
Abstract
Molecular targets of tyrosine kinase inhibitors are not restricted to the B-cell compartment but also regulate functions in the tumor microenvironment. Increasing evidence suggests that B-cell receptor-associated kinases like protein kinase C (PKC)-β is essential for the formation of a microenvironment supporting leukemic growth. Here we describe the effect of Idelalisib on the PKCβ/NF-κB and Notch pathway in stromal cells upon contact to primary chronic lymphocytic leukemia cells (CLL). There is no Idelalisib-dependent regulation of the Notch expression in stromal cells, whereas Idelalisib induces PKCβ expression and activates the canonical NF-κB pathway. Idelalisib deregulates important immune-modulatory proteins in activated stromal cells, which might provoke the patient’s side effects. Additionally, we established a 3D-stroma/leukemia model, that can give us a more defined look into the communication between tumor and stromal cells than standard cell cultures. This opens up the possibility to improve therapies, especially in the context of minimal-residual disease.
Keywords:
Introduction
The circular contact with the bone marrow is one of the hallmarks of chronic lymphocytic leukemia (CLL) [Citation1]. The interaction between the microenvironment and malignant B-cell is a dynamic process and provides protection from spontaneous apoptosis [Citation2,Citation3]. The expression of PKCβ and the subsequent activation of NF-κB in bone marrow stromal cells (BMSCs) are prerequisites to support the survival of malignant B-cells [Citation4]. Moreover, the resistance of CLL cells toward drug-induced apoptosis is also mediated by stromal contact, clinically recognized as a minimal-residual disease (MRD) [Citation5]. Recently, in vivo experiments demonstrated that the inhibition of PKCβ in stromal cells enhances chemosensitivity in CLL and overcomes drug resistance [Citation6]. It could be shown that stromal contact activates Notch signaling [Citation7] indicating that Notch might play a distinct role in the communication between CLL cells and the microenvironment. Notably, stromal Notch2-dependent mechanisms control non-autonomous WNT-signaling in CLL cells. Pharmacological inhibition of the WNT pathway impairs stromal-mediated survival of the tumor cells [Citation8].
Tyrosine kinase inhibitors (TKIs), targeting signals downstream B-cell receptor (BCR) have rapidly advanced clinical trials in patients with CLL. Their molecular targets are not restricted to the B-cell compartment but regulate key functions in the microenvironment [Citation9,Citation10]. Idelalisib, an inhibitor of the phosphatidylinositol-3-kinase p110δ subunit (PI3Kδ), is approved for treating relapsed/refractory chronic lymphocytic leukemia, follicular lymphoma and small lymphocytic lymphoma [Citation11]. The drug is associated with hematopoietic and pulmonary toxicities and can lead to CMV reactivations in patients, which limit its clinical use. However, the toxicity mechanisms are not completely elucidated [Citation10,Citation12]. Malignant B-cells modify infiltrating tissues and manipulate surrounding cells to generate a protective microenvironment. Moreover, they modify cells devoted to natural and adaptive immune response, inducing an immunosuppressive behavior [Citation9,Citation13]. TKIs reduce the lymphoid mass very efficiently, but can obtain a complete response only in a small minority of cases [Citation9]. Analyzing the effects of TKIs on the microenvironment of B-cell malignancies is relevant to understand phenomena observed in the clinical setting and will help to define how protective niches are modified.
In the present study, we describe Idelalisib-induced effects on the PKCβ/NF-κB pathway and the Notch regulation in stromal cells of CLL. The effect of Idelalisib shows deregulation of important immune-modulatory proteins in stromal cells. Additionally, findings of inhibitor combination studies in a new 3D-stroma/leukemia model could be exploited to generate new approaches to therapeutically overcome stroma-mediated drug resistance.
Experimental procedures
Patient samples
After informed patients' consent and in accordance with the Helsinki declaration, peripheral blood was obtained from untreated, infectious-free patients with a diagnosis of CLL. Studies were approved by the Ethics Committee of the University of Erlangen-Nürnberg (number: 219_14B, addendum 59_17 Bc). PBMC isolation was performed as previously described [Citation4]. A list of patient samples used in each experiment is shown in Supplement Table.
Cell culture conditions
The culture conditions of CLL cells, EL08-1D2 and HS-5 cells (CRL-11882, ATCC) were previously described in detail [Citation4]. Purified CD19+ CLL cells were cocultured on EL08-1D2 or HS-5 cells for 5 d. CLL cells were physically removed from stromal cells by repeated washing resulting in minimal cross-contamination numbers. Inflow cytometry analysis CLL cells can easily be distinguished using a CD19+ lymphocyte gate. Stromal cells were further purified using anti-CD19 magnetic beads to eliminate any minimal CLL contamination, especially in qPCR analysis.
3D cell culture
The scaffolds were initially prepared as suggested by the manufacturer (Alvetex, Reprocell Europe, Durham, UK). EL08-1D2 cell suspension was added to the center of the scaffold. After 3 h the medium was applied. After 24 h 3 × 106 CLL cells/ml were added in their appropriate medium to the EL08-1D2 cells on the scaffold for 5 d. The CLL cells were harvested by collecting the entire medium. With two additional washing steps in medium, the CLL cells sitting on stromal cells on the outside of the scaffold were released by gently pipetting. To retrieve the cells out of the inner core of the scaffold, we unclip the inserts, remove the scaffold using flat-ended forceps and cut the scaffold into pieces. The small pieces were transferred in 0.25% Trypsin/EDTA and incubated for 15 min at 37 °C, 5% CO2 on a tube rolling mixer. This CLL cell isolation step from the inner core of the scaffolds was repeated twice. CLL cells can be separated from stromal cells of this region using a CD19+ lymphocyte gate in flow cytometry.
RT2 assays
After coculture, CLL and stromal cells were separated with anti-CD19 magnetic beads and RNA of stromal cells was extracted. Stromal cDNA was prepared (RT2 First Strand Kit, Qiagen) and subjected to pathway-focused gene expression analyses of 84 genes involved in cancer inflammation and immunity crosstalk (RT2 Profiler PCR Arrays, Qiagen, Hilden, Germany).
CRISPR/Cas9 plasmids
Single-guide RNA sequences (sgRNA) were cloned into lentiCRISPRv2-GFP according to Zhang et al. LentiCRISPRv2GFP was a gift from David Feldser [Citation14] (Addgene plasmid # 82,416; http://n2t.net/addgene:82,416; RRID:Addgene_82,416). Lentiviral infection of EL08-1D2 cells with two different Notch2-sgRNA sequences was performed (). Following the separation of GFP positive cells, EL08-1D2 cells were negatively sorted for Notch2 expression. The experiments were performed with EL08ΔNotch2 using sgRNA Notch2 #1.
Table 3. sgRNA sequences.
HS-5 stromal cells were infected with lentiviral sgRNA (Edit-R Human NOTCH2 hEF1a-EGFP All-in-one Set, Horizon Discovery, Cambridge, UK) with three different Notch2-sgRNA sequences (). Following the separation of GFP positive cells, HS-5 cells were negatively sorted for Notch2 expression. The experiments were performed with HS-5ΔNotch2 using sgRNA VSGH12180-249451941.
GM-CSF ELISA
After coculture of murine EL08-1D2 or human HS-5 cells with primary CLL cells, the supernatant was used for detection of GM-CSF in a mouse or human-specific enzyme-linked immunosorbent assay (R&D Systems, Minneapolis, USA) according to manufacturer’s protocol.
NF-κB assay
After coculture of EL08-1D2 and primary CLL cells for 5 d, nuclear-cytoplasmatic protein separation was performed of the stromal pellet (SP) using a Nuclear Extraction Kit (Abcam, Cambridge, UK). CLL cells were pretreated with the caspase inhibitor Z.vad.fmk (100 µM) for 30 min to suppress spontaneous apoptosis. The nuclear protein pellet of the stromal cells was analyzed with the colorimetric NF-κB Transcription Factor Assay Kit (Abcam, Cambridge, UK) following the manufacturer’s manual.
Immunofluorescence staining of 3D-stroma/leukemia cell culture
After 5 d of coculturing stromal cells with CLL cells on scaffolds in 8-well chamber slides (Ibidi, Gräfelfing, Germany), the medium was removed. 3D cell culture was washed twice with PBS and fixed by incubation within 4% paraformaldehyde for 15 min at room temperature. The 3D culture was permeabilized in 1% Triton-X100 diluted in PBS for 15 min at room temperature. Specific binding of primary antibody (CD19-antibody, ) and CellMask Green plasma membrane stain was achieved by incubating overnight at 4 °C. After washing, nuclei were counterstained with DAPI before the 3D cultures were mounted on glass slides within SlowFade antifade mountant medium (Invitrogen, Germany). Confocal imaging was performed on a Zeiss Spinning Disk Axio Z1 live-cell observer (Zeiss, Jena, Germany).
Z-stack data of confocal images were reconstructed using Fiji [Citation15] to obtain three-dimensional representations of the stromal cell distribution within the 3D-stroma/leukemia model.
Statistical analysis
All experiments were repeated at least three times. The sample sizes for each experiment were provided in the figure legends. Statistical analyses were performed by one-way ANOVA for multiple comparisons. Two-tailed Student t-tests with unpaired or paired analysis were performed based on the distribution levels using GraphPad Prism Version 8 (GraphPad Prism Software Inc., La Jolla, CA). Throughout the manuscript, statistical significance was defined as p < 0.0001 (****), p < 0.001 (***), p < 0.01 (**), p < 0.05 (*) or ns (statistically non-significant).
Results
Idelalisib reduces the expression of distinct proteins of the immunity crosstalk in activated stromal cells
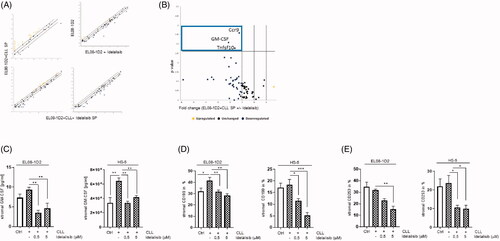
We previously established a coculture system to study heterotypic cell-cell interactions between stromal cells and primary CLL cells [Citation4]. The EL08-1D2 cells are primary stromal cells derived from mouse embryonic liver (E11), which has been carefully characterized as stromal cells that are able to support human hematopoietic stem cell activity [Citation16]. We analyzed the relative gene expression of proteins that are related to inflammation and immunity in stromal cells to gain insight into a possible transcriptional regulation of Idelalisib treatment. We found Idelalisib-induced changes in the expression level of stromal genes like granulocyte-macrophage colony-stimulating factor 2 (GM-CSF), tumor necrosis factor (ligand) superfamily member 10 (Tnfsf10/CD253) and chemokine receptor 9 (Ccr9/CD199). These genes were downregulated in EL08-1D2 stromal cells upon CLL contact due to incubation of Idelalisib (). To confirm the gene regulatory data, we detected the GM-CSF levels in the supernatant of EL08-1D2 and HS-5 monocultures (Ctrl) in comparison to cocultures with CLL cells using a mouse- and human-specific GM-CSF ELISA. Idelalisib reduced the GM-CSF protein expression in murine EL08-1D2 stromal cells as well as in human HS-5 stromal cells (). The same effect was assessed by cell surface expression of stromal CD199 and CD253 on EL08-1D2 and HS-5 cells using FACS analysis ().
Figure 1. Idelalisib induced changes in the immunity crosstalk in stromal cells after CLL contact. EL08-1D2 cells were cocultivated with CLL cells for 5 d. Idelalisib (5 µM) was added 24 h prior separation of the cell types. The stromal cells were subjected to gene expression analyses focusing on the stromal immunity crosstalk (RT2Profiler PCR Array, Qiagen). (A) Scatter Blots from EL08-1D2 cells in monoculture ± Idelalisib and EL08-1D2 cells after coculture with CLL cells (SP) for 5 d with CLL cells from 3 individual patients ± Idelalisib were shown (n = 3). (B) The Volcano plot was calculated using the data derived from the scatter plots (https://geneglobe.qiagen.com/de/my-geneglobe/analysis). It displayed Idelalisib induced differentially expressed genes in EL08-1D2 stromal cells upon CLL contact (n = 3). Significant downregulated genes like C–C Motif Chemokine Receptor 9 (Ccr9/CD199), Tumor Necrosis Factor Ligand Superfamily Member 10 (Tnfsf10/Trail/CD253) and Colony Stimulating Factor 2 (Csf2/GM-CSF) were found within the upper left quadrant (blue box). (C) Concentration of the stromal GM-CSF was assessed in the supernatant from EL08-1D2 and HS-5 stromal cells before and after CLL contact using a species-specific GM-CSF ELISA (n = 6). (D) Density of stromal CD199 and (E) CD253 was detected on EL08-1D2 and HS-5 cells upon CLL contact under baseline conditions and after Idelalisib treatment by FACS analysis (n = 6). Bars indicate the standard error of the mean. *p < 0.05; **p < 0.01; ***p < 0.001. Abbreviations: CLL: chronic lymphocytic leukemia cells; SP: stromal cell pellet after CLL contact; Ctrl: control CLL cells with stromal contact without treatment; p; p-value.
Notch signaling in stromal cells after CLL contact is not affected by Idelalisib treatment
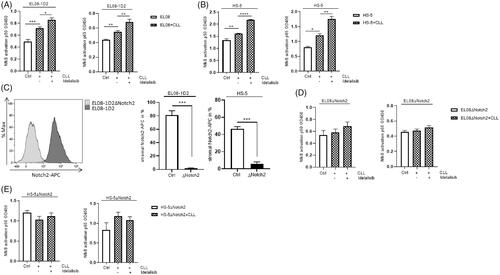
Since inhibition of the Notch pathway diminishes the survival of stroma-protected CLL cells in vitro and disease engraftment in vivo, we investigated the impact of Idelalisib on the stromal Notch pathway. First, we applied Idelalisib together with the Notch pathway inhibitor DAPT, which blocks the function of the γ-secretase, on EL08-1D2/CLL and HS-5/CLL cocultures and detected the CLL survival rate using Annexin-V/PI staining. Interestingly, the combination of Idelalisib and DAPT did not induce a higher apoptosis rate in CLL cells compared to the single agents (). Additionally, we analyzed the gene expression of the Notch receptors in stromal cells in monoculture (white column) and after CLL contact (filled column) using qPCR studies (, ). Notch3 and Notch4 were not regulated after Idelalisib or DAPT treatment (Supplement Figure 1(A,B)). In line with Notch protein expression analysis published by Mangolini et al. [Citation8], we detected a decrease in Notch1 and a clear increase in Notch2 gene expression in EL08-1D2 cells after CLL contact (). Moreover, CLL contact-induced an upregulation of the transcriptional factor Hes1 in EL08-1D2 and HS-5 stromal cells indicating activation of functional Notch target genes ( right panel). However, Idelalisib had no impact on the gene regulation of Notch receptors and Hes1 in stromal cells after CLL contact. This is in line with the protein expression of the Notch receptors on the cell surface of EL08-1D2 cells after coculture with CLL cells (Supplement Figure 1(C)). In agreement with a previous report, we could show that Notch1 and Notch2 had by far the highest surface expression in EL08-1D2 cells (Supplement Figure 1C) [Citation8]. This indicated that Idelalisib has no impact on the Notch pathway in stromal cells during CLL contact, also in lower concentrations (Supplement Figure 2).
Figure 2. Notch signaling in stromal cells after CLL contact is not affected by Idelalisib treatment. (A) Apoptotic CLL cells were cultured on EL08-1D2 and HS-5 stromal cells under baseline conditions or after addition of Idelalisib (5 µM, 24 h) or DAPT (10 µM, 4 d) or combination of both. CLL cells were assessed by Annexin-V/PI staining (n = 7). (B) EL08-1D2 and (C) HS-5 cells were cocultivated with CLL cells for 5 d under basal conditions. Idelalisib (5 µM, 24 h) or DAPT (10 µM, 4 d) or combination of both was added. After separation changes in the relative gene expression of the stromal Notch receptors Notch1, Notch2 and the transcription factor Hes1 were analyzed by qPCR in stromal cells (n = 6) (grey columns). The results were normalized to monocultured EL08-1D2 or HS-5 cells as control (white column). Bars indicate the standard error of the mean. ns (not significant); *p < 0.05; **p < 0.01; ****p < 0.0001. Abbreviations: CLL: chronic lymphocytic leukemia cells; DAPT: N-[N-(3,5-Difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester; p: p-value.
![Figure 2. Notch signaling in stromal cells after CLL contact is not affected by Idelalisib treatment. (A) Apoptotic CLL cells were cultured on EL08-1D2 and HS-5 stromal cells under baseline conditions or after addition of Idelalisib (5 µM, 24 h) or DAPT (10 µM, 4 d) or combination of both. CLL cells were assessed by Annexin-V/PI staining (n = 7). (B) EL08-1D2 and (C) HS-5 cells were cocultivated with CLL cells for 5 d under basal conditions. Idelalisib (5 µM, 24 h) or DAPT (10 µM, 4 d) or combination of both was added. After separation changes in the relative gene expression of the stromal Notch receptors Notch1, Notch2 and the transcription factor Hes1 were analyzed by qPCR in stromal cells (n = 6) (grey columns). The results were normalized to monocultured EL08-1D2 or HS-5 cells as control (white column). Bars indicate the standard error of the mean. ns (not significant); *p < 0.05; **p < 0.01; ****p < 0.0001. Abbreviations: CLL: chronic lymphocytic leukemia cells; DAPT: N-[N-(3,5-Difluorophenacetyl)-L-alanyl]-S-phenylglycine t-butyl ester; p: p-value.](/cms/asset/b635fc33-8c4b-476b-96e9-225902fd3b99/ilal_a_1927019_f0002_c.jpg)
PKCβ inhibitor enzastaurin support the Idelalisib-induced CLL cell death despite stromal contact
Direct contact of CLL cells activates stromal cells by inducing PKCβ expression [Citation4]. To assess to what extend Idelalisib manipulates the PKCβ regulation in the stromal compartment of CLL, we first tested if Idelalisib has an effect on the expression of PKCβ in EL08-1D2 and HS-5 stromal cells. The stromal cells were analyzed for PKCβ protein expression after 5 d of coculture using an intracellular FACS staining. As expected, CLL contact-induced a PKCβ expression in stromal cells. With the addition of Idelalisib in increasing concentrations, the PKCβ expression was further increased in stromal cells (, left panel). Similar to murine EL08-1D2 stromal cells, the PKCβ expression in human HS-5 stromal cells was elevated after Idelalisib treatment during CLL contact (, right panel).
Figure 3. PKCβ inhibitor Enzastaurin support the Idelalisib-induced CLL cell death despite stromal contact. (A) Stromal PKCβ expression was assessed in EL08-1D2 cells (left panel) (n = 9) and HS-5 cells (right panel) (n = 8) before (Ctrl; white column) and after CLL contact (grey column) under baseline conditions and after addition of Idelalisib (0,5µM, 5 µM, 24 h) by FACS analysis. (B) CLL cells were cultured in medium as control (Ctrl) and were exposed to Idelalisib (5 µM) alone or in combination with Sotrastaurin (5 µM) or Enzastaurin (5 µM) for 24 h. Apoptotic CLL cells were stained with Annexin-V/PI (n = 6). (C) CLL cells were cultured in contact to EL08-1D2 and HS-5 stromal cells for 5 d (Ctrl). The cocultures were treated with Idelalisib (5 µM) or in combination with Sotrastaurin (5 µM) or Enzastaurin (5 µM) for 24 h. Annexin-V/PI staining detected CLL viability by FACS analysis (n = 6). (D) Stromal PKCβ expression was assessed in EL08-1D2 (left panel) and HS-5 cells (right panel) after CLL contact under baseline conditions and after addition of Sotrastaurin (5 µM) or Enzastaurin (5 µM) for 24 h by FACS analysis (n = 8). Bars indicate the standard error of the mean *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. Abbreviations: CLL: chronic lymphocytic leukemia cells; Ctrl: control CLL cells with stromal contact without treatment; APC: Allophycocyanin; p: p-value.
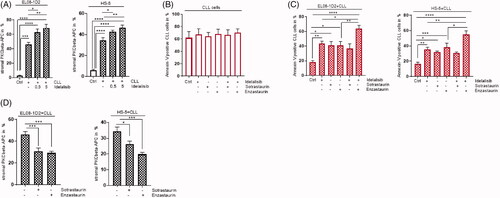
Next, we tested if Idelalisib could overcome the protective effect of stromal cells on CLL cells. We analyzed apoptosis using Annexin-V/PI staining in primary CLL cells cultured as monoculture (Ctrl) for 24 h to detect the direct effect of Idelalisib avoiding cell death because of missing stromal contact (). Stromal cells were in contact with CLL cells for 5 d. The coculture was exposed to Idelalisib for 24 h before assessing CLL survival. As expected CLL cells cultured on EL08-1D2 or HS-5 cells had a clear survival benefit compared to suspension cells in monoculture (; Ctrl). Notably, CLL cells were not protected from the cytotoxic effect of Idelalisib despite stromal contact (). Furthermore, the PKCβ inhibitor Enzastaurin or pan-PKC inhibitor Sotrastaurin was applied separately or in combination to Idelalisib for 24 h on CLL cells in monoculture () and cocultures (). Both compounds have the ability to inhibit the PKCβ expression in stromal cells upon CLL contact (). Interestingly, only the PKCβ-specific inhibitor Enzastaurin in combination with Idelalisib showed a significantly higher apoptotic CLL rate on protective EL08-1D2 and HS-5 stromal cells (). The enhanced PKCβ expression seems not to be relevant for CLL survival during Idelalisib treatment. However, PKCβ inhibition using Enzastaurin supported the Idelalisib-induced CLL cell death despite stromal contact.
The Idelalisib-induced NF-κB activation in stromal cells in response to CLL contact is Notch2 dependent
It is known that stromal PKCβ activates NF-ĸB through the canonical pathway, which is indispensable for CLL survival [Citation4]. Due to the fact that we detected an Idelalisib-dependent PKCβ induction in stromal cells (), we assessed downstream NF-κB activation in stromal cells. After 5 d of coculture with and without Idelalisib treatment for 24 h, the nuclear fractions from stromal cells were isolated and analyzed with an ELISA-based NF-κB Transcription Factor Activation Assay. CLL contact clearly induced NF-κB p50 and p65 subunits in stromal cells, which was further enhanced by Idelalisib treatment (). This NF-κB activation was not detected in PKCβ knockout stromal cells upon CLL contact and Idelalisib treatment (data not shown). Numerous reports also described regulation of NF-κB by Notch and vice versa through different mechanisms [Citation17]. To investigate if Idelalisib can affect the NF-κB activation without regulating the Notch pathway, we generated Notch2 deleted EL08-1D2 (EL08ΔNotch2) and HS-5 cells (HS5ΔNotch2) using CRISPR/Cas9 technology (). Notably, the NF-κB activation of p50 and p65 in EL08ΔNotch2 and HS5ΔNotch2 cells was not altered due to CLL contact indicating that Notch2 signaling is required for NF-κB activation through the canonical pathway. Furthermore, Idelalisib treatment could not activate NF-κB in Notch2 deleted stromal cells supporting our data that the Notch pathway is not affected by Idelalisib ().
Figure 4. The Idelalisib-induced NF-κB activation in stromal cells in response to CLL contact is Notch2 dependent. (A) CLL cells were pretreated with the caspase inhibitor Z.vad.fmk (100 nM, 30 min) and cultured on EL08-1D2 and (B) HS-5 stromal cells for 5 d with and without Idelalisib treatment (5 µM, 24 h). After separation with CD19 beads the nuclear fractions of the stromal cells were isolated. The activation of the NF-κB transcription factor p50 and p65 in stromal was quantified using a colorimetric ELISA (OD450 nm). Stromal cells without CLL contact were designate as control (Ctrl; white column) (n = 6). (C) Notch2 was deleted in EL08-1D2 (EL08ΔNotch2) and HS-5 (HS-5ΔNotch2) cells using CRISPR/Cas9 technology. Exemplary histogram of Notch2 in EL08-1D2 cells (Ctrl) and EL08-1D2ΔNotch2 is shown (left panel). Flow cytometry of stromal Notch2 expression on the cell surface of EL08-1D2 and HS-5 cells ± CRISPR/Cas9 deletion of Notch2 was detected (n = 3). (D) NF-κB transcription factor p50 and p65 expression in EL08ΔNotch2 and (E) HS-5ΔNotch2 stromal cells was quantified using a colorimetric ELISA. Notch2-deleted stromal cells without CLL contact were designate as control (Ctrl; white column) (n = 6). Bars indicate the standard error of the mean. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. Abbreviations: CLL: chronic lymphocytic leukemia cells; Ctrl: control CLL cells with stromal contact without treatment; EL08ΔNotch2: Notch2 deleted EL08-1D2 cells via CRISPR/Cas9; APC: Allophycocyanin; p: p-value.
Idelalisib treatment combined with PKCβ inhibition can overcome stromal-mediated drug resistance in 3D cell culture
We generate a 3D cell culture system as a stroma-based leukemia model cultivating stromal cells on a scaffold together with malignant B-cells to mimic the in vivo situation. Numerous key findings obtained from 2D cell cultures can be validated in these 3D stroma niches to rule out optional treatment approaches for patients. Particularly, the effectiveness of inhibitors could be evaluated directly in 3D cell culture. Using this model, we aimed to analyze if inhibitor combinations with Idelalisib have the same effect on CLL survival and can overcome stromal-mediated protection as in standard 2D cell cultures. Tumor cells were exposed to different inhibitors alone (Supplement Figure 3(A,B)) and in combination with Idelalisib to assess their impact on viability in established cocultures. These experiments were also performed at lower doses of Idelalisib (Supplement Figure 4(A–E)). After adding Idelalisib in combination with Fludarabine (purine analog), ABT-199 (B cell lymphoma 2 (BCL2) inhibitor) or Ibrutinib (inhibitor of B cell receptor-induced kinase), CLL survival was investigated by Annexin-V/PI staining. As expected, the contact with EL08-1D2 or HS-5 stromal cells in 2D cell culture enhanced the survival of CLL cells in the medium control (Ctrl) () compared to CLL cells in monoculture (Supplement Figure 5(A,B,C)). Idelalisib treatment was able to overcome this protective stromal effect. Adding Idelalisib in combination with ABT-199 (), Fludarabine () and Ibrutinib (), an additive effect on CLL cell death that was comparable to the CLL cell death in monocultures (Supplement Figure 5(A,B,C)) was detected.
Figure 5. Idelalisib treatment combined with PKCβ inhibition can overcome stromal-mediated drug resistance in 3D cell culture. Apoptotic CLL cells were quantified by Annexin-V/PI staining after contact to EL08-1D2 and HS-5 cells (Ctrl) for 5 d with 24 h exposure to (A) Idelalisib (5 µM) ± ABT-199 (10 nM) (B) Idelalisib (5 µM) ± Fludarabine (40 µg/ml), (C) Idelalisib (5 µM) ± Ibrutinib (10 µM) in a standard 2D cell culture (n = 6). (D) Illustration of the scaffold-based 3D-model consisting of stromal cells and malignant B-cells with the core region and the peripheral regions. (E) CLL cell survival was measured using Annexin-V/PI staining after coculture in 3D cell culture conditions and 24 h inhibitor exposure to Idelalisib (5 µM) ± Enzastaurin (5 µM), ± ABT-199 (10 nM), ± Fludarabine (40 µg/ml) and ± Ibrutinib (10 µM). CLL cells were in direct contact to stromal cells (EL08-1D2, left panel; HS-5, right panel) and isolated from the peripheral regions of the scaffold (n = 6). (F) Using confocal laser microscopy, the distribution of the stromal cells in the different regions of the scaffold was determined. Serial sections EL08-1D2 (left panel) and HS-5 stromal cells (right panel) in coculture with CLL cells were stained for CD19 (red) and CellMask plasma membrane stain (green). Nuclei were stained with DAPI (blue) (n = 3). Scale bars: 100 μm. Z-stacks of stained 3D cultures were imaged by a confocal spinning disk microscope with an axial distance of 270 nm and reconstructed for 3D visualization. (G) CLL cell survival was measured using Annexin-V/PI staining after coculture in 3D cell culture conditions and 24 h inhibitor exposure to Idelalisib (5 µM) ± Enzastaurin (5 µM), ± ABT-199 (10 nM), ± Fludarabine (40 µg/ml) and ± Ibrutinib (10 µM). CLL cells were isolated after 5 d of coculture in direct contact to stromal cells (EL08-1D2, left panel; HS-5, right panel) from the core region of the scaffold (n = 6). Bars indicate the standard error of the mean. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001. Abbreviations: CLL: chronic lymphocytic leukemia cells; Ctrl: control CLL cells with stromal contact without treatment; p: p-value.
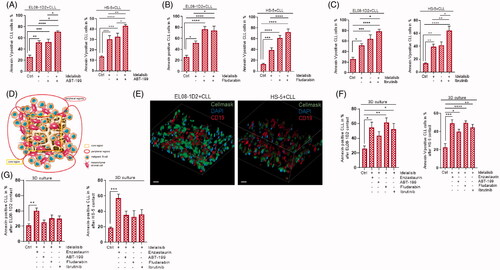
Interestingly, using the 3D leukemia model we can differentiate between two regions - the peripheral regions of the 3D stromal complex and the inside core region (). Importantly, stromal cells can migrate and grow in all regions of the scaffold (). Z-stack data of the confocal images were reconstructed using Fiji (3D script plugin) to obtain 3 D representations showing the distribution of the stromal cells upon CLL contact within the 3 D-stroma/leukemia model (Supplement Video 1, 2). We isolated the CLL cells that were in contact with stromal cells from the outside region of the scaffold and analyzed the CLL survival using Annexin-V/PI staining. As expected, the CLL cells profit from the stromal mediated protection (; Ctrl). The combination treatments of Idelalisib with Enzastaurin, Fludarabine or Ibrutinib were able to overcome stroma-mediated drug resistance resulting in increased CLL cell death (), which was comparable to the 2D situation ( 5(B,C)). The combination with ABT-199 exhibited a trend in elevated CLL cell death but was not as effective as in the 2D cell culture (). Notably, analysis of the CLL cells sitting in the inside core region of the 3D cell culture model, showed a different survival pattern. Only Idelalisib in combination with the PKCβ inhibitor Enzastaurin seems to be effective by diminishing the protective stromal effect on CLL cell survival in the more secure core region of the 3D cell culture ().
Discussion
TKIs can induce an immune-tolerant behavior in surrounding cells [Citation9]. Interestingly, we monitored Idelalisib-induced changes in the immunity crosstalk in stromal cells after CLL contact. Stromal GM-CSF, Ccr9 and Tnfsf10 expression were reduced due to Idelalisib treatment upon CLL contact. GM-CSF has a profound role in regulating the immune response and maintaining immunological tolerance [Citation18]. A lower level of GM-CSF could lead to lower activation of macrophages, a process crucial for fighting infections. This could be evidence for the patient’s side effects like pulmonary toxicity and susceptibility to infections, like mycosis. Ccr9 is involved in T-cell development in the thymus and in the gut-associated immune response, which might explain gastrointestinal symptoms in patients. Tnfsf10 activates the caspase 8-dependent apoptosis in tumor cells. Tnfsf10 receptor-targeted therapy has so far been disappointing in the clinic because only a small proportion of patients responded due to the development of resistance to Tnfsf10 [Citation19]. However, targeting the Tnfsf10 receptor in combinational therapies or adding GM-CSF to compensate immune-modulatory effects of Idelalisib to counterpart side effects might be helpful.
Stromal cells get activated by direct CLL contact leading to stromal PKCβ expression followed by NF-κB activation [Citation4]. Additionally, CLL cells induce Notch2 in stromal cells that in turn activates the canonical WNT pathway in CLL cells [Citation8]. The manipulation of both pathways in stromal cells leads to CLL survival demonstrating that CLL cells can change the function of surrounding cells. A recent study underlines the importance of PKCβ expression in the stromal compartment to protect malignant B-cells from cytotoxic therapies in vivo enrolling a PKCβ-dependent effect in environment-mediated drug resistance of CLL cells [Citation6]. The PI3K inhibitor Idelalisib demonstrated promising clinical efficiency in different B-cell malignancies [Citation9]. Here, we describe that Idelalisib can stimulate the PKCβ/NF-κB pathway in stromal cells after direct contact with CLL cells. Of note, the canonical NF-κB pathway was not activated in PKCβ-deficient stromal cells (data not shown). Interestingly, the effect of Idelalisib can overcome the stroma-mediated protection despite a secondary increased PKCβ/NF-κB expression in stromal cells.
Notably, the stromal Notch pathway seems not to be affected by Idelalisib. However, adding the PKCβ inhibitor Enzastaurin in combination with Idelalisib diminishes stroma protection and leads to CLL cell death.
Cultivating primary CLL cells in a 3D leukemia model, where they are able to get in contact with stromal cells in multiple dimensions facilitating more natural interactions, can help us to get a defined look into the communication of different cell types. 3D cultures can reveal dramatic differences in cellular responses compared to monolayer culture representing the in vivo situation even better [Citation20–23]. The combination of Idelalisib with Enzastaurin, Fludarabine or Ibrutinib can overcome stromal protection in the 3D constellation () comparable to the 2D cell culture. Interestingly, we could show that CLL cells sitting on stromal cells in the core region of the 3D cell culture model exhibit a higher drug resistance compared to CLL cells that were in direct contact with stromal cells located in the peripheral regions. Of note, stromal cells are evenly distributed in the scaffold (). CLL cells in the core region demonstrate a clear maintaining stromal protection visualized by low numbers of apoptotic cells despite ABT-199, Fludarabine or Ibrutinib treatment in combination with Idelalisib (). In contrast, Idelalisib in combination with Enzastaurin can overcome the protective effect in the core area. This indicates that, on the one hand, CLL cells can frequent specially protected areas of stromal cells to survive the cytotoxic action of several drugs, which might lead to MDR. On the other hand, PKCβ inhibition can overcome also stromal-mediated drug resistance in the more secure regions of the 3D cell culture. This symbolizes a clear difference to the 2D cell culture and might be an important point concerning the perspective of therapeutic efficiency and drug resistance of combination therapy (33). Moreover, this effect clearly shows that the drug diffusion into the whole high-porous scaffold is possible. Specifically, these results point to an essential role of the CLL cell distribution in a 3 D niche. 3D cell cultures and organoids have already been proven to be an excellent platform for drug screening and give the opportunity to mimic the complexity in signaling and tissue formation [Citation20–27].
In summary, our study provides new insight into the Idelalisib-mediated changes of the PKCβ and NF-κB activation in stromal cells upon CLL contact. Additionally, we detected deregulation of immune-modulatory proteins in the stromal cells after contact to CLL, which can be responsible for specific side effects. Furthermore, PKCβ inhibition can overcome the protective stromal effect on CLL survival in the core region of the 3D-stroma/leukemia model. The findings based on the 3D model point to an essential role of a more in vivo like the situation to improve therapies against the stromal microenvironment, especially in the context of MRD in patients.
Author contributions
S.H.: Collection and analysis of data, manuscript writing. F.v.H.: Collection and assembly of data. S.V.: Provision of patient material. R.A.J.O.: Provision of study material. J.W.: Provision of patient material. A.N.K.: interpretation of data and final approval of the manuscript. A.M.: Administrative support, final approval of manuscript and financial support. G.L-G.: Conception and design, financial support, interpretation of data and the manuscript writing.
Table 1. Antibody list.
Table 2: Primer.
Acknowledgements
The present work was performed in fulfillment of the requirements for obtaining the degree “Dr. med.” The project was supported by the Deutsche Forschungsgemeinschaft (DFG) – Project number LU2181/1-1 and by GILEAD (Förderprogramm Onkologie). We acknowledge the kind assistance of the University of Cambridge, UK for providing blood samples from patients (MTA). The CRISPR vector was designed by David Feldser. Cell sorting and immunofluorescence analysis has been performed at the Core Units “Cell Sorting and Immunomonitoring” and the “Optical Imaging Center” Erlangen.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Chiorazzi N, Rai KR, Ferrarini M. Chronic lymphocytic leukemia. N Engl J Med. 2005;352(8):804–815.
- Lagneaux L, Delforge A, Bron D, et al. Chronic lymphocytic leukemic B cells but not normal B cells are rescued from apoptosis by contact with normal bone marrow stromal cells. Blood. 1998;91(7):2387–2396.
- Ding W, Nowakowski GS, Knox TR, et al. Bi-directional activation between mesenchymal stem cells and CLL B-cells: implication for CLL disease progression. Br J Haematol. 2009;147(4):471–483.
- Lutzny G, Kocher T, Schmidt-Supprian M, et al. Protein kinase c-beta-dependent activation of NF-kappaB in stromal cells is indispensable for the survival of chronic lymphocytic leukemia B cells in vivo. Cancer Cell. 2013;23(1):77–92.
- Meads MB, Gatenby RA, Dalton WS. Environment-mediated drug resistance: a major contributor to minimal residual disease. Nat Rev Cancer. 2009;9(9):665–674.
- Park E, Chen J, Moore A, et al. Stromal cell protein kinase C-beta inhibition enhances chemosensitivity in B cell malignancies and overcomes drug resistance. Sci Transl Med. 2020;12(526):eaax9340.
- Jitschin R, Braun M, Qorraj M, et al. Stromal cell-mediated glycolytic switch in CLL cells involves Notch-c-Myc signaling. Blood. 2015;125(22):3432–3436.
- Mangolini M, Gotte F, Moore A, et al. Notch2 controls non-autonomous Wnt-signalling in chronic lymphocytic leukaemia. Nat Commun. 2018;9(1):3839.
- Maffei R, Fiorcari S, Martinelli S, et al. Targeting neoplastic B cells and harnessing microenvironment: the "double face" of ibrutinib and idelalisib. J Hematol Oncol. 2015;8:60.
- Cheah CY, Fowler NH. Idelalisib in the management of lymphoma. Blood. 2016;128(3):331–336.
- Zirlik K, Veelken H. Idelalisib. Recent Results Cancer Res. 2018;212:243–264.
- George JA, Alshebli Z, Alneyadi A, et al. Idelalisib induces apoptosis in the lymphoid tissues and impairs lung function in mice. J Chemother. 2020;32(2):88–97.
- ten Hacken E, Burger JA. Microenvironment dependency in chronic lymphocytic leukemia: the basis for new targeted therapies. Pharmacol Ther. 2014;144(3):338–348.
- Walter DM, Venancio OS, Buza EL, et al. Systematic in vivo inactivation of chromatin-regulating enzymes identifies setd2 as a potent tumor suppressor in lung adenocarcinoma. Cancer Res. 2017;77(7):1719–1729.
- Schindelin J, Arganda-Carreras I, Frise E, et al. Fiji: an open-source platform for biological-image analysis. Nat Methods. 2012;9(7):676–682.
- Oostendorp RA, Harvey KN, Kusadasi N, et al. Stromal cell lines from mouse aorta-gonads-mesonephros subregions are potent supporters of hematopoietic stem cell activity. Blood. 2002;99(4):1183–1189.
- Osipo C, Golde TE, Osborne BA, et al. Off the beaten pathway: the complex cross talk between Notch and NF-kappaB. Lab Invest. 2008;88(1):11.
- Bhattacharya P, Thiruppathi M, Elshabrawy HA, et al. GM-CSF: an immune modulatory cytokine that can suppress autoimmunity. Cytokine. 2015;75(2):261–271.
- Dimberg LY, Anderson CK, Camidge R, et al. On the TRAIL to successful cancer therapy? Predicting and counteracting resistance against TRAIL-based therapeutics. Oncogene. 2013;32(11):1341–1350.
- Walsh AJ, Cook RS, Sanders ME, et al. Quantitative optical imaging of primary tumor organoid metabolism predicts drug response in breast cancer. Cancer Res. 2014;74(18):5184–5194
- Neal JT, Li X, Zhu J, et al. Organoid modeling of the tumor immune microenvironment. Cell. 2018;175(7):1972–1988.
- Holloway EM, Capeling MM, Spence JR. Biologically inspired approaches to enhance human organoid complexity. Development. 2019;146(8):dev166173.
- Wimmer RA, Leopoldi A, Aichinger M, et al. Human blood vessel organoids as a model of diabetic vasculopathy. Nature. 2019;565(7740):505–510.
- Dekkers JF, Wiegerinck CL, de Jonge HR, et al. A functional CFTR assay using primary cystic fibrosis intestinal organoids. Nat Med. 2013;19(7):939–945.
- Seifert M, Lubitz A, Trommer J, et al. Crosstalk between immune cells and mesenchymal stromal cells in a 3D bioreactor system. Int J Artif Organs. 2012;35(11):986–995.
- Czerniecki SM, Cruz NM, Harder JL, et al. High-throughput screening enhances kidney organoid differentiation from human pluripotent stem cells and enables automated multidimensional phenotyping. Cell Stem Cell. 2018;22(6):929–940.
- Eliopoulos N, Francois M, Boivin MN, et al. Neo-organoid of marrow mesenchymal stromal cells secreting interleukin-12 for breast cancer therapy. Cancer Res. 2008;68(12):4810–4818.