
Abstract
We describe a new and unusual vertebrate trackway from the Middle Jurassic Scalby Formation of the Cleveland Basin, Yorkshire, United Kingdom. The Enigmatic Burniston Trackway (EBT) is the first and only example of such a trackway known from this region. The best preserved EBT print, belonging to a pentadactyl tetrapod, does not resemble any known Middle Jurassic ichnogenus, but shares features with Triassic and Cretaceous archosaur and synapsid ichnotaxa. EBT most closely resembles the Triassic ichnogenus Synaptichnium in having the longest digit III, shortest digits I and V, and digit V positioned posterior to the other digits. Synaptichnium has been assigned to various trackmakers, including crocodylomorphs, and early archosaurs (‘thecodonts’ and aetosaurs). However, the anteriorly oriented digits and reduced and posterolaterally placed digit V of EBT also resemble Sederipes from the Cretaceous, and Dicynodontipus from the Permian-Triassic (both representing large-bodied synapsid or ‘mammal’ trackmakers). Unlike most traces assigned to cynodont (including mammalian) or crocodylomorph makers, EBT has low total digit divergence. Digit imprints end in short claws and the pes is rotated by ∼20° outwards in relation to the manus. Preservation is fragmentary for most of the trackway and the specimen could not be collected, but we record it here and provide comparative context within the existing ichnofossil record. We discuss the issues regarding ‘mammal’ ichnofossil literature, including taxonomic nomenclature, inconsistent diagnostic criteria, and assumptions made previously about Mesozoic synapsid body mass.
Introduction
Fossil trackways provide invaluable information about extinct faunal assemblages, and the palaeodistribution, foot morphology, locomotion and behaviour of extinct animals (Alexander, Citation1976; Falkingham, Citation2014; Sarjeant, Citation1995). Middle Jurassic trackways are relatively well known from the British Isles, for example there are extensive sites in Yorkshire (Romano & Whyte, Citation2003a), Oxfordshire (Day et al., Citation2004), and the Isle of Skye (e.g., dePolo et al., Citation2018, Citation2020; Romano et al., Citation2018). These mostly comprise dinosaur trackways and single prints, preserved in fluvial and lagoon environments. Although the traces of some other trackmakers are known (Romano & Whyte, Citation2003a, Citation2003b, Citation2010; Whyte et al., Citation2007), non-dinosaur vertebrate tracks are comparatively rare.
Herein we describe a unique trackway from the Middle Jurassic of the British Isles, identified during a field excursion of the Leeds Geological Association (led by MR) in 2015 to outcrops of the Ravenscar Group (Cleveland Basin), Burniston Bay, Yorkshire (). It comprises up to six separate prints of varied preservation (), with one well-preserved probable manus print belonging to a quadrupedal, pentadactyl, tetrapod trackmaker ().
Figure 1. Map of Burniston Bay showing the approximate location (asterisk) the specimen was found, and geological context (stratigraphy adapted from Rawson & Wright, Citation2018).
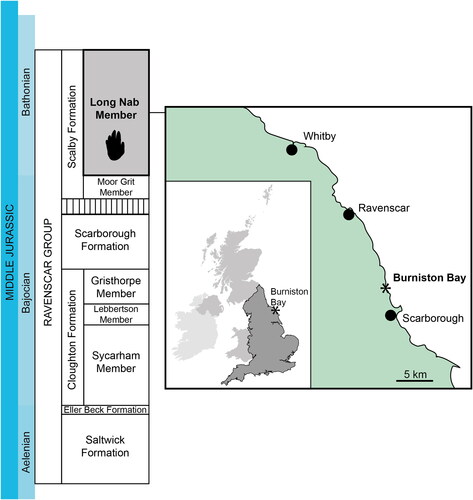
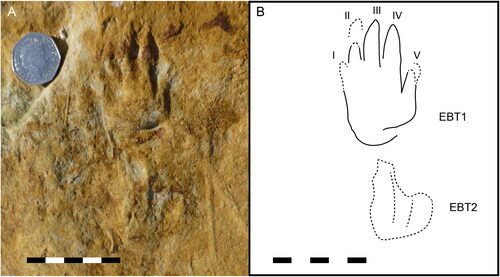
Figure 2. EBT (Enigmatic Burniston Trackway): A, preserved as a positive hypichnion; B, interpretive drawing. Scale 100 mm, same throughout.

Figure 3. Close up of best preserved print of EBT, a left manus EBT1, as shown in . A, print; B, interpretive drawing. Scale 50 mm, same throughout.
Of the vertebrate footprints known from the Ravenscar Group the majority are assigned to dinosaur trackmakers, with only a few having chelonian or crocodylomorph makers (Romano & Whyte, Citation2003a, Citation2010; Whyte et al., Citation2007) (). Some show behaviours, such as swimming, comprising digit drags of up to four digits in soft sediment (Romano & Whyte, Citation2010). Apart from the large sauropod pes prints, none preserve five digit imprints in either the pes or manus prints. Due to its size and number of digits alone, the track from Burniston likely represents a new ichnotaxon for this locality.
Figure 4. Ichnofossils assigned to synapsid and crocodylomorph trackmakers comparison: A, Synaptichnium (from Avanzini & Mietto, Citation2008); B, Synaptichnium (King et al., Citation2005); C, Dicynodontipus (from Retallack, Citation1996); D, Sederipes ‘Morphotype C’ (from Stanford et al., Citation2018); E, ‘Morphotype Civ; (from Romano & Whyte, Citation2010); F, Crocodylopodus (from Lockley & Meyer, Citation2004); G, Pooleyichnus (from Sarjeant, Citation1975); H, Schadipes (from Lockley & Foster, Citation2003); I, Koreasaltipes (from Kim et al., Citation2017); J, Catocapes (from Mateus et al., Citation2017); K, Batrachopus, (from Avanzini et al., Citation2007); L, Ameghinichnus (from deValais, 2009). Scale equals 50 mm in C, same scale as in C for: C-D, F-H, J-L. Scale equals 100 mm in E. Scale equals 10 mm in I.

The previous recordings of crocodylomorph prints from the Cleveland Basin of Yorkshire (Romano & Whyte, Citation2003a, Citation2010) filled an important gap in the crocodylomorph ichnological record for this time globally. The only trackway known from the Middle Jurassic of the British Isles suggested to have been made by a ‘mammal’, is a purported mammaliaform print briefly described by Sarjeant (Citation1975), Pooleyichnus, based on a specimen collected in 1886 from the Taynton Limestone Formation (Stonesfield Slate).
The new Burniston trackway is described herein, and the possible trackmaker affinities assessed. We address problems with previous diagnoses for ‘mammal’ ichnogenera, and discuss issues concerning body mass and the evolution of pedal morphology in synapsids, and their bearing on the Burniston track and the synapsid ichnofossil record globally.
Materials and methods
The new vertebrate trackway is here named Enigmatic Burniston Trackway, or EBT. The moniker refers to the difficultly in interpreting the track, coupled with the locality in which it was found. Individual prints in the trackway are referred to with a number suffix- EBT1, EBT2 etc. ( and ). EBT was discovered during a Leeds Geological Association field excursion in 2015. It was on the surface of a fallen block in the middle of Burniston Bay, approximately 5 km north of Scarborough (). The block came from the Long Nab Member of the Scalby Formation, which is Bathonian (Middle Jurassic) in age (Barron et al., Citation2012; Rawson & Wright, Citation2018). Unfortunately it was not possible to collect the specimen, and the block on which the trackway was preserved has since been destroyed by coastal erosion.
Images of EBT were measured from photographs using measuring tools in the visualization software ImageJ (Rasband, 1997–Citation2018). Data for other comparative tracks were compiled from the literature (cites herein). Tracks were included where manus vs pes identification was unknown, but where identification was known the manus measurements were favoured to facilitate comparison with the best-preserved Burniston track, which represents a manus (see below). Measurement methodology, and ichnological terminology, follow Leonardi (Citation1987).
We use crown-group cladistical terminology throughout this text, in recognition of its commonly accepted use in the current palaeontological literature. Archosauria refers to the group that includes the common ancestor of living birds and crocodiles, and their extinct relatives. This includes Pseudosuchia (Aetosauria, Rauisuchia, Crocodylomorpha and relatives) and Ornithosuchia (Ornithosuchidae and Ornithodira and relatives). We use Crocodylomorpha (which includes Crocodyliformes, Mesoeucrocodylia, Neosuchia, Eusuchia and Crocodylia) rather than ‘crocodilian’ (see Lockley et al., Citation2010 for discussion of this nomenclature in relation to ichnofossils). For simplicity, herein we use the term archosaur to refer to any member of this clade, unless referring to a specific group.
The term ‘mammal’ has previously been used to refer to prints assigned to trackmakers ranging from non-mammalian synapsids to crown mammalians (Lockley & Foster, Citation2003). This lack of clarity partly owes to a legacy of previously used terminology (for example the dubious term ‘mammaloid’, or the now defunct term ‘mammal-like reptiles’), and changes in the commonly used nomenclature for the mammal lineage. Mammals were previously defined as animals including Sinoconodon and all descendants, but most Mesozoic mammal palaeontologists have now adopted the crown-based definition (Rowe, Citation1988), which recognises Sinoconodon and all descendants as Mammaliaformes, with the term Mammalia restricted to the common ancestor of Monotremata and Eutheria and all descendants (). The closest non-mammaliaform relatives, such as Tritylodonta, belong to Mammaliamorpha, which is nested in Cynodontia, within Synapsida.
Figure 5. Simplified relationships and terminology for Synapsida (adapted from Angielczyk, Citation2009).
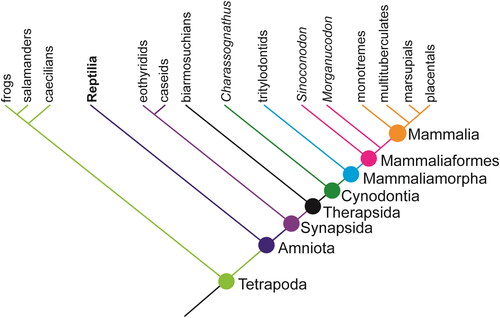
The lack of specificity in some of the previous ichnological literature creates confusion over what authors are referring to when they assign ‘mammal’ trackways. This presents problems for interpretation, particularly regarding variation among Mesozoic cynodonts (see discussion). In recognition of this uncertainty, and to avoid further taxonomic confusion, we use terminology carefully, and specifically refer to groups wherever possible when discussing prints made by trackmakers from the synapsid lineage. For further information on terminology among Synapsida, see Angielczyk (Citation2009).
Geological context
The ichnofossils of the Cleveland Basin, Yorkshire, are globally recognized as comprising a significant megatracksite (Whyte et al., Citation2007). The tracks occur in the Ravenscar Group, which represents a period of predominantly non-marine deposition spanning approximately 10 million years of the Aalenian to the Bathonian (Barron et al., Citation2012; Romano & Whyte, Citation2003a). The Ravenscar Group comprise a series of thin-bedded sequences of silts, clays and sands representing freshwater to brackish floodplain and lacustrine environments, with occasional marine intercalations (Barron et al., Citation2012; Whyte et al., Citation2007). Freshwater bivalves (Unio), the burrow Lockeia, and the presence of Equisetum (Whyte et al., Citation2007) characterise the non-marine units.
Although the stratigraphical context of the slab containing the footprints cannot be known with total certainty because it is now lost, it can be inferred from its location, appearance, and previous ichnofossil discoveries. Based on these factors, it is certain that the slab originated from the Long Nab Member of the Scalby Formation (). The Long Nab Member is underlain by the Moor Grit Member, and overlain disconformably by the Callovian aged Abbotsbury Cornbrash Formation (Rawson & Wright, Citation2018; Romano & Whyte, Citation2003a). Part of the Long Nab Member includes the sandstone ‘Burniston Footprint Bed’ (Hargreaves, Citation1914; Romano & Whyte, Citation2003a), a unit long known to be rich in dinosaur footprints (Hargreaves, Citation1914; Ivens & Watson, Citation1994; Romano & Whyte, Citation2003a).
ICHNOTAXONOMY
The new trackway from Burniston Bay described here was not collected or recorded using photogrammetry, and the figures herein provide the only source for comparison. We therefore choose not to assign a formal name. However, as the prints are considered to be a new ichnotaxon for this locality, and possibly even a new ichnogenus, they are assigned an alphanumerical tag in the hope that they may be referred to unambiguously in the future, and so avoid being lost in the wealth of vertebrate ichnotaxonomic literature.
Material: Four to six tracks preserved as positive hypichnia on the underside of a fallen thick-bedded sandstone block. The bedding plane on which the tracks are preserved is covered with a number of vertebrate and invertebrate traces. The tracks could not be collected at the time they were found, and have since been destroyed by coastal erosion.
Horizon and locality: The loose block on which the tracks occur comes from the Long Nab Member, Scalby Formation, Ravenscar Group, Middle Jurassic (Bathonian). It was located in the middle of Burniston Bay approximately 4 km north of Scarborough (TA0285 9375).
Description: A well-preserved footprint, EBT1, is located at the ‘anterior’ end of an indistinct trackway consisting of four to six prints. Only this single print from EBT permits detailed description. It is approximately 64 mm long and 38 mm wide, with digit proportions III > II > IV > V > I (although the difference in length between digits II and IV is very slight) (). EBT1 has a nearly straight posterior margin, sub-parallel digits, and a slightly more divergent digit V, set back posterolaterally to the rest of the digits. The print has low total digit divergence of only ∼40°, but poor preservation of digit I makes this angle uncertain. Digit imprints terminate with acuminate claw imprints. Pes prints are immediately behind manus prints, and appear rotated slightly laterally (outwards). They are indistinct, shallow impressions showing two to four scratch-like digit traces.
Table 1. Measurements of the well-preserved EBT1 manus print, alongside selected Permian and Mesozoic synapsid ichnogenera.
We interpret EBT1 as a left manus print. Our identification of this as the left-hand side of a trackway is further supported by the lack of prints in the centre of the slab. An alternative interpretation of EBT is that the paired tracks represent both halves of a narrow trackway, and that EBT1 could represent an over-stepped right pes impression. If so, numbering of the digits of this print would be reversed (the shortest digit would be I, not V). However, in the field it was observed that faint tracks were apparent along the broken longest edge of the slab, which may have represented the right side of the tracks (MR, pers. obs.), although these are not visible in the photographic record.
In EBT1 there is no clear evidence of pads, and the digit imprints are similar in width along their length, narrowing rapidly anteriorly and ending in a sharp point. The print immediately posterior to EBT1, EBT2, is faint, but there are traces of at least three digit imprints aligned at a ∼∼20° angle outwards to those of the print in front, indicating the foot was turned slightly outwards relative to the midline of the trackway. Other elements in the trackway are not well-preserved enough to permit detailed comparison, but are interpreted as belonging to the same trackway. Another pair of traces, EBT3 and EBT4, are positioned 126 mm posterior to the two prints described above. A putative track, EBT5, lies 102 mm behind this. Damage to the sandstone surface makes identification of this as a track less certain. Another 113 mm posterior to that, at the edge of the sandstone slab, lies a final potential print (EBT6), also poorly preserved. If their identification as tracks is correct, this provides a complete trackway length of ∼558 mm, and a stride length of approximately 230 mm.
Remarks
The morphology of EBT is different from any other vertebrate track from the Middle Jurassic Ravenscar Group of the United Kingdom. The two ichnotaxa most similar to EBT are Synaptichnium and Sederipes, neither of which are Jurassic (they are Triassic and Cretaceous respectively).
EBT1 resembles the manus of Synaptichnium pseudosuchoides in being pentadactyl, mesaxonic, the digits end in claws, digit III is longest, digit I and V are the shortest, and digit V is straight, set back from the other digits and situated proximally at an angle of c. 40° to the long axis through digit III, and the impressions of digits I and V are less distinct in the print than the other digit impressions (King et al., Citation2005). The posterior margin is rarely preserved in Synaptichnium, but where it is (e.g., King et al., [Citation2005:fig 8b]), the manus appears to narrow slightly posteriorly, unlike EBT1, which has a straight posterior margin of similar width to the rest of the manus ().
Sederipes print m23, ‘Morphotype C’ (Stanford et al., Citation2018) has subequal digits II-V and a shorter digit I, a distinctively posteriorly set lateral digit (identified as digit I in Stanford et al., [Citation2007, Citation2018]). The posterior position of digit V in EBT1 is similar to that seen in Sederipes, but this becomes less significant if the digit notation is reversed in Sederipes as we suspect (see discussion).
The low total digit divergence of EBT1 (∼40°), is unlike that seen in crocodylomorph traces Batrachopus, Crocodylopodus, or Angolaichnus (Kim et al., Citation2020; Lockley & Meyer, Citation2004; Mateus et al., Citation2017; Olsen & Padian, Citation1986), or mammaliamorph synapsid tracks, such as Schadipes, Ameghinichnus and Catocapes (de Valais, Citation2009; Lockley & Foster, Citation2003; Mateus et al., Citation2017), which show wider digit divergence. The digit notation is different from that seen in Batrachopus, in which digit II points anteriorly and V points posteriorly (Olsen & Padian, Citation1986). EBT resembles some features of Dicynodontipus, being plantigrade and pentadactyl, with digits that are oriented anteriorly and digit V slightly more divergent and posteriorly shifted (Melchor & de Valais, Citation2006; Marchetti et al., Citation2019). However, Dicynodontipus can also have more divergent manus digits (e.g., Retallack, Citation1996). For further discussion of these ichnotaxa, see below.
Discussion of potential trackmakers
Trackmakers known from the Ravenscar Group include dinosaurs, chelonians, crocodylomorphs, and pterosaurs (Romano et al., Citation2018; Whyte et al., Citation2007; Whyte & Romano, Citation2014). EBT1 clearly does not conform to the autopodial morphology of any of these groups except possibly crocodylomorphs (see below). It is obviously not from a dinosaur. Chelonian trackways typically consisting of short, subparallel digit imprints with deep scratch marks that widen posteriorly (Avanzini et al., Citation2005; Lockley & Foster, Citation2006; Romano & Whyte, Citation2010), not congruent with the morphology of EBT.
Other terrestrial groups represented by body fossil material from the Middle Jurassic of the British Isles include squamates, lepidosauromorphs, pterosaurs, and mammaliamorphs (this includes mammaliaforms and mammalians, see discussion below) (Evans et al., Citation2006; Evans & Milner, Citation1994; Panciroli et al., Citation2017a, Citation2017b, Citation2018a, Citation2018b). All taxa known from these groups are too small to have produced EBT - with the possible exception of pterosaurs, but pterosaur tracks are distinctly different, with a four-toed manus and pes with an elongate morphology of the pes (Lockley et al., Citation1995; Padian & Olsen, Citation1984).
An alternative interpretation of EBT is that it was made by a lacertilian trackmaker (see below re. Pooleyichnus). An indentation anterior to our assigned digit II could be suggested to be the true tip of this digit, which would require re-interpretation of this as an elongate digit IV on a right manus. Lockley et al. (Citation2004 :91) noted that the smaller manus of the ichnogenus Rhynchosauroides could be confused with the pes print of a synapsid. However, interpretation of EBT as lacertilian is unlikely for the following reasons: lacertilian digits usually curve medially, and those of EBT do not. There is also no tail drag, which is often associated with lacertilian trackways (but not a diagnostic feature of them), although tail drags are also seen in cynodont tracks, e.g., Ameghinichnus (de Valais, Citation2009). A line that could be interpreted as a tail drag near the well-preserved print EBT1 is too close to this print to be left by a tail, and as noted previously, there are no tracks on the rest of the block (which would represent the left-hand side of the trackway), leading us to exclude a right-hand print. However, we note that poor preservation means a tail drag may not be preserved in EBT. Another possible argument for a lacertilian affinity for EBT is the similar length of digits II, III and IV ( and ) (Leonardi, Citation1987). However, many synapsid tracks show similar proportional digit lengths (see discussion below). Finally, the claws at the tip of each digit could indicate a lacertilian trackmaker, and could be considered an identifying criteria, but claw marks are also present in some cynodont tracks from the Triassic (e.g., Lockley et al., Citation2004), Jurassic (e.g., Lockley et al., Citation2004) and from the Cretaceous ( Schadipes, e.g., Lockley & Foster, Citation2003; Contessi, Citation2013 ), so this is also not a diagnostic criterion.
There are however, multiple similarities between EBT1 and manus prints assigned to the Triassic ichnogenus Synaptichnium (Nopcsa, Citation1923; King et al., Citation2005), belonging to the ichnofamily Chirotheriidae (). This ichnogenus has been attributed to a wide range of trackmakers, including lacertilians, crocodylomorphs, pseudosuchians, aetosaurs, and most recently, ‘thecodont’ (early archosaur) trackmakers (King et al., Citation2005 and references therein). Only one ichnospecies, S. pseudosuchoides, is known from the British Isles, but only from a fragmented holotype and lectotype. Although the similarity between EBT1 and S. pseudosuchoides is striking, there is a large temporal gap between this Early-Late Triassic ichnogenus and the Middle Jurassic EBT1. It does suggest, however, that a related trackmaker may have been responsible. The only archosaur groups surviving into the Middle Jurassic are crocodylomorphs and ornithodirans such as dinosaurs and pterosaurs. As the latter two groups have already been discounted on morphological grounds, if EBT is assignable to Synaptichnium, this supports this print interpretation as a possible crocodylomorph trackmaker. However, as EBT is lost, we consider there to be insufficient support for assigning it to Synaptichnium at this time.
Potential Crocodylomorph Trackmaker
Crocodylomorph trackways are attributed to the ichnofamily Batrachopodidae ( Lull, Citation1904; revised by Lockley & Meyer, Citation2004 ) and have a range extending throughout the Mesozoic. The diagnosis for Batrachopodidae includes: 1) trackway of a quadruped with pronounced heteropody; 2) manus pentadactyl, digitigrade and much smaller than the pes (heteropody); 3) pes tetradactyl, plantigrade with four digits, sometimes terminating in acuminate claw impressions; 4) pes digits II and IV subequal in length and slightly shorter than digit III; 5) pes digit I shortest; 6) trackway narrow, about twice pes width; 7) inner margin of pes tracks falls on or near trackway axis; 8) outward pes rotation 24–33˚ based on alignment of digit III (foot axis) in relation to trackway axis; 9) placement of manus close to trackway axis and typically immediately in front of the pes; 10) step about 2–3 times pes length; and 11) pes pace angulation about 150–160˚. Crocodylomorph tracks also have a phalangeal pad formula of 2-3-4-4-2 or 3(0) (although Protosuchidae have 2-3-4-5-3(4)) (Leonardi, Citation1987). Discussions of crocodylomorph trackways often use modern crocodylians as extant homologs for understanding Mesozoic tracks. While this provides some useful information, it fails to account for the distinctively different morphologies of early crocodylomorphs, particularly Triassic forms (Lockley et al., Citation2010). In extant studies, the modern crocodylian manus imprints have a digit divergence between 60°-179° (Farlow et al., Citation2018; Kubo, Citation2008; Milan & Hildegaard, 2010).
By the above diagnosis, EBT only partially meets diagnostic criteria 1, 8 and 9, although not all criteria can be assessed due to the poor preservation of all but the manus print. Regarding the extant crocodylian trackways, EBT is less divergent in the interdigital angle (∼40°) than the range outlined by other authors. Many of the criteria listed above are also criteria included in the diagnoses of other trackways, and so are not exclusive to crocodylomorph tracks.
There are few Middle Jurassic crocodylomorph tracks globally, but some have been reported from the Ravenscar Group (Romano & Whyte, Citation2010). EBT does not match the other crocodylomorph tracks from the Ravenscar Group, which are more rounded in morphology with a wide mediolateral width at the base of the digits. The best preserved print, morphotype Civ, a probable manus (Romano & Whyte, Citation2010, ), is larger than the EBT trace from Burniston (78 mm length by 66 mm width) and has shorter digits, <35% of the print length versus ∼50% in EBT ( and 6).
The ichnofossil literature for crocodylomorph trackways is somewhat confused, but ichnotaxa include Sustenodactylus (Lull, Citation1904) from the Early Jurassic of North America, which is possibly a junior synonym (Olsen & Padian, Citation1986) of Batrachopus () (Hitchcock, Citation1845) from the Early Jurassic of North America and France; Crocodylopodus () from the Jurassic-Cretaceous boundary of Spain, and Angolaichnus from the Cretaceous of Angola (Mateus et al., Citation2017). EBT does not match the morphology of any of these crocodylomorph ichnotaxa. In the revised diagnosis for Batrachopus, Olsen and Padian (Citation1986) included the rotation of the manus such that digit II pointed forward and digit V pointed backwards, with a wide total divergence of the digits. These features are not shared with EBT. The diagnosis for Crocodylopodus includes: slender, well-separated, clawed digits; wide divergence of digits I and V; manus digit II oriented almost parallel to the trackway axis with digits III and IV oriented laterally; narrow trackway about twice pes width (Lockley & Meyer, Citation2004). Although the manus digit imprints of EBT1 are also clawed, it meets none of the other diagnostic criteria for the ichnogenus Crocodylopodus. The Cretaceous crocodylomorph ichnogenus Angolaichnus also does not match the morphology of EBT: although the manus is plantigrade in this ichnotaxon, it is also rotated laterally relative to the trackway axis, and the digits are more divergent than in EBT (Mateus et al., Citation2017).
Setting aside the heteropody of crocodylomorph tracks, which cannot be assessed with certainty in the EBT track from Burniston due to the poor preservation of the pes, it is clear that this track does not conform to any of these crocodylomorph ichnotaxa, nor closely enough to the diagnosis of Batrachopodidae to assign it to this ichnofamily with confidence.
Potential Synapsid Trackmaker
For an outline of current synapsid terminology and understanding of their relationships, see Materials and Methods and . There is confusion in the literature over assigning tracks to ‘mammal’ trackmakers, a term that has been used to include everything from Permian-Triassic synapsids through to Late Cretaceous crown mammalians (Lockley & Foster, Citation2003). This is partly due to the difficulty of recognizing distinct trackmakers from their tracks, a common problem recognised by ichnologists due to the nature of footprint evidence (Baird, Citation1957; Leonardi, Citation1987). The taxonomic confusion when discussing ‘mammals’ is further complicated by the temporal overlap of multiple synapsid lineages, particularly during the Late Triassic to Early Cretaceous, when synapsids and non-mammalian cynodonts persisted alongside emerging and proliferating lineages of mammaliamorphs, mammaliaforms, and mammalians (see Kemp, Citation2005 for an overview of this diversity). This makes assigning tracks to a true mammalian trackmaker, rather than a non-mammalian synapsid or other early mammal relative, problematic.
Some key criteria for identification of Mesozoic ‘mammal’ trackways, as named in the literature, include: 1) tracks are pentadactyl and mesaxonic (the axis of the foot is formed by digit III); 2) digits II-IV are divergent, with digits I and V shorter and more divergent; 3) phalangeal formula 2-3-3-3-3; and 4) trackways are relatively small, including small digit lengths (Contessi, Citation2013; Lockley & Foster, Citation2003). The EBT tracks meet the first criteria of being pentadactyl and mesaxonic, and partially meet the second in having a shorter digit I and IV. However, the other two criteria are subjective and mainly applicable to mammaliaforms. Triassic non-mammalian cynodonts did not all have exceptionally divergent digits (Kümmell & Frey, Citation2012). Cynodonts have a phalangeal formula of 2-3-4-4-3, and the subjective criterion of size is based on (incorrect) assumptions about cynodont body mass in the Mesozoic (see below).
Criteria for making the distinction between crown-group mammalian tracks versus non-mammalian cynodonts were proposed by de Valais (Citation2009): 1) an overall similarity to conservative mammal tracks; 2) pentadactyl manus and pes; 3) digit impressions subequal in size and shape; 4) mesaxonic tracks; 5) no sprawling gait; 6) advanced gait indicating important locomotion development (e.g., running, galloping or hopping gaits), related with 5); and 7) relatively small-sized tracks. These provide a useful guide, but may not take into account the range of posture and movement possible in earlier mammaliaforms and non-mammalian cynodonts, and difficulties interpreting the morphology of trackmakers from the ichnofossil record.
Multiple authors state that most Mesozoic ‘mammals’ were small, rarely reaching over 30 g in body mass (e.g., Lockley et al., Citation2004:97), and their postcranial fossil record is poor. However, discoveries in the last two decades have transformed our understanding of the morphology and ecology of cynodonts in the Mesozoic, and vastly improved the postcranial fossil record, especially for mammaliaforms. Although many cynodonts underwent a reduction in body size in the Late Triassic, including the lineage that led to mammaliaforms, not all members of the synapsid lineage were diminutive, and those that were, did not all remain so. For example, Lisowicia bojani was a therapsid from the Late Triassic of Poland that stood at 4.5 m in height and had a body mass of 9 tons (Sulej & Niedźwiedzki, Citation2019). The tritylodontids, an incredibly successful group of mammaliamorphs considered to be the sister-group to mammaliaforms, lived alongside mammals into the Early Cretaceous (Matsuoka et al., Citation2016; Panciroli et al., Citation2017a), and some reached as much as 42 kg (as estimated for Kayentatherium, Hoffman & Rowe, Citation2018). Among mammaliaforms, examples of larger body mass include the Middle Jurassic docodontan Castorocauda, estimated at around 800 g (Ji et al., Citation2006), and the Early Cretaceous gobiconodontid crown mammalian, Repenomamus giganticus, at 14 kg (Hu et al., Citation2005), both from China.
Another seldom addressed but key issue in the interpretation of Mesozoic synapsid trackways is the evolution of foot morphology. Many previous studies assume a ‘rodent-like’ track for all mammals in the Mesozoic (Lockley et al., Citation2004). However, the posture and stance of mammaliaform groups, such as the morganucodontans, docodonts and haramiyidans remained morphologically somewhat conserved from their earlier cynodont relatives (Jenkins, Citation1971; Kühne, Citation1956; Luo & Wible, Citation2005; Zhou et al., Citation2013). These groups also retain a more rigid pectoral girdle and comparatively more sprawling stance than later crown mammals (Kielan-Jaworowska & Hurum, Citation2006) – with the exception of monotremes – which may be reflected in their tracks. For this reason, direct comparison with modern crown mammals has limitations. Broad similarities in foot morphology between many branches of Synapsida (Kümmell & Frey, Citation2012) suggests basal mammaliaform and non-mammalian cynodont tracks may usefully be compared to geologically older synapsid trackways, such as the Permian Chelichnus (Jardine, Citation1850) or Permian-Triassic Dicynodontipus (Rühle von Lilienstern, Citation1944) (see below). Indeed the elevation of the middle part of the digit arcade – a feature typical in many modern mammals - can be seen in ichnofossil tracks made by stem-mammals as far back as the Permian (Kümmell & Frey, Citation2012) - although it is not present in EBT.
The first track to be assigned to a ‘mammal’ trackmaker in the Jurassic of the British Isles was Pooleyichnus burfordensis (Sarjeant, Citation1975) (), based on a single natural cast from the Stonesfield Slate of the Taynton Limestone Formation, Oxfordshire. Although collected in 1886 it remained undescribed in the Natural History Museum of London until the 1970s, when it was attributed to a ‘mammalian’ trackmaker. This was based primarily on: the length of digit III, as the longest digit; and the wide divergence of the digits (interdigital divergence of 12-38°, total 87°); and only having three digit segments in digits III and IV. It was identified as a right manus or pes in the text (Sarjeant, Citation1975:275), but is measured as a left and depicted as left in the figures (Sarjeant, Citation1975:274). EBT does not conform to the morphology of Pooleyichnus. In Pooleyichnus the digit imprints are more divergent and elongate, and digit I and V are longer in relation to the rest of the digits than in the ‘Burniston Track’ (). The digits of Pooleyichnus also comprise up to 63% of the whole print, with no elongate palm/heel as seen in EBT.
An alternative interpretation of Pooleyichnus is that it belongs to a lacertilian trackmaker. While in the original description digit III was described as the longest digit, impressions anterior to digit II could be interpreted as the tip of an elongated digit (EP pers. obs.), suggesting a reinterpretation of this as digit IV and making this the asymmetrical right pes print typical of a small reptilian trackmaker. It could be re-evaluated as belonging to an ichnotaxon akin to Rhynchosauroides; an ichnogenus that has been attributed to lepidosauromorph (Haubold, Citation1971a; Lockley & Hunt, Citation1995), prolacertiform (Avanzini & Renesto, Citation2002; Diedrich, Citation2002) or sphenodontid (Baird, Citation1957) reptiles. Based on the body fossils from other Middle Jurassic sites in the British Isles, we know that lepidosauromorphs were present at this time, together with stem squamates (Evans et al., Citation2006; Evans & Milner, Citation1994; Panciroli et al., Citation2018b, Citation2020).
A rich assemblage of Middle Jurassic mammaliamorphs, mammaliaforms and mammalians are known from sites in England and Scotland (Evans et al., Citation2006; Evans & Milner, Citation1994; Panciroli et al., Citation2018b, Citation2020). None of these are of sufficient body mass to be identified as the EBT trackmaker. Assessing the size of a wider range of synapsid prints from the Mesozoic ichnofossil record, EBT1 is among the larger prints, but fits comfortably within the range of, known ichnogenera (). EBT1 is smaller than Dicynodontipus (Rühle von Lilienstern, Citation1944) and Sederipes (Stanford et al., Citation2018).
Figure 6. Measurements for ichnogenera attributed to selected Permian and Mesozoic trackmakers. Measurements in .
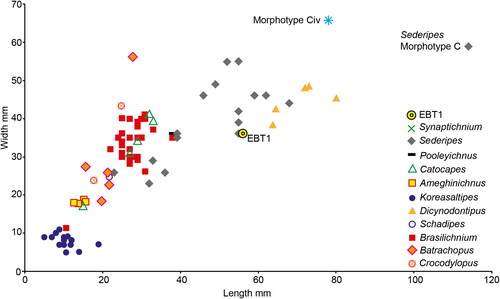
Dicynodontipus is Permian to Triassic in age, and attributed to a cynodont trackmaker (Haubold, Citation1971b; Marchetti et al., Citation2019) (not the dicynodont Lystrosaurus as suggested by Retallack, Citation1996). Like EBT, Dicynodontipus is plantigrade (although the manus is diagnosed as semi-plantigrade by Marchetti et al., Citation2019) and pentadactyl, and the manus is placed slightly anterior to pes (although it can be overstepped during faster gaits) (). The digits are oriented anteriorly, and digit V is slightly more divergent and posteriorly shifted (Melchor & de Valais, Citation2006) – although less so than in EBT1. Some tracks assigned to Dicynodontipus have more divergent manus digits (e.g., Retallack, Citation1996) which do not match the morphology of EBT1, but overall the similarities outlined above between this ichnogenus and EBT1 could suggest an affinity. We do not suggest assigning EBT to this ichnogenus due to the relatively longer digits of EBT1, as well as the temporal difference between their occurrences.
The tracks of Sederipes were found in the Early Cretaceous Potomac Group of Maryland, USA, and are assigned to a crown-mammalian trackmaker by Stanford et al. (Citation2018). Their large size would make the trackmaker the largest crown-mammal known from either the ichno- or body fossil record (but not the largest cynodont) (). Like EBT1, Sederipes is pentadactyl, but due to problems differentiating between manus and pes prints, these were not diagnosed separately, limiting comparisons with EBT. Sederipes tracks have subequal digits II-V and a shorter digit I, and a convex margin to the heel, with tracks often arranged in pairs that may indicate a “sitting position” (Stanford et al., Citation2018:8). Although the latter behaviour is not seen in EBT from Burniston (and is of limited utility as a diagnostic feature), Sederipes shares features with EBT. The most striking resemblance is found between print m23, ‘Morphotype C’ from Stanford et al. (Citation2018) (), previously figured in Stanford et al. (Citation2007), and EBT1. Print m23 is significantly larger than any of the other synapsid trackways known, including the others assigned to Sederipes, and EBT ( and ). It resembles EBT1 in having a distinctively posteriorly set lateral digit (identified as digit I in Stanford et al., [Citation2007, Citation2018]), and subequal central digits (identified as digits II-V in Stanford et al., [Citation2007, Citation2018]). Track m23 is an isolated print, making it difficult to determine whether it is left or right, manus or pes. If the short, posteriorly positioned digit is re-interpreted as a digit V, it matches much of the morphology of EBT1. Digit pads are present in m23, and are a common feature in many synapsid ichnofossils (Kümmell & Frey, Citation2012). Although EBT1 does not have digit pads, this could be a preservational difference rather than a morphological one, and pads are not present in all synapsid ichnofossils, including mammaliforms (e.g., some Schadipes and Ameghinichnus, [Kümmell & Frey, Citation2012]).
Most other Mesozoic synapsid tracks are smaller than EBT, and are likely to represent cynodonts (including tritylodontids, mammaliaforms and mammalians) (). These include the Late Triassic to Middle Jurassic Ameghinichnus (Casamiquela, Citation1961), Late Triassic to Early Cretaceous Brasilichnium (Leonardi, Citation1981), and Cretaceous Schadipes, Catocapes (Mateus et al., Citation2017), and Koreasaltipes (Kim et al., Citation2017). Multiple prints were also named by Ellenberger (Citation1974) from the Late Triassic to Early Jurassic of South Africa and Lesotho, however, the profusion of ichnotaxa without distinct and exclusive diagnoses makes these tracks problematic.
We do not find sufficient similarities between EBT and any other Mesozoic cynodont fossil prints to assign our track to these ichnogenera. Other cynodont tracks have wider digit divergence (); in Schadipes digits I and V are more divergent and short, with digits II-IV subequal in length, and in trackway UCM220.28 (Lockley & Foster, Citation2003: ) the digits are more anteriorly oriented, but the total digit divergence for most prints in this ichnogenus is still >100° (Lockley & Foster, Citation2003) (). The pes print is positioned lateral to the manus in Ameghinichnus (de Valais, Citation2009) as in EBT, but manus and pes prints are wider than they are long in Ameghinichnus, and also exhibit a greater total digit divergence, >100° (). Digit V is short in Catocapes, a recently named ichnogenus from Angola (Mateus et al., Citation2017) (), but the narrowest total digit divergence of this ichnogenus is still higher than in EBT1, at 80°, and the track is wider than it is long, with the digits representing >60% of the track length.
Koreasaltipes and Brasilichnium both include saltating (hopping) gaits, which are unlikely in larger-bodied Mesozoic taxa such as the animal responsible for EBT. Koreasaltipes is very small and diagnosed as bipedal, with pes width wider than length (Kim et al., Citation2017) (). Brasilichnium has a larger size range and is quadrupedal with a variable gait, but again tracks are mostly much smaller than EBT, wider than they are long, lack proportionally long digits, and like Koreasaltipes, tend to be preserved in eolian environments (Kim et al., Citation2017; Porchetti et al., Citation2017), whereas EBT at Burniston was found in fluvial sandstone.
Conclusions
These new tracks are unique in the rocks of the Cleveland Basin of Yorkshire, and reinforce the significance of the British Isles for the study of Middle Jurassic fauna and ecosystems. Although the classification of the Enigmatic Burniston Track, EBT, remains uncertain, the prints do not closely resemble other trackmakers known from the Middle Jurassic of the British Isles. We rule out dinosaur, pterosaur, chelonian and lacertilian trackmakers, and consider the track congruent with either an archosaur - namely crocodylomorph - trackmaker or a synapsid (cynodont) trackmaker. If they were made by a crocodylomorph, they represent a previously unknown ichnotaxon. If they were made by a synapsid (incl. mammalian or non-mammalian cynodonts), and if Pooleyichnus does indeed represent a lacertilian trackmaker, this would make the Burniston track the first and only ‘mammal’ track recorded in the British Isles, representing a rare record of large-bodied cynodont.
The most similar ichnotaxa are Synaptichnium from the Triassic of Europe and North America, and m23 Morphotype C (?Sederipes) from the Early Cretaceous Potomac Group of Maryland, USA. But the temporal gap between these ichnotaxa and EBT, and the loss of the original trackway described here from Burniston, suggest caution in attributing the Middle Jurassic track to either ichnotaxon. Globally the increasingly good record for Mesozoic cynodonts indicates many basal groups persisted longer, and that mammaliaforms in particular achieved greater body masses than previously recognised. Non-mammalian cynodonts, mammaliaforms and mammals are all known from body fossil record of the Middle Jurassic of the British Isles, and it is certainly likely that they were part of the ecosystem in the Cleveland Basin during the Middle Jurassic, but confirmation of this must await future finds.
The discovery of the Enigmatic Burniston Track augments our knowledge of the Middle Jurassic vertebrate assemblages of this period and region. Although the specimen of EBT has been lost, we hope the discovery and recording of this trackway will initiate a search for further material, which will clarify the identification and permit a more confident assignment of higher level ichnotaxonomy in the future.
Acknowledgments
Thanks to the Leeds Geological Association for sharing their photographs of their discovery and allowing us to publish on it. EP was supported by NERC (NE/L002558/1) during the initial phases on manuscript preparation. Thanks to Professor Martin Lockley for providing MR with publications, and to our reviewers J Farlow, H Klein and SH Lucas, and our editor L Buatois, for their helpful and constructive feedback, which greatly improved this manuscript.
Disclosure statement
No potential conflict of interest was reported by the authors.
References
- Alexander, R. M. (1976). Estimates of speeds of dinosaurs. Nature , 261 (5556), 129–130.
- Angielczyk, K. D. (2009). Dimetrodon is not a dinosaur: Using tree thinking to understand the ancient relatives of mammals and their evolution. Evolution: Education and Outreach , 2 , 257–271.
- Avanzini, M. , & Renesto, S. (2002). A review of Rhynchosauroides tyrolicus Abel, 1926 ichnospecies (Middle Triassic: Anisian-Ladinian) and some inferences on Rhynchosauroides trackmaker. Rivista Italiana di Paleontologia e Stratigrafia , 108 , 51–66.
- Avanzini, M. , García-Ramos, J. C. , Lires, J. , Menegon, M. , Piñuela, L. , & Fernández, L. A. (2005). Turtle tracks from the Late Jurassic of Asturias. Spain. Acta Palaeontological Polonica , 50 , 743–755.
- Avanzini, M. García-Ramos, J. C., Lires, J., Piñuela, L., & Lockley, M. G. (2007). Crocodylomorph tracks from the late Jurassic of Asturias (Spain). Ichnos, 14(1–2), 143–153.
- Avanzini, M. , & Mietto, P. (2008). The occurrence of the vertebrate ichnogenus Synaptichnium in the Anisian (Middle Triassic) of Southern Alps. Studi Trentini di Scienze Naturali Acta Geologica , 83 , 13–19.
- Baird, D. (1957). Triassic reptile faunules from Milford, New Jersey. Bulletin of the Museum of Comparative Zoology , 117 , 449–520.
- Barron, A. J. M. , Lott, G. K. , & Riding, J. B. (2012). Stratigraphical framework for the Middle Jurassic strata of Great Britain and the adjoining continental shelf . British Geological Survey, 177p.
- Casamiquela, R. M. (1961). Sobre la presencia de un mamífero en el primer elenco (icnológico) de vertebrados del Jurásico de la Patagonia. Physis , 22 , 225–233.
- Contessi, M. (2013). First report of mammal-like tracks from the Cretaceous of North Africa (Tunisia). Cretaceous Research , 42 , 48–54.
- Day, J. J. , Norman, D. B. , Gale, A. S. , Upchurch, P. , & Powell, H. P. (2004). A Middle Jurassic dinosaur trackway site from Oxfordshire. Palaeontology , 47 (2), 319–348.
- de Valais, S. (2009). Ichnotaxonomic revision of Ameghinichnus, a mammalian ichnogenus from the middle Jurassic La Matilde formation, Santa Cruz province, Argentina. Zootaxa , 2203 (1), e21–21.
- dePolo, P. E. , Brusatte, S. L. , Challands, T. J. , Foffa, D. , Ross, D. A. , Wilkinson, M. , & Yi, H. Y. (2018). A sauropod-dominated tracksite from Rubha nam Brathairean (Brothers’ Point), Isle of Skye, Scotland. Scottish Journal of Geology , 54 (1), 1–12.
- dePolo, P. E. , Brusatte, S. L. , Challands, T. J. , Foffa, D. , Wilkinson, M. , Clark, N. D. L. , Hoad, J., Pereira, P. V. L. G. C., Ross, D. A., & Wade, T. J. (2020). Novel track morphotypes from new tracksites indicate increased Middle Jurassic dinosaur diversity on the Isle of Skye, Scotland. PlosONE , 15(3), e0229640.
- Diedrich, C. (2002). Vertebrate track bed stratigraphy at new megatrack sites in the Upper Wellenkalk member and orbicularis member (Muschelkalk, Middle Triassic) in carbonate tidal flat environments of the western Germanic Basin. Palaeogeography, Palaeoclimatology, Palaeoecology , 183 (3–4), 185–208.
- Ellenberger, P. (1974). Contribution a la classification des pistes de vertebres du 650 Trias; les types du Stormberg d’Afrique du Sud, 2e partie. Palaeovertebrata, Memoire Extraordinaire, Laboratoire de Paleontologie des Vertebres, Montpellier, 170p.
- Evans, S. , Barrett, P. , Hilton, J. , Butler, R. J. , Jones, M. E. H. , Liang, M.-M. , Parish, J., Rayfield, E. J., Sigogneau-Russell, D., & Underwood, C. J. (2006). The Middle Jurassic vertebrate assemblage of Skye, Scotland. In P. Barrett & S. Evans (Eds.), Proceedings of the Ninth Symposium on Mesozoic Terrestrial Ecosystems and Biota , Natural History Museum, London.
- Evans, S. E. , & Milner, A. R. (1994). Middle Jurassic microvertebrate assemblages from the British Isles. In N. C. Fraser & H.-D. Sues (Eds.), In the shadow of the dinosaurs: Early Mesozoic tetrapods . Cambridge University Press, 429p.
- Falkingham, P. L. (2014). Interpreting ecology and behaviour from the vertebrate fossil track record. Journal of Zoology , 292 (4), 222–228.
- Farlow, J. O. , Robinson, N. J. , Kumagai, C. J. , Paladino, F. V. , Falkingham, P. L. , Elsey, R. M. , & Martin, A. J. (2018). Trackways of the American crocodile (Crocodylus acutus) in northwestern Costa Rica: Implications for crocodylian ichnology. Ichnos , 25 (1), 30–65.
- Hargreaves, J. A. (1914). Fossil footprints near Scarborough. The Naturalist , 688 , 154–156.
- Haubold, H. (1971a). Ichnia amphibiorum et reptiliorum fossilium. In O. Kuhn (Ed.), Handbuch der Paläoherpetologie, part. 18 . Gustav Fischer Verlag, 121p.
- Haubold, H. (1971b). Die Tetrapodenfährten des Buntsandsteins in der Deutschen Demokratischen Republik und in Westdeutschland und ihre Äquivalente in der gesamten Trias. Paläontologische Abhandlungen , 4 , 3952013548.
- Hitchcock, E. (1845). An attempt to name, classify, and describe the animals that made the fossil footmarks of New England. In Proceedings of the 6th Annual Meeting of the Association of American Geologists and Naturalists, New Haven, CT (Vol. 6, pp. 23–25).
- Hoffman, E. A. , & Rowe, T. B. (2018). Jurassic stem-mammal perinates and the origin of mammalian reproduction and growth. Nature , 561 , 104–108.
- Hu, Y. , Meng, J. , Wang, Y. , & Li, C. (2005). Large Mesozoic mammals fed on young dinosaurs. Nature , 433 , 149–152.
- Ivens, C. R. , & Watson, N. G. (1994). Records of dinosaurs of the North East Yorkshire coast 1895–1993 . Rosebury Publications, 20p.
- Jardine, W. (1850). Note to Mr. Harkness’s paper “On the position of the impressions of footsteps in the Bunter sandstones of Dumfriesshire. Annals and Magazine of Natural History , 6 , 208–209.
- Jenkins, F. A. Jr. (1971). The postcranial skeleton of African cynodonts. Peabody Museum of. Natural History Bulletin , 36 , 1–216.
- Ji, Q. , Luo, Z. X. , Yuan, C. X. , & Tabrum, A. R. (2006). A swimming mammaliaform from the Middle Jurassic and ecomorphological diversification of early mammals. Science , 311 (5764), 1123–1127.
- Kemp, T. S. (2005). The origin and evolution of mammals . Oxford University Press, 342p.
- Kielan-Jaworowska, Z. , & Hurum, J. H. (2006). Limb posture in early mammals: Sprawling or parasagittal. Acta Palaeontologica Polonica , 51 , 393–406.
- Kim, K. S. , Lim, J. D. , Lockley, M. G. , Xing, L. , & Choi, Y. (2017). Korean trackway of a hopping, mammaliform trackmaker is first from the Cretaceous of Asia. Cretaceous Research , 74 , 188–191.
- Kim, K. S. , Lockley, M. G. , Lim, J. D. , Bae, S. M. , & Romilio, A. (2020). Trackway evidence for large bipedal crocodylomorphs from the Cretaceous of Korea. Scientific Reports , 10 , 1–13.
- King, M. J. , Sarjeant, W. A. S. , Thompson, D. B. , & Tresise, G. (2005). A revised systematic ichnotaxonomy and review of the vertebrate footprint ichnofamily Chirotheriidae from the British Triassic. Ichnos , 12 (4), 241–299.
- Kubo, T. (2008). In quest of the Pteraichnus trackmaker: Comparisons to modern crocodilians. Acta Palaeontologica Polonica , 53 (3), 405–412.
- Kühne, W. G. (1956). The liassic therapsid Oligokyphus. British Museum of Natural History, 174p.
- Kümmell, S. B. , & Frey, E. (2012). Digital arcade in the autopodia of Synapsida: Standard position of the digits and dorsoventral excursion angle of digital joints in the rays II–V. Palaeobiodiversity and Palaeoenvironments , 92 (2), 171–196.
- Leonardi, G. (1981). Novo icnogenero de tetrapode mesozoico da formacao Botucatu, Araraquara, SP. Anais da Academia Brasileira de Ciências , 53 , 793–805.
- Leonardi, G. (1987). Glossary and manual of tetrapod footprint palaeoichnology. Departamento Nacional da Produçao Mineral, 137p.
- Lockley, M. G. , & Foster, J. (2006). Dinosaur and turtle tracks from the Morrison Formation (Upper Jurassic) of Colorado National Monument, with observations on the taxonomy of vertebrate swim tracks. New Mexico Museum of Nature and Science, Bulletin , 36 , 193–198.
- Lockley, M. , & Foster, J. (2003). Late Cretaceous mammal tracks from North America. Ichnos , 10 (2-4), 269–276.
- Lockley, M. G. , & Hunt, A. P. (1995). Dinosaur tracks and other fossil footprints of the western United States . Columbia University Press, 338p.
- Lockley, M. G. , Logue, T. J. , Moratalla, J. J. , Hunt, A. P. , Schultz, R. J. , & Robinson, J. W. (1995). The fossil trackway Pteraichnusis is pterosaurian, not crocodilian: Implications for the global distribution of pterosaurs tracks. Ichnos, 4 (1), 7–20.
- Lockley, M. G. , Lucas, S. G. , Hunt, A. P. , & Gaston, R. (2004). Ichnofaunas from the Triassic-Jurassic boundary sequences of the Gateway area, western Colorado: Implications for faunal composition and correlations with other areas. Ichnos , 11 (1–2), 89–102.
- Lockley, M. , Lucas, S. , Milàn, J. , Harris, J. , Avanzini, M. , Foster, J. R. , & Spielmann, J. (2010). The fossil record of crocodilian tracks and traces: An overview. In J. Milàn , S. Lucas , M. Lockley , J. Spielmann (Eds.), Crocodyle tracks and traces (Vol. 51). New Mexico Museum of Natural History Bulletin, 244p.
- Lockley, M. G. , & Meyer, C. (2004). Crocodylomorph trackways from the Jurassic to Early Cretaceous of North America and Europe: implications for ichnotaxonomy. Ichnos , 11 (1–2), 167–178.
- Lull, R. S. (1904). Fossil Footprints of the Jura-Trias of North America. Memoirs of the Boston Society of Natural History , 5 , 461–557.
- Luo, Z.-X. , & Wible, J. R. (2005). A Late Jurassic digging mammal and early mammalian diversification. Science , 308 , 103–107.
- Marchetti, L. , Klein, H. , Buchwitz, M. , Ronchi, A. , Smith, R. M. , De Klerk, W. J. , Sciscio, L., & Groenewald, G. H. (2019). Permian-Triassic vertebrate footprints from South Africa: Ichnotaxonomy, producers and biostratigraphy through two major faunal crises. Gondwana Research , 72 , 139–168.
- Mateus, O. , Marzola, M. , Schulp, A. S. , Jacobs, L. L. , Polcyn, M. J. , Pervov, V. , Gonçalves A. O., & Morais, M. L. (2017). Angolan ichnosite in a diamond mine shows the presence of a large terrestrial mammaliamorph, a crocodylomorph, and sauropod dinosaurs in the Early Cretaceous of Africa. Palaeogeography, Palaeoclimatology , Palaeoecology , 471 , 220–232.
- Matsuoka, H. , Kusuhashi, N. , & Corfe, I. J. (2016). A new Early Cretaceous tritylodontid (Synapsida, Cynodontia, Mammaliamorpha) from the Kuwajima Formation (Tetori Group) of central Japan. Journal of Vertebrate Paleontology , 36 (4), e1112289.
- Melchor, R. N. , & de Valais, S. (2006). A review of Triassic tetrapod track assemblages from Argentina. Palaeontology , 49 (2), 355–379.
- Milan, J. , & Hedegaard, R. (2010). Interspecific variation in tracks and trackways from extant crocodylians. In J. Milàn , S. Lucas , M. Lockley , & J. Spielmann (Eds.), Crocodyle tracks and traces (Vol. 51). New Mexico Museum of Natural History Bulletin, 244p.
- Nopcsa, F. (1923). Die Familen der Reptilien. Fortschritte Der Geologie Und Paläeontologie , 2 , 1–210.
- Olsen, P. E. , & Padian, K. (1986). Earliest records of Batrachopus from the southwestern United States, and a revision of some Early Mesozoic crocodylomorph ichnogenera. In K. Padian (ed.), The beginning of the age of the dinosaurs . Cambridge University Press, 404p.
- Padian, K. , & Olsen, P. E. (1984). The fossil trackway Pteraichnus: Not pterosaurian, but crocodilian. Journal of Paleontology , 58 , 178–184.
- Panciroli, E. , Benson, R. B. J. , & Walsh, S. (2017b). The dentary of Wareolestes rex (Megazostrodontidae): A new specimen from Scotland and implications for morganucodontan tooth replacement. Papers in Palaeontology , 3 (3), 373–386.
- Panciroli, E. , Benson, R. B. J. , & Walsh, S. (2018b). The mammal-rich freshwater assemblage of the Middle Jurassic Kilmaluag Formation, Isle of Skye, Scotland. Abstracts 13th Mesozoic Terrestrial Ecosystems and Biota, Bonn, Germany.
- Panciroli, E. , Benson, R. B. J. , Walsh, S. , Butler, R. J. , Castro, T. A. , Jones, M. E. H. , & Evans, S. E. (2020). Diverse vertebrate assemblage of the Kilmaluag Formation (Bathonian, Middle Jurassic) of Skye, Scotland. Earth and Environmental Transactions of the Royal Society of Edinburgh , 111, 135–156.
- Panciroli, E. , Schultz, J. A. , & Luo, Z.-X. (2018a). Morphology of the petrosal and stapes of Borealestes (Mammaliaformes, Docodonta) from the Middle Jurassic of Skye, Scotland. Papers in Palaeontology , 5 (1), 139–156.
- Panciroli, E. , Walsh, S. , Fraser, N. C. , Brusatte, S. L. , & Corfe, I. (2017a). A reassessment of the postcanine dentition and systematics of the tritylodontid Stereognathus (Cynodontia, Tritylodontidae, Mammaliamorpha), from the Middle Jurassic of the UK. Journal of Vertebrate Paleontology , 37 (5), e1351448.
- Porchetti, S. D. O. , Bertini, R. J. , & Langer, M. C. (2017). Walking, running, hopping: Analysis of gait variability and locomotor skills in Brasilichnium elusivum Leonardi, with inferences on trackmaker identification. Palaeogeography, Palaeoclimatology, Palaeoecology , 465 , 14–29.
- Rasband, W. S. (1997–2018). ImageJ, U. S. National Institutes of Health, Bethesda, Maryland, USA. https://imagej.nih.gov/ij/.
- Rawson, P. F. , & Wright, J. K. (2018). Geology of the Yorkshire Coast (4th ed.). Geologists’ Association Guide 34, 178pp.
- Retallack, G. J. (1996). Early Triassic therapsid footprints from the Sydney basin. Alcheringa: An Australasian Journal of Palaeontology , 20 (4), 301–314.
- Romano, M. , Clark, N. D. L. , & Brusatte, S. L. (2018). A comparison of the dinosaur communities from the Middle Jurassic of the Cleveland (Yorkshire) and Hebrides basins, based on their ichnites. Geosciences , 8 (327), 1–22.
- Romano, M. , & Whyte, M. A. (2003a). Jurassic dinosaur tracks and trackways of the Cleveland Basin, Yorkshire: Preservation, diversity and distribution. Proceedings of the Yorkshire Geological Society , 54, 185–215.
- Romano, M. , & Whyte, M. A. (2003b). The first record of xiphosurid (arthropod) trackways from the Saltwick Formation, Middle Jurassic of the Cleveland Basin, Yorkshire. Palaeontology , 46 (2), 257–269.
- Romano, M. , & Whyte, M. A. (2010). Crocodilian and other non-dinosaurian tracks and trackways from the Ravenscar Group (Middle Jurassic) of the Cleveland Basin, Yorkshire, UK. In J. Milàn , S. Lucas , M. Lockley , & J. Spielmann (Eds.), Crocodyle tracks and traces (Vol. 51, 244p). New Mexico Museum of Natural History Bulletin.
- Rowe, T. B. (1988). Definition, diagnosis and origin of Mammalia. Journal of Vertebrate Paleontology , 8 (3), 241–264.
- Rowland, S. M. , & Mercadante, J. M. (2014). Trackways of a gregarious, dunefield-dwelling, Early Jurassic therapsid in the Aztec Sandstone of Southern Nevada. PALAIOS , 29 (10), 539–552.
- Rühle von Lilienstern, H. (1944). Eine Dicynodontierführten aus dem Chiretheriumsandstein von Hessberg. Päläontologische Zeitschrift , 23 , 368–385.
- Sarjeant, W. A. S. (1975). A vertebrate footprint from the Stonesfield Slate (Middle Jurassic) of Oxfordshire. Mercian Geologist , 5 , 273–277.
- Sarjeant, W. A. S. (1995). Footprints in the sands of time. Vertebrate footprints and the interpretation of past environments. Geoscience Canada , 21 , 77–87.
- Stanford, R. , Lockley, M. G. , Tucker, C. , Godfrey, S. , & Stanford, S. M. (2018). A diverse mammal-dominated, footprint assemblage from wetland deposits in the Lower Cretaceous of Maryland. Scientific Reports , 8(741), 1–12.
- Stanford, R. , Lockley, M. , & Weems, R. (2007). Diverse dinosaur-dominated ichnofaunas from the potomac group (Lower Cretaceous) Maryland. Ichnos , 14 (3–4), 155–173.
- Sulej, T. , & Niedźwiedzki, G. (2019). An elephant-sized Late Triassic synapsid with erect limbs. Science , 363 (6422), 78–80.
- Whyte, M. A. , & Romano, M. (2014). First record of the pterosaur footprint Pteraichnus from the Saltwick Formation (Aalenian) of the Cleveland Basin, Yorkshire, UK. Proceedings of the Yorkshire Geological Society , 60, 19–27.
- Whyte, M. A. , Romano, M. , & Elvidge, D. J. (2007). Reconstruction of Middle Jurassic dinosaur-dominated communities from the vertebrate ichnofauna of the Cleveland Basin of Yorkshire, UK. Ichnos , 14 (1–2), 117–129.
- Zhou, C. F. , Wu, S. , Martin, T. , & Luo, Z. X. (2013). A Jurassic mammaliaform and the earliest mammalian evolutionary adaptations. Nature , 500 (7461), 163–167.