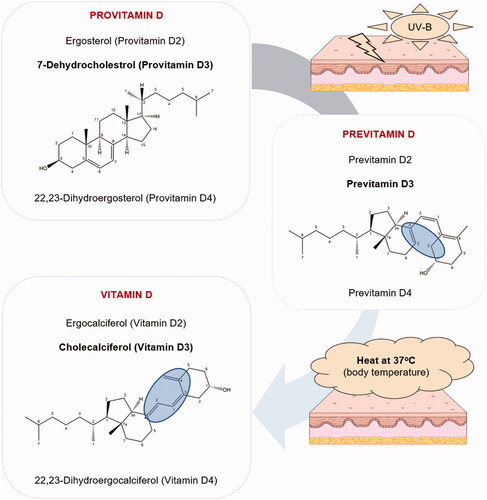
Figure 2. Synthesis of vitamin D. Upon UV-B radiation, the provitamins D ergosterol, 7-dehydrocholesterol, and 22,23-dihydroergosterol are respectively converted to pre-vitamins D2, D3, and D4, which are further thermally transformed into ergocalciferol (vitamin D2), cholecalciferol (vitamin D3), and 22,23-dihydroergocalciferol (vitamin D4). Only structures of provitamin D3, pre-vitamin D3, and vitamin D3 are depicted for better lucidity.
A number of effects are attributed to vitamin D. The most known and studied effect is linked to calcium and phosphate homeostasis, with a crucial impact on bone metabolism. In addition to mineral homeostasis, current research has been investigating a plethora of different activities of vitamin D. VDR, which interacts with the active form of vitamin D, calcitriol or 1,25-dihydroxycholecalciferol (1,25(OH)2D3), has been identified in almost all human cells, even in those that do not play a role in calcium metabolism, including the skin, brain, immune cells, prostate cancer cells, and pancreas [Citation17,Citation18]. Many preclinical, clinical, and epidemiological studies have demonstrated a beneficial effect of vitamin D on cardiovascular diseases [Citation19], diabetes [Citation20,Citation21], cancer [Citation22], depression [Citation23], cognition [Citation24], multiple sclerosis [Citation25], and even incidence of falls in the elderly people [Citation26]. Last but not least, in the recent COVID-19 pandemic, the most cited benefit is its immune-modulating effect [Citation27]. These data will be further discussed in more detail in this review.
Current human living conditions differ significantly from those of our evolutionary ancestors from equatorial Africa, avoiding direct sun exposure through the usage of clothes and living in a wide range of climate conditions [Citation28]. Therefore, deficiency of vitamin D is a relatively common problem in modern civilization [Citation29,Citation30] and supplementation may be beneficial in many cases. The goal of this article is to provide insight into vitamin D pathophysiology, pharmacognosy, basic and clinical pharmacology, detection, and toxicity.
Endogenous formation – photosynthesis of vitamin D3 in human skin
During the exposure to sunlight, UV-B photons penetrate the epidermis and the absorbed energy causes the photolysis of 7-dehydrocholesterol, present in the plasma membrane of keratinocytes, into the previtamin D3 () [Citation31–33]. The formed previtamin D3 is thermodynamically unstable and rapidly isomerizes to vitamin D3 [Citation34]. At 37 °C, 80% of previtamin D3 is isomerized to vitamin D3 within 8 h [Citation32]. Generated vitamin D3 is then released from the plasma membrane into the extracellular space, wherefrom it moves into the capillary bed and binds to plasma proteins [Citation35]. As mentioned above, skin synthesis can create 80–90% of the body's vitamin D pool under ideal conditions [Citation36].
Exogenous intake – dietary sources
Unfortunately, there are not many rich natural sources of vitamin D. Vitamin D2 is synthesized almost exclusively by fungi, which includes both microscopic species and fungi that form macroscopic fruiting bodies. UV-B radiation is also needed for the synthesis of vitamin D2 from ergosterol (), as in the case of conversion of 7-dehydrocholesterol to vitamin D3. Additionally, various levels of vitamin D2 have been reported in algae, and its traces can also be found in plants, probably as a result of fungal contamination. Contrarily, vitamin D3 occurs mainly in animal sources but can also be found in algae and plants. Due to the symbiotic coexistence of fungi and microscopic algae, both types of vitamin D are found in lichens [Citation37–39].
The main food sources of vitamin D vary largely according to eating habits and age. While meat and meat products (excluding fish), followed by other animal products, predominate in the UK adult population, fish by far outweighs vitamin D intake from other sources in Japan and chicken eggs in the Czech Republic [Citation40–42]. For non-breastfed infants, the main source of vitamin D is formula milk, which is later replaced by milk and dairy products. From the age of 4 to 10, meat and meat products, together with cereals and fat spreads, begin to predominate [Citation41].
An overview of the most important sources of vitamin D () shows that vitamin D3 dominates over vitamin D2 in natural dietary intake. Although fatty fish and fish oil are the richest sources of vitamin D3, they do not account for a large proportion of vitamin D intake in many countries (cod liver oil as a traditional preparation for the treatment of rickets was described in the eighteenth century) [Citation64]. Liver oil from wild cod fish contains up to 1,250 µg of vitamin D3 per 100 g. Other fish liver oil, especially various tuna species, has even higher vitamin D content than that from the cod. Interestingly, in comparison with other fish (e.g. mackerel, salmon, herring), cod and tuna flesh usually contain lower amounts of vitamin D3 [Citation43–45,Citation48]. Of the freshwater fish, rainbow trout and tilapia have a significant content of vitamin D3. Wild fish (marine and freshwater) usually have a higher vitamin content than farmed fish. An example is a salmon, where the content of vitamin D3 in wild specimens is up to four times higher than in farmed [Citation44,Citation47]. Other aquatic animals, such as mussels, oysters, or shrimps, contain negligible amounts of vitamin D3 compared to fish [Citation45].
Table 1. Vitamin D content in selected foodstuff and preparations.
Another important source of vitamin D3 is eggs, in which the majority is contained in the yolk. There are no significant differences in vitamin D3 contents between the individual animal species whose eggs are commonly consumed (e.g. chicken, duck, goose, quail), with values ranging from 2 to 5 µg/100 g of yolk. In poultry, vitamin D3 supplementation is of huge importance, as a significant increase in egg yolk vitamin D content can be achieved [Citation44,Citation51,Citation52,Citation65].
Compared to eggs, chicken meat and liver have a significantly lower vitamin D content. Of livestock meat, the highest amount of vitamin D3 is reported in pork. However, the variance of values is very wide because it depends on the part of the body from which the meat is taken as well as the animal’s diet and sun exposure. Most vitamin D is in fat, where it is stored and retained for several weeks after discontinuation of vitamin D3 supplementation. Concerning beef, there is large variability in the content of vitamin D3 among studies. Despite the fact that meat is not a rich source of vitamin D3 compared to fish, its consumption contributes significantly to the total daily intake of this vitamin due to the relatively large amount consumed [Citation44,Citation53–55,Citation66].
Dairy products are also an important source of vitamin D, especially in younger individuals. Milk naturally does not contain as much vitamin D3 as other sources, but some fatty dairy products like butter, whipping cream, and cheese, contain significant amounts and usually also contain some vitamin D2. Dairy products are also among the foods most often fortified with vitamin D3. In some countries, such as Canada, milk fortification is mandatory, whereas elsewhere it is optional [Citation44,Citation55,Citation67–70].
Although some plants contain significant amounts of vitamin D3 (e.g. tomato and potato plants), they cannot be considered dietary sources because the vitamin is contained in inedible parts [Citation38]. The main dietary source of vitamin D2 is mushrooms (). The number of vitamins differ enormously because it strictly depends on UV-B exposure. Most cultivated mushrooms are grown in the dark, where ergosterol is not converted to ergocalciferol, and therefore does not contain any vitamin D2. However, this conversion can also be achieved by post-harvest exposure. Exposure of mushrooms to the sun for 15 min can result in significant production of vitamin D2. The degree of conversion depends on the intensity and length of UV-B radiation. Artificial UV-B radiation is used for the production of vitamin D2 fortified mushrooms. Unlike cultivated mushrooms, wild mushrooms contain vitamin D2 naturally, but the amount varies considerably depending on growing conditions [Citation58–63]. Processing can also affect vitamin D2 levels. Except for sun-dried mushrooms, where the level of vitamin increases, there is typically a loss of vitamin D2 during drying and storage. Regardless, dried mushrooms can still contain almost 70% of the original vitamin content after 18-month storage [Citation60,Citation71].
In the case of vitamin D, relatively high retention (60–100%) has been observed after various types of heat treatment, such as baking, cooking, frying, or microwaving [Citation72–74]. There is generally no difference between natural sources and fortified foods in terms of heat-related vitamin D loss. Vitamin D is also stable during the long-term maturation of cheese and storage of fortified milk and dairy products with long shelf lives [Citation68–70,Citation73].
As vitamin D deficiency is still an important issue worldwide, there is an effort to promote vitamin D intake by fortifying foods. Vitamin D3 for fortification is made mainly from cholesterol, which comes from lanolin. It is used as a feed supplement for livestock and for fortification of foods for human consumption. The most frequently fortified foods are dairy products, but breakfast cereals, margarine, fruit juices, veggie alternatives of dairy products, and other foods are also enriched. Vitamin D of animal origin is not acceptable for some specific diets, especially the vegan diet, which lacks sufficient sources of this vitamin. Plant-based alternatives such as vitamin D obtained from lichens or algae are thus available. UV-B irradiated baker's yeasts are also a very rich source of vitamin D2 (), which can be used for supplementation of various food such as bread. Yeast-derived ergosterol in bread can be converted to vitamin D2 by UV-B even after baking [Citation57,Citation67,Citation75–79].
Oral absorption
As mentioned above, the current human population is more dependent on dietary intake of vitamin D to achieve the recommended serum level due to the modern way of life [Citation15]. Since vitamin D is liposoluble, it is assumed to share the fate of lipids, especially with its precursor cholesterol, in the human gastrointestinal tract, including emulsification, solubilization in mixed micelles, diffusion through the water layer, and permeation through the enterocyte membrane. However, this theory does not fully correspond to observed kinetics, because the absorption efficiency of vitamin D is lower and more variable than that of triacylglycerols [Citation12,Citation80].
Vitamin D absorption begins in the stomach, where pepsin plays a role by releasing the associated protein fraction. In the duodenum, proteases, amylases, and lipases continue the process of vitamin D release from the food matrix [Citation12]. Bile acids initiate the emulsification and formation of mixed micelles containing fat-soluble substances that are then absorbed by enterocytes [Citation81,Citation82]. Hydroxylated forms of vitamin D have a different fate. They have better water-solubility not requiring bile acids for absorption. Interestingly, oral 25(OH)D3 reached 3–4 fold higher plasma levels than oral vitamin D3 within 6 h [Citation83]. However, the exact absorption mechanisms of the hydroxylated form have yet to be discovered. Regardless, this form may be utilized in the vitamin D supplementation of the patients with fat malabsorption. Patients with intestinal resection, Crohn’s disease, cholestasis, or cystic fibrosis have significantly lower absorption of non-hydroxylated vitamin D in contrast to absorption of hydroxylated form, which is relatively well-preserved [Citation84–88].
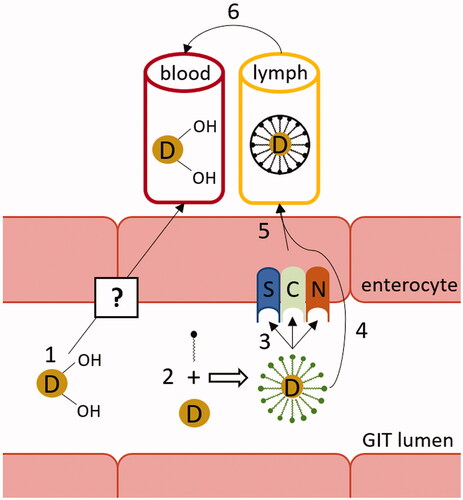
Vitamin D is absorbed in the small intestine, but the precise part of the intestine mediating the absorption is not known in humans. In rats, the main site of absorption is the ileum. The uptake of non-hydroxylated vitamin D at dietary concentrations is protein-mediated (). Proteins involved at the apical side include three intestinal cell membrane proteins: scavenger receptor class B type 1 (SR-B1), cluster of differentiation 36 (CD36), and Niemann-Pick C1-Like 1 (NPC1L1) [Citation89]. However, at pharmacological concentrations (>μM), passive diffusion also plays an important role in absorption. Based on a recent study with colorectal-derived Caco-2 cells, the existence of active efflux of the absorbed vitamin D from the intestine into the lumen was also suggested [Citation90].
Figure 3. Vitamin D absorption. Hydroxylated forms of vitamin D are absorbed directly into the vena portae by an unknown mechanism (1). Intact vitamin D is first built into mixed micelles (2). The uptake of vitamin D at dietary concentrations is protein-mediated (3), while at higher pharmacological concentrations it is absorbed through passive diffusion as well (4). Chylomicrons containing vitamin D are then secreted into the lymphatic capillaries (5) before reaching systemic circulation (6). C: cluster determinant 36 (CD36); D: vitamin D; GIT: gastrointestinal; N: Niemann-Pick C1-Like 1 (NPC1L1); S: scavenger receptor class B type 1 (SR-B1).
Absorbed vitamin D is then incorporated into chylomicrons secreted into the lymphatic capillaries, hence bypassing the first-pass metabolism. A fraction of vitamin D contained in chylomicrons might be transported and immediately stored in skeletal muscles and adipose tissues due to the action of lipoprotein lipase [Citation83].
Although vitamins D2 and D3 differ significantly in terms of the production of active metabolites, they possess equivalent intestinal absorption efficiency [Citation8,Citation89,Citation91]. Most data suggest that the food matrix has no significant effect on this parameter but there are other several factors that may affect vitamin D bioavailability [Citation12]. Some drugs used to inhibit lipid absorption may impair vitamin D absorption, for example, the anti-obesity drug orlistat [Citation92]. Plant phytosterols also compete with vitamin D for micelle incorporation and apical uptake in the intestine [Citation93]. Similarly, other fat-soluble vitamins A, E, and K may compete with vitamin D during absorption. The interaction between absorption of vitamin D and vitamin E was confirmed by Reboul et al. [Citation89]. As proteins are involved in vitamin D absorption, genetic mutations may also affect this process. Specifically, mutations in the gene promoter may affect the protein expression [Citation94] and mutations in amino acid sequence may affect the protein activity [Citation95].
Interestingly, most of the vitamin D, either taken orally or synthesized in the skin, fails to become 25(OH)D. Animal data showed that three-quarters of the vitamin D dose taken orally is not used for 25(OH)D synthesis [Citation96]. A small fraction of unmetabolized vitamin D is stored in adipose tissue and muscles. Less than 5% of vitamin D synthesized in the skin was subsequently found in fat tissue in shaven mice, and the biggest portion of vitamin D entering circulation appeared to be excreted into the bile [Citation97]. The half-life of unmetabolized vitamin D in circulation is 2 days [Citation28]. However, in healthy individuals, the biological half-life is much longer. Even with no supply, vitamin D3 can be continuously released from storage tissues for a period of 2–3 months [Citation98]. A plasma half-life of 25(OH)D is about 2 weeks [Citation99]. But again, the biological half-life is much longer due to synthesis from vitamin D3 stored in the body. The biological half-life of the active form, calcitriol, is 12 h [Citation28].
Distribution
Like steroid hormones, vitamin D and its metabolites exhibit high binding affinity to plasma proteins. Vitamin D binding protein (vDBP), earlier also known as a group-specific component or transcalciferin, was described in 1959, but its transport function was discovered in 1975 [Citation100–102]. The main physiological function of vDBP is the regulation of total and free circulating levels of vitamin D metabolites. The vDBP acts as their circulating reservoir [Citation103]. Dietary vitamin D is slowly transferred from chylomicrons to vDBP. In contrast, vitamin D synthesized in the skin is almost exclusively bound to vDBP [Citation104].
vDBP binds both unmetabolized vitamin D2/3 and all their metabolites, being commonly present in plasma in 50-fold excess over all vitamin D forms [Citation105]. The tightest binding to the vDPB is exhibited by 25(OH)D [Citation28]. vDBP affinity to vitamin D2 metabolites is lesser than that for D3 metabolites [Citation8].
vDBP has a half-life of 1.7 days, which is much shorter than that of 25(OH)D, indicating that 25(OH)D molecules are intensively recycled during their existence in the body. vDBP, either free or loaded with vitamin D or its metabolites, is removed from circulation mainly by the liver and kidneys. Thus, these organs have preferential access to vDBP-bound metabolites of vitamin D [Citation28,Citation106].
In normal individuals, approximately 85% of circulating vitamin D metabolites are strongly bound to vDBP, 15% are bound to the albumin with a much lower affinity, and 0.03% of 25(OH)D and 0.4% of the biologically active form calcitriol are present in the free form in serum. Most tissues are able to take up only free forms of calcitriol or 25(OH)D, therefore the regulation at this level is an important step. This is in contrast to the kidneys, which are capable of the uptake of 25(OH)D bound to vDBP due to the presence of an active receptor-based transport mechanism (megalin/cubulin complex) [Citation107–110].
The distribution volume of vitamin D metabolites decreases with higher water-solubility, with calcitriol being present only in the plasma compartment [Citation28]. Although the highest total concentration of vitamin D and its metabolites occurs in plasma, the larger pool of vitamin D and 25(OH)D is found in fat and muscles [Citation111,Citation112].
As obesity is commonly associated with low levels of serum vitamin D metabolites, several studies were conducted on this topic. Results showed that body mass index inversely correlates with peak vitamin D serum concentrations. Since the cutaneous production and intestinal uptake are not impaired, it was concluded that this phenomenon is caused simply by a larger body mass associated with the larger volume of distribution [Citation113,Citation114].
Other physiological and pathological conditions can also influence vDBP levels. For example, during pregnancy levels of vDBP in the maternal blood increase. This is associated with higher levels of 25(OH)D bound to vDBP in the mother. Contrarily, the concentration of free 25(OH)D is slightly higher in the cord serum [Citation115]. On the other hand, during critical illness and inflammation, including COVID-19, levels of vDBP go down [Citation116–118].
Metabolism
Both dietary and endogenously formed vitamin D2/3 are two steps away from their active forms. The first activation step is the conversion of vitamin D2/3 to its 25-hydroxylated form, calcifediol, denoted 25(OH)D2/3, in the liver (). There are many enzymes with 25-hydroxylase activity, but evidence indicates that microsomal cytochrome P450 2R1 (CYP2R1) is the principal vitamin D hydroxylase in humans. Mitochondrial CYP27A1 can also carry out this reaction but only in the case of vitamin D3 [Citation119–122]. There is also a unique alternative activation pathway for vitamin D2 by hepatic 24-hydroxylase (CYP24A1) since formed 24(OH)D2 may undergo further activation to 1,24(OH)D2 in the kidney [Citation123,Citation124]. Contrarily, hydroxylation of biologically active 1,25(OH)2D3 in position 24 results in its deactivation, as mentioned below.
Figure 4. Systemic vitamin D3 metabolism and its regulation. FGF23: fibroblast growth factor 23; PTH: parathyroid hormone.
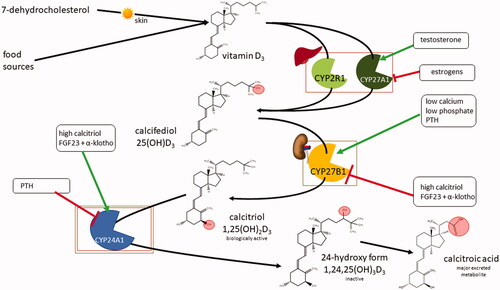
The second step in the activation of vitamin D is the conversion of 25(OH)D2/3 to its biologically active form calcitriol [1,25(OH)2D2/3] by the CYP27B1. This conversion takes place either in the kidneys or in a number of extrarenal tissues. Inactivation of calcitriol is ensured by the 24-hydroxylation, promoted by CYP24A1 (). This enzyme is expressed in most cells and is induced by elevation in calcitriol plasma concentrations, representing negative feedback protection against hypercalcemia. The formed 1,24,25(OH)3D has low biological activity and is further metabolized in the liver and kidneys into calcitroic acid, the major inactive vitamin D metabolite, which is excreted into the bile [Citation125].
Moreover, all the vitamin D metabolites may undergo epimerization in position C3 by the enzyme 3-epimerase. It is known that 3-epi-calcitriol has lower calcemic effects than its non-epimeric form. This is likely due to the fact that the affinity of C3-epimers to the VDR is about 2–3% of the respective calcitriol [Citation126–128]. This change in affinity is due to the different configurations of one functional group at a carbon C3 [Citation128]. 3-epi-calcitriol is the most potent one among all epimeric forms. Interestingly, the final effects related to calcemia and other vitamin D functions were comparable to calcitriol as it suppressed the parathyroid hormone secretion in bovine parathyroid cells [Citation129] and stimulated the synthesis of surfactant phospholipids and proteins in pulmonary alveoli [Citation127], both with the same potency as calcitriol. Contrarily, the CYP24A1 gene is activated by 3-epi-calcitriol at a 7–8 times lower rate than parent calcitriol [Citation126]. On the other hand, 3-epi-calcitriol has been proposed to have higher metabolic stability than its parent compound [Citation126,Citation130]. C3-epimers of calcifediol and calcitriol bind to the vDBP about 60% less often than corresponding non-epimers [Citation128]. The proportion of epimers to the total concentration of vitamin D metabolites is significantly higher in adult pregnant women and infants, and the concentration of epimers is also inversely proportional to age in children [Citation131,Citation132]. It is in these two groups where the epimer metabolites concentration may have clinical relevance. When epimer concentration was omitted from the assessment of vitamin D levels, 38% of women and 80% of newborns were classified as having an insufficient concentration. However, with epimer concentration included in the measurement of insufficiency, 33% of women and 73% of neonates were found to have sufficient levels of vitamin D [Citation133]. Finally, it is necessary to mention that the biological activity of C3-epimers has been demonstrated mainly with in vitro models and their physiological functions remain unclear. However, epimers are not the only vitamin D metabolites. The list of potential metabolic cascades of vitamin D is not yet fully deciphered, for example, 20-hydroxycholecalciferol produced locally by CYP11A1 induces keratinocyte differentiation [Citation134].
Returning to the main cascade, the production of calcitriol in the kidneys is crucial for mineral homeostasis and bone metabolism, and it is strictly controlled in humans. Calcitriol directly inhibits CYP27B1 in the kidneys [Citation135–138]. On the other hand, parathyroid hormone (PTH), for which release is induced by hypocalcemia, stimulates CYP27B1, resulting in elevation of calcitriol production. As negative feedback, calcitriol suppresses PTH release from the parathyroid glands by upregulation of calcium-sensing receptors and increasing calcium levels in serum [Citation139]. Direct inhibition of PTH release by calcitriol/VDR complex has also been discovered [Citation140]. The other regulation pathway is through fibroblast growth factor 23 (FGF23), produced by osteoblasts and osteocytes, that acts as a regulator of vitamin D metabolism through phosphate serum level. During hyperphosphatemia, FGF23 together with its cofactor α-klotho stimulates phosphate excretion in the kidneys, inhibits CYP27B1, and increases expression of CYP24A1, therefore lowering calcitriol concentration in plasma. In turn, calcitriol stimulates FGF23 expression [Citation141–143]. It has been shown that CYP27A1 is downregulated by estrogens and upregulated by testosterone in liver-based HepG2 cells [Citation144]. However, it must be emphasized that the regulation by sex hormones is by far more complicated, and contrarily, a decrease in calcium absorption and an increase in urinary calcium loss have been described in post-menopausal women [Citation145]. Of note, it was reported that estrogen loss may reduce VDR expression [Citation146], but evidence also shows that the estrogen effects in calcium absorption are vitamin D-independent, mediated by nuclear estrogen receptors [Citation147]. In addition, the metabolism of vitamin D is dependent on magnesium level, which is a cofactor for CYP2R1, 27B1, and 24A1 [Citation148].
As mentioned previously, the regulation of plasma levels of calcitriol is very meticulous. The enzymes involved are able to maintain stable calcitriol levels even during a number of pathophysiological processes. Moreover, circulating calcitriol is the result of renal metabolic activity and does not correspond to local calcitriol concentrations in the extrarenal tissues. These tissues rely on the availability of the precursor metabolite 25(OH)D in the plasma [Citation149]. Therefore, the measurement of calcitriol levels is of relatively low predictive value during the symptoms of vitamin D deficiency (see analytical methods section).
As mentioned previously, calcitriol produced by the kidney is responsible for the regulation of life-sustaining calcium homeostasis. However, other described non-calcemic functions of vitamin D are consequences of the local paracrine formation of calcitriol in non-renal tissues, for which plasma levels of free 25(OH)D3 are much more important than calcitriol levels [Citation28]. Aside from in the kidneys, the synthesis of calcitriol in non-renal tissues expressing CYP27B1 is also regulated in other ways. For example, the regulation is principally driven by cytokines in epithelial and immune cells [Citation150]. The extrarenal tissues are also the first that experience low levels of vitamin D. Extrarenal tissues usually do not express the megalin/cubulin system, therefore the uptake of 25(OH)D depends on passive diffusion, which is severely limited when serum levels of 25(OH)D are low [Citation151–154].
Malfunctions of any mentioned enzyme can cause human disease [Citation155]. With increasing age, the ability of kidneys to activate 25(OH)D3 declines. Also, increased expression of CYP24A1 and increased calcitriol clearance have been described [Citation156–158]. These factors may contribute to age-related bone thinning. Inborn diseases can also affect vitamin D metabolism. Vitamin D-dependent rickets (VDDR) type 1 is caused by an inactivating mutation in the CYP27B1 gene, subsequently leading to insufficient 1α-hydroxylation and thus reduced activation of 25(OH)D3 [Citation159]. There is also a rare hereditary variant of VDDR called type 1 b in which the loss-of-function mutation occurs in CYP2R1, leading to low levels of 25(OH)D [Citation160]. VDDR type 3 is associated with the gain-of-function mutation in the substrate recognition site of CYP3A4, which thereafter starts to extensively deactivate calcitriol [Citation161]. The mechanisms of vitamin D metabolism-mediated rickets as well as other genetically based rickets are schematically depicted in and summarized in .
Figure 5. Summary of genetic disorders in vitamin D3 metabolism. Vitamin D-dependent rickets (VDDR) type 1 is caused by the hypofunction of the activating enzyme CYP27B1. VDDR type 1 b arises one step earlier due to the hypofunction of the activating enzyme CYP2R1. VDDR type 2, also called vitamin D-resistant rickets, is caused by dysfunction of the substrate recognition site of the vitamin D receptor (VDR). VDDR type 3 is caused by the gain-of-function mutation of CYP3A4, which starts to deactivate vitamin D metabolites with even greater activity than CYP24A1.
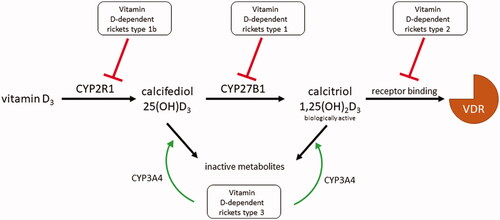
Table 2. Types of rickets induced by alterations in vitamin D metabolism.
Mechanism of action
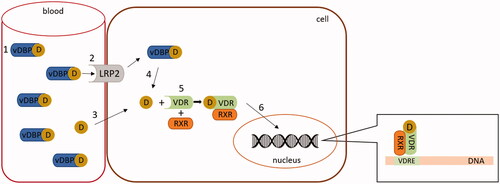
From a molecular perspective, the effects of vitamin D are mediated by VDR, a nuclear receptor. After entering the cell, calcitriol triggers heterodimerization of VDR with retinoid X receptor (RXR) and recruits other necessary regulatory molecules. The formed VDR/RXR dimer interacts with specific DNA sequences, known as vitamin D response elements (VDREs), in regulated genes and either activates or represses DNA transcription () [Citation162]. Recent research has discovered that VDREs are present not only at proximal promoters of the target gene but can also be situated within introns or intergenic regions with many kilobases in front or behind the regulated gene [Citation138,Citation163].
Figure 6. Mechanism of vitamin D (calcitriol) action. In this figure vitamin D should be understood as its active form - calcitriol. The majority of circulating vitamin D is bound to vitamin D binding protein (vDBP) (1). This complex may only enter cells with the megalin/cubulin system (LRP2) (2). Free vitamin D can enter any cell through passive diffusion (3). vDBP-bound vitamin D is released inside the cells (4). In the cytoplasm, vitamin D interacts with its receptor (VDR) and creates a heterodimer with retinoid X receptor (RXR) (5). The active VDR complex enters the nucleus (6) and binds to the responsive elements (VDRE) of regulated genes.
Biological effects of vitamin D
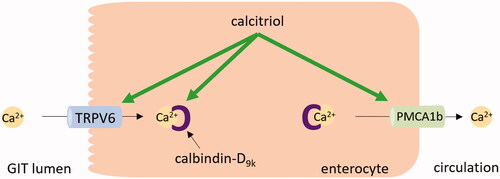
The most well-known physiological role of vitamin D is in the control of calcium homeostasis. When hypocalcemia occurs, the increase in calcium levels mediated by vitamin D is ensured by several pathways. In the intestine, the increased absorption of calcium is crucially dependent on vitamin D. With low levels of vitamin D, the small intestine absorbs only 10–15% of dietary calcium. When vitamin D levels are adequate, the absorption rises to 30–40% [Citation164,Citation165]. Calcitriol stimulates the synthesis of transient receptor potential vanilloid channel 6 (TRPV6), an apical epithelial calcium channel that facilitates calcium entry into the enterocyte, and calbindin-D9k, an intracellular calcium-binding protein that buffers potentially toxic levels of calcium in the enterocytes (). Both these proteins are expressed in the duodenum [Citation166]. In experiments with TRPV6 and calbindin-D9k knockout mice, the absorbed amount of the calcium was reduced but not fully diminished [Citation167]. This suggests another calcium absorption pathway, which is likely independent of vitamin D. Moreover, most of the ingested calcium is absorbed in the distal intestine where VDR and common transcellular mediators are also expressed [Citation168,Citation169]. These findings suggest that calcium absorption mediated by vitamin D in the intestine is a very complex process and more involved proteins have yet to be discovered [Citation170]. In the final step, the plasma membrane calcium pump (PMCA), whose isoform PMCA1b in the intestine is upregulated by calcitriol, transports calcium from enterocytes into the circulation [Citation171,Citation172]. In the kidney, calcitriol stimulates calcium reabsorption in the distal tubule. This process is achieved by an increase in expression of the apical calcium channel TRPV5 and protein calbindin-D28k [Citation137,Citation173].
Figure 7. Calcium absorption in the enterocyte. In hypocalcemia, calcitriol upregulates calcium transient receptor potential vanilloid channel 6 (TRPV6) and calbindin-D9k in the enterocyte, thus stimulating the calcium absorption in the intestine. Also, the expression of plasma membrane calcium pump type 1b (PMCA1b) is increased by calcitriol. GIT: gastrointestinal tract.
When increased absorption is not sufficient to maintain calcium homeostasis, preservation of blood calcium levels is prioritized over skeletal integrity. Calcitriol upregulates the receptor activator of nuclear factor κΒ ligand in osteoblastic cells, increasing osteoclastogenesis and promoting the release of calcium from the bones [Citation137,Citation138,Citation174]. The antirachitic effect of vitamin D is primarily indirect, caused by sufficient calcium plasma levels.
VDR is present in numerous tissues and cells that are not involved in calcium homeostasis (e.g. immune cells, the pancreas, brain, colon, breast, and skin). However, data on the non-classical effects of vitamin D are less documented. Calcitriol actively participates in the upregulation of genes encoding proteins required for the tight, gap, and adherens junctions located in epithelial cells in the skin, gut, respiratory and urinary tract, thus, boosting their barrier function [Citation175–178]. Moreover, it induces the expression of cathelicidin, a peptide with a broad range of antimicrobial activity in monocytes, lung and intestinal epithelium cells, keratinocytes, and placenta [Citation179,Citation180]. The active form of vitamin D activates hydrogen peroxide secretion in human monocytes, increasing oxidative burst potential [Citation178,Citation181]. Calcitriol also influences adaptive immunity. Specifically, it suppresses the production of inflammatory cytokines (IL-2, interferon-γ, IL-12, and IL-17), suppresses antigen-presentation by dendritic cells, and stimulates T-regulatory cells involved in the inhibition of inflammation [Citation137,Citation179,Citation182]. These effects are likely responsible for the beneficial effect of vitamin D in some autoimmune diseases [Citation182].
The most recent discoveries suggest that vitamin D may have a more specific effect in patients with SARS-CoV-2 infection than just boosting immunity, as a fatty acid-binding pocket was discovered in the spike protein trimer of the virus. The binding of linoleate in this pocket stabilizes the spike protein in the “locked” conformation and reduces the chance of receptor-mediated cell entry of SARS-CoV-2 via binding to angiotensin-converting enzyme-2, thus reducing infectivity [Citation183]. Therefore, the fatty acid-binding pocket represents one of the therapeutic targets in COVID-19 patients, and in silico simulations identified vitamin D as a potential ligand of this pocket [Citation184]. Further research must be conducted on this topic to verify the real clinical impact of these findings.
In an animal model, calcitriol exhibited a suppressing effect on cancer cell growth, more specifically on skin, colon, mammary, and prostate cancer cells [Citation185]. The topical application of calcitriol is beneficial also in psoriasis, a disease that is characterized by hyperproliferation and abnormal differentiation of keratinocytes [Citation186]. In the skin, vitamin D regulates keratinocyte proliferation, differentiation, and apoptosis through VDR [Citation187]. The effects are dose-dependent. The physiological concentration of vitamin D promotes keratinocyte growth and protects against apoptosis. However, pharmacological concentrations inhibit keratinocyte proliferation and stimulate their selective apoptosis [Citation188]. The regulation of keratinocytes is also influenced by local metabolism, since 20-hydroxycholecalciferol produced by CYP11A1 induces keratinocyte differentiation [Citation134], as previously mentioned. Moreover, vitamin D corrects the altered distribution of CD26, intercellular adhesion molecule 1 (ICAM-1), and human leukocyte antigen DR isotype (HLA-DR) integrins in psoriatic skin [Citation189].
Many other effects of vitamin D have been proposed but extensive clinical studies are needed to verify these hypotheses and thorough research has to be carried out in order to identify the potential biological relevance of these novel data.
Vitamin D deficiency: causes and symptoms
Although vitamin D3 can be endogenously synthesized in the human body, there are many factors that could impair the synthesis of the vitamin. The major cause of vitamin D deficiency is insufficient sun exposure and thus reduced skin synthesis. Limited sun exposure can occur due to a number of factors: (i) season and high latitude, where above 40° latitude the UV-B photons reaching the skin surface are reduced by more than 80% in the winter months [Citation190]. In general, low intensity of sunshine inadequate for sufficient vitamin D production lasts from October to March in the northern hemisphere at latitudes greater than 40° north and from April to September in the southern hemisphere above latitude 40° south; (ii) increased amount of skin pigment melanin [Citation191]; (iii) whole-body clothing related to climate or cultural tradition; and (iv) impaired mobility [Citation192]. Sunscreen with a sun protection factor of only 15 decreases the vitamin D synthetic capacity of the skin by 98% [Citation193]. On the other hand, sunscreens protect against burn injury, which substantially lowers the synthesis of vitamin D even after proper sun exposure [Citation194]. Additionally, aging decreases the concentration of 7-dehydrocholesterol, the vitamin D3 precursor, in human skin, therefore vitamin D synthesis is also reduced with age [Citation195]. The impact of sun exposure on vitamin D levels in infants and the elderly is very important and thus intake of this vitamin from dietary sources or supplements is very relevant for these populations [Citation40,Citation41,Citation44]. As the majority of foods containing vitamin D are of animal origin, individuals on a vegetarian and/or vegan diet may be at higher risk of vitamin D deficiency. Additional causes of vitamin D deficiency include various gastrointestinal and renal disorders, genetic mutations of involved enzymes, vDBP polymorphisms, tumors, pregnancy and therapeutic drug interactions, as briefly discussed below.
Orally administered vitamin D is absorbed in the gut; therefore, gastrointestinal disorders may cause vitamin D malabsorption and subsequent deficiency. In addition to bile acid production needed for vitamin D absorption, the liver is crucial for vitamin D metabolism due to the 25-hydroxylation step. Some examples of gastrointestinal disorders that may lead to vitamin D deficiency are obstructive liver disease, food allergies, celiac disease, cholestasis, biliary obstructions, inflammatory bowel syndrome, and cystic fibrosis. Patients after gastrectomy may also experience vitamin D deficiency due to the loss of acidity and malfunction of the proximal part of the small intestine [Citation196,Citation197].
Since the kidneys are the main site of vitamin D activation, chronic renal diseases are also associated with a low vitamin D activation rate, particularly due to loss of CYP27B1 and suppression of this enzyme as a consequence of hyperphosphatemia [Citation198]. Moreover, patients with nephrotic syndrome may develop vitamin D deficiency due to loss of 25(OH)D bound to vDBP in urine [Citation199,Citation200].
As shown in , genetic mutations of enzymes responsible for vitamin D metabolism may cause vitamin D deficiency due to low synthesis or high catabolism of active metabolites. The most common causes are a mutation of CYP27B1, 2R1, and 3A4 leading to VDDR type 1, 1 b, and 3, respectively [Citation159–161]. Another site of potential detrimental mutations is VDR. In this case, serum levels of vitamin D are usually preserved, however, the response of target tissues is insufficient due to defective VDR. This disease is classified as VDDR type 2 [Citation201]. Other possibilities are the mutations in the FGF23 gene, leading to high concentrations of FGF23 and therefore decreased kidney synthesis of calcitriol [Citation202,Citation203].
There are at least 120 identified isoforms of vDBP [Citation204]. These isoforms differ in the affinity to vitamin D metabolites and as a consequence, the level of free 25(OH)D in the plasma is different [Citation205,Citation206]. The clinical impact of these polymorphisms is not yet fully understood. However, the presence of certain isoforms is an additional risk factor for vitamin D deficiency independent of other factors like age, sex, or bodyweight [Citation207]. Vitamin D insufficiency can also be caused by small and otherwise often benign tumors that produce FGF23 [Citation208].
Primary hyperparathyroidism affects vitamin D metabolism, decreasing the the serum 25(OH)D level [Citation209]. Hyperthyroidism also reduces 25(OH)D plasma concentration due to enhanced catabolism of the metabolites [Citation210,Citation211].
Moreover, during pregnancy, vitamin D is transported across the placenta into the fetus and increased intake of vitamin D is needed to prevent deficiency in both the mother and fetus. Also, infants receiving solely breast milk are at risk of vitamin D deficiency due to its low content [Citation212,Citation213].
Lastly, certain drugs are capable of altering vitamin D metabolism and hence cause vitamin D deficiency. Phenytoin, carbamazepine, isoniazid, and rifampicin can bring about vitamin D deficiency due to induction CYP enzymes and subsequently enhanced catabolism of 25(OH)D and calcitriol. Cholestyramine and similar drugs that decrease absorption of fats as well as orlistat may reduce vitamin D absorption. In addition, ketoconazole blocks the activation of 25(OH)D in the kidneys and higher doses of vitamin D are needed during concomitant administration to achieve proper plasma levels of the active metabolite [Citation214–216].
The diagnosis of vitamin D deficiency is often carried out by measuring plasma calcifediol. An optimal concentration of 25(OH)D is still the subject of debate, although 32 ng/mL (80 nmol/L) or higher is considered by scientific consensus to be sufficient to fulfill its physiological functions [Citation217,Citation218]. Around this concentration, the level of PTH starts to drop and intestinal calcium absorption is about 60% more effective than at a 25(OH)D level of 20 ng/mL [Citation219,Citation220]. If we accept the optimal level of calcifediol to be 32 ng/mL, the prevalence of vitamin D deficiency is about 1 billion people worldwide [Citation217,Citation221].
Symptoms of vitamin D deficiency are logically linked to the malfunction of vitamin D-dependent processes. The deficit causes decreased absorption of dietary calcium and phosphate, which affects the quality of bones. In childhood, vitamin D deficiency manifests as rickets. This disease is characterized by a delay in closure of the fontanels, bowing of long bones, malformations in knees and wrists, scoliosis or kyphosis, and poor growth [Citation192,Citation213,Citation222]. Additionally, hypocalcemic seizures may often occur in the first year of life [Citation222]. In adults, vitamin D deficiency causes osteomalacia and osteoporosis, diseases characterized by demineralization of bones with an increased risk of fractures [Citation216,Citation223]. Bone deformities are not common symptoms in adults, in contrast to children [Citation192]. Unlike osteoporosis, osteomalacia is accompanied by isolated or generalized bone pain [Citation224,Citation225].
Other vitamin D deficiency symptoms are muscle weakness and fatigue [Citation26,Citation213,Citation216], increased susceptibility to infectious diseases [Citation226–228], and slower healing of epidermal wounds [Citation229]. Several studies have shown that profound vitamin D deficiency is even associated with the atrophy of type II muscle fibers [Citation230,Citation231]. Localized hair loss (i.e. alopecia areata) has also been reported [Citation232].
In the brain, VDR and vitamin D metabolizing enzymes are expressed, especially in the hypothalamus and large neurons of the substantia nigra. Since vitamin D regulates calcium transients in the brain and neuronal development as well as protects against reactive oxygen species, this vitamin deficiency may be involved in neurological disorders such as multiple sclerosis [Citation233].
Use of vitamin D in therapeutics
Guidelines for vitamin D supplementation are partly based on the recommended intake values to prevent vitamin D deficiency or to achieve target 25(OH)D levels. In addition, some more recent findings show that in addition to maintaining certain levels of 25(OH)D, it is also important to limit any significant fluctuations. This is because extrarenal tissue that lacks complex regulation mechanisms like the kidneys is sensitive to those fluctuations, and even bolus doses of 25(OH)D may lead to adverse effects [Citation234,Citation235].
Vitamins D2 and D3 are generally preferred for the treatment of simple vitamin D deficiencies, including those due to inadequate exposure of the skin to UV sunlight or lack of vitamin D in the diet. Individuals susceptible to these factors include the elderly with limited mobility and thus decreased sunlight exposure, obese patients, and people with fat malabsorption syndromes or chronic liver diseases [Citation236,Citation237]. The primary prevention of vitamin D deficiency is debated across national health policies, even though undiagnosed deficiency of vitamin D is not uncommon and the risk of bone disease as well as other chronic health disturbances may occur [Citation221,Citation238]. In particular, vitamin D deficiency may appear in infants who are breastfed without supplemental vitamin D; therefore, vitamin D supplementation of 400 IU/day for up to six months in infants at higher risk of lack of vitamin D reduces the risk of vitamin D insufficiency. These infants can also be supported by maternal vitamin D supplementation. Nevertheless, the influence of vitamin D supplementation on bone markers is still unclear [Citation239].
Vitamin D in all its forms together with calcium is employed as a standard treatment of vitamin D deficiency and hypocalcemia due to primary or secondary hypoparathyroidism [Citation240]. Secondary hypoparathyroidism can be commonly caused by thyroidectomy when patients with severe preoperative vitamin D deficiency are at much higher risk of permanent hypoparathyroidism. However, the evidence that supplementation of vitamin D in patients undergoing this surgery can reduce this risk is not robust [Citation241]. When large doses or fast potent onset of vitamin D action are required in hypoparathyroidism, it is preferable to use one of the more potent derivatives. In particular, the use of vitamin D derivatives, such as synthetic drugs (i.e. alfacalcidol or calcitriol), is necessary for conditions when renal function is impaired (e.g. hyperparathyroidism associated with chronic renal failure) as they do not require renal hydroxylation for activation [Citation240,Citation242,Citation243].
The important use of vitamin D plus calcium lies in the management of bone mass disorders, such as the prevention of osteoporosis fractures or osteomalacia in men and women [Citation244]. Vitamin D derivatives can serve as alternatives in these conditions in overcoming resistance to calcitriol due to age-related decline in the expression of VDR, especially in postmenopausal women. On the other hand, they can be related to a risk of hypercalcemia with hypercalciuria [Citation245,Citation246]. The risk of hypercalcemia caused by vitamin D and its derivates can be even higher in the primary prevention of osteoporosis in people without specific risk factors of vitamin D deficiency, and in these cases, supplementation is rather inappropriate because of side effects such as nephrolithiasis or calcinosis [Citation247–249]. Among the active vitamin D analogs, calcitriol and alfacalcidol have been introduced to the European market, particularly for postmenopausal osteoporosis and renal bone disease indications. Additionally, they can be used in off-label regimens for bone loss due to chronic long-term corticosteroid or antiepileptic drug administration or for male osteoporosis [Citation248,Citation250].
Vitamin D supplements may play a beneficial role in the direct protection of bones and skeletal muscles, increasing bone mineral density and muscle strength, performance, and balance, thus decreasing the incidence of falls and fractures [Citation248]. However, this issue is still controversial as it is unknown whether and to what extent vitamin D may influence muscle strength since the indication of fall prophylaxis in the elderly is mentioned just by some international databases [Citation251]. Of note, muscle strength and performance are associated with the lower range of serum 25(OH)D levels and may not necessitate elevations to the normal or higher range [Citation252]. Moreover, the use of excessive daily doses (2000 or 4000 IU) of vitamin D3 should even be reevaluated in the elderly due to an apparent relationship with increased risk of falls in this population [Citation253].
Other diseases may require increased vitamin D intake. Patients with severe renal failure can suffer from impaired activation of vitamin D. A special risk group is represented by patients with moderate to severe chronic renal insufficiency, dialysis patients, and patients after kidney transplantation, respectively. Kidney Disease: Improving Global Outcomes (KDIGO) recommends supplementing vitamin D2/3 with individualized monitoring of vitamin D levels, usually with an annual frequency. Indeed, the importance of vitamin D supplementation in these patients is highlighted because they must maintain significant dietary restrictions and avoid sunbathing, the latter of which causes reduced cutaneous synthesis of vitamin D [Citation254–256]. If patients have concomitant elevation in serum PTH levels, it is recommended to directly supplement active forms of vitamin D [Citation257].
Lastly, topical therapies with vitamin D or analogues (e.g. calcitriol) belong among the first-line treatment in the management of mild to moderate psoriasis. For better efficacy, a combination with synergistic topical corticosteroids can be used [Citation258]. Moreover, oral forms of vitamin D, such as vitamin D2, vitamin D3, or calcitriol have been reported as effective and safe treatment options for plaque psoriasis, similar to UV-B phototherapy and sunshine [Citation259].
Vitamin D in specific health conditions
In addition to the proven indications of vitamin D mentioned above, the clinical use of this vitamin in other health conditions or diseases is currently being intensively investigated.
COVID-19
To mitigate the huge health and socioeconomic consequences of the COVID-19 pandemic, the immunopathology of COVID-19 requires new therapeutic options such as vitamin D, which could be an effective and safe strategy for the management of this disease [Citation260]. As described in this review, apart from maintaining bone integrity, vitamin D can stimulate immune cell maturation and regulate inflammatory processes in the body. Experimental animal studies suggested a possible higher risk of acute viral infections with vitamin D depletion [Citation261]. A current meta-analysis of randomized controlled trials (RCTs) supports that regular vitamin D supplementation can prevent such infections and its efficacy was more pronounced in people with deficiency [Citation262]. Nevertheless, studies on the use of vitamin D in humans to prevent viral respiratory infections are inconsistent and the results, which are influenced by diverse variables and limits arising from the variability of in vitro and in vivo studies and the design of clinical trials, cannot be extrapolated to all types of infections, including SARS-CoV-2, or to all population groups [Citation263]. So far, there is no clear evidence of the clinical benefit of vitamin D supplementation in COVID-19 patients due to a lack of RCTs [Citation260].
Cardiovascular diseases
Vitamin D deficiency plays an important role in the etiology and pathogenesis of cardiovascular disease; however, systematic reviews have brought conflicting conclusions in relation to vitamin D supplementation in cardiovascular disease prevention. Lack of vitamin D has been described as a risk factor for arterial hypertension, but no clinically significant effect on blood pressure was observed during vitamin D supplementation in hypertensive patients with low levels of 25(OH)D [Citation264]. The relationship between vitamin D deficiency and the risk of heart disease has also been demonstrated, although administration of vitamin D has not been shown to be an effective therapeutic intervention. Currently published reviews and meta-analyses of RCTs did not confirm the benefit of vitamin D supplementation for incidence of coronary heart disease and stroke, risk of serious cardiovascular adverse events, individual cardiovascular disease outcomes (myocardial infarction, stroke, cardiovascular mortality), or all-cause mortality [Citation265–267]. Although, positive impacts may be expected in patients with a history of heart failure as vitamin D supplementation improved left ventricular function. Nevertheless, future RCTs are needed to investigate if patients without vitamin D deficiency can benefit from routine vitamin D administration [Citation268]. One of the reasons vitamin D supplementation fails to elicit cardiovascular protection could be the fact that both high and low levels of vitamin D can be associated with vascular calcification [Citation269].
Inflammatory bowel disease
Inflammatory bowel disease (IBD) is a multifactorial disease and vitamin D deficiency is considered a risk factor [Citation270]. The prevalence of vitamin D deficiency in IBD is about 30–40%. It may be caused by limited sun exposure, malabsorption, or inadequate dietary intake, the latter of which may be partly related to the fact that patients with IBD must be careful with their diet and may hence avoid food fortified with vitamin D [Citation271,Citation272]. Although a recent meta-analysis demonstrated that vitamin D supplementation significantly increased 25(OH)D levels in patients with IBD [Citation273], the impact of this adjuvant treatment on inflammation and the course of IBD is not yet clearly proven, nor is the target level of vitamin D known. Regardless, vitamin D supplementation should be considered in patients with vitamin D deficiency or those taking corticosteroids [Citation274].
Rheumatoid arthritis
It is assumed that vitamin D does not play an important role in the pathogenesis of rheumatoid arthritis. This fact was supported by a recent meta-analysis, which showed that there are insufficient data to demonstrate the effect of vitamin D on rheumatoid arthritis activity (DAS-28 or pain) or flares [Citation275]. On the contrary, vitamin D supplementation can be employed to prevent musculoskeletal complications in patients suffering from rheumatoid arthritis with a deficiency of this vitamin.
Cancer
The role of vitamin D in the pathogenesis of cancer has been discussed for a long time, but the conclusions are still unclear. The strongest evidence was demonstrated in colorectal cancer [Citation276,Citation277]. A meta-analysis of epidemiological studies has shown that vitamin D deficiency is associated with a higher incidence of colorectal cancer, and the highest risk was found in individuals with 25(OH)D levels lower than 20 ng/ml (50 nmol/L) [Citation278]. Indeed, causality modeling supports a relationship between low levels of 25(OH)D and a higher risk of colorectal cancer [Citation279]. These findings align with a review of prospective cohort studies, in which circulating 25(OH)D levels were measured and showed a high prevalence of vitamin D deficiency in colorectal cancer patients [Citation280]. On the contrary, patients with higher levels of 25(OH)D had longer survival, although this phenomenon may be related to the stage of colorectal cancer [Citation281]. The longer survival of patients with colorectal cancer treated with vitamin D was confirmed by a recent meta-analysis of RCTs [Citation282] and long-term vitamin D supplementation was shown to confer some additional benefits to patients suffering from this cancer [Citation283]. Still, all these results need to be further analyzed in larger high-quality cohort studies, also focusing on determining an adequate daily dose of vitamin D for these patients. Currently, the therapeutic administration of vitamin D3 and its analogues is considered appropriate in patients with a history of clinically significant vitamin D deficiency. Regarding the preventive administration of vitamin D3, even a daily dose of 2000 IU was not associated with a reduced risk of invasive cancer compared to a placebo, although a non-significant benefit on the incidence of colorectal cancer was reported [Citation284].
For other types of cancers, clear evidence for the therapeutic administration of vitamin D is missing. As an example, a systematic review and meta-analysis by Shahvazi et al. demonstrated no benefit of vitamin D in patients with prostate cancer [Citation285]. In contrast, a study by Nair-Shalliker et al. indicated the therapeutic potential of vitamin D even with an impact on the mortality of patients with prostate cancer [Citation286]. Therefore, any discrepancy needs to be resolved in the future by summarizing data from large RCTs. Finally, it should be mentioned that the administration of vitamin D is suitable for androgen deprivation therapy in patients with prostate cancer because vitamin D reduces the adverse effects of this therapy on the musculoskeletal system [Citation287,Citation288].
Vitamin D toxicity
Vitamin D displays a wide therapeutic window, with toxicity being observed only at extremely high doses [Citation289]. As such, the number of cases of vitamin D intoxication reported is low but affects all age groups (see intoxication cases section) [Citation290–292].
To date, there is no consensus on the dose of vitamin D that causes toxicity or the upper safe limit of serum/plasma levels of 25(OH)D3 [Citation293], possibly as distinct populations are differently affected by several factors (e.g. geography, lifestyle, genetics). Nonetheless, most studies have shown that hypercalcemia appears when serum 25(OH)D3 concentration is higher than 150 ng/mL (375 nmol/L) [Citation294–296]. As such, a concentration of 100 ng/mL is accepted by the Endocrine Society [Citation297] as posing no risk of developing hypercalcemia. In terms of intake, the 2011 report on dietary reference intakes for calcium and vitamin D from the Institute of Medicine set the no observed adverse effect level at 10,000 IU/day, from which a tolerable upper intake level of 4000 IU/day could be extrapolated for adults (), reflecting acceptable long-term chronic intake for general public health [Citation298].
Figure 8. Vitamin D overdose. Excessive intake of vitamin D due to intentional or inadvertent incorrect dosing (e.g. prescribing errors, supplementation with products that have low or no quality control) may increase plasma 25-hydroxyvitamin D [25(OH)D] to concentrations susceptible to causing toxicity. NOAEL: no observed adverse effect level; UL: tolerable upper intake level.
![Figure 8. Vitamin D overdose. Excessive intake of vitamin D due to intentional or inadvertent incorrect dosing (e.g. prescribing errors, supplementation with products that have low or no quality control) may increase plasma 25-hydroxyvitamin D [25(OH)D] to concentrations susceptible to causing toxicity. NOAEL: no observed adverse effect level; UL: tolerable upper intake level.](/cms/asset/546aa3cb-ca08-41ef-9f4e-e64abcac4cb9/ilab_a_2070595_f0008_c.jpg)
Certain diseases make patients more prone to vitamin D toxicity. Individuals suffering from idiopathic infantile hypercalcemia, lymphoma, and granulomatous disorders such as sarcoidosis, tuberculosis, leprosy, fungal diseases, infantile subcutaneous fat necrosis, giant cell polymyositis, and berylliosis are hypersensitive to vitamin D increases both from exogenous sources or endogenous synthesis [Citation290,Citation299]. In granulomatous diseases, hypervitaminosis D and hypercalcemia are the results of abnormal local synthesis of calcitriol in macrophages [Citation296,Citation300]. The rise in the active form of vitamin D in idiopathic infantile hypercalcemia patients is related to the malfunction of deactivating enzyme CYP24A1, while in patients with lymphoma, the causes of vitamin D toxicity are not yet fully understood [Citation290].
Since the number of scientific reports on the potential benefits of vitamin D on different diseases is large and continues to increase, fortified food and vitamin D supplements have become easily obtainable over-the-counter in pharmacies, supermarkets, and online stores [Citation296], and this market has been growing all over the world [Citation301]. As such, in addition to iatrogenic factors, the causes of vitamin D overdose and subsequent toxicity are mainly associated with self-medication, accidental incorrect doses (for instance due to prescribing errors), or the use of unlicensed and/or poorly standardized products [Citation289,Citation295]. The latter was associated with the urgent need for vitamin D products to be certified by highly harmonized analytical methodologies [Citation302]. Several validated methods and standardized protocols have been provided by international authorities, including the United States Pharmacopeia in the Vitamin D Assay Monography (581) [Citation303] and European Standard approved by the European Committee on Standardization [Citation304], to increase the quality of dietary supplements. These guidelines summarize analytical procedure conditions for various formulations, which are described in detail with acceptable tolerance limits [Citation305]. Nevertheless, it should be emphasized, that the regulation and quality control of dietary supplements differ in individual countries based on the current law and recommendations, thus in some cases, there is a lack of dietary supplement control that can lead to poor quality products. Indeed, two adult cases of chronic vitamin D intoxication caused by manufacturing errors have been reported [Citation289,Citation295]. In both cases, the real vitamin D level was 1000-fold higher than the daily dose level declared by the manufacturer, which points to a failure in quality control.
Figure 9. Clinical manifestations of hypervitaminosis D.
Clinical manifestations
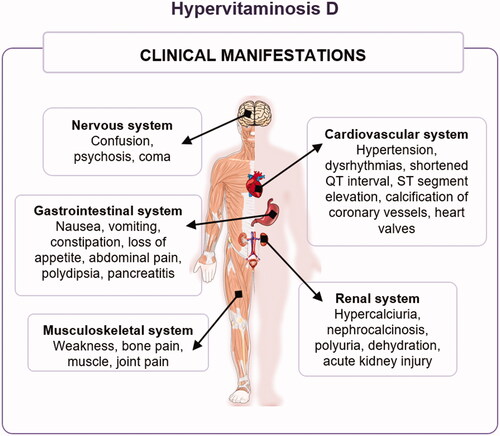
Clinical signs of vitamin D toxicity () can vary from asymptomatic to coma [Citation296]. An excessive high intake of vitamin D2/3 is correlated to increased 25(OH)D in the blood, which may lead to augmented calcium absorption by the gut and bone resorption [Citation41,Citation295]. For these reasons, the main conditions associated with hypervitaminosis D are hypercalcemia and hypercalciuria; subsequent hyperphosphatemia also commonly occurs [Citation291,Citation306].
The deleterious consequences of increasing circulating calcium vary and are specific to each organ system (). These include (i) gastrointestinal symptoms such as nausea, vomiting, constipation, loss of appetite, abdominal pain, polydipsia, and pancreatitis; (ii) other effects such as hypercalciuria, nephrocalcinosis, polyuria, dehydration, and acute kidney injury; (iii) hypertension, dysrhythmias, shortened QT interval and ST-segment elevation, calcification of coronary vessels and heart valves; (iv) musculoskeletal weakness, bone, muscle, and joint pains; as well as (v) neurological effects such as confusion, psychosis, and coma [Citation289,Citation301,Citation307].
Mechanisms of toxicity
As proposed by Jones et al. [Citation307] and supported by other authors [Citation290,Citation296,Citation301,Citation307,Citation308], there are three major theories to explain the mechanisms underlying vitamin D toxicity (). All of these theories are related to high plasma concentrations of vitamin D metabolites and the activation of nuclear VDR in the target cells, stimulating transcriptional machinery.
Figure 10. Theories proposed by Jones et al. [Citation307] for the mechanisms of toxicity of vitamin D. The first mechanism proposed for explaining vitamin D toxicity involves a plasma increase in calcitriol [1,25(OH)2D]. This active form of vitamin D has low affinity to the vitamin D binding protein (vDBP) and high affinity to the vitamin D receptor (VDR), leading to a critical increase in calcitriol in the target cells and subsequent overstimulation of the gene expression machinery. A second theory proposes an increase in plasma vitamin D metabolites to concentrations that saturate vDBP, allowing high free levels of these metabolites to enter the target cells, in particular 25-hydroxyvitamin D [25(OH)D] that has a greater affinity to VDR. The last mechanism is related to the release of calcitriol from vDBP because it has the lowest affinity for this plasma protein compared to other vitamin D metabolites. 24,25(OH)2D: 24,25-dihydroxyvitamin D; 25,26(OH)2D: 25,26-dihydroxyvitamin D; 25(OH)D-26,23-lactone: 25-hydroxyvitamin D-26,23-lactone.
![Figure 10. Theories proposed by Jones et al. [Citation307] for the mechanisms of toxicity of vitamin D. The first mechanism proposed for explaining vitamin D toxicity involves a plasma increase in calcitriol [1,25(OH)2D]. This active form of vitamin D has low affinity to the vitamin D binding protein (vDBP) and high affinity to the vitamin D receptor (VDR), leading to a critical increase in calcitriol in the target cells and subsequent overstimulation of the gene expression machinery. A second theory proposes an increase in plasma vitamin D metabolites to concentrations that saturate vDBP, allowing high free levels of these metabolites to enter the target cells, in particular 25-hydroxyvitamin D [25(OH)D] that has a greater affinity to VDR. The last mechanism is related to the release of calcitriol from vDBP because it has the lowest affinity for this plasma protein compared to other vitamin D metabolites. 24,25(OH)2D: 24,25-dihydroxyvitamin D; 25,26(OH)2D: 25,26-dihydroxyvitamin D; 25(OH)D-26,23-lactone: 25-hydroxyvitamin D-26,23-lactone.](/cms/asset/ca9b824d-0752-4516-a14e-4c16d5177d56/ilab_a_2070595_f0010_c.jpg)
The first theory involves an increase in plasma calcitriol concentration with a subsequent increase in the target cells, for instance, due to the inability to suppress the 1-hydroxylase in response to high 25(OH)D levels [Citation299]. Calcitriol has low affinity to the transport protein, vDBP, and high affinity to VDR, leading to critical overstimulation of the gene expression machinery. This hypothesis appears to be the most probable for explaining vitamin D toxicity in patients with already elevated plasma calcitriol levels (e.g. certain granulomatous disorders with unregulated 1-hydroxylase). Of note, disturbances in the calcitriol catabolic system (e.g. genetic defects in 24-hydroxylase) also make certain individuals particularly susceptible to vitamin D toxicity. The “free calcitriol concept” was clinically substantiated by Pettifor et al. [Citation309] who showed that free calcitriol was responsible for toxicity, despite no elevated plasma concentration of the active metabolite being observed in their cases of vitamin D toxicity, in line with other human and animal data.
The second theory postulates the increased plasma levels of vitamin D metabolites following vitamin D intoxication, especially 25(OH)D, to concentrations that saturate vDBP, allowing high levels of free 25(OH)D to enter the target cells. Compared to others, this metabolite has a greater affinity to VDR, stimulating gene expression in a concentration-dependent manner [Citation279].
The last hypothesis is related to the presence of vitamin D and metabolites at levels so high that vDBP is saturated [Citation273,Citation279]. Of note, in such a case, calcitriol is released from vDBP due to its lower affinity for this protein compared to other vitamin D metabolites, including 25(OH)D, 24,25(OH)2D, 25,26(OH)2D, and 25(OH)D-26,23-lactone, or even vitamin D itself, which will be found at higher concentrations after vitamin D intake. The active metabolite is then free to enter cells and bind to VDR [Citation299]. Nevertheless, Deluca et al. [Citation310] reported that both wild-type controls and 1-hydroxylase knockout mice (i.e. unable to produce calcitriol) suffered from vitamin D toxicity after a high dose of vitamin D, suggesting that other vitamin D metabolites may also contribute to hypercalcemia. In fact, VDDR type 1 patients ( or ) respond therapeutically to extreme doses of 25(OH)D [Citation311,Citation312].
Treatment of intoxication
The treatment of vitamin D toxicity includes different approaches (), taking into consideration several factors, such as the patient’s age, physiological status (e.g. pregnancy, presence of other comorbidities), and duration and severity of hypercalcemia [Citation313].
Figure 11. Treatment approaches for hypervitaminosis D.
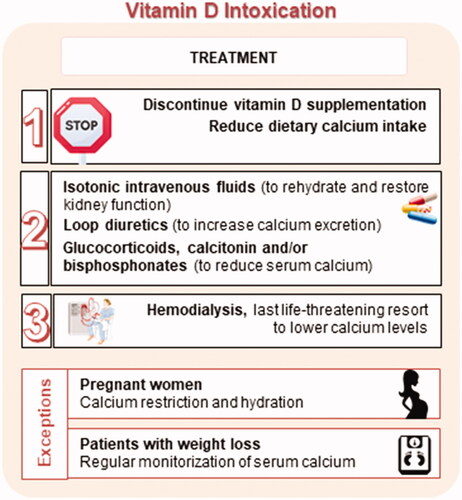
As a first-line approach, the patient should stop taking vitamin D and reduce calcium intake from the diet. For granulomatous disorders, lymphoma, and idiopathic intracranial hypertension patients, sunlight and other UV-B light exposures are not recommended [Citation290].
To treat hypercalcemia, isotonic intravenous fluids to correct dehydration and restore kidney function should be considered. Furthermore, loop diuretics (e.g. furosemide) can be added to increase calcium excretion; however, this approach has its limitations due to potential adverse reactions [Citation313]. Therapy with glucocorticoids (e.g. prednisone) can also be successfully applied to reduce serum calcium levels. These steroid hormones prevent active reabsorption of calcium in the kidneys and also alter vitamin D metabolism, favoring the synthesis of inactive metabolites, which lowers plasma calcitriol concentration and consequently reduces intestinal calcium absorption. Nevertheless, it should be noted that chronic glucocorticoid treatments are also associated with adverse effects, such as secondary osteoporosis, osteonecrosis, and muscle weakness, among others. Calcitonin and bisphosphonate therapies (e.g. pamidronate and alendronate) can be useful in severe cases to reduce calcium serum levels by inhibiting bone resorption. In some reports, bisphosphonates are described as the most effective treatment of vitamin D toxicity in children. As a last resort, when no other treatment has been successful, hemodialysis can be used to rapidly lower calcium levels [Citation314].
The treatment for pregnant women is more complicated as some of the available medicines are not indicated or are even contraindicated in this case. Therefore, it seems judicious to focus on calcium restriction and hydration [Citation315]. Importantly, vitamin D accumulates, especially after megadoses, in adipose tissue due to its lipophilicity. Consequently, in cases of significant weight loss, vitamin D is mobilized from fat and slowly released into the circulation, with toxicity symptoms such as hypercalcemia lasting for a long period of time, from several weeks to up to 18 months, even after vitamin D discontinuation [Citation290]. Hence, it is prudent to follow up with such patients regularly, paying particular attention to serum calcium levels [Citation301,Citation314].
Intoxication cases
As discussed, the reports of vitamin D intoxication present in the literature are rare, with recent publications describing mild toxicity cases [Citation316] and intoxications in adults [Citation289,Citation295]. A summary of these intoxications is presented in , with further information on cases prior to 1999 compiled in the publication from Vieth [Citation340]. Overall, the majority of vitamin D intoxications occurred in children younger than one year and consisted of accidental overdoses resulting from administration of a high dose carried out by the parents, by virtue of misinterpretation of the product label or physician instructions (e.g. erroneous administration of the product by the full milliliter dropper instead of giving it by the drop [Citation341]), as well as deliberated self-medication cases, where parents thought that vitamin administration would be beneficial and harmless to children. There are also a few reported cases concerning iatrogenic intoxications. Overall, analysis of these case reports highlights the urge to establish cutoff limits, especially for the youngest population, tightly regulate the production and sale of vitamin D products, and increase the awareness of the medical and non-medical population on the risks of vitamin D intoxication [Citation295,Citation335].
Table 3. Cases of vitamin D toxicity reported in the literature.
Analytical methods for the determination of vitamin D and its metabolites with emphasis to clinical practice
Only a few vitamin D metabolites are analyzed in common clinical practice despite the fact that more metabolites have been described in the last decades, including monohydroxylated, dihydroxylated, and trihydroxylated forms with their epimers, esters, lactones, and sulfates [Citation342]. Among them, 25(OH)D3 and 25(OH)D2 are the most frequently studied vitamin D metabolites in human serum or plasma [Citation343–345]. They were assumed as the main vitamin D status biomarkers due to their longer half-life in plasma, better stability, and relatively high concentration compared to other vitamin D metabolites [Citation345]. Other biological fluids, including saliva, urine, cerebrospinal fluid, milk, and tissues can be also used for vitamin D analysis, but they are not typically used in clinical practice [Citation346]. Despite calcitriol being the active metabolite of vitamin D, it is not recommended to be used as a biomarker due to its short half-life and tight physiological regulation of its concentration [Citation347]. In fact, Biancuzzo et al. [Citation91] demonstrated that the serum calcitriol concentration measured in volunteers did not significantly vary across the 11-week period of 1000 IU daily administration of vitamin D2 or D3. Indeed, calcitriol will be within the reference range even if the test subject is vitamin D deficient with associated symptoms. However, specific clinical scenarios may require the examination of 1,25(OH)2D plasma level, including (i) suspicion of 1α-hydroxylase deficiency, (ii) exclusion of disorders presenting as hypophosphatemic syndromes mediated by FGF23, where normal to low 1,25(OH)2D concentrations are found, from non-FGF23-mediated disorders with normal to high 1,25(OH)2D concentrations, and (iii) presence of excessive extrarenal 1α-hydroxylation by granulomatous or lymphoproliferative diseases or hereditary vitamin D-resistant rickets (VDDR type 2, see or ) where very high concentrations of calcitriol occur [Citation348]. On the other hand, as 25(OH)D3 reflects the variations resulting from both dietary intake and endogenous synthesis, it is considered a good biomarker to monitor vitamin D status in adults and children [Citation347].
The concentration of 25(OH)D2/3 in serum is often expressed as a sum of both, reflecting a total of 25(OH)D. Since the 2000s, there has been an increase in clinical testing of 25(OH)D2/3 levels in serum due to accumulated experimental and clinical evidence that vitamin D deficiency can be associated with several diseases. Thus, the Endocrine Society Task Force recommends supplementation, depending on age and clinical circumstances, when low vitamin D levels are measured via the 25(OH)D2/3 diagnostic test [Citation289,Citation349]. To allow clinical decision-making, the limit of the quantification should be <25 nmol/L and the imprecision should be <10% in the concentration range of 50–150 nmol/L, according to the guideline EP05 from the Clinical Laboratory Standard Institute that is mandatory in the clinical analysis [Citation350]. The guideline EP05 differs from guidelines typically used for the validation of bioanalytical methods such as the European Agency Medicine Guideline for bioanalytical method validation, where accuracy and precision with different requirements are defined [Citation351]. In recent years, there has been growing interest in other metabolites, including 24,25(OH)2D, and the C3-epimer of 25(OH)D, denoted 3-epi-25(OH)D [Citation346,Citation352]. In addition to free 25(OH)D circulating in plasma, the vitamin D metabolite ratio, calculated as a ratio of 24,25(OH)2D and total 25(OH)D concentrations, has been proposed as a new serum biomarker [Citation345,Citation353]. Indeed, there is a strong correlation between 24,25(OH)2D and 25(OH)D as a consequence of vitamin D metabolism, where 25(OH)D and 1,25(OH)2D are degraded by CYP24A1 into 24,25(OH)2D. The catabolism of 25(OH)D increases due to the increased activation of VDR. Thus, only small variations in 24,25(OH)2D have a dramatic effect on this ratio. Furthermore, the ratio can predict the 25(OH)D increase in response to vitamin D supplementation and can hence have clinical utility as a marker for vitamin D catabolism as well as a CYP24A1 deficiency, including various mutations [Citation353,Citation354].
In general, analysis of vitamin D in clinical routine is challenging, regardless of the method used, due to several issues such as (i) lipophilic nature of compounds with log P varying in the range of 4.49–7.03 (values predicted by ChemDraw 20.0 software), (ii) very low concentrations in biological fluids, (iii) short biological half-life in blood, (iv) binding to plasma proteins, (v) presence of multiple vitamin D metabolites as well as isobaric and isomeric interferences that can lead to inaccurate results, and (vi) chemical instability toward the light, oxygen, and temperature [Citation342,Citation344,Citation355,Citation356]. The measurement of vitamin D in serum is currently carried out using different analytical approaches such as competitive protein-binding assay (CPBA), radioimmunoassay (RIA), chemiluminescence immunoassay (CLIA), enzyme-linked immunoassay (ELISA), liquid chromatography (LC) with UV detection, and LC with mass spectrometry (LC-MS) or tandem mass spectrometry (LC-MS/MS) detection [Citation344,Citation357].
The first immunoassays enabling the measurement of a total of 25(OH)D included RIA and CPBA [Citation358,Citation359]. In RIA, 25(OH)D is separated from vDBP by an organic solvent, typically acetonitrile, and a radionucleotide is used as a tracer [Citation353,Citation357]. RIA is commercially available from DiaSorin (Italy), Tecan (Switzerland), IBL America (USA), and Demeditec (Germany) for the determination of a total of 25(OH)D in plasma. CPBA based on the competition of labeled and unlabeled ligand binding to target protein exhibited several drawbacks, including manual processing, cross-reaction with other vitamin D metabolites, need for solvent extraction and chromatography prior to the assay, and variable results. Moreover, the use of radionucleotides in RIA was found to be disadvantageous. Thus, these approaches have been mostly replaced by automatic assays that are more suitable for clinical practice [Citation353].
Currently, CLIA is designed based on the competitive assay format. 25(OH)D present in serum competes with labeled 25(OH)D conjugate for binding to the specifier, designated as a 25(OH)D binding component, which is typically an antibody against 25(OH)D or vDBP. The unbound material is removed after incubation, and a complex of conjugate-binding components is detected after the chemiluminescent reaction. The signal decreases with increasing 25(OH)D levels [Citation350,Citation353,Citation360]. Several commercial CLIA kits using different reagents are available from different vendors, including Immunodiagnostic Systems (IDS, United Kingdom, Germany), Abbott (USA), Siemens Healthineers (Germany), Tosoh (India), Roche (Germany), Fujirebio (Japan), and Bio-Ras Laboratories (USA) [Citation353]. Most of the assays measure the total amount of 25(OH)D and are not able to distinguish between D3 and D2 forms, thus they can lead to the overestimation or underestimation of results [Citation353]. It needs to be emphasized that immunoassays differ from each other in cross-reactivity with different D3 metabolites such as 25(OH)D3, calcitriol, 24,25(OH)2D3, as well as C3-epimers. ELISA is also based on a competitive design. The endogenous 25(OH)D dissociated from vDBP competes for binding with standards or controls and a fixed amount of conjugated biotin-25(OH)D with 25(OH)D antibody-coated wells. Free biotin-25(OH)D is detected after a washing step, with streptavidin-HRP using spectrophotometry. The concentration of a total 25(OH)D increases with a decrease in streptavidin-HRP [Citation355,Citation357]. Similar to CLIA, a total of 25(OH)D levels are determined and the cross-reactivity with other D metabolites is reported [Citation355,Citation360]. ELISA kits are available from Diagnostika GmbH (Germany), Beckman Coulter (USA), and ABCam (United Kingdom) [Citation353]. Calcitriol assays, including RIA and ELISA, that enable the determination of pmol/L in human serum or plasma, are also commercially available. These kits are burdened with similar problems such as overestimation and underestimation caused by cross-reactivity [Citation346]. To get more detailed information about single assays, the reader is referred to these references [Citation353,Citation355,Citation357]. Despite the mentioned disadvantages, automated immunoassays still belong among the most frequently used methods in clinical laboratories, providing very rapid results.
The first chromatography-based methods for vitamin D determination were developed in the 1970s, using both normal phases silica columns and reversed-phase C18 columns with UV detection at 254 nm [Citation361–363]. These methods enabled simultaneous determination of vitamin D2, vitamin D3, 25(OH)D2, and 25(OH)D3 in biological sample extracts. Nowadays, the C18 reversed-phase [Citation364–366] and a combination of acetonitrile with methanol in different ratios, alone or with phosphate buffer as a mobile phase in isocratic elution, are used for the simultaneous analysis of vitamin D2, vitamin D3, 25(OH)D2, and 25(OH)D3 in human serum at the selected wavelength of 265 nm. However, the selectivity in high-performance liquid chromatography with UV detection (HPLC-UV) is achieved only via chromatographic resolution of observed compounds and suffers from samples variability and nonspecific interferences such as lipids and epimers, despite the fact that methods using carefully optimized conditions can fully distinguish 25(OH)D2 and 25(OH)D3 as well as vitamin D2, vitamin D3, and the dihydroxylated metabolites. Thus, there is a risk of misinterpreting spurious peaks and overestimation of results [Citation355].
In recent years, LC-MS/MS with a triple quadrupole or quadrupole-linear ion trap has been considered the gold standard method for the quantification of vitamin D and its metabolites in human samples. The selectivity of this method is increased due to the detection of analytes based on their mass-to-charge ratio (m/z). Typically, the m/z of two different product ions, derived from a precursor, are monitored as selected reaction monitoring transitions. This data acquisition mode enables a significant increase in method selectivity. LC-MS/MS thus allows us to distinguish multiple compounds with various masses (384–428 Da) in one run compared to traditional immunoassays, which suffer from cross-reactivity between different D2 and D3 forms [Citation356,Citation367,Citation368]. Vitamin D and its metabolites are small naturally lipophilic compounds that lack protonation sites and charged groups that could facilitate ionization, which is necessary for the first step of MS detection [Citation369]. As a result, they exhibit low ionization efficiency under electrospray ionization (ESI) and atmospheric pressure chemical ionization (APCI) [Citation342]. Nevertheless, API (atmospheric pressure ionization) techniques are the most frequently used ionization techniques in vitamin D analysis. It is necessary to emphasize that LC-MS/MS is still able to provide very sensitive results, especially due to important technological advances in current instrumentation that have improved the limits of detection. A chemical derivatization step is another way to improve sensitivity. Typically, Diels-Alder derivatization with Cookson-type reagent such as 4-phenyl-1,2,4-triazoline-3,5-dione (PTAD) [Citation369–374] and 4-(4-dimethylaminophenyl)-1,2,4-triazoline-3,5-dione (DAPTAD) [Citation375] is used to introduce a polar ionizable group to the vitamin D structure, which finally enables 100- to 1000-fold increase in sensitivity [Citation356]. In addition, m/z shifts to higher values with lower chemical noise from solvent ions, and characteristic fragment ions unique to derivatized compounds have been observed [Citation356,Citation367]. However, the derivatization technique is a laborious sample preparation step, and thus it is not very suitable for routine clinical practice. The formation of two epimers after PTAD derivatization that can lead to the loss of signal has also been reported [Citation374,Citation376]. The use of atmospheric pressure photoionization (APPI) has been shown as a tool to significantly increase sensitivity compared to APCI [Citation377], but more studies are needed to confirm the results. Despite the specificity and sensitivity of LC-MS/MS, another challenge in vitamin D quantification is the presence of interfering compounds, including epimers and isobars that have the same exact mass and fragmentation pattern as the compounds being analyzed. C3-epimers of 25(OH)D are naturally produced by metabolism and they are present in all samples. Their higher concentration has been confirmed in infants, contributing to 8.7–61.1% of the total 25(OH)D [Citation352,Citation378]. Exogenous pharmaceutical compound, D3-calcitriol [1α,25(OH)2D3], and endogenous bile acid precursor, 7α-OH-4-cholestene-3-one, are two known isobaric interferences of 25(OH)D3 [Citation128,Citation379]. To avoid overestimation of 25(OH)D levels, baseline separation and sufficient resolution have to be achieved for these known interferences. Thus, various column chemistries, including pentafluorophenyl (PFP) [Citation373,Citation375,Citation380–383] and cyanopropyl [Citation384], provide different selectivity and are commonly used in practice to distinguish vitamin D forms. On the other hand, C18 is used when epimeric and isobaric forms are not being monitored [Citation365,Citation370–372,Citation374,Citation385–389]. C12 [Citation390], C8 [Citation391], and PFP-propyl [Citation371] also provide different selectivity compared to the previously mentioned chemistries but are not commonly used. A carefully optimized gradient can be also helpful to achieve separation. Typically, gradient elution is used for the separation, with methanol or acetonitrile as an organic component of the mobile phase and water with additives such as formic acid, ammonium formate, and methanoic acid as an aqueous component [Citation367]. Supercritical fluid chromatography (SFC) with MS/MS detection also has the potential to distinguish all commonly analyzed forms and C3-epimer forms in a very short time [Citation373,Citation392,Citation393]. SFC is a complementary method to LC that uses compressed nonpolar carbon dioxide as a major part of the mobile phase, usually with the addition of a polar organic modifier that affects the physical and chemical properties of the mobile phase, solubility of analytes, and selectivity of the separation [Citation394]. Despite the undoubted advantages, SFC-MS/MS is still niche in clinical applications.
Common interferences present in human plasma and serum such as lipids, phospholipids, and salts often cause ion suppression of target analytes, especially in ESI and APCI LC-MS and SFC-MS. To avoid this phenomenon, sample preparation is crucial. Protein precipitation (PP) using acetonitrile, methanol, and ethanol, alone or in combination with selective liquid-liquid extraction (LLE), supported liquid extraction (SLE), or solid-phase extraction (SPE), represent the preferable methods for sample preparation. Hexane, heptane, ethyl-acetate, methanol, chloroform, and their combinations enable direct, fast, and easy LLE or SLE from liquid sample or precipitate. SPE can be carried out in off-line or on-line modes. C8, C18, and hydrophilic-lipophilic balanced copolymer sorbents are typically used for the extraction [Citation345,Citation395,Citation396]. Finally, stable an isotopically labeled internal standard (SIL-IS) should be used for vitamin D LC-MS quantification to compensate for losses during the sample preparation step, prevent matrix effects, and increase method selectivity [Citation352,Citation368]. However, it is necessary to emphasize that SIL-IS are very expensive and some are not commercially available. The main benefits and limitations of vitamin D assays used in clinical practice are summarized in .
Table 4. Summary of the main benefits and limitations of the different immunoassays and chromatographic approaches to analyze vitamin D and its metabolitesa.
With the increasing clinical measurement and variable results between laboratories, the Vitamin D External Quality Assessment Scheme (DEQAS) was established in 1989 to reduce the variability and poor performance of the 25(OH)D assays [Citation368]. In recent years, the number of in-house optimized LC-UV and LC-MS/MS methods for vitamin D analysis has increased, followed by studies showing discrepancies such as overestimation and underestimation of 25(OH)D levels between immunoassay, HPLC, and LC-MS/MS methods [Citation397–402]. These data revealed the need to harmonize all developed procedures. The Vitamin D Standardization Program was established in 2010 to standardize laboratory 25(OH)D measurements as well as improve the accuracy and comparability of commercial or in-house assays used in clinical and research laboratories [Citation403]. Thus, vitamin D standard reference materials (SRMs) and reference method procedures (RMPs) were developed by the National Institute of Standards and Technology (NIST). NIST SRMs include solvent calibrators and pooled serum material. SRM 2972a 25(OH)D Calibration Solutions is a set of ethanolic solutions for the preparation of LC-UV and LC-MS/MS instrument calibration curves. The most recent NIST SRM for method validation is SRM 972a, which includes 25(OH)D3, 25(OH)D2, 3-epi-25(OH)D3, and 24,25(OH)2D3 in pooled serum materials at different concentration levels, and SRM 973, which contains only 25(OH)D3 and 3-epi-25(OH)D3 at high concentration levels. Both of these SRMs can also be used as quality control samples for all optimized methods. The SRMs are commercially available, and they are supplied with certified reference values assigned using NIST RPMs [Citation368,Citation403]. Vitamin D analysis in clinical practice is not an easy task. However, with the available SRMs, increasing use of the LC-MS/MS method, good laboratory practice compliance, and complex knowledge about vitamin D metabolism, it can provide very accurate, precise, and reliable results.
Conclusions
Vitamin D is one of the oldest vitamins made in living organisms, as the earliest life forms were capable of its synthesis. Since humans are able to produce vitamin D in the skin, it is often referred to as a hormone. Vitamin D is crucial for the growth and development of healthy bones from birth until death. Its major function has always been considered the control of calcium homeostasis. However, its non-calcemic functions are of crucial importance too. The ability to synthesize vitamin D in the skin should have been an evolutionary advantage for maintaining adequate plasma levels of vitamin D. However, the modern lifestyle might not be associated with sufficient endogenous synthesis of vitamin D by sun exposure. Surprisingly, even in the twenty-first century with all the knowledge and advances in medicine, we still do not know the precise quantity of vitamin D that is needed for normal homeostasis and its associated calcium-dependent and calcium-independent effects. Regardless, chronic vitamin D deficiency does not only cause rickets in children and osteomalacia/osteoporosis in adults but may have severe long-lasting adverse consequences such as the increased risk of hypertension, type 1 diabetes, multiple sclerosis, and several cancer types.
With the outburst of the SARS-CoV-2 pandemic, vitamin D came into massive interest among researchers and the lay public for its immunity-boosting effect and a number of new findings were reported. The supplementation of vitamin D may be beneficial or even necessary depending on the season and latitude. However, it must be emphasized that high doses of vitamin D are not always associated with positive impacts on human health and can very rarely lead to intoxication with serious health consequences.
In conclusion, vitamin D is crucial for overall health. These days, it might be difficult to achieve the ideal plasma level in a natural way. However, supplementation is not always associated with a benefit; therefore, supplementation and proper dosage should be discussed with a health professional.
| Abbreviations | ||
| 25(OH)D | = | 25-hydroxyvitamin D, calcifediol, or calcidiol |
| APCI | = | atmospheric pressure chemical ionization |
| APPI | = | atmospheric pressure photoionization |
| CD26/36 | = | cluster of differentiation 26/36 |
| CLIA | = | chemiluminescence immunoassay |
| CPBA | = | competitive protein-binding assay |
| DAPTAD | = | 4-(4-dimethylaminophenyl)-1,2,4-triazoline-3,5-dione |
| DEQAS | = | Vitamin D External Quality Assessment Scheme |
| ELISA | = | enzyme-linked immunoassay |
| ESI | = | electrospray ionization |
| FGF23 | = | fibroblast growth factor 23 |
| FGFR | = | fibroblast growth factor 23 receptor |
| HLA-DR | = | human leukocyte antigen DR isotype |
| HPLC | = | high performance liquid chromatography |
| HPLC-UV | = | high performance liquid chromatography with ultraviolet detection |
| HRMS | = | high resolution mass spectrometry |
| IBD | = | inflammatory bowel disease |
| ICAM1 | = | intercellular adhesion molecule 1 |
| KDIGO | = | Kidney Disease: Improving Global Outcomes |
| LC | = | liquid chromatography |
| LC-MS | = | liquid chromatography with mass spectrometry |
| LC-MS/MS | = | liquid chromatography with tandem mass spectrometry |
| LLE | = | liquid-liquid extraction |
| LRP2 | = | megalin/cubulin system |
| m/z | = | mass-to-charge ratio |
| NIST | = | National Institute of Standards and Technology |
| NPC1L1 | = | Niemann-Pick C1-like 1 |
| PHEX | = | phosphate-regulating neutral endopeptidase, X-linked |
| PMCA | = | plasma membrane calcium pump |
| PP | = | protein precipitation |
| PTAD | = | 4-phenyl-1,2,4-triazoline-3,5-dione |
| PTH | = | parathyroid hormone |
| RCT | = | randomized controlled trial |
| RIA | = | radioimmunoassay |
| RMP | = | reference method procedure |
| RXR | = | retinoid X receptor |
| SIL-IS | = | stable isotopically labeled internal standard |
| SLE | = | supported liquid extraction |
| SFC | = | supercritical fluid chromatography |
| SPE | = | solid-phase extraction |
| SR-B1 | = | scavenger receptor class B type 1 |
| SRM | = | standard reference material |
| TRPV6 | = | transient receptor potential vanilloid channel 6 |
| UV | = | ultraviolet |
| vDBP | = | vitamin D binding protein |
| VDDR | = | vitamin D-dependent rickets |
| VDR | = | vitamin D receptor |
| VDRE | = | vitamin D response element |
Disclosure statement
The authors declare no conflict of interest.
Additional information
Funding
References
- Owen I. Geographical distribution of rickets, acute and subacute rheumatism, chorea, cancer and urinary calculus in the British islands. Br Med J. 1889;1:113–116.
- Palm TA. The geographical distribution and aetiology of rickets. Practitioner. 1890;45(4):270–342.
- Trousseau A. Clinique médicale de l'Hôtel-Dieu de Paris v. 3. Vol. 3. JB Baillière1865.
- McCollum EV, Simmonds N, Becker JE, et al. Studies on experimental rickets XXI. An experimental demonstration of the existence of a vitamin which promotes calcium deposition. J Biol Chem. 1922;53(2):293–312.
- Chick H, Dalyell E, Hume M, et al. The aetiology of rickets in infants: prophylactic and curative observations at the Vienna University Kinderklinik. Lancet. 1922;200(5157):7–11.
- Huldschinsky K. Die behandlung der rachitis durch ultraviolettbestrahlung. Z Orthop Chir. 1920;39:426.
- Carpenter KJ, Zhao L. Forgotten mysteries in the early history of vitamin D. J Nutr. 1999;129(5):923–927.
- Armas LA, Hollis BW, Heaney RP. Vitamin D2 is much less effective than vitamin D3 in humans. J Clin Endocrinol Metab. 2004;89(11):5387–5391.
- Tjellesen L, Christiansen C, Rodbro P, et al. Different metabolism of vitamin D2 and vitamin D3 in epileptic patients on carbamazepine. Acta Neurol Scand. 1985;71(5):385–389.
- Tjellesen L, Gotfredsen A, Christiansen C. Different actions of vitamin D2 and D3 on bone metabolism in patients treated with phenobarbitone/phenytoin. Calcif Tissue Int. 1985;37(3):218–222.
- Tjellesen L, Hummer L, Christiansen C, et al. Serum concentration of vitamin D metabolites during treatment with vitamin D2 and D3 in normal premenopausal women. Bone Miner. 1986;1(5):407–413.
- Borel P, Caillaud D, Cano N. Vitamin D bioavailability: state of the art. Crit Rev Food Sci Nutr. 2015;55(9):1193–1205.
- Krause R, Bühring M, Hopfenmüller W, et al. Ultraviolet B and blood pressure. Lancet. 1998;352(9129):709–710.
- Stamp T, Haddad J, Twigg C. Comparison of oral 25-hydroxycholecalciferol, vitamin D, and ultraviolet light as determinants of circulating 25-hydroxyvitamin D. Lancet. 1977;1(8026):1341–1343.
- Wimalawansa SJ. Vitamin D in the new millennium. Curr Osteoporos Rep. 2012;10(1):4–15.
- Phillips KM, Horst RL, Koszewski NJ, et al. Vitamin D4 in mushrooms. PLoS One. 2012;7(8):e40702–e40702.
- Wang Y, Zhu J, DeLuca HF. Where is the vitamin D receptor? Arch Biochem Biophys. 2012;523(1):123–133.
- Christakos S, DeLuca HF. Minireview: vitamin D: is there a role in extraskeletal health? Endocrinology. 2011;152(8):2930–2936.
- Judd S, Tangpricha V. Vitamin D deficiency and risk for cardiovascular disease. Circulation. 2008;117(4):503.
- von Hurst PR, Stonehouse W, Coad J. Vitamin D supplementation reduces insulin resistance in South asian women living in New Zealand who are insulin resistant and vitamin D deficient–a randomised, placebo-controlled trial. Br J Nutr. 2010;103(4):549–555.
- Mathieu C, Gysemans C, Giulietti A, et al. Vitamin D and diabetes. Diabetologia. 2005;48(7):1247–1257.
- Garland CF, Garland FC, Gorham ED, et al. The role of vitamin D in cancer prevention. Am J Public Health. 2006;96(2):252–261.
- Jorde R, Sneve M, Figenschau Y, et al. Effects of vitamin D supplementation on symptoms of depression in overweight and obese subjects: randomized double blind trial. J Intern Med. 2008;264(6):599–609.
- Oudshoorn C, Mattace-Raso F, van der Velde N, et al. Higher serum vitamin D3 levels are associated with better cognitive test performance in patients with Alzheimer's disease. Dement Geriatr Cogn Disord. 2008;25(6):539–543.
- Burton J, Kimball S, Vieth R, et al. A phase I/II dose-escalation trial of vitamin D3 and calcium in multiple sclerosis. Neurology. 2010;74(23):1852–1859.
- Bischoff-Ferrari HA, Dawson-Hughes B, Willett WC, et al. Effect of vitamin D on falls: a meta-analysis. JAMA. 2004;291(16):1999–2006.
- Urashima M, Segawa T, Okazaki M, et al. Randomized trial of vitamin D supplementation to prevent seasonal influenza a in schoolchildren. Am J Clin Nutr. 2010;91(5):1255–1260.
- Vieth R. The pharmacology of vitamin D. In: Vitamin D. 3rd ed. Boston (MA): Elsevier Academic Press; 2011. p. 1041–1066.
- Utiger RD. The need for more vitamin D. Mass Medical Soc. 1998;338(12):828–829.
- Holick MF, Chen TC. Vitamin D deficiency: a worldwide problem with health consequences. Am J Clin Nutr. 2008;87(4):1080S–1086S.
- Holick MF, Chen TC, Lu Z, et al. Vitamin D and skin physiology: AD‐lightful story. J Bone Miner Res. 2007;22(S2):V28–V33.
- Holick MF. Environmental factors that influence the cutaneous production of vitamin D. Am J Clin Nutr. 1995;61(3 Suppl):638S–645S.
- Holick MF, MacLaughlin J, Clark M, et al. Photosynthesis of previtamin D3 in human skin and the physiologic consequences. Science. 1980;210(4466):203–205.
- Holick MF, Tian XQ, Allen M. Evolutionary importance for the membrane enhancement of the production of vitamin D3 in the skin of poikilothermic animals. Proc Natl Acad Sci USA. 1995;92(8):3124–3126.
- Whyte MP, Haddad JJ, Walters DD, et al. Vitamin D bioavailability: serum 25-hydroxyvitamin D levels in man after oral, subcutaneous, intramuscular, and intravenous vitamin D administration. J Clin Endocrinol Metab. 1979;48(6):906–911.
- Kowalówka M, Główka AK, Karaźniewicz-Łada M, et al. Clinical significance of analysis of vitamin D status in various diseases. Nutrients. 2020;12(9):2788.
- Wang T, Bengtsson G, Kärnefelt I, et al. Provitamins and vitamins D2and D3in Cladina spp. over a latitudinal gradient: possible correlation with UV levels. J Photochem Photobiol B. 2001;62(1–2):118–122.
- Jäpelt RB, Jakobsen J. Vitamin D in plants: a review of occurrence, analysis, and biosynthesis. Front Plant Sci. 2013;4:136–136.
- Göring H. Vitamin D in nature: a product of synthesis and/or degradation of cell membrane components. Biochemistry Moscow. 2018;83(11):1350–1357.
- Nakamura K, Nashimoto M, Okuda Y, et al. Fish as a major source of vitamin D in the Japanese Diet. Nutrition. 2002;18(5):415–416.
- Spiro A, Buttriss JL. Vitamin D: an overview of vitamin D status and intake in Europe. Nutr Bull. 2014;39(4):322–350.
- Bischofova S, Dofkova M, Blahova J, et al. Dietary intake of vitamin D in the Czech population: a comparison with dietary reference values, main food sources identified by a total diet study. Nutrients. 2018;10(10):1452.
- Takeuchi A, Okano T, Ayame M, et al. High-performance liquid chromatographic determination of vitamin D3 in fish liver oils and eel body oils. J Nutr Sci Vitaminol. 1984;30(5):421–430.
- Schmid A, Walther B. Natural vitamin D content in animal products. Adv Nutr. 2013;4(4):453–462.
- Byrdwell WC, Horst RL, Phillips KM, et al. Vitamin D levels in fish and shellfish determined by liquid chromatography with ultraviolet detection and mass spectrometry. J Food Compost Anal. 2013;30(2):109–119.
- Polzonetti V, Pucciarelli S, Vincenzetti S, et al. Dietary intake of vitamin D from dairy products reduces the risk of osteoporosis. Nutrients. 2020;12(6):1743.
- Lu Z, Chen TC, Zhang A, et al. An evaluation of the vitamin D3 content in fish: is the vitamin D content adequate to satisfy the dietary requirement for vitamin D? J Steroid Biochem Mol Biol. 2007;103(3–5):642–644.
- Mattila P, Piironen V, Uusi-Rauva E, et al. Cholecalciferol and 25-Hydroxycholecalciferol contents in fish and fish products. J Food Compost Anal. 1995;8(3):232–243.
- Kjerstad M, Larssen WE, Midtbø LK. Belly flap from Norwegian spring-spawning herring (Clupea harengus L.): a potentially new product with high content of vitamin D, EPA and DHA. Heliyon. 2020;6(10):e05239.
- Stancheva M, Dobreva DA. Bulgarian marine and freshwater fishes as a source of Fat-Soluble vitamins for a healthy human diet. Foods. 2013;2(3):332–337.
- Kühn J, Schutkowski A, Hirche F, et al. Non-linear increase of vitamin D content in eggs from chicks treated with increasing exposure times of ultraviolet light. J Steroid Biochem Mol Biol. 2015;148:7–13.
- Mattila P, Lehikoinen K, Kiiskinen T, et al. Cholecalciferol and 25-hydroxycholecalciferol content of chicken egg yolk as affected by the cholecalciferol content of feed. J Agric Food Chem. 1999;47(10):4089–4092.
- Thompson JN, Plouffe L. Determination of cholecalciferol in meat and fat from livestock fed normal and excessive quantities of vitamin D. Food Chem. 1993;46(3):313–318.
- Strobel N, Buddhadasa S, Adorno P, et al. Vitamin D and 25-hydroxyvitamin D determination in meats by LC-IT-MS. Food Chem. 2013;138(2–3):1042–1047.
- Mattila PH, Piironen VI, Uusi-Rauva EJ, et al. Contents of cholecalciferol, ergocalciferol, and their 25-Hydroxylated metabolites in milk products and raw meat and liver as determined by HPLC. J Agric Food Chem. 1995;43(9):2394–2399.
- Roseland JM, Patterson KY, Andrews KW, et al. Interlaboratory trial for measurement of vitamin D and 25-Hydroxyvitamin D [25(OH)D] in foods and a dietary supplement using liquid chromatography-mass spectrometry. J Agric Food Chem. 2016;64(16):3167–3175.
- EFSA Panel on Dietetic Products N, Allergies. Scientific opinion on the safety of vitamin D-enriched UV-treated baker's yeast. Efsa J. 2014;12(1):3520.
- Huang S-J, Lin C-P, Tsai S-Y. Vitamin D2 content and antioxidant properties of fruit body and mycelia of edible mushrooms by UV-B irradiation. J Food Compost Anal. 2015;42:38–45.
- Taofiq O, Fernandes Â, Barros L, et al. UV-irradiated mushrooms as a source of vitamin D2: a review. Trends Food Sci Technol. 2017;70:82–94.
- Cardwell G, Bornman JF, James AP, et al. A review of mushrooms as a potential source of dietary vitamin D. Nutrients. 2018;10(10):1498.
- Simon RR, Borzelleca JF, DeLuca HF, et al. Safety assessment of the post-harvest treatment of button mushrooms (agaricus bisporus) using ultraviolet light. Food Chem Toxicol. 2013;56:278–289.
- Rangel-Castro JI, Staffas A, Danell E. The ergocalciferol content of dried pigmented and albino cantharellus cibarius fruit bodies. Mycol Res. 2002;106(1):70–73.
- Phillips K. A nutritionally meaningful increase in vitamin D in retail mushrooms is attainable by exposure to sunlight prior to consumption. J Nutr Food Sci. 2013;3:236.
- Hernigou P, Auregan JC, Dubory A. Vitamin D: part II; cod liver oil, ultraviolet radiation, and eradication of rickets. Int Orthop. 2019;43(3):735–749.
- Réhault-Godbert S, Guyot N, Nys Y. The golden egg: nutritional value, bioactivities, and emerging benefits for human health. Nutrients. 2019;11(3):684.
- Larson-Meyer DE, Ingold BC, Fensterseifer SR, et al. Sun exposure in pigs increases the vitamin D nutritional quality of pork. PLOS ONE. 2017;12(11):e0187877–e0187877.
- Calvo MS, Whiting SJ. Survey of current vitamin D food fortification practices in the United States and Canada. J Steroid Biochem Mol Biol. 2013;136:211–213.
- Wagner D, Rousseau D, Sidhom G, et al. Vitamin D3 fortification, quantification, and long-term stability in cheddar and low-fat cheeses. J Agric Food Chem. 2008;56(17):7964–7969.
- Banville C, Vuillemard JC, Lacroix C. Comparison of different methods for fortifying cheddar cheese with vitamin D. Int Dairy J. 2000;10(5–6):375–382.
- Kaushik R, Sachdeva B, Arora S. Vitamin D2 stability in milk during processing, packaging and storage. LWT. 2014;56(2):421–426.
- Sławińska A, Fornal E, Radzki W, et al. Study on vitamin D2 stability in dried mushrooms during drying and storage. Food Chem. 2016;199:203–209.
- Ložnjak P, Jakobsen J. Stability of vitamin D3 and vitamin D2 in oil, fish and mushrooms after household cooking. Food Chem. 2018;254:144–149.
- Jakobsen J, Knuthsen P. Stability of vitamin D in foodstuffs during cooking. Food Chem. 2014;148:170–175.
- Hrncirik K. Stability of fat-soluble vitamins and PUFA in simulated shallow-frying. Lipid Technol. 2010;22(5):107–109.
- Rogerson D. Vegan diets: practical advice for athletes and exercisers. J Int Soc Sports Nutr. 2017;14:36.
- EFSA Panel on Dietetic Products N, Allergies. Scientific opinion on the safety of UV-treated bread as a novel food EFSA panel on dietetic products, nutrition and allergies. Efsa J. 2015;13(7):4148.
- Hayes A, Cashman KD. Food-based solutions for vitamin D deficiency: putting policy into practice and the key role for research. Proc Nutr Soc. 2017;76(1):54–63.
- Mangels AR. Bone nutrients for vegetarians. Am J Clin Nutr. 2014;100(suppl_1):469S–475S.
- Liu J. Vitamin D content of food and its contribution to vitamin D status: a brief overview and Australian focus. Photochem Photobiol Sci. 2012;11(12):1802–1807.
- Tso P, Fujimoto K. The absorption and transport of lipids by the small intestine. Brain Res Bull. 1991;27(3–4):477–482.
- Mulligan GB, Licata A. Taking vitamin D with the largest meal improves absorption and results in higher serum levels of 25‐hydroxyvitamin D. J Bone Miner Res. 2010;25(4):928–930.
- Rautureau M, Rambaud J. Aqueous solubilisation of vitamin D3 in normal man. Gut. 1981;22(5):393–397.
- Compston JE, Merrett AL, Hammett F, et al. Comparison of the appearance of radiolabelled vitamin D3 and 25-hydroxy-vitamin D3 in the chylomicron fraction of plasma after oral administration in man. Clin Sci. 1981;60(2):241–243.
- Sitrin MD, Bengoa JM. Intestinal absorption of cholecalciferol and 25-hydroxycholecalciferol in chronic cholestatic liver disease. Am J Clin Nutr. 1987;46(6):1011–1015.
- Maislos M, Shany S. Bile salt deficiency and the absorption of vitamin D metabolites. In vivo study in the rat. Isr J Med Sci. 1987;23(11):1114–1117.
- Leichtmann GA, Bengoa JM, Bolt M, et al. Intestinal absorption of cholecalciferol and 25-hydroxycholecalciferol in patients with both Crohn's Disease and intestinal resection. Am J Clin Nutr. 1991;54(3):548–552.
- Heubi JE, Hollis BW, Specker B, et al. Bone disease in chronic childhood cholestasis. I. Vitamin D absorption and metabolism. Hepatology. 1989;9(2):258–264.
- Farraye F, Nimitphong H, Stucchi A, et al. Use of a novel vitamin D bioavailability test demonstrates that vitamin D absorption is decreased in patients with quiescent Crohn's Disease. Inflamm Bowel Dis. 2011;17(10):2116–2121.
- Reboul E, Goncalves A, Comera C, et al. Vitamin D intestinal absorption is not a simple passive diffusion: evidences for involvement of cholesterol transporters. Mol Nutr Food Res. 2011;55(5):691–702.
- Margier M, Collet X, Le May C, et al. ABCB1 (P-glycoprotein) regulates vitamin D absorption and contributes to its transintestinal efflux. Faseb J. 2019;33(2):2084–2094.
- Holick MF, Biancuzzo RM, Chen TC, et al. Vitamin D2 is as effective as vitamin D3 in maintaining circulating concentrations of 25-hydroxyvitamin D. J Clin Endocrinol Metab. 2008;93(3):677–681.
- McDuffie JR, Calis KA, Booth SL, et al. Effects of orlistat on fat‐soluble vitamins in obese adolescents. Pharmacotherapy. 2002;22(7):814–822.
- Goncalves A, Gleize B, Bott R, et al. Phytosterols can impair vitamin D intestinal absorption in vitro and in mice. Mol Nutr Food Res. 2011;55(S2):S303–S311.
- Hernández-Romano J, Martínez-Barnetche J, Valverde-Garduño V. Polymorphisms in gene regulatory regions and their role in the physiopathology of complex disease in the post-genomic era. Salud Pública Méx. 2009;51:S455–S62.
- Lindqvist A, Sharvill J, Sharvill DE, et al. Loss-of-function mutation in carotenoid 15,15'-monooxygenase identified in a patient with hypercarotenemia and hypovitaminosis A. J Nutr. 2007;137(11):2346–2350.
- Shephard RM, Deluca HF. Plasma concentrations of vitamin D3 and its metabolites in the rat as influenced by vitamin D3 or 25-hydroxyvitamin D3 intakes. Arch Biochem Biophys. 1980;202(1):43–53.
- Lawson D, Sedrani S, Douglas J. Interrelationships in rats of tissue pools of cholecalciferol and 25-hydroxycholecalciferol formed in UV light. Biochem J. 1986;233(2):535–540.
- Dlugos D, Perrotta P, Horn W. Effects of the submarine environment on renal-stone risk factors and vitamin D metabolism. Undersea Hyperb Med. 1995;22(2):145–152.
- Vicchio D, Yergey A, O'Brien K, et al. Quantification and kinetics of 25-hydroxyvitamin D3 by isotope dilution liquid chromatography/thermospray mass spectrometry. Biol Mass Spectrom. 1993;22(1):53–58.
- Daiger SP, Schanfield MS, Cavalli-Sforza L. Group-specific component (Gc) proteins bind vitamin D and 25-hydroxyvitamin D. Proc Natl Acad Sci USA. 1975;72(6):2076–2080.
- Hirschfeld J. Immune-electrophoretic demonstration of qualitative differences in human sera and their relation to the haptoglobins. Acta Pathol Microbiol Scand. 1959;47(2):160–168.
- Bouillon R, Schuit F, Antonio L, et al. Vitamin D binding protein: a historic overview. Front Endocrinol. 2019;10:910.
- Safadi FF, Thornton P, Magiera H, et al. Osteopathy and resistance to vitamin D toxicity in mice null for vitamin D binding protein. J Clin Invest. 1999;103(2):239–251.
- Haddad JG, Matsuoka LY, Hollis BW, et al. Human plasma transport of vitamin D after its endogenous synthesis. J Clin Invest. 1993;91(6):2552–2555.
- Vieth R. Simple method for determining specific binding capacity of vitamin D-binding protein and its use to calculate the concentration of" free" 1, 25-dihydroxyvitamin D. Clin Chem. 1994;40(3):435–441.
- Haddad J, Fraser D, Lawson D. Vitamin D plasma binding protein. Turnover and fate in the rabbit. J Clin Invest. 1981;67(5):1550–1560.
- Bikle DD, Schwartz J. Vitamin D binding protein, total and free vitamin D levels in different physiological and pathophysiological conditions. Front Endocrinol. 2019;10:317.
- Moestrup SK, Verroust PJ. Megalin- and cubilin-mediated endocytosis of protein-bound vitamins, lipids, and hormones in polarized epithelia. Annu Rev Nutr. 2001;21(1):407–428.
- Willnow TE, Nykjaer A. Pathways for kidney-specific uptake of the steroid hormone 25-hydroxyvitamin D3. Curr Opin Lipidol. 2002;13(3):255–260.
- Willnow TE, Nykjaer A. Cellular uptake of steroid carrier proteins-mechanisms and implications. Mol Cell Endocrinol. 2010;316(1):93–102.
- Mawer EB, Backhouse J, Holman CA, et al. The distribution and storage of vitamin D and its metabolites in human tissues. Clin Sci. 1972;43(3):413–431.
- Rosenstreich SJ, Rich C, Volwiler W. Deposition in and release of vitamin D3 from body fat: evidence for a storage site in the rat. J Clin Invest. 1971;50(3):679–687.
- Drincic AT, Armas LA, Van Diest EE, et al. Volumetric dilution, rather than sequestration best explains the low vitamin D status of obesity. Obesity. 2012;20(7):1444–1448.
- Wortsman J, Matsuoka LY, Chen TC, et al. Decreased bioavailability of vitamin D in obesity. Am J Clin Nutr. 2000;72(3):690–693.
- Bouillon R, van Baelen H, de Moor P. 25-Hydroxyvitamin D and its binding protein in maternal and cord serum. J Clin Endocrinol Metab. 1977;45(4):679–684.
- Madden K, Feldman HA, Chun RF, et al. Critically ill children have low vitamin D–binding protein, influencing bioavailability of vitamin D. Ann Am Thorac Soc. 2015;12(11):1654–1661.
- Ghafouri B, Carlsson A, Holmberg S, et al. Biomarkers of systemic inflammation in farmers with musculoskeletal disorders; a plasma proteomic study. BMC Musculoskelet Disord. 2016;17(1):1–11.
- Smolders J, van den Ouweland J, Geven C, et al. Letter to the editor: Vitamin D deficiency in COVID-19: mixing up cause and consequence. Metab Clin Exp. 2021;115:154434.
- Cheng JB, Motola DL, Mangelsdorf DJ, et al. De-orphanization of cytochrome P450 2R1: a microsomal vitamin D 25-hydroxilase. J Biol Chem. 2003;278(39):38084–38093.
- Cheng JB, Levine MA, Bell NH, et al. Genetic evidence that the human CYP2R1 enzyme is a key vitamin D 25-hydroxylase. Proc Natl Acad Sci USA. 2004;101(20):7711–7715.
- Thacher TD, Fischer PR, Singh RJ, et al. CYP2R1 mutations impair generation of 25-hydroxyvitamin D and cause an atypical form of vitamin D deficiency. J Clin Endocrinol Metab. 2015;100(7):E1005–E1013.
- Jensen MB. Vitamin D metabolism, sex hormones, and male reproductive function. Reproduction. 2012;144(2):135–152.
- Horst RL, Koszewski NJ, Reinhardt TA. 1. Alpha.-hydroxylation of 24-hydroxyvitamin D2 represents a minor physiological pathway for the activation of vitamin D2 in mammals. Biochemistry. 1990;29(2):578–582.
- Bailie GR, Johnson CA. Comparative review of the pharmacokinetics of vitamin D analogues. Semin Dial. 2002;15(5):352–357.
- Yu OB, Arnold LA. Calcitroic acid-A review. ACS Chem Biol. 2016;11(10):2665–2672.
- Kamao M, Tatematsu S, Hatakeyama S, et al. C-3 epimerization of vitamin D3 metabolites and further metabolism of C-3 epimers: 25-hydroxyvitamin D3 is metabolized to 3-epi-25-hydroxyvitamin D3 and subsequently metabolized through C-1alpha or C-24 hydroxylation. J Biol Chem. 2004;279(16):15897–15907.
- Rehan VK, Torday JS, Peleg S, et al. 1Alpha,25-dihydroxy-3-epi-vitamin D3, a natural metabolite of 1alpha,25-dihydroxy vitamin D3: production and biological activity studies in pulmonary alveolar type II cells. Mol Genet Metab. 2002;76(1):46–56.
- Shah I, James R, Barker J, et al. Misleading measures in vitamin D analysis: a novel LC-MS/MS assay to account for epimers and isobars. Nutr J. 2011;10(1):1–9.
- Brown A, Ritter C, Slatopolsky E, et al. 25‐Dihydroxy‐3‐epi‐vitamin D3, a natural metabolite of 1α, 25‐dihydroxyvitamin D3, is a potent suppressor of parathyroid hormone secretion. J Cell Biochem. 1999;73(1):106–113.
- Astecker N, Reddy GS, Herzig G, et al. 1α, 25-Dihydroxy-3-epi-vitamin D3 a physiological metabolite of 1α, 25-dihydroxyvitamin D3: its production and metabolism in primary human keratinocytes. Mol Cell Endocrinol. 2000;170(1–2):91–101.
- Singh RJ, Taylor RL, Reddy GS, et al. C-3 epimers can account for a significant proportion of total circulating 25-Hydroxyvitamin D in infants, complicating accurate measurement and interpretation of vitamin D status. J Clin Endocrinol Metab. 2006;91(8):3055–3061.
- van den Ouweland JM, Beijers AM, van Daal H. Overestimation of 25-hydroxyvitamin D3 by increased ionisation efficiency of 3-epi-25-hydroxyvitamin D3 in LC-MS/MS methods not separating both metabolites as determined by an LC-MS/MS method for separate quantification of 25-hydroxyvitamin D3, 3-epi-25-hydroxyvitamin D3 and 25-hydroxyvitamin D2 in human serum. J Chromatogr B Analyt Technol Biomed Life Sci. 2014;967:195–202.
- Aghajafari F, Field CJ, Rabi D, et al. Plasma 3-Epi-25-Hydroxycholecalciferol can alter the assessment of vitamin D status using the current reference ranges for pregnant women and their newborns. J Nutr. 2016;146(1):70–75.
- Zbytek B, Janjetovic Z, Tuckey RC, et al. 20-Hydroxyvitamin D3, a product of vitamin D3 hydroxylation by cytochrome P450scc, stimulates keratinocyte differentiation. J Invest Dermatol. 2008;128(9):2271–2280.
- Jones G, Prosser DE, Kaufmann M. Cytochrome P450-mediated metabolism of vitamin D. J Lipid Res. 2014;55(1):13–31.
- Bikle DD. Vitamin D metabolism, mechanism of action, and clinical applications. Chem Biol. 2014;21(3):319–329.
- Christakos S, Dhawan P, Verstuyf A, et al. Vitamin D: metabolism, molecular mechanism of action, and pleiotropic effects. Physiol Rev. 2016;96(1):365–408.
- Pike JW, Christakos S. Biology and mechanisms of action of the vitamin D hormone. Endocrinol Metab Clin North Am. 2017;46(4):815–843.
- Canaff L, Hendy GN. Human calcium-sensing receptor gene. Vitamin D response elements in promoters P1 and P2 confer transcriptional responsiveness to 1,25-dihydroxyvitamin D. J Biol Chem. 2002;277(33):30337–30350.
- Demay MB, Kiernan MS, DeLuca HF, et al. Sequences in the human parathyroid hormone gene that bind the 1,25-dihydroxyvitamin D3 receptor and mediate transcriptional repression in response to 1,25-dihydroxyvitamin D3. Proc Natl Acad Sci USA. 1992;89(17):8097–8101.
- Quarles LD. Skeletal secretion of FGF-23 regulates phosphate and vitamin D metabolism. Nat Rev Endocrinol. 2012;8(5):276–286.
- Hu MC, Shiizaki K, Kuro-o M, et al. Fibroblast growth factor 23 and klotho: physiology and pathophysiology of an endocrine network of mineral metabolism. Annu Rev Physiol. 2013;75:503–533.
- Henry HL. Regulation of vitamin D metabolism. Best Pract Res Clin Endocrinol Metab. 2011;25(4):531–541.
- Tang W, Norlin M, Wikvall K. Regulation of human CYP27A1 by estrogens and androgens in HepG2 and prostate cells. Arch Biochem Biophys. 2007;462(1):13–20.
- Riggs BL, Khosla S, Melton IL. Sex steroids and the construction and conservation of the adult skeleton. Endocr Rev. 2002;23(3):279–302.
- Shao A, Wood R, Fleet J. Increased vitamin D receptor level enhances 1,25-dihydroxyvitamin D3-mediated gene expression and calcium transport in Caco-2 cells. J Bone Miner Res. 2001;16(4):615–624.
- van Abel M, Hoenderop JG, van der Kemp AW, et al. Regulation of the epithelial Ca2+ channels in small intestine as studied by quantitative mRNA detection. Am J Physiol Gastrointest Liver Physiol. 2003;285(1):G78–G85.
- Rosanoff A, Dai Q, Shapses SA. Essential nutrient interactions: does low or suboptimal magnesium status interact with vitamin D and/or calcium status? Adv Nutr. 2016;7(1):25–43.
- Glendenning P, Inderjeeth CA. Controversy and consensus regarding vitamin D: recent methodological changes and the risks and benefits of vitamin D supplementation. Crit Rev Clin Lab Sci. 2016;53(1):13–28.
- Bikle DD, Patzek S, Wang Y. Physiologic and pathophysiologic roles of extra renal CYP27b1: case report and review. Bone Rep. 2018;8:255–267.
- Chen TC, Schwartz GG, Burnstein KL, et al. The in vitro evaluation of 25-hydroxyvitamin D3 and 19-nor-1α, 25-dihydroxyvitamin D2 as therapeutic agents for prostate cancer. Clin Cancer Res. 2000;6(3):901–908.
- Reichel H, Koeffler HP, Norman AW. 25-Hydroxyvitamin D3 metabolism by human T-lymphotropic virus-transformed lymphocytes. J Clin Endocrinol Metab. 1987;65(3):519–526.
- Reichel H, Bishop JE, Koeffler HP, et al. Evidence for 1, 25-dihydroxyvitamin D3 production by cultured porcine alveolar macrophages. Mol Cell Endocrinol. 1991;75(2):163–167.
- Vieth R, McCarten K, Norwich KH. Role of 25-hydroxyvitamin D3 dose in determining rat 1,25-dihydroxyvitamin D3 production. Am J Physiol. 1990;258(5 Pt 1):E780–E789.
- Jones G, Kottler ML, Schlingmann KP. Genetic diseases of vitamin D metabolizing enzymes. Endocrinol Metab Clin North Am. 2017;46(4):1095–1117.
- Tsai K-S, Heath H, Kumar R, et al. Impaired vitamin D metabolism with aging in women. Possible role in pathogenesis of senile osteoporosis. J Clin Invest. 1984;73(6):1668–1672.
- Armbrecht H, Zenser T, Davis B. Effect of age on the conversion of 25-hydroxyvitamin D3 to 1,25-dihydroxyvitamin D3 by kidney of rat. J Clin Invest. 1980;66(5):1118–1123.
- Johnson JA, Beckman MJ, Pansini-Porta A, et al. Age and gender effects on 1, 25-dihydroxyvitamin D3-regulated gene expression. Exp Gerontol. 1995;30(6):631–643.
- Kim CJ, Kaplan LE, Perwad F, et al. Vitamin D 1alpha-hydroxylase gene mutations in patients with 1alpha-hydroxylase deficiency. J Clin Endocrinol Metab. 2007;92(8):3177–3182.
- Molin A, Wiedemann A, Demers N, et al. Vitamin D-dependent rickets type 1B (25-Hydroxylase Deficiency): a rare condition or a misdiagnosed condition? J Bone Miner Res. 2017;32(9):1893–1899.
- Roizen JD, Li D, O'Lear L, et al. CYP3A4 mutation causes vitamin D-dependent rickets type 3. J Clin Invest. 2018;128(5):1913–1918.
- Christakos S, Li S, De La Cruz J, et al. New developments in our understanding of vitamin metabolism, action and treatment. Metabolism. 2019;98:112–120.
- Pike JW, Meyer MB, Benkusky NA, et al. Genomic determinants of vitamin D-regulated gene expression. Vitamins & hormones. Vol. 100. Boston (MA): Elsevier; 2016. p. 21–44.
- Holick MF. Sunlight and vitamin D for bone health and prevention of autoimmune diseases, cancers, and cardiovascular disease. Am J Clin Nutr. 2004;80(6 Suppl):1678S–1688S.
- Holick MF. Vitamin D: the underappreciated D-lightful hormone that is important for skeletal and cellular health. Curr Opin Endocrinol Diabetes Obes. 2002;9(1):87–98.
- Christakos S, Lieben L, Masuyama R, et al. Vitamin D endocrine system and the intestine. BoneKEy Rep. 2014;3:496.
- Benn BS, Ajibade D, Porta A, et al. Active intestinal calcium transport in the absence of transient receptor potential vanilloid type 6 and calbindin-D9k. Endocrinology. 2008;149(6):3196–3205.
- Wasserman R. Vitamin D and the dual processes of intestinal calcium absorption. J Nutr. 2004;134(11):3137–3139.
- Xue Y, Fleet JC. Intestinal vitamin D receptor is required for normal calcium and bone metabolism in mice. Gastroenterology. 2009;136(4):1317–1327.
- Lee SM, Riley EM, Meyer MB, et al. 1,25-Dihydroxyvitamin D3 controls a cohort of Vitamin D receptor target genes in the proximal intestine that is enriched for calcium-regulating components. J Biol Chem. 2015;290(29):18199–18215.
- Ryan ZC, Craig TA, Filoteo AG, et al. Deletion of the intestinal plasma membrane calcium pump, isoform 1, Atp2b1, in mice is associated with decreased bone mineral density and impaired responsiveness to 1, 25-dihydroxyvitamin D3. Biochem Biophys Res Commun. 2015;467(1):152–156.
- Glendenning P, Ratajczak T, Dick IM, et al. Calcitriol upregulates expression and activity of the 1b isoform of the plasma membrane calcium pump in immortalized distal kidney tubular cells. Arch Biochem Biophys. 2000;380(1):126–132.
- Ajibade D, Benn BS, Christakos S. Mechanism of action of 1, 25-Dihydroxyvitamin D 3 on intestinal calcium absorption and renal calcium transport. Vitamin D. New York (NY): Springer; 2010. p. 175–187.
- Yasuda H, Shima N, Nakagawa N, et al. Osteoclast differentiation factor is a ligand for osteoprotegerin/osteoclastogenesis-inhibitory factor and is identical to TRANCE/RANKL. Proc Natl Acad Sci USA. 1998;95(7):3597–3602.
- Clairmont A, Tessman D, Stock A, et al. Induction of gap junctional intercellular communication by vitamin D in human skin fibroblasts is dependent on the nuclear Induction of gap junctional intercellular communication by vitamin D in human skin fibroblasts is dependent on the nuclear vitamin D receptor. Carcinogenesis. 1996;17(6):1389–1391.
- Gniadecki R. Vitamin D–a modulator of cell proliferation. Retinoids. 1997;13:55–59.
- Pálmer HG, González-Sancho JM, Espada J, et al. Vitamin D(3) promotes the differentiation of colon carcinoma cells by the induction of E-cadherin and the inhibition of beta-catenin signaling. J Cell Biol. 2001;154(2):369–388.
- Schwalfenberg GK. A review of the critical role of vitamin D in the functioning of the immune system and the clinical implications of vitamin D deficiency. Mol Nutr Food Res. 2011;55(1):96–108.
- Wei R, Christakos S. Mechanisms underlying the regulation of innate and adaptive immunity by vitamin D. Nutrients. 2015;7(10):8251–8260.
- Vandamme D, Landuyt B, Luyten W, et al. A comprehensive summary of LL-37, the factotum human cathelicidin peptide. Cell Immunol. 2012;280(1):22–35.
- Cohen MS, Mesler DE, Snipes RG, et al. 1, 25-Dihydroxyvitamin D3 activates secretion of hydrogen peroxide by human monocytes. J Immunol. 1986;136(3):1049–1053.
- Vanherwegen A-S, Gysemans C, Mathieu C. Regulation of immune function by vitamin D and its use in diseases of immunity. Endocrinol Metab Clin North Am. 2017;46(4):1061–1094.
- Toelzer C, Gupta K, Yadav SK, et al. Free fatty acid binding pocket in the locked structure of SARS-CoV-2 spike protein. Science. 2020;370(6517):725–730.
- Shoemark DK, Colenso CK, Toelzer C, et al. Molecular simulations suggest vitamins, retinoids and steroids as ligands of the free fatty acid pocket of the SARS‐CoV‐2 spike protein. Angew Chem. 2021;133(13):7174–7186.
- Welsh J. Cellular and molecular effects of vitamin D on carcinogenesis. Arch Biochem Biophys. 2012;523(1):107–114.
- Schön MP, Boehncke W-H. Psoriasis. N Engl J Med. 2005;352(18):1899–1912.
- Soleymani T, Hung T, Soung J. The role of vitamin D in psoriasis: a review. Int J Dermatol. 2015;54(4):383–392.
- Trémezaygues L, Reichrath J. Vitamin D analogs in the treatment of psoriasis: where are we standing and where will we be going? Dermatoendocrinol. 2011;3(3):180–186.
- Savoia P, Novelli M, De Matteis A, et al. Effects of topical calcipotriol on the expression of adhesion molecules in psoriasis. J Cutan Pathol. 1998;25(2):89–94.
- Webb AR, Kline L, Holick MF. Influence of season and latitude on the cutaneous synthesis of vitamin D3: exposure to winter sunlight in Boston and Edmonton will not promote vitamin D3 synthesis in human skin. J Clin Endocrinol Metab. 1988;67(2):373–378.
- Clemens T, Henderson S, Adams J, et al. Increased skin pigment reduces the capacity of skin to synthesise vitamin D3. Lancet. 1982;1(8263):74–76.
- Sahay M, Sahay R. Rickets-vitamin D deficiency and dependency. Indian J Endocrinol Metab. 2012;16(2):164–176.
- Matsuoka LY, Ide L, Wortsman J, et al. Sunscreens suppress cutaneous vitamin D3 synthesis. J Clin Endocrinol Metab. 1987;64(6):1165–1168.
- Klein GL, Chen TC, Holick MF, et al. Synthesis of vitamin D in skin after burns. Lancet. 2004;363(9405):291–292.
- MacLaughlin J, Holick MF. Aging decreases the capacity of human skin to produce vitamin D3. J Clin Invest. 1985;76(4):1536–1538.
- Dibble J, Sheridan P, Losowsky M. A survey of vitamin D deficiency in gastrointestinal and liver disorders. Q J Med. 1984;53(209):119–134.
- American gastroenterological association medical position statement: guidelines on osteoporosis in gastrointestinal diseases. Gastroenterology. 2003;124(3):791–794.
- Elder GJ, Mackun K. 25‐Hydroxyvitamin D deficiency and diabetes predict reduced BMD in patients with chronic kidney disease. J Bone Miner Res. 2006;21(11):1778–1784.
- Vaziri N. Endocrinological consequences of the nephrotic syndrome. Am J Nephrol. 1993;13(5):360–364.
- Goldstein DA, Haldimann B, Sherman D, et al. Vitamin D metabolites and calcium metabolism in patients with nephrotic syndrome and normal renal function. J Clin Endocrinol Metab. 1981;52(1):116–121.
- Takeda E, Yamamoto H, Taketani Y, et al. Vitamin D-dependent rickets type I and type II. Acta Paediatr Jpn. 1997;39(4):508–513.
- Shimada T, Mizutani S, Muto T, et al. Cloning and characterization of FGF23 as a causative factor of tumor-induced osteomalacia. Proc Natl Acad Sci USA. 2001;98(11):6500–6505.
- Perwad F, Azam N, Zhang MY, et al. Dietary and serum phosphorus regulate fibroblast growth factor 23 expression and 1, 25-dihydroxyvitamin D metabolism in mice. Endocrinology. 2005;146(12):5358–5364.
- Cleve H, Constans J. The mutants of the vitamin-D-binding protein: more than 120 variants of the GC/DBP system. Vox Sang. 1988;54(4):215–225.
- Arnaud J, Constans J. Affinity differences for vitamin D metabolites associated with the genetic isoforms of the human serum carrier protein (DBP). Hum Genet. 1993;92(2):183–188.
- Schwartz JB, Gallagher JC, Jorde R, et al. Determination of free 25 (OH) D concentrations and their relationships to total 25 (OH) D in multiple clinical populations. J Clin Endocrinol Metab. 2018;103(9):3278–3288.
- Thongthai P, Chailurkit L-o, Chanprasertyothin S, et al. Vitamin D binding protein gene polymorphism as a risk factor for vitamin D deficiency in Thais. Endocr Pract. 2015;21(3):221–225.
- Econs MJ. Disorders of phosphate metabolism: autosomal dominant hypophosphatemic rickets, tumor induced osteomalacia, fibrous dysplasia, and the pathophysiological relevance of FGF23. Vitamin D. 2nd ed. Boston (MA): Elsevier Academic Press; 2005. p. 1189–1195.
- Silverberg SJ. Vitamin D deficiency and primary hyperparathyroidism. J Bone Miner Res. 2007;22(S2):V100–V104.
- Xu M-Y, Cao B, Yin J, et al. Vitamin D and graves’ disease: a meta-analysis update. Nutrients. 2015;7(5):3813–3827.
- Planck T, Shahida B, Malm J, et al. Vitamin D in graves disease: levels, correlation with laboratory and clinical parameters, and genetics. Eur Thyroid J. 2018;7(1):27–33.
- Hollis BW, Wagner CL. Vitamin D requirements during lactation: high-dose maternal supplementation as therapy to prevent hypovitaminosis D for both the mother and the nursing infant. Am J Clin Nutr. 2004;80(6 Suppl):1752S–1758S.
- Holick MF. Resurrection of vitamin D deficiency and rickets. J Clin Invest. 2006;116(8):2062–2072.
- Sotaniemi EA, Hakkarainen HK, Puranen JA, et al. Radiologic bone changes and hypocalcemia with anticonvulsant therapy in epilepsy. Ann Intern Med. 1972;77(3):389–394.
- Hahn TJ. 6 Drug-induced disorders of vitamin D and mineral metabolism. Clin Endocrinol Metab. 1980;9(1):107–129.
- Holick MF. High prevalence of vitamin D inadequacy and implications for health. Mayo Clin Proc. 2006;81(3):353–373.
- Dawson-Hughes B, Heaney RP, Holick MF, et al. Estimates of optimal vitamin D status. Osteoporos Int. 2005;16(7):713–716.
- Hollis BW. Circulating 25-hydroxyvitamin D levels indicative of vitamin D sufficiency: implications for establishing a new effective dietary intake recommendation for vitamin D. J Nutr. 2005;135(2):317–322.
- Chapuy M-C, Preziosi P, Maamer M, et al. Prevalence of vitamin D insufficiency in an adult normal population. Osteoporos Int. 1997;7(5):439–443.
- Heaney RP, Dowell MS, Hale CA, et al. Calcium absorption varies within the reference range for serum 25-hydroxyvitamin D. J Am Coll Nutr. 2003;22(2):142–146.
- Holick MF. Vitamin D deficiency. N Engl J Med. 2007;357(3):266–281.
- Robinson PD, Högler W, Craig ME, et al. The re-emerging burden of rickets: a decade of experience from sydney. Arch Dis Child. 2005;91(7):564–568.
- Chapuy MC, Arlot ME, Duboeuf F, et al. Vitamin D3 and calcium to prevent hip fractures in elderly women. N Engl J Med. 1992;327(23):1637–1642.
- Gloth FM, Lindsay JM, Zelesnick LB, et al. Can vitamin D deficiency produce an unusual pain syndrome? Arch Intern Med. 1991;151(8):1662–1664.
- Malabanan AO, Turner AK, Holick MF. Severe generalized bone pain and osteoporosis in a premenopausal black female: effect of vitamin D replacement. J Clin Densitom. 1998;1(2):201–204.
- Najada AS, Habashneh MS, Khader M. The frequency of nutritional rickets among hospitalized infants and its relation to respiratory diseases. J Trop Pediatr. 2004;50(6):364–368.
- Zasloff M. Fighting infections with vitamin D. Nat Med. 2006;12(4):388–390.
- Zanetti M. Cathelicidins, multifunctional peptides of the innate immunity. J Leukoc Biol. 2004;75(1):39–48.
- Oda Y, Tu C-L, Menendez A, et al. Vitamin D and calcium regulation of epidermal wound healing. J Steroid Biochem Mol Biol. 2016;164:379–385.
- Ceglia L, Harris SS. Vitamin D and its role in skeletal muscle. Calcif Tissue Int. 2013;92(2):151–162.
- Dawson-Hughes B. Vitamin D and muscle function. J Steroid Biochem Mol Biol. 2017;173:313–316.
- Aksu Cerman A, Sarikaya Solak S, Kivanc Altunay I. Vitamin D deficiency in alopecia areata. Br J Dermatol. 2014;170(6):1299–1304.
- Föcker M, Antel J, Ring S, et al. Vitamin D and mental health in children and adolescents. Eur Child Adolesc Psychiatry. 2017;26(9):1043–1066.
- Mazess RB, Bischoff‐Ferrari HA, Dawson‐Hughes B. Vitamin D: bolus is bogus-A narrative review. JBMR Plus. 2021;5(12):e10567.
- Vieth R. How to optimize vitamin D supplementation to prevent cancer, based on cellular adaptation and hydroxylase enzymology. Anticancer Res. 2009;29(9):3675–3684.
- Martindale SS. The Complete Drug Reference.: London: Pharmaceutical Press. (electronic version), IBM Watson Health (Healthcare), Greenwood Village, Colorado, USA; [cited 2021 Mar 15]. Available from: https://www.micromedexsolutions.com/
- Pazirandeh S, Burns DL. Overview of vitamin D. 2019 [cited 2021 Mar 30]. Available from: https://www.uptodate.com/contents/overview-of-vitamin-d
- Amrein K, Scherkl M, Hoffmann M, et al. Vitamin D deficiency 2.0: an update on the current status worldwide. Eur J Clin Nutr. 2020;74(11):1498–1513.
- Tan ML, Abrams SA, Osborn DA. Vitamin D supplementation for term breastfed infants to prevent vitamin D deficiency and improve bone health. Cochrane Database Syst Rev. 2020;(12):CD013046.
- Shoback D. Clinical practice. Hypoparathyroidism. N Engl J Med. 2008;359(4):391–403.
- Vaitsi KD, Anagnostis P, Veneti S, et al. Preoperative vitamin D deficiency is a risk factor for postthyroidectomy hypoparathyroidism: a systematic review and meta-analysis of observational studies. J Clin Endocrinol Metab. 2021;106(4):1209–1224.
- Brown AJ, Coyne DW. Vitamin D analogs: new therapeutic agents for secondary hyperparathyroidism. Treat Endocrinol. 2002;1(5):313–327.
- Cunningham J. New vitamin D analogues for osteodystrophy in chronic kidney disease. Pediatr Nephrol. 2004;19(7):705–708.
- Kanis J, Burlet N, Cooper C, et al. European guidance for the diagnosis and management of osteoporosis in postmenopausal women. Osteoporos Int. 2008;19(4):399–428.
- Avenell A, Mak JC, O'Connell D. Vitamin D and vitamin D analogues for preventing fractures in post‐menopausal women and older men. Cochrane Database Syst Rev. 2014;(4):CD000227.
- Aspray TJ, Bowring C, Fraser W, et al. National osteoporosis society vitamin D guideline summary. Age Ageing. 2014;43(5):592–595.
- Kahwati LC, Weber RP, Pan H, et al. Vitamin D, calcium, or combined supplementation for the primary prevention of fractures in community-dwelling adults: evidence report and systematic review for the US preventive services task force. JAMA. 2018;319(15):1600–1612.
- Cianferotti L, Cricelli C, Kanis JA, et al. The clinical use of vitamin D metabolites and their potential developments: a position statement from the European Society for Clinical and Economic Aspects of Osteoporosis and Osteoarthritis (ESCEO) and the International Osteoporosis Foundation (IOF). Endocrine. 2015;50(1):12–26.
- Reid IR, Bolland MJ, Grey A. Effects of vitamin D supplements on bone mineral density: a systematic review and meta-analysis. Lancet. 2014;383(9912):146–155.
- Deng J, Silver Z, Huang E, et al. The effect of calcium and vitamin D compounds on bone mineral density in patients undergoing glucocorticoid therapies: a network meta-analysis. Clin Rheumatol. 2021;40(2):725–734.
- IBM Watson Health GV, Colorado, USA. Micromedex® (electronic version) [cited 2021 Mar 15]. Available from: https://www.micromedexsolutions.com/
- Mithal A, Bonjour J-P, Boonen S, et al. Impact of nutrition on muscle mass, strength, and performance in older adults. Osteoporos Int. 2013;24(5):1555–1566.
- Smith LM, Gallagher JC, Suiter C. Medium doses of daily vitamin D decrease falls and higher doses of daily vitamin D3 increase falls: a randomized clinical trial. J Steroid Biochem Mol Biol. 2017;173:317–322.
- Group KDIGOC-MW. KDIGO clinical practice guideline for the diagnosis, evaluation, prevention, and treatment of Chronic Kidney Disease-Mineral and bone disorder (CKD-MBD). Kidney Int Suppl. 2009;76(113):S1–S130.
- Wheeler DC, Winkelmayer WC. KDIGO 2017 clinical practice guideline update for the diagnosis, evaluation, prevention, and treatment of Chronic Kidney Disease-Mineral and Bone Disorder (CKD-MBD) foreword. Kidney Int Suppl. 2017;7(1):1–59.
- Christodoulou M, Aspray TJ, Schoenmakers I. Vitamin D supplementation for patients with chronic kidney disease: a systematic review and meta-analyses of trials investigating the response to supplementation and an overview of guidelines. Calcif Tissue Int. 2021;109(2):157–178.
- Uhlig K, Berns JS, Kestenbaum B, et al. KDOQI US commentary on the 2009 KDIGO clinical practice guideline for the diagnosis, evaluation, and treatment of CKD-Mineral and Bone Disorder (CKD-MBD). Am J Kidney Dis. 2010;55(5):773–799.
- Health NIF, Excellence C. Psoriasis: assessment and management. National Institute for Health and Care Excellence (NICE); 2017.
- McCullough PJ, McCullough WP, Lehrer D, et al. Oral and topical vitamin D, sunshine, and UVB phototherapy safely control psoriasis in patients with normal pretreatment serum 25-Hydroxyvitamin D concentrations: a literature review and discussion of health implications. Nutrients. 2021;13(5):1511.
- Banerjee A, Ganguly U, Saha S, et al. Vitamin D and immuno-pathology of COVID-19: many interactions but uncertain therapeutic benefits. Expert Rev anti Infect Ther. 2021;19(10):1245–1258.
- Nonnecke B, McGill J, Ridpath J, et al. Acute phase response elicited by experimental bovine diarrhea virus (BVDV) infection is associated with decreased vitamin D and E status of vitamin-replete preruminant calves. J Dairy Sci. 2014;97(9):5566–5579.
- Martineau AR, Jolliffe DA, Hooper RL, et al. Vitamin D supplementation to prevent acute respiratory tract infections: systematic review and meta-analysis of individual participant data. BMJ. 2017;356:i6583.
- Lee C. Controversial effects of vitamin D and related genes on viral infections, pathogenesis, and treatment outcomes. Nutrients. 2020;12(4):962.
- Pilz S, Gaksch M, Kienreich K, et al. Effects of vitamin D on blood pressure and cardiovascular risk factors: a randomized controlled trial. Hypertension. 2015;65(6):1195–1201.
- Nudy M, Krakowski G, Ghahramani M, et al. Vitamin D supplementation, cardiac events and stroke: a systematic review and meta-regression analysis. Int J Cardiol Heart Vasc. 2020;28:100537.
- Barbarawi M, Kheiri B, Zayed Y, et al. Vitamin D supplementation and cardiovascular disease risks in more than 83 000 individuals in 21 randomized clinical trials: a meta-analysis. JAMA Cardiol. 2019;4(8):765–776.
- Zittermann A, Trummer C, Theiler-Schwetz V, et al. Vitamin D and cardiovascular disease: an updated narrative review. IJMS. 2021;22(6):2896.
- Naghedi A, Haghaninejad H, Varastehravan H, et al. Effect of vitamin D supplements on left ventricular ejection fraction in patients with heart failure: a systematic review and meta-analysis of randomized controlled trials. Rev Port Cardiol. 2021;40(6):447–455.
- Wang J, Zhou JJ, Robertson GR, et al. Vitamin D in vascular calcification: a double-edged sword? Nutrients. 2018;10(5):652.
- Gubatan J, Chou ND, Nielsen OH, et al. Systematic review with meta-analysis: association of vitamin D status with clinical outcomes in adult patients with inflammatory bowel disease. Aliment Pharmacol Ther. 2019;50(11–12):1146–1158.
- Kaplan GG. The global burden of IBD: from 2015 to 2025. Nat Rev Gastroenterol Hepatol. 2015;12(12):720–727.
- Pappa HM, Grand RJ, Gordon CM. Report on the vitamin D status of adult and pediatric patients with inflammatory bowel disease and its significance for bone health and disease. Inflamm Bowel Dis. 2006;12(12):1162–1174.
- Guzman-Prado Y, Samson O, Segal JP, et al. Vitamin D therapy in adults with inflammatory bowel disease: a systematic review and meta-analysis. Inflamm Bowel Dis. 2020;26(12):1819–1830.
- Lamb CA, Kennedy NA, Raine T, et al. British society of gastroenterology consensus guidelines on the management of inflammatory bowel disease in adults. Gut. 2019;68(Suppl 3):s1–s106.
- Nguyen Y, Sigaux J, Letarouilly J-G, et al. Efficacy of oral vitamin supplementation in inflammatory rheumatic disorders: a systematic review and meta-analysis of randomized controlled trials. Nutrients. 2020;13(1):107.
- Autier P, Boniol M, Pizot C, et al. Vitamin D status and ill health: a systematic review. Lancet Diabetes Endocrinol. 2014;2(1):76–89.
- Zgaga L, Theodoratou E, Farrington SM, et al. Plasma vitamin D concentration influences survival outcome after a diagnosis of colorectal cancer. J Clin Oncol. 2014;32(23):2430–2439.
- Ma Y, Zhang P, Wang F, et al. Association between vitamin D and risk of colorectal cancer: a systematic review of prospective studies. J Clin Oncol. 2011;29(28):3775–3782.
- Zgaga L, Agakov F, Theodoratou E, et al. Model selection approach suggests causal association between 25-hydroxyvitamin D and colorectal cancer. PLoS One. 2013;8(5):e63475.
- Maalmi H, Walter V, Jansen L, et al. Association between blood 25-hydroxyvitamin D levels and survival in colorectal cancer patients: an updated systematic review and meta-analysis. Nutrients. 2018;10(7):896.
- Yuan C, Ng K. Vitamin D supplementation: a potential therapeutic agent for metastatic colorectal cancer. Br J Cancer. 2020;123(8):1205–1206.
- Vaughan-Shaw PG, Buijs LF, Blackmur JP, et al. The effect of vitamin D supplementation on survival in patients with colorectal cancer: systematic review and meta-analysis of randomised controlled trials. Br J Cancer. 2020;123(11):1705–1712.
- Xu Y, Qian M, Hong J, et al. The effect of vitamin D on the occurrence and development of colorectal cancer: a systematic review and meta-analysis. Int J Colorectal Dis. 2021;36(7):1329–1344.
- Manson JE, Cook NR, Lee I-M, et al. Vitamin D supplements and prevention of cancer and cardiovascular disease. N Engl J Med. 2019;380(1):33–44.
- Shahvazi S, Soltani S, Ahmadi SM, et al. The effect of vitamin D supplementation on prostate cancer: a systematic review and meta-analysis of clinical trials. Horm Metab Res. 2019;51(1):11–21.
- Nair-Shalliker V, Bang A, Egger S, et al. Post-treatment levels of plasma 25-and 1, 25-dihydroxy vitamin D and mortality in men with aggressive prostate cancer. Sci Rep. 2020;10(1):1–11.
- Datta M, Schwartz GG. Calcium and vitamin D supplementation during androgen deprivation therapy for prostate cancer: a critical review. Oncologist. 2012;17(9):1171–1179.
- Kim TJ, Koo KC. Pathophysiology of bone loss in patients with prostate cancer receiving androgen-deprivation therapy and lifestyle modifications for the management of bone health: a comprehensive review. Cancers. 2020;12(6):1529.
- Galior K, Grebe S, Singh R. Development of vitamin D toxicity from overcorrection of vitamin D deficiency: a review of case reports. Nutrients. 2018;10(8):953.
- Marcinowska-Suchowierska E, Kupisz-Urbańska M, Łukaszkiewicz J, et al. Vitamin D toxicity–a clinical perspective. Front Endocrinol. 2018;9:550.
- Kennel KA, Drake MT, Hurley DL. Vitamin D deficiency in adults: when to test and how to treat. Mayo Clin Proc. 2010;85(8):752–757.
- Lee JP, Tansey M, Jetton JG, et al. Vitamin D toxicity: a 16-year retrospective study at an academic medical center. Lab Med. 2018;49(2):123–129.
- Bouillon R. Comparative analysis of nutritional guidelines for vitamin D. Nat Rev Endocrinol. 2017;13(8):466–479.
- Dudenkov DV, Yawn BP, Oberhelman SS, et al. Changing incidence of serum 25-hydroxyvitamin D values above 50 ng/mL: a 10-year population-based study. Mayo Clin Proc. 2015;90(5):877–886.
- Taylor PN, Davies JS. A review of the growing risk of vitamin D toxicity from inappropriate practice. Br J Clin Pharmacol. 2018;84(6):1121–1127.
- Lim K, Thadhani R. Vitamin D toxicity. J Bras Nefrol. 2020;42(2):238–244.
- Holick MF, Binkley NC, Bischoff-Ferrari HA, et al. Evaluation, treatment, and prevention of vitamin D deficiency: an endocrine society clinical practice guideline. J Clin Endocrinol Metab. 2011;96(7):1911–1930.
- Ross AC, Manson JE, Abrams SA, et al. The 2011 report on dietary reference intakes for calcium and vitamin D from the institute of medicine: what clinicians need to know. J Clin Endocrinol Metab. 2011;96(1):53–58.
- Vieth R. The mechanisms of vitamin D toxicity. Bone Miner. 1990;11(3):267–272.
- Ketha H, Wadams H, Lteif A, et al. Iatrogenic vitamin D toxicity in an infant–a case report and review of literature. J Steroid Biochem Mol Biol. 2015;148:14–18.
- Misgar RA, Sahu D, Bhat MH, et al. Vitamin D toxicity: a prospective study from a Tertiary Care Centre in Kashmir Valley. Indian J Endocrinol Metab. 2019;23(3):363–366.
- Sarma N, Giancaspro G, Venema J. Dietary supplements quality analysis tools from the United States pharmacopeia. Drug Test Analysis. 2016;8(3–4):418–423.
- Vitamin D assay. The United States Pharmacopeia – the national formulary. Rockville: United States Pharmacopeial Convention, Inc.; 2021.
- Foodstuffs-Determination of vitamin D by high performance liquid Chromatography—Measurement of cholecalciferol (D3) or ergocalciferol (D2). Brussels: Comité Européen de Normalisation; 2009. Standard No.: CEN 12821.
- Temova Rakuša Ž, Roškar R. Vitamin D in supplements and medicines. In Yaseen Sofi N, Mandal A, Amiri W, editors. Vitamin D deficiency: causes & treatment. Las Vegas, NV, USA: OPEN ACCESS EBOOKS; 2018. p. 1–19.
- Deb S, Reeves AA, Lafortune S. Simulation of physicochemical and pharmacokinetic properties of vitamin D3 and its natural derivatives. Pharmaceuticals. 2020;13(8):160.
- Alshahrani F, Aljohani N. Vitamin D: deficiency, sufficiency and toxicity. Nutrients. 2013;5(9):3605–3616.
- Jones G. Pharmacokinetics of vitamin D toxicity. Am J Clin Nutr. 2008;88(2):582S–586S.
- Pettifor JM, Bikle DD, Cavaleros M, et al. Serum levels of free 1,25-dihydroxyvitamin D in vitamin D toxicity. Ann Intern Med. 1995;122(7):511–513.
- DeLuca HF, Prahl JM, Plum LA. 1,25-Dihydroxyvitamin D is not responsible for toxicity caused by vitamin D or 25-hydroxyvitamin D. Arch Biochem Biophys. 2011;505(2):226–230.
- Acar S, Demir K, Shi Y. Genetic causes of rickets. J Clin Res Pediatr Endocrinol. 2017;9(Suppl 2):88–105.
- Levine MA. Diagnosis and management of vitamin D dependent rickets. Front Pediatr. 2020;8:315.
- Waterbury S. Implications of vitamin D toxicity & deficiency. Nurse Pract. 2018;43(5):22–30.
- Vogiatzi MG, Jacobson-Dickman E, DeBoer M. Vitamin D supplementation and risk of toxicity in pediatrics: a review of current literature. J Clin Endocrinol Metab. 2014;99(4):1132–1141.
- Tebben PJ, Singh RJ, Kumar R. Vitamin D-mediated hypercalcemia: mechanisms, diagnosis, and treatment. Endocr Rev. 2016;37(5):521–547.
- Spiller HA, Good TF, Spiller NE, et al. Vitamin D exposures reported to US poison centers 2000-2014: temporal trends and outcomes. Hum Exp Toxicol. 2016;35(5):457–461.
- Allen SH, Shah JH. Calcinosis and metastatic calcification due to vitamin D intoxication. A case report and review. Horm Res. 1992;37(1–2):68–77.
- Alonso Canal L, Ruiz Herrero J, Villalobos Reales J, et al. [Vitamin D intoxication in infants born from Latin-American immigrants. Series of 3 cases]. An Pediatr. 2011;74(6):409–412.
- Atabek ME, Pirgon O, Sert A. Oral alendronate therapy for severe vitamin D intoxication of the infant with nephrocalcinosis. J Pediatr Endocrinol Metab. 2006;19(2):169–172.
- Barrueto F, Wang-Flores HH, Howland MA, et al. Acute vitamin D intoxication in a child. Pediatrics. 2005;116(3):e453–e456.
- Bereket A, Erdogan T. Oral bisphosphonate therapy for vitamin D intoxication of the infant. Pediatrics. 2003;111(4 Pt 1):899–901.
- Çağlar A, Çağlar HT. Vitamin D intoxication due to misuse: 5-year experience. Arch Pediatr. 2021;28(3):222–225.
- Chambellan-Tison C, Horen B, Plat-Wilson G, et al. [Severe hypercalcemia due to vitamin D intoxication]. Arch Pediatr. 2007;14(11):1328–1332.
- Chatterjee M, Speiser PW. Pamidronate treatment of hypercalcemia caused by vitamin D toxicity. J Pediatr Endocrinol Metab. 2007;20(11):1241–1248.
- Conti G, Chirico V, Lacquaniti A, et al. Vitamin D intoxication in two brothers: be careful with dietary supplements. J Pediatr Endocrinol Metab. 2014;27(7–8):763–767.
- Davies M, Adams P. The continuing risk of vitamin-D intoxication. Lancet. 1978;2(8090):621–623.
- Ezgu FS, Buyan N, Gündüz M, et al. Vitamin D intoxication and hypercalcaemia in an infant treated with pamidronate infusions. Eur J Pediatr. 2004;163(3):163–165.
- Feige J, Salmhofer H, Hecker C, et al. Life-threatening vitamin D intoxication due to intake of ultra-high doses in multiple sclerosis: a note of caution. Mult Scler. 2019;25(9):1326–1328.
- Gurkan F, Davutoglu M, Bosnak M, et al. Pamidronate treatment in acute vitamin D intoxication. J Endocrinol Invest. 2004;27(7):680–682.
- Joshi R. Hypercalcemia due to hypervitaminosis D: report of seven patients. J Trop Pediatr. 2009;55(6):396–398.
- Kara C, Gunindi F, Ustyol A, et al. Vitamin D intoxication due to an erroneously manufactured dietary supplement in seven children. Pediatrics. 2014;133(1):e240–e244.
- Marins TA, Galvão TdFG, Korkes F, et al. Vitamin D intoxication: case report. Einstein. 2014;12(2):242–244.
- Mawer EB, Hann J, Berry JL, et al. Vitamin D metabolism in patients intoxicated with ergocalciferol. Clin Sci. 1985;68(2):135–141.
- Orbak Z, Doneray H, Keskin F, et al. Vitamin D intoxication and therapy with alendronate (case report and review of literature). Eur J Pediatr. 2006;165(8):583–584.
- Rajakumar K, Reis EC, Holick MF. Dosing error with over-the-counter vitamin D supplement: a risk for vitamin D toxicity in infants. Clin Pediatr. 2013;52(1):82–85.
- Rizzoli R, Stoermann C, Ammann P, et al. Hypercalcemia and hyperosteolysis in vitamin D intoxication: effects of clodronate therapy. Bone. 1994;15(2):193–198.
- Sezer RG, Guran T, Paketçi C, et al. Comparison of oral alendronate versus prednisolone in treatment of infants with vitamin D intoxication. Acta Paediatr. 2012;101(3):e122–e125.
- Talarico V, Barreca M, Galiano R, et al. Vitamin D and risk for vitamin a intoxication in an 18-month-old boy. Case Rep Pediatr. 2016;2016:1395718.
- Zhou L, Taylor-Miller T, Zacharin M, et al. Extreme hypercalcaemia due to accidental vitamin D intoxication. J Paediatr Child Health. 2019;55(1):104–106.
- Vieth R. Vitamin D supplementation, 25-hydroxyvitamin D concentrations, and safety. Am J Clin Nutr. 1999;69(5):842–856.
- Al-Kandari A, Sadeq H, Alfattal R, et al. Vitamin D intoxication and nephrocalcinosis in a young breastfed infant. Case Rep Endocrinol. 2021;2021:3286274.
- Muller MJ, Volmer DA. Mass spectrometric profiling of vitamin D metabolites beyond 25-hydroxyvitamin D. Clin Chem. 2015;61(8):1033–1048.
- Saponaro F, Saba A, Zucchi R. An update on vitamin D metabolism. Int J Mol Sci. 2020;21(18):6573–6589.
- Stokes CS, Lammert F, Volmer DA. Analytical methods for quantification of vitamin D and implications for research and clinical practice. Anticancer Res. 2018;38(2):1137–1144.
- Zhang Y, Bala V, Mao Z, et al. Quantification of fat-soluble vitamins and their metabolites in biological matrices: an updated review. Bioanalysis. 2020;12(9):625–640.
- Fraser WD, Tang JCY, Dutton JJ, et al. Vitamin D measurement, the debates continue, new analytes have emerged, developments have variable outcomes. Calcif Tissue Int. 2020;106(1):3–13.
- Turck D, Bresson JL, Burlingame B, et al. Dietary reference values for vitamin K. Efsa J. 2017;15(5):e04780.
- Dirks NF, Ackermans MT, Lips P, et al. The when, what & how of measuring vitamin D metabolism in clinical medicine. Nutrients. 2018;10(4):482.
- Shahangian S, Alspach TD, Astles JR, et al. Trends in laboratory test volumes for Medicare part B reimbursements, 2000–2010. Arch Pathol Lab Med. 2014;138(2):189–203.
- Ofenloch-Haehnle B. Approaches to measurement of vitamin D concentrations – immunoassays. Scand J Clin Lab Invest Suppl. 2012;243:50–53.
- European Medicine Agency, Guideline on bioanalytical method validation; 2011.
- van den Ouweland JMW, Vogeser M, Bächer S. Vitamin D and metabolites measurement by tandem mass spectrometry. Rev Endocr Metab Disord. 2013;14(2):159–184.
- Altieri B, Cavalier E, Bhattoa HP, et al. Vitamin D testing: advantages and limits of the current assays. Eur J Clin Nutr. 2020;74(2):231–247.
- Wagner D, Hanwell HE, Schnabl K, et al. The ratio of serum 24, 25-dihydroxyvitamin D3 to 25-hydroxyvitamin D3 is predictive of 25-hydroxyvitamin D3 response to vitamin D3 supplementation. J Steroid Biochem Mol Biol. 2011;126(3–5):72–77.
- Farrell CJ, Herrmann M. Determination of vitamin D and its metabolites. Best Pract Res Clin Endocrinol Metab. 2013;27(5):675–688.
- Yin S, Yang Y, Wu L, et al. Recent advances in sample preparation and analysis methods for vitamin D and its analogues in different matrices. Trends Analyt Chem. 2019;110:204–220.
- Rezayi M, Ghayour-Mobarhan M, Tavakoly Sany SB, et al. A comparison of analytical methods for measuring concentrations of 25-hydroxy vitamin D in biological samples. Anal Methods. 2018;10(47):5599–5612.
- Hollis BW, Kamerud JQ, Selvaag SR, et al. Determination of vitamin D status by radioimmunoassay with an 125I-labeled tracer. Clin Chem. 1993;39/(3):529–533.
- Haddad JG, Chyu KJ. Competitive protein-binding radioassay for 25-hydroxycholecalciferol. J Clin Endocrinol Metab. 1971;33(6):992–995.
- Arneson WL, Arneson DL. Current methods for routine clinical laboratory testing of vitamin D levels. Lab Med. 2013;44(1):e38–e42.
- Eisman JA, Shepard RM, DeLuca HF. Determination of 25-Hydroxyvitamin D2 and 25-Hydroxyvitamin D3 in human plasma using High-Pressure liquid chromatography. Anal Biochem. 1977;80(1):298–305.
- Jones G. Assay of vitamins D2 and D3, and 25-hydroxyvitamins D2 and D3 in human plasma by high-performance liquid chromatography. Clin Chem. 1978;24(2):287–298.
- Gilbertson TJ, Stryd RP. High-performance liquid chromatographic assay for 25-hydroxyvitamin D3 in serum. Clin Chem. 1977;23(9):1700–1704.
- Mata-Granados JM, Quesada Gómez JM, Luque de Castro MD. Fully automatic method for the determination of fat soluble vitamins and vitamin D metabolites in serum. Clin Chim Acta. 2009;403(1–2):126–130.
- Keyfi F, Nahid S, Mokhtariye A, et al. Evaluation of 25-OH vitamin D by high performance liquid chromatography: validation and comparison with electrochemiluminescence. J Anal Sci Technol. 2018;9(1):25–30.
- Abu el Maaty MA, Hanafi RS, Aboul-Enein HY, et al. Design-of-experiment approach for HPLC analysis of 25-hydroxyvitamin D: a comparative assay with ELISA. J Chromatogr Sci. 2015;53(1):66–72.
- Gathungu RM, Flarakos CC, Reddy GS, et al. The role of mass spectrometry in the analysis of vitamin D compounds. Mass Spectrom Rev. 2013;32(1):72–86.
- Couchman L, Moniz CF. Analytical considerations for the biochemical assessment of vitamin D status. Ther Adv Musculoskelet Dis. 2017;9(4):97–104.
- Abu Kassim NS, Gomes FP, Shaw PN, et al. Simultaneous quantitative analysis of nine vitamin D compounds in human blood using LC-MS/MS. Bioanalysis. 2016;8(5):397–411.
- Duan X, Weinstock-Guttman B, Wang H, et al. Ultrasensitive quantification of serum vitamin D metabolites using selective solid-phase extraction coupled to microflow liquid chromatography and isotope-dilution mass spectrometry. Anal Chem. 2010;82(6):2488–2497.
- Best CM, Riley DV, Laha TJ, et al. Vitamin D in human serum and adipose tissue after supplementation. Am J Clin Nutr. 2021;113(1):83–91.
- Lipkie TE, Janasch A, Cooper BR, et al. Quantification of vitamin D and 25-hydroxyvitamin D in soft tissues by liquid chromatography-tandem mass spectrometry. J Chromatogr B Analyt Technol Biomed Life Sci. 2013;932:6–11.
- Oberson JM, Benet S, Redeuil K, et al. Quantitative analysis of vitamin D and its main metabolites in human milk by supercritical fluid chromatography coupled to tandem mass spectrometry. Anal Bioanal Chem. 2020;412(2):365–375.
- Aronov PA, Hall LM, Dettmer K, et al. Metabolic profiling of major vitamin D metabolites using Diels-Alder derivatization and ultra-performance liquid chromatography-tandem mass spectrometry. Anal Bioanal Chem. 2008;391(5):1917–1930.
- Rola R, Kowalski K, Bieńkowski T, et al. Development of a method for multiple vitamin D metabolite measurements by liquid chromatography coupled with tandem mass spectrometry in dried blood spots. Analyst. 2018;144(1):299–309.
- Volmer DA, Mendes LR, Stokes CS. Analysis of vitamin D metabolic markers by mass spectrometry: current techniques, limitations of the "gold standard" method, and anticipated future directions. Mass Spectrom Rev. 2015;34(1):2–23.
- Adamec J, Jannasch A, Huang J, et al. Development and optimization of an LC-MS/MS-based method for simultaneous quantification of vitamin D2, vitamin D3, 25-hydroxyvitamin D2 and 25-hydroxyvitamin D3. J Sep Sci. 2011;34(1):11–20.
- Al-Zohily B, Al-Menhali A, Gariballa S, et al. Epimers of vitamin D: a review. Int J Mol Sci. 2020;21(2):470–485.
- Bailey D, Veljkovic K, Yazdanpanah M, et al. Analytical measurement and clinical relevance of vitamin D(3) C3-epimer. Clin Biochem. 2013;46(3):190–196.
- Abouzid M, Karaźniewicz-Łada M, Pawlak K, et al. Measurement of plasma 25-hydroxyvitamin D2, 25-hydroxyvitamin D3 and 3-epi-25-hydroxyvitamin D3 in population of patients with cardiovascular disease by UPLC-MS/MS method. J Chromatogr B Analyt Technol Biomed Life Sci. 2020;1159:122350.
- Albarhani AA, Collier F, Greaves RF, et al. Vitamins D and a can be successfully measured by LC-MS/MS in cord blood diluted plasma. Clin Biochem. 2015;48(16–17):1105–1112.
- Clarke MW, Tuckey RC, Gorman S, et al. Optimized 25-hydroxyvitamin D analysis using liquid–liquid extraction with 2D separation with LC/MS/MS detection, provides superior precision compared to conventional assays. Metabolomics. 2013;9(5):1031–1040.
- Geib T, Meier F, Schorr P, et al. A simple micro-extraction plate assay for automated LC-MS/MS analysis of human serum 25-hydroxyvitamin D levels. J Mass Spectrom. 2015;50(1):275–279.
- Bruce SJ, Rochat B, Beguin A, et al. Analysis and quantification of vitamin D metabolites in serum by ultra-performance liquid chromatography coupled to tandem mass spectrometry and high-resolution mass spectrometry-a method comparison and validation. Rapid Commun Mass Spectrom. 2013;27(1):200–206.
- Mena-Bravo A, Priego-Capote F, Luque de Castro MD. Study of blood collection and sample preparation for analysis of vitamin D and its metabolites by liquid chromatography-tandem mass spectrometry. Anal Chim Acta. 2015;879:69–76.
- Midttun O, Ueland PM. Determination of vitamins A, D and E in a small volume of human plasma by a high-throughput method based on liquid chromatography/tandem mass spectrometry. Rapid Commun Mass Spectrom. 2011;25(14):1942–1948.
- Tiwari A, Dawkhar B, Bagul M, et al. Development and validation of liquid chromatohraphy-electro-spray ionization-mass spectrometry method for the quantitation of ergocalciferol in human plasma. Int J Pharm Sci Res. 2018;9(9):3863–3869.
- Yang MY, Huang CY, Chiu THT, et al. Using gas chromatography and mass spectrometry to determine 25-hydroxyvitamin D levels for clinical assessment of vitamin D deficiency. J Food Drug Anal. 2019;27(2):494–501.
- Kobold U. Approaches to measurement of vitamin D concentrations – mass spectrometry. Scand J Clin Lab Invest Suppl. 2012;243:54–59.
- Hojskov CS, Heickendorff L, Moller HJ. High-throughput liquid-liquid extraction and LCMSMS assay for determination of circulating 25(OH) vitamin D3 and D2 in the routine clinical laboratory. Clin Chim Acta. 2010;411(1–2):114–116.
- Knox S, Harris J, Calton L, et al. A simple automated solid-phase extraction procedure for measurement of 25-hydroxyvitamin D3 and D2 by liquid chromatography-tandem mass spectrometry. Ann Clin Biochem. 2009;46(Pt 3):226–230.
- Jumaah F, Larsson S, Essen S, et al. A rapid method for the separation of vitamin D and its metabolites by ultra-high performance supercritical fluid chromatography-mass spectrometry. J Chromatogr A. 2016;1440:191–200.
- Liu TT, Cheong LZ, Man QQ, et al. Simultaneous profiling of vitamin D metabolites in serum by supercritical fluid chromatography-tandem mass spectrometry (SFC-MS/MS). J Chromatogr B Analyt Technol Biomed Life Sci. 2019;1120:16–23.
- Pilařová V, Plachká K, Khalikova MA, et al. Recent developments in supercritical fluid chromatography – mass spectrometry: is it a viable option for analysis of complex samples? Trends Analyt Chem. 2019;112:212–225.
- Musteata ML, Musteata FM. Overview of extraction methods for analysis of vitamin D and its metabolites in biological samples. Bioanalysis. 2011;3(17):1987–2002.
- Kasalova E, Aufartova J, Krcmova LK, et al. Recent trends in the analysis of vitamin D and its metabolites in milk–a review. Food Chem. 2015;171:177–190.
- Saleh L, Mueller D, von Eckardstein A. Analytical and clinical performance of the new Fujirebio 25-OH vitamin D assay, a comparison with liquid chromatography-tandem mass spectrometry (LC-MS/MS) and three other automated assays. Clin Chem Lab Med. 2016;54(4):617–625.
- Roth HJ, Schmidt-Gayk H, Weber H, et al. Accuracy and clinical implications of seven 25-hydroxyvitamin D methods compared with liquid chromatography-tandem mass spectrometry as a reference. Ann Clin Biochem. 2008;45(Pt 2):153–159.
- Spanaus K, von Eckardstein A. Evaluation of two fully automated immunoassay based tests for the measurement of 1α,25-dihydroxyvitamin D in human serum and comparison with LC-MS/MS. Clin Chem Lab Med. 2017;55(9):1305–1314.
- Shin SY, Kwon MJ, Song J, et al. Measurement of serum total vitamin D (25-OH) using automated immunoassay in comparison [corrected] with liquid chromatography tandem-mass spectrometry. J Clin Lab Anal. 2013;27(4):284–289.
- Jafri L, Khan AH, Siddiqui AA, et al. Comparison of high performance liquid chromatography, radio immunoassay and electrochemiluminescence immunoassay for quantification of serum 25 hydroxy vitamin D. Clin Biochem. 2011;44(10–11):864–868.
- Denimal D, Ducros V, Dupre T, et al. Agreement of seven 25-hydroxy vitamin D(3) immunoassays and three high performance liquid chromatography methods with liquid chromatography tandem mass spectrometry. Clin Chem Lab Med. 2014;52(4):511–520.
- Sempos CT, Betz JM, Camara JE, et al. General steps to standardize the laboratory measurement of serum total 25-Hydroxyvitamin D. J AOAC Int. 2017;100(5):1230–1233.