
Abstract
Large animal tracks, unequivocally attributable to terrestrial mammals, are reported for the first time in sediment from uppermost Bed I (Tuff IF; ∼1.803 million years ago) at Olduvai Gorge, Tanzania. One track in particular (attributed to the ichnogenus Pecoripeda) retains an exceptional level of detail, demonstrating the excellent trackway-preserving potential of the volcanic ash fall (tuff) layers at this important hominin archaeological locality. Olduvai Gorge is renowned for its abundant Plio-Pleistocene (zoo)archaeological discoveries and fossiliferous deposits vis-à-vis studies of human evolution. Fossil trackways, and trace fossils more widely, provide an important additional tool for characterizing ancient ecosystems, which remain underexplored at Olduvai. Considered together with fossil hominin remains, information derived from coeval fossil animal tracks provides additional insight into our ancestors’ behaviour and their interactions with the surrounding palaeoenvironment. A range of large herbivore tracks indicates the availability of nearby resources (i.e., freshwater, vegetation preferred by grazers/browsers). These newly-discovered tracks are of archaeological and palaeontological significance because they highlight the potential for future discovery of animal or hominin tracks and trackways preserved in tuff at Olduvai and in other archaeological localities.
Introduction
Fossil assemblages of bones and teeth provide an excellent basis for interpreting ancient ecosystems. However, the processes involved in the burial and fossilization of animal material often results in biased assemblages. Over time, taphonomic processes (i.e., mode of deposition, scavenging, chemical and physical weathering at surface, and post-burial diagenesis) alter the fossil assemblage from that of the original animal community. True species abundances are commonly distorted and species richness is underrepresented (Behrensmeyer, Citation1978). For example, disarticulated skeletal remains can be transported long distances (i.e., by water or predators) or a specific species may be over-represented in one location (i.e., predator bias). These processes can obscure the original ecological provenience of an ancient animal. Fossil tracks and trackways associated with fossil bone assemblages can provide additional information about faunal communities and help alleviate some of the taphonomic biases.
Tracks and other trace fossils provide direct evidence that the track maker was present in a specific environment or location. Tracks can also convey aspects of behaviour, such as mode and speed of locomotion, and even inter/intra-species interactions if multiple individuals are represented on a single tracking surface (Cohen et al., 1993; Lockley, Citation1998). Despite the preservation bias towards larger, heavier animals, tracks and trackway assemblages provide complementary palaeoecological information that helps to eliminate some preservation biases associated with fossil assemblages.
Fossil trackways in Africa
Despite their overall rarity, animal and hominin trackways are well described from Plio-Pleistocene sediments from East and South Africa (). Some track sites are critical to our understanding of hominin evolution. The 3.6-million-year-old Pliocene hominin trackways preserved in volcanic ash at the Laetoli site (40 km south of Olduvai Gorge) remain the earliest unequivocal evidence of bipedalism in early human ancestors (Day & Wickens, Citation1980; Leakey, Citation1978a; Citation1978b; Raichlen et al., Citation2010). Hundreds of posited Homo erectus/ergaster tracks preserved in ancient lakeshore sediments from the Ileret site (Koobi Fora Formation) near Lake Turkana, Kenya demonstrate that these early human ancestors utilized lakeshore habitats and evolved an essentially modern style of locomotion and foot function by 1.5 million years ago (Bennett et al., Citation2009; Roach et al., Citation2016; Citation2018). Fossil footprints attributed to Homo erectus are reported from the Danakil Desert, Eritrea from an ancient lake shore deposit dated to ∼800 ka (Archaeology News Network, Citation2016). Additional sites within the Koobi Fora Formation record Pleistocene bipedal hominin and large vertebrate tracks and trackways - including rare examples of trackways produced by swimming hippopotami (Behrensmeyer & Laporte, Citation1981; Bennett et al., Citation2009; Citation2014; Brown et al., Citation2006; Hatala et al., Citation2017; Roach et al., Citation2016).
Figure 1. African Plio-Pleistocene and Holocene records of hominin and large animal fossil track sites. 1 – Danakil Desert, Eritrea (∼800 ka) (Archaeology News Network, Citation2016); 2 – Melka Kunture (Upper Awash Valley), Ethiopia (∼700 ka) (Altamura et al., Citation2018); 3 – Koobi Fora (∼1.5 Ma), Ileret (∼1.52 Ma), and GaJi10 sites (∼1.43 Ma) (Okote Member, Koobi Fora Formation), Lake Turkana, Kenya (Behrensmeyer & Laporte, Citation1981; Bennett et al., Citation2009; Citation2014; Brown et al., Citation2006; Hatala et al., Citation2017; Roach et al., Citation2016); 4 – Sandai Plain (Loboi Silts), Lake Bogoria, Kenya (Late Pleistocene) (Scott et al., Citation2008); 5 – Engare Sero, Lake Natron, Tanzania (5760 ± 30 to 19.1 ± 3.1 ka) (Balashova et al., Citation2016; Hatala et al., Citation2020; Liutkus-Pierce et al., Citation2016; Zimmer et al., Citation2018); 6 – Laetoli, Tanzania (∼3.6 Ma) (Day & Wickens, Citation1980; Leakey, Citation1978b; Raichlen et al., Citation2010); 7 – Nahoon Site, Eastern Cape Province, South Africa (∼124 ka) (Jacobs & Roberts, Citation2009; Roberts, Citation2008); 8 – Brenton-on-Sea, Western Cape Province, South Africa (∼90 ka) (Helm, McCrea, et al., Citation2018); 9 – Still Bay, Western Cape Province, South Africa (∼90 ka) (Helm, Cawthra, de Vynck, et al., Citation2019; Roberts et al., Citation2008); 10 – Langbaan Lagoon, Western Cape Province, South Africa (∼117 ka) (Berger & Hilton-Barber, Citation2000; Roberts & Berger, Citation1997); 11 – Namib Sand Sea, Walvis Bay, Namibia (late Holocene) (Morse et al., Citation2013); 12 – Cape south coast sites, South Africa (Late Pleistocene) (Helm, Cawthra, Combrink, et al., Citation2020; Helm, Cawthra, Cowling, et al., Citation2020; Helm, Cawthra, de Vynck, et al., Citation2019; Helm, Cawthra, et al., Citation2018; Helm et al., Citation2017; Helm et al., Cawthra, Hattingh, 2019; Roberts, Citation2008; Roberts et al., Citation2008).
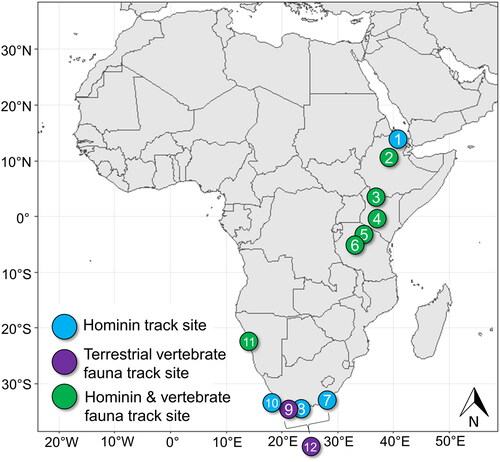
Additional records of Pleistocene hominin/early Homo sapiens and vertebrate fauna track sites from East Africa are rare. Tracks are typically preserved in palaeolake margin settings or ancient volcanic ash. Known sites include a single hominin track at the Sandai Plain site at Lake Bogoria, Kenya and the Engare Sero site at Lake Natron, Tanzania (Balashova et al., Citation2016; Hatala et al., Citation2020; Liutkus-Pierce et al., Citation2016; Scott et al., Citation2008; Zimmer et al., Citation2018). The Engare Sero site yielded over 400 human footprints attributed to a (predominantly female) group of 16 individuals, providing insight into early human group composition and dynamics (Hatala et al., Citation2020). An extensive record of hominin and animal (bovid, hippopotamus, equid and bird) tracks are reported from the ∼700 ka Melka Kunture site (Upper Awash Valley) in Ethiopia (Altamura et al., Citation2018). The surface may indicate the presence very young children and is preserved alongside coeval archaeological evidence of flint knapping and a butchered hippopotamus carcass (Altamura et al., Citation2018).
Abundant Quaternary fossil track sites are reported from South Africa from the coastal aeolianites of the Cape south coast. These include a single record of forty hominin tracks (90 ka), making it the best preserved late Pleistocene archive of its kind for the region (Helm, McCrea, et al., Citation2018; Roberts, Citation2008). Additional late Pleistocene human track sites from the region include Nahoon (∼124 ka) in Eastern Cape Province, and Brenton-on-Sea (∼90 ka) and Langbaan Lagoon (∼117 ka), both in Western Cape Province (Berger & Hilton-Barber, Citation2000; Helm, McCrea, et al., Citation2018; Jacobs & Roberts, Citation2009; Roberts, Citation2008; Roberts & Berger, Citation1997). Numerous fossil animal tracks and trackway sites preserved in aeolianites (i.e., from Witsand to Robberg Nature Reserve) that represent contemporary animal genera were also discovered along the Cape south coast (Helm, Cawthra, Combrink, et al., Citation2020; Helm, Cawthra, Cowling, et al., Citation2020; Helm, Cawthra, de Vynck, et al., Citation2019; Helm, Cawthra, Hattingh, et al., 2019; Helm, Cawthra, et al., Citation2018; Helm et al., Citation2017; Roberts, Citation2008; Roberts et al., Citation2008). Megafauna are well-represented at these sites (e.g., artiodactyls, giraffe, elephant, rhinoceros, long-horned buffalo, giant Cape horse), as well as reptiles and birds (Helm, Cawthra, Combrink, et al., Citation2020; Helm, Cawthra, de Vynck, et al., Citation2019; Helm et al., Cawthra, Hattingh, 2019). Records are currently sparse for the west coast of Africa, although a late Holocene human and animal (bovid, giraffe, elephant, carnivore, and bird) track site is reported from the Namib Sand Sea, near Walvis bay, Namibia (Morse et al., Citation2013).
Palaeogeography and Ecology of Olduvai Gorge
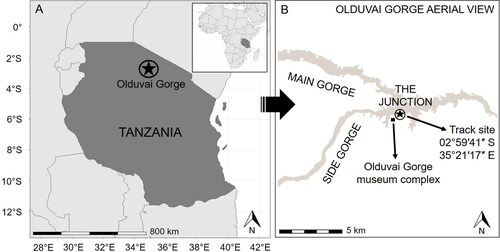
Olduvai Gorge is on the western margin of the East African Rift Valley system (). The contemporary gorge is situated between the Serengeti Plains to the west and the Ngorongoro volcanic highlands to the east. Prior to incision of the modern gorge by fluvial processes ∼50 ka, the palaeoenvironment included a saline-alkaline lake and less saline wetland lake-margin habitats.
Figure 2. A. Olduvai Gorge, northern Tanzania. B. Olduvai Gorge, with the location of the fossil track site indicated by a star.
Saline-alkaline lake and lakeshore habitats dominated the Olduvai Gorge landscape during Bed I time (∼2.1 to 1.75 Ma). Throughout this interval, the Bed I sedimentary record indicates that a mosaic of habitats existed, including lake, lake-margin, wetland, woodland (>80% tree [canopy] cover), and grassland (<10% tree cover) (Ashley et al., Citation2009; Hay, Citation1976; Magill et al., Citation2013; Citation2016). Groundwater springs were also present close to the lake. Palaeo-lake Olduvai expanded and contracted with seasonal and global changes in rainfall patterns (Ashley & Hay, Citation2002; Hay, Citation1976; Hay & Kyser, Citation2001; McHenry, Citation2012). The surrounding palaeoenvironment transitioned through time from closed woodland habitat during middle Bed I time, to more open woodland and grassland during lower Bed II time (Bibi et al., Citation2018; Fernández-Jalvo et al., Citation1998). However, this transition was not gradual or unidirectional (Magill et al., Citation2013). The overall trend towards aridity in East Africa coincided with high latitude glacial cycles (DeMenocal, Citation1995; Citation2004; Shackleton, Citation1995).
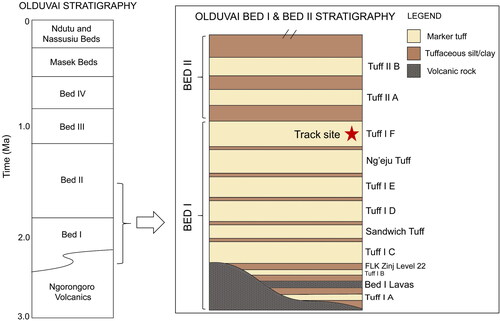
At Olduvai Gorge, the lacustrine and near-shore sedimentary record from the late Pliocene to the Holocene is rich in faunal remains, early hominin fossils (Paranthropus boisei, Homo habilis, Homo erectus), and stone tools from the Oldowan and Acheulian industries (Bibi et al., Citation2018; Clarke, Citation2012; Diez-Martín et al., Citation2015; Domínguez-Rodrigo et al., Citation2013; Egeland, Citation2007; Kimura, Citation2002; Kovarovic et al., Citation2013; Leakey, Citation1971). The stratigraphic succession () comprises alternating weathered clays and well-dated volcanic ash deposits (marker tuffs). A diverse Pleistocene faunal community occupied the landscape at Olduvai Gorge during Bed I and II times. The large herbivore fossil assemblage is composed of Bovidae (Alcelaphini, Antilopini, Tragelaphini, and Bovini tribes), Suidae, Proboscidea, Hippopotamidae, Giraffidae, and Rhinocerotidae. Large-bodied carnivores – Canidae, Felidae, and Hyaenidae – preyed upon the diverse range of herbivores (Egeland, Citation2007). This well-dated stratigraphic succession provides an important window into early human evolution and the palaeoecology of the East African landscape over the past 4 million years.
Figure 3. A simplified stratigraphic column for Bed I and Lower Bed II at Olduvai Gorge (based on geological information from Reck, Citation1914, Hay, Citation1963, Citation1976, Leakey, Citation1978a, McHenry, Citation2012, Bibi et al., Citation2018, and Uno et al., Citation2018).
Methods
A single locality that exposed track-bearing surface(s) within Tuff IF was identified and surveyed in June 2019. The tracks and track casts were photographed and measured, while stratigraphic context was identified on site.
Results
Locality: Olduvai Gorge, Ngorongoro Conservation Area, Northern Tanzania
Stratigraphic context: Tuff IF, upper Bed I
Outcrop coordinates: 02°59′41″ S and 35°21′17″ E
Site elevation: 1430 m
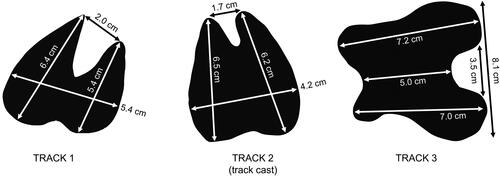
Multiple, large-bodied terrestrial animal tracks were discovered in an exposure of Tuff IF, upper Bed I at Olduvai Gorge, Tanzania ( and ). Tracks and track dimensions are presented in . Tuff IF is an ∼1.80 Ma deposit that contains reworked, high-energy pyroclastic material (Bibi et al., Citation2018; Deino, Citation2012). Tuff IF caps the stratigraphic sequence of Bed I and is sandwiched between the Ng’eju tuff and the base of Bed II ().
Figure 4. Block of pyroclastic tuff containing Track 1. The fossil track (A) and corresponding track cast (B) are preserved with a high degree of detail. The marginal ridge is indicated by a white arrow.

Figure 5. A. Block of pyroclastic tuff containing Track 2. Only the fossil track cast was located. B. The concave reverse impression created by the marginal ridge surrounding Track 2 (indicated with dashed white line). The impression left by the keratinous ridge on the underside of the hoof is also visible (indicated by a white arrow).

Figure 6. Block of pyroclastic tuff containing Track 3. The fossil track (A) and corresponding track cast (B) are preserved with minimal detail.

Figure 7. Track measurements.
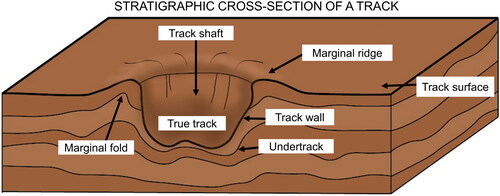
The exposure is approximately 345 m northeast of the Olduvai Gorge museum building complex. Three individual blocks of laminated tuff containing fossil animal tracks were discovered eroding out of the same exposure (). The three separate tracks were found within 10 m of one another. Animal tracks preserved in fine-grained sediment produce morphologically distinct patterns (). This can include true tracks that formed in contact with the foot of the animal, undertracks, and various styles of sediment deformation peripheral to the track. None of the discovered tracks appeared to be over-printed.
Figure 8. The exposure of Tuff IF, Bed I, where the tracks were discovered. A block of tuff containing a footprint is indicated by arrow ‘A’, and scale is indicated by geological hammer (32 cm long) at arrow ‘B’. Photograph taken facing East.

Figure 9. A simplified block diagram illustrating the morphological features of a track in cross-section. Modified from Allen (Citation1997) and Melchor (Citation2015).
The degree of preservation and the level of detail varies among the three separate tracks. This could indicate i) the presence of multiple track surfaces within the same ash layer or ii) that the same continuous track-bearing surface had spatially-variable consistency (i.e., more or less cohesive) at the time the tracks were made.
Discussion
Ichnotaxonomy
Ichnogenus Pecoripeda (Vyalov, Citation1965; Citation1966)
Ichnospecies Pecoripeda isp.
Descriptions:
Track 1: Single print produced by an artiodactyl (cloven-hoofed animal) (). The lateral digit impression is larger than the medial digit impression, which is indicative of a left-side track. Track and corresponding track cast () were discovered near one another. This substrate was cohesive at the time of print formation, as demonstrated by the crisp detail and the presence of a marginal ridge surrounding the print (). In addition, the sharp edge of the underside of the keratinous hoof-wall has left a clear indentation in the tuff.
Track 2: Single track cast produced by an artiodactyl . The indentations created by the anterior tips of the hooves are clearly visible (). The track surface substrate was cohesive at the time of print formation, and a concave reverse impression of a marginal ridge is visible (). The keratinous ridge on the underside of the tips of the hooves is also visible (). A stratigraphically lower surface preserving silicified roots was present in the same block of tuff.
The Cenozoic vertebrate ichnotaxonomic group Pecoripeda incorporates all footprints of ruminant artiodactyls that possess horns and antlers (except for Tylopoda, or camels and camelids) (Lucas & Hunt, Citation2007; Vyalov, Citation1965; Citation1966). This ichnogenus includes giraffes, bovids, and antelopes. Tracks from this ichnogenus are characterized by elongated, wedge-shaped tracks with two mirror-image hooves separated by an interdigit space (Bromley et al., Citation2009; Morgan & Williamson, Citation2000; Sarjeant & Langston, Citation1994; Vyalov, Citation1965).
Track 3: Single print produced by a large even-toed ungulate. Track and corresponding track cast were discovered together (). The track was infilled with coarse-grained sediment. The track is poorly preserved with minimal detail, and the marginal ridge is very weakly developed.
The asymmetrical track shape and dimensions suggest a large track maker belonging to the ichnogenus Megapecoripeda (Hágen et al., Citation2014; Kordos, Citation1985). The Cenozoic vertebrate ichnotaxonomic grouping Megapecoripeda incorporates the larger even-toed ungulates. Tracks from this ichnogenus are larger than Pecoripeda and are characterized by a medial digit shifted distally and slightly diverging toes (Kordos, Citation1985, Hágen et al., Citation2014).
Vertebrate palaeoecology
Tracks unequivocally attributable to large terrestrial mammals have not been previously reported from Pleistocene deposits at Olduvai Gorge. Undulating palaeo-wetland sediments at Olduvai have been attributed to widespread trampling (bioturbation) by large vertebrates, and large trough-shaped structures in lowermost Bed II claystones are hypothesized to be ancient hippo trails (Ashley, Citation2003). However, the only confirmed fossil animal tracks at Olduvai occur in the Bird Print Tuff (BPT) in lower Bed II. The BPT is a yellow laminated vitric tuff that contains abundant footprints of shore birds (Hay, Citation1976).
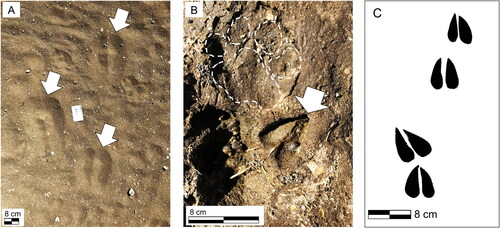
All three tracks appear morphologically consistent with those made by members of Artiodactyla (even-toed ungulates). This is unsurprising, given that members of Bovidae dominate the fossil assemblages of Bed I and II (Bibi et al., Citation2018; Egeland, Citation2007; Gentry & Gentry, Citation1978). Tracks 1 and 2 are morphologically consistent with those made by medium-bodied members of Artiodactyla, possibly Bovidae. Comparison with modern African Bovidae tracks (), both in the field and in reference guides, suggest that Track 1 was made by an animal similar in size and hoof-morphology to a modern Waterbuck (weighing 130 kg or more) (Liebenberg, Citation1990). A smaller member of Bovidae, perhaps belonging to Antilopini, may have made Track 2.
Figure 10. Modern artiodactyl tracks from Olduvai Gorge region. A. Fresh giraffe (Giraffa camelopardalis) footprints in loose sandy sediment from the plains surrounding Olduvai Gorge. The tracks were produced by a small herd of individuals of different ages/sizes. B. Arrow indicating medium-sized bovid footprint (probably wildebeest, gazelle, or antelope) in the soft muddy lakeshore sediments of the present-day saline/alkaline Lake Ndutu, Tanzania (situated approximately 35 km west of Olduvai Gorge). Dashed white outline indicates a carnivore track in the same image. Photographs from June 2019. C. Outline of a modern Waterbuck trackway from Tanzania. Silhouette modified from photograph (2019).
Track 3 is the largest and preserved in the least degree of detail. The track is larger and asymmetrical, suggesting a large artiodactyl with a complex foot structure.
Although more specific track maker associations cannot be constructed based on current evidence, the fossil record at Olduvai Gorge supports our initial ichnotaxonomic interpretations. Artiodactyla is one of the most abundant mammal groups found in fossil deposits from Olduvai Gorge (Gentry & Gentry, Citation1978). Bovidae fossils are abundant in Bed I and II at Olduvai Gorge (Gentry & Gentry, Citation1978), where Bovinae, Hippotraginae, Alcelaphinae, Antilopinae, and Caprinae are well represented.
Palaeoecological implications
Tracks of large herbivores provide direct evidence that essential resources occurred at Olduvai Gorge during the mid-Pleistocene. Today, the nearby Serengeti Plains are a well-established migratory pathway for wildebeest, zebra, antelope, and gazelle. These animals follow the rains in order to access nutritious grazing and better quality water sources. Many African species of Bovidae require open grasslands or shrub grasslands to survive. The presence of tracks attributed to these taxa imply that grassland and an adequate supply of freshwater occurred (at least seasonally) at Olduvai Gorge during the early to mid-Pleistocene. This agrees with what is known about early hominin site choice. Hominin archaeological sites and freshwater springs and lakes are closely associated with the rift valley axis and fault zone, where groundwater springs remain available even when there is a seasonal scarcity of surface water (Cuthbert et al., Citation2017).
Future research directions
Further investigation of the Olduvai Gorge track site is required to identify the precise trackway surface within Tuff IF and determine if it is continuous or if multiple tracked surfaces exist. Such surfaces record a snapshot in time and allow rare, direct observation of ancient inter- and intra-species interaction and behaviour. The discovery of trackways (instead of isolated tracks) would allow additional research into the locomotion style and speed of track makers at Olduvai (Falkingham, Citation2014; Lockley, Citation1998). Tracks and skeletal fossils are rarely found in the same strata, and the discovery of additional tracks and trackways would further our palaeoecological understanding of Olduvai Gorge during the early- to mid-Pleistocene.
The discovery of well-preserved fossil tracks has important implications for palaeoanthropological research at Olduvai Gorge. Habitat diversity and crucial resources (i.e., raw materials and freshwater) attracted a variety of hominin species to Olduvai Gorge during the late Pliocene and throughout the Pleistocene. This new discovery highlights the potential to discover faunal and hominin tracks or trackways from well-dated tuff deposits that have direct stratigraphic association with well-described fossil and tool assemblages from Olduvai Gorge.
Conclusions
Tracks belonging to the ichnogenera Pecoripeda and Megapecoripeda were discovered in volcanic ash Tuff IF, upper Bed I at Olduvai Gorge. Track morphology suggests track-makers were medium to large-bodied members of Artiodactyla (c.f., Bovidae). One track retains an exceptional level of detail and demonstrates the optimal trackway preservation potential of Tuff IF. The individual tracks show variable levels of detail, indicating that substrate consistency varied in time and space, or perhaps that tracks were made on multiple surfaces. Further investigation is required to identify the precise trackway surface(s) within Tuff IF. The preservation of these three tracks indicates there is the potential for future discovery of in situ fauna and hominin tracks or trackways in tuff deposits at Olduvai Gorge.
Acknowledgements
We express our deepest thanks to the Tanzania Antiquities Department, Ngorongoro Conservation Area Authority (NCAA), and The Olduvai Paleoanthropology and Paleoecology Project (TOPPP) for supplying permits and fieldwork support for our research. We also extend thanks to L. Stancampiano for offering geological field insights amid track impression excavations, and A. G. Plint for editorial advice.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Allen, J. R. L. (1997). Subfossil mammalian tracks (Flandrian) in the Severn Estuary, SW Britain: Mechanics of formation, preservation and distribution. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences , 352 (1352), 481–518.
- Altamura, F. , Bennett, M. R. , D’Août, K. , Gaudzinski-Windheuser, S. , Melis, R. T. , Reynolds, S. C. , & Mussi, M. (2018). Archaeology and ichnology at Gombore II-2, Melka Kunture, Ethiopia: Everyday life of a mixed-age hominin group 700,000 years ago. Scientific Reports , 8 (1), 2815.
- Archaeology News Network . (2016, June 15). Earliest footprints of Homo Erectus found in Eritrea. Archaeology News Network. Retrieved August 28, 2020, from <https://archaeologynewsnetwork.blogspot.com/2016/06/earliest-footprints-of-homo-erectus.html>.
- Ashley, G. M. (2003). Tracks, trails and trampling by large vertebrates in a rift valley paleo–wetland, lowermost Bed II, Olduvai Gorge, Tanzania. Ichnos , 9 (1-2), 23–32.
- Ashley, G. M. , & Hay, R. L. (2002). Sedimentation patterns in a Plio–Pleistocene volcaniclastic rift–platform basin, Olduvai Gorge. Tanzania. Special Publication SEPM , 73 , 107–122.
- Ashley, G. M. , Tactikos, J. C. , & Owen, R. B. (2009). Hominin use of springs and wetlands: Paleoclimate and archaeological records from Olduvai Gorge (∼1.79–1.74 Ma). Palaeogeography, Palaeoclimatology, Palaeoecology , 272 (1-2), 1–6.
- Balashova, A. , Mattsson, H. B. , Hirt, A. M. , & Almqvist, B. S. (2016). The Lake Natron Footprint Tuff (northern Tanzania): volcanic source, depositional processes and age constraints from field relations. Journal of Quaternary Science , 31 (5), 526–537.
- Behrensmeyer, A. K. (1978). Taphonomic and ecologic information from bone weathering. Paleobiology , 4 (2), 150–162.
- Behrensmeyer, A. K. , & Laporte, L. F. (1981). Footprints of a Pleistocene hominid in northern Kenya. Nature , 289 (5794), 167–169.
- Bennett, M. R. , Harris, J. W. , Richmond, B. G. , Braun, D. R. , Mbua, E. , Kiura, P. , Olago, D. , Kibunjia, M. , Omuombo, C. , Behrensmeyer, A. K. , & Huddart, D. (2009). Early hominin foot morphology based on 1.5-million-year-old footprints from Ileret. Kenya. Science , 323 , 1197–1201.
- Bennett, M. R. , Morse, S. A. , & Falkingham, P. L. (2014). Tracks made by swimming Hippopotami: An example from Koobi Fora (Turkana Basin, Kenya). Palaeogeography, Palaeoclimatology, Palaeoecology , 409 , 9–23.
- Berger, L. R. , & Hilton-Barber, B. (2000). In the footsteps of Eve: The mystery of human origins . National Geographic Adventure Press.
- Bibi, F. , Pante, M. , Souron, A. , Stewart, K. , Varela, S. , Werdelin, L. , Boisserie, J. R. , Fortelius, M. , Hlusko, L. , Njau, J. , & de la Torre, I. (2018). Paleoecology of the Serengeti during the Oldowan–Acheulean transition at Olduvai Gorge, Tanzania: The mammal and fish evidence. Journal of Human Evolution , 120 , 48–75.
- Bromley, R. G. , Milan, J. , Uchman, A. , & Hansen, K. S. (2009). Rheotactic Macaronichnus, and human and cattle trackways in Holocene beachrock, Greece: Reconstruction of paleoshoreline orientation. Ichnos , 16 (1-2), 103–117.
- Brown, F. H. , Haileab, B. , & McDougall, I. (2006). Sequence of tuffs between the KBS Tuff and the Chari Tuff in the Turkana Basin, Kenya and Ethiopia. Journal of the Geological Society , 163 (1), 185–204.
- Clarke, R. (2012). A Homo habilis maxilla and other newly–discovered hominid fossils from Olduvai Gorge, Tanzania. Journal of Human Evolution , 63 (2), 418–428.
- Cuthbert, M. O. , Gleeson, T. , Reynolds, S. C. , Bennett, M. R. , Newton, A. C. , McCormack, C. J. , & Ashley, G. M. (2017). Modelling the role of groundwater hydro-refugia in East African hominin evolution and dispersal. Nature Communications , 8 (1), 1–11.
- Day, M. H. , & Wickens, E. H. (1980). Laetoli Pliocene hominid footprints and bipedalism. Nature , 286 (5771), 385–387.
- Deino, A. L. (2012). 40Ar/39Ar dating of Bed I, Olduvai Gorge, Tanzania, and the chronology of early Pleistocene climate change. Journal of Human Evolution , 63 (2), 251–273.
- DeMenocal, P. B. (1995). Plio–Pleistocene African climate. Science , 270 (5233), 53–59.
- DeMenocal, P. B. (2004). African climate change and faunal evolution during the Pliocene–Pleistocene. Earth and Planetary Science Letters , 220 (1-2), 3–24.
- Diez-Martín F. , Yustos, P. S. , Uribelarrea, D. , Baquedano, E. , Mark D. F. , Mabulla, A. , Fraile, C. , Duque, J. , Díaz, I. , Pérez–González, A. , & Yravedra, J. (2015). The origin of the Acheulean: The 1.7 million–year–old site of FLK West, Olduvai Gorge (Tanzania). Scientific Reports , 5 , 17839.
- Domínguez-Rodrigo, M. , Pickering, T. R. , Baquedano, E. , Mabulla, A. , Mark, D. F. , Musiba, C. , Bunn, H. T. , Uribelarrea, D. , Smith, V. , Diez–Martin, F. , & Pérez–González, A. (2013). First partial skeleton of a 1.34 million year old Paranthropus boisei from Bed II, Olduvai Gorge, Tanzania. Plos One , 8 (12), e80347.
- Egeland, C. P. (2007). Zooarchaeological and taphonomic perspectives on hominid and carnivore interactions at Olduvai Gorge, Tanzania [PhD dissertation, Indiana University]. Indiana University Theses and Dissertations. http://hdl.handle.net/2022/7566.
- Falkingham, P. L. (2014). Interpreting ecology and behaviour from the vertebrate fossil track record. Journal of Zoology , 292 (4), 222–228.
- Fernández-Jalvo, Y. , Denys, C. , Andrews, P. , Williams, T. , Dauphin, Y. , & Humphrey, L. (1998). Taphonomy and palaeoecology of Olduvai Bed–I (Pleistocene, Tanzania. Journal of Human Evolution , 34 (2), 137–172. ).
- Gentry, A. W. , & Gentry, A. (1978). Fossil Bovidae (Mammalia) of Olduvai Gorge, Tanzania. Part I. Bulletin of the British Museum (Natural History), Geology Series , 30 , 1–83.
- Hágen, A. , Horváth, D. , & Stromp, M. (2014). Dynamics of the terrestrial mammals of Ipolytarnóc (Northern Hungary). Ichnos , 21 (3), 205–210.
- Hatala, K. G. , Harcourt-Smith, W. E. H. , Gordon, A. D. , Zimmer, B. W. , Richmond, B. G. , Pobiner, B. L. , Green, D. J. , Metallo, A. , Rossi, V. , & Liutkus-Pierce, C. M. (2020). Snapshots of human anatomy, locomotion, and behavior from Late Pleistocene footprints at Engare Sero. Scientific Reports , 10 (1), 7740.
- Hatala, K. G. , Roach, N. T. , Ostrofsky, K. R. , Wunderlich, R. E. , Dingwall, H. L. , Villmoare, B. A. , Green, D. J. , Braun, D. R. , Harris, J. W. K. , Behrensmeyer, A. K. , and Richmond, B. G. (2017). Hominin track assemblages from Okote Member deposits near Ileret, Kenya, and their implications for understanding fossil hominin paleobiology at 1.5 Ma. Journal of Human Evolution , 112 , 93–104.
- Hay, R. L. (1963). Stratigraphy of Beds I through IV, Olduvai Gorge, Tanganyika. Science , 139 (3557), 829–833.
- Hay, R. L. (1976). Geology of the Olduvai Gorge: A study of sedimentation in a semiarid basin. Science , 193 , 1234.
- Hay, R. L. , & Kyser, T. K. (2001). Chemical sedimentology and paleoenvironmental history of Lake Olduvai, a Pliocene lake in northern Tanzania. Geological Society of America Bulletin , 113 (12), 1505–1521.
- Helm, C. W. , Anderson, R. J. , Buckley, L. G. , Cawthra, H. C. , & de Vynck, J. C. (2017). Biofilm assists recognition of avian trackways in Late Pleistocene coastal aeolianites. South Africa. Palaeontologia Africana , 52 , 78–84.
- Helm, C. W. , Cawthra, H. C. , Combrink, X. , Helm, C. J. , Rust, R. , Stear, W. , & Heever, A. V. D. (2020). Pleistocene large reptile tracks and probable swim traces on South Africa’s Cape south coast. South African Journal of Science , 116 (3/4), 1–8.
- Helm, C. W. , Cawthra, H. C. , Cowling, R. M. , De Vynck, J. C. , Lockley, M. G. , Marean, C. W. , Thesen, G. H. , & Venter, J. A. (2020). Pleistocene vertebrate tracksites on the Cape south coast of South Africa and their potential palaeoecological implications. Quaternary Science Reviews , 235 , 105857.
- Helm, C. W. , Cawthra, H. C. , de Vynck, J. C. , Lockley, M. G. , McCrea, R. T. , & Venter, J. (2019). The Pleistocene fauna of the Cape south coast revealed through ichnology at two localities. South African Journal of Science , 115 (1/2), 1–9.
- Helm, C. W. , Cawthra, H. C. , Hattingh, R. , Hattingh, S. , McCrea, R. T. , & Thesen, G. H. (2019). Pleistocene vertebrate trace fossils of Robberg Nature Reserve. Palaeontologia Africana , 54 , 36–47.
- Helm, C. W. , McCrea, R. T. , Cawthra, H. C. , Lockley, M. G. , Cowling, R. M. , Marean, C. W. , Thesen, G. H. , Pigeon, T. S. , & Hattingh, S. (2018). A new Pleistocene hominin tracksite from the Cape south coast. Scientific Reports , 8 (1), 3772.
- Helm, C. , Cawthra, H. , Cowling, R. , De Vynck, J. , Marean, C. , McCrea, R. , & Rust, R. (2018). Palaeoecology of giraffe tracks in Late Pleistocene aeolianites on the Cape south coast. South African Journal of Science , 114 (1/2), 1–8.
- Jacobs, Z. , & Roberts, D. L. (2009). Last Interglacial Age for aeolian and marine deposits and the Nahoon fossil human footprints, Southeast Coast of South Africa. Quaternary Geochronology , 4 (2), 160–169.
- Kimura, Y. (2002). Examining time trends in the Oldowan technology at Beds I and II, Olduvai Gorge. Journal of Human Evolution , 43 (3), 291–321.
- Kordos, L. (1985). Footprints in the Lower Miocene sandstone at Ipolytarnóc. Geologica Hungarica, Series Paleontologica , 46 , 257–415.
- Kovarovic, K. , Slepkov, R. , & McNulty, K. P. (2013). Ecological continuity between Lower and Upper Bed II, Olduvai Gorge, Tanzania. Journal of Human Evolution , 64 (6), 538–555.
- Leakey, M. D. (1971). Olduvai Gorge: Volume 3, excavations in beds I and II, 1960–1963 . Cambridge University Press.
- Leakey, M. D. (1978a). Olduvai Gorge 1911–75: A history of the investigations. Geological Society, London, Special Publications, 6 (1), 151–155.
- Leakey, M. D. (1978b). Pliocene footprints at Laetolil, Northern Tanzania. Antiquity , 52 (205), 133.
- Liebenberg, L. (1990). A field guide to the animal tracks of southern Africa . David Philip Publishers.
- Liutkus-Pierce, C. M. , Zimmer, B. W. , Carmichael, S. K. , McIntosh, W. , Deino, A. , Hewitt, S. M. , McGinnis, K. J. , Hartney, T. , Brett, J. , Mana, S. , & Deocampo, D. (2016). Radioisotopic age, formation, and preservation of Late Pleistocene human footprints at Engare Sero, Tanzania. Palaeogeography, Palaeoclimatology, Palaeoecology , 463 , 68–82.
- Lockley, M. G. (1998). The vertebrate track record. Nature , 396 (6710), 429–432.
- Lucas, S. G. , & Hunt, A. (2007). Ichnotaxonomy of camel footprints. In Cenozoic Vertebrate Tracks and Traces (Bulletin 42, pp. 155–168). New Mexico Museum of Natural History and Science.
- Magill, C. R. , Ashley, G. M. , & Freeman, K. H. (2013). Ecosystem variability and early human habitats in eastern Africa. Proceedings of the National Academy of Sciences , 110, 1167–1174.
- Magill, C. R. , Ashley, G. M. , Domínguez-Rodrigo, M. , & Freeman, K. H. (2016). Dietary options and behavior suggested by plant biomarker evidence in an early human habitat. Proceedings of the National Academy of Sciences , 113, 2874–2879.
- McHenry, L. J. (2012). A revised stratigraphic framework for Olduvai Gorge Bed I based on tuff geochemistry. Journal of Human Evolution , 63 (2), 284–299.
- Melchor, R. N. (2015). Application of vertebrate trace fossils to palaeoenvironmental analysis. Palaeogeography, Palaeoclimatology, Palaeoecology , 439 , 79–96.
- Modern vertebrate tracks from Lake Manyara, Tanzania and their paleobiological implications. Paleobiology , 19 (4), 433–458. ‒
- Morgan, G. S. , & Williamson, T. E. (2000). Middle Miocene (Late Barstovian) vertebrates from the Benavidez Ranch local fauna, Albuquerque Basin, New Mexico. New Mexico Museum of Natural History and Science, Bulletin , 16 , 195–207.
- Morse, S. A. , Bennett, M. R. , Liutkus‐Pierce, C. , Thackeray, F. , McClymont, J. , Savage, R. , & Crompton, R. H. (2013). Holocene footprints in Namibia: The influence of substrate on footprint variability. American Journal of Physical Anthropology , 151 (2), 265–279.
- Raichlen, D. A. , Gordon, A. D. , Harcourt-Smith, W. E. H. , Foster, A. D. , & Haas, W. R. Jr (2010). Laetoli footprints preserve earliest direct evidence of human–like bipedal biomechanics. PLOS One. , 5 (3), e9769.
- Reck, H. (1914). Erste vorläufige Mitteilung über den Fund eines fossilen Menschenskelets aus zentral Afrika. Gesellschaft Naturforschender Freunde zu Berlin , 3 , 81–95.
- Roach, N. T. , Du, A. , Hatala, K. G. , Ostrofsky, K. R. , Reeves, J. S. , Braun, D. R. , Harris, J. W. , Behrensmeyer, A. K. , & Richmond, B. G. (2018). Pleistocene animal communities of a 1.5 million–year–old lake margin grassland and their relationship to Homo erectus paleoecology. Journal of Human Evolution , 122 , 70–83.
- Roach, N. T. , Hatala, K. G. , Ostrofsky, K. R. , Villmoare, B. , Reeves, J. S. , Du, A. , Braun, D. R. , Harris, J. W. Behrensmeyer , & Richmond, B. G. (2016). Pleistocene footprints show intensive use of lake margin habitats by Homo erectus groups. Scientific Reports , 6 (1)
- Roberts, D. L. (2008). Last interglacial hominid and associated vertebrate fossil trackways in coastal eolianites, South Africa. Ichnos , 15 , 190–207.
- Roberts, D. L. , Bateman, M. D. , Murray-Wallace, C. V. , Carr, A. S. , & Holmes, P. J. (2008). Last interglacial fossil elephant trackways dated by OSL/AAR in coastal aeolianite, Still Bay, South Africa. Palaeogeography, Palaeoclimatology, Palaeoecology , 257 (3), 261–279.
- Roberts, D. , & Berger, L. R. (1997). Last interglacial (c. 117 kyr) human footprints from South Africa. South African Journal of Science , 93 , 349–350.
- Sarjeant, W. A. S. , & Langston, W. Jr. (1994). Vertebrate footprints and invertebrate traces from the Chadronian (Late Eocene) of Trans-Pecos Texas. The Bulletin of the Texas Memorial Museum. Bulletin , 36 , 1–86.
- Scott, J. J. , Renaut, R. W. , & Owen, R. B. (2008). Preservation and paleoenvironmental significance of a footprinted surface on the Sandai Plain, Lake Bogoria, Kenya Rift Valley. Ichnos , 15: 208–231
- Shackleton, N. J. (1995). New data on the evolution of Pliocene climatic variability. Paleoclimate and evolution with emphasis on human origins . Yale University Press.
- Uno, K. T. , Rivals, F. , Bibi, F. , Pante, M. , Njau, J. , & de la Torre, I. (2018). Large mammal diets and paleoecology across the Oldowan–Acheulean transition at Olduvai Gorge, Tanzania from stable isotope and tooth wear analyses. Journal of Human Evolution , 120 , 76–91.
- Vyalov, O. S. (1965). Stratigrafiya Neogenovykh molass Predkarpatskovo progriba. (Stratigraphy of the Neogene molasse of the Pre-Carpathian Basin). Akademiya Nauk Ukrainskoy SSR Institut Geologii I Geokhimii Goryuchikh Inskopayemykh: 165.
- Vyalov, O. S. (1966). Sledy zhiznedeyatelnosti organizmov i ikh paleontologicheskoe znacheniye (Traces of living organisms and their paleontological significance). Akademiya Nauk Ukrainskoy SSR Institut Geologii I Geokhimii Goryuchikh Inskopayemykh: 219.
- Zimmer, B. , Liutkus-Pierce, C. , Marshall, S. T. , Hatala, K. G. , Metallo, A. , & Rossi, V. (2018). Using differential structure-from-motion photogrammetry to quantify erosion at the Engare Sero footprint site, Tanzania. Quaternary Science Reviews , 198 , 226–241.