
 ?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.
?Mathematical formulae have been encoded as MathML and are displayed in this HTML version using MathJax in order to improve their display. Uncheck the box to turn MathJax off. This feature requires Javascript. Click on a formula to zoom.Abstract
Cattle are sensitive to temperature fluctuations but adapt well to inclement weather conditions. When environmental temperatures exceed specific thresholds, heat stress becomes a critical concern for cattle. The TRPM2 gene, which resides on cattle chromosome 1 encodes a TRP channel protein, holding a unique capacity to sense temperature changes and facilitate rapid response to avoid heat stress. Here, we utilized the Bovine Genome Variation Database (BGVD) (http://animal.omics.pro/code/index.php/BosVar), and identified a missense mutation site, c.805A > G: p. Met269Val (rs527146862), within the TRPM2 gene. To elucidate the functional assessment of this mutation in temperature adaptation attributes of Chinese cattle, we genotyped 407 samples from 20 distinct breeds representing diverse climatic zones across China. The association analysis incorporates three temperature parameters and revealed compelling insights in terms of allele frequency. Interestingly, the prevalence of the wild-type allele A was notably higher among northern cattle breeds and this trend diminished gradually as observed in southern cattle populations. Conversely, the mutant-type allele G demonstrated a contrasting trend. Moreover, southern cattle exhibited markedly higher frequencies of GG and GA genotypes (P < 0.01). The presence of heterozygous and homozygous mutations appears to confer an enhanced capacity for adaptation to elevated temperatures. These results provide unequivocal correlation evidence between TRPM2 genotypes (AA, GA, GG) and environmental temperature parameters and comprehend the genetic mechanisms governing temperature adaptation in cattle. This provides valuable insights for strategic breed selection across diverse climatic regions, thereby aiding livestock production amid evolving climate challenges.
SIMPLE SUMMARY
The TRPM2 gene encodes TRP channel protein that helps animals in combating heat stress. Twenty Chinese local cattle breeds were genotyped, and association analysis was performed. This investigation encompasses the distribution pattern of the missense mutation locus rs527146862 of the TRPM2 gene in southern, northern, and central cattle populations. The results demonstrated a significant relationship between rs527146862 locus and temperature adaptation attributes in Chinese cattle.
Introduction
China has a unique geographical location that forms the intersection area of Bos taurus and Bos indicus cattle in the Central Plains.Citation1 The geographical distribution pattern of domestic cattle across the world is closely related to the climatic background.Citation2 B. Taurus cattle are cold-resistant and adapt to temperate and cold climates and are predominantly distributed in the northern hemisphere of the globe, with a wide distribution range. Whereas, B. Indicus cattle are heat and drought-resistant cattle and are well-adapted to tropical and subtropical climates. It is mainly distributed in the equatorial region and the southern hemisphere, including South Asia, Southeast Asia, Africa, and America.Citation3 B. Taurus was introduced into northern China between 5000 and 4000 years ago,Citation4 while B. indicus was introduced into southern China from the Indian subcontinent between 3500 and 2500 years ago.Citation5 After that, population migration and geographical environment changes caused B. taurus and B. indicus to exchange genes across these two divergent populations. The analysis of the whole genome involving 20 local breeds across distinct regions of China has substantiated the origin of local Chinese cattle breeds from both B. taurus and B. indicus. Notably, the pedigree component of B. indicus exhibited a gradual increase from northern to southern regions, with a pronounced effect of Bos indicus cattle from South and Southwest China. Conversely, the prevalence of B. taurus was more prominent, and cattle from the northern regions exhibited nearly negligible lineage of Bos indicus. Citation6 In addition, the maternal and paternal studies on Chinese cattle further confirmed this conclusion.Citation7,Citation8
The domestication and breeding efforts of Chinese farmers across diverse landscapes have resulted in a valuable genetic resource for local cattle in China. The genetic diversity of most cattle gene pools is influenced by both B. taurus and B. indicus, contributing to the rich diversity.Citation9 Based on geographical locations, cattle are affected by B. taurus and B. indicus to certain degrees and can be divided into northern cattle, Central Plains cattle, and southern cattle. Purebred B. taurus are mainly found in the Tibetan Plateau, the Northeast of China, and the Mongolian Plateau. Mongolian cattle originating from B. taurus are representative of the northern group, with characteristics of excellent cold hardiness traits.Citation10 The pure Zebu cattle(B. indicus) group herds are mainly found in the south, such as Hainan cattle, and are adaptive to humid and hot environments exposed to strong ultraviolet rays.Citation11 Cattle from other intermediate regions are crossbreeds of B. taurus and B. indicus such as Qinchuan cattle in the Central Plains, which bear certain hybridization advantages and more excellent environmental adaptability.Citation12,Citation13
The capacity to perceive warmth and heat is critical for the survival of all animals enabling to identification of suitable thermal environments for various life-sustaining activities and preventing damage from extreme temperatures.Citation14 The ancestors of cattle inhabited colder regions, and their prolonged evolutionary process rendered them more tolerant to cold than heat. Consequently, investigating the mechanisms behind heat tolerance in cattle holds scientific relevance in light of the ongoing global temperature increase. Moreover, it can be helpful in considerations for cattle breeding environments, and the diversification of their habitats. Many genes related to cattle heat tolerance traits have been selected employing Y-SNP labelling, mitochondrial fragments, low-density gene chip technology, and whole-genome resequencing technology.Citation15 Transient receptor potential (TRP) is a superfamily of ion channel proteins that exist in the membrane of a cell or intracellular organelles. The transient receptor potential (TRP) family was first discovered in Drosophila melanogasterCitation16 which encodes a temperature-related protein involved in the environmental temperature sensing of the organism and is called the Thermo TRP motif.Citation17 The TRPM2 gene is located on chromosome 1 in cattle and as a subfamily member of the TRP channel superfamily, the TRPM2 protein is a nonselective cationic channel involved in oxidative stress responses as a redox receptor. It is a protein with two functions: cation channel and adenosine diphosphate ribose hydrolase activity.Citation18 TRPM2 channel proteins are involved in forming homopolymers;Citation19 regulating body temperature, promote to produce inflammation and apoptosis, and stimulate insulin secretion.Citation20,Citation21 The TRPM2 protein, initially identified in the human brain in 1998,Citation22 shares structural similarities with other TRP family channel proteins that consist of six transmembrane structural domains and form a channel between transmembrane structural domains 5 and 6. The TRPM2 protein comprises 1503 amino acid residuesCitation23 and exhibits widespread expression in various tissues and cell types across organisms such as worms, fruit flies, zebrafish, mice, and humans.Citation24 Additionally, the TRPM2 gene mobilizes Ca2+ from both extracellular and intracellular levels, so its biological significance is related to cellular function regulated by Ca2+. Moreover, TRPM2 serves as a calcium-permeable, non-selective cation channel, exerting pathological influences in diverse neuroinflammatory diseases.Citation25 Its pivotal role extends to the regulation of key transcription factors and target genes associated with mitochondrial function, biological processes, antioxidant response, and autophagy, thereby influencing the proliferation and survival of acute myeloid leukaemia cells.Citation26 The inhibition of TRPM2 expression or function leads to reduced tumour proliferation and viability across several malignancies, encompassing breast, gastric, pancreatic, prostate, head and neck cancers, melanoma, neuroblastoma, and T-cell acute myelogenous leukaemia.Citation27 Notably, in rat insulinoma RIN-5F cells, temperatures exceeding 35 °C have been shown to activate TRPM2 channelsCitation28 or promote their activation through its activator ADPR (cARDPR).Citation29 This provides a theoretical foundation for exploring the correlation between genetic polymorphisms of the TRPM2 gene and climatic adaptation parameters in cattle. Given the functional context of the TRPM2 gene and diverse climatic adaptations observed in Chinese cattle populations, our study was crafted to explore mutations in the TRPM2 gene and their association with varying temperature zones across China. We aimed to identify a missense mutation site, c.805A > G:p. Met269Val (rs527146862), in the TRPM2 gene by selecting representative 20 cattle breeds spanning from south to north in China to examine the frequency distribution of this mutation site and correlation with the temperature and humidity indices of local cattle breeds and to elucidate how this mutant locus influences the temperature adaptation of different local cattle breeds in China.
Materials and methods
Ethics statement
All animal experiments were conducted following applicable laws and guidelines. Furthermore, these experiments were approved by the College of Animal Science, and the Ethical and Welfare Committee of Northwest A&F University (FAPWC-NWAFU; WAFAC1008).
Data collection, DNA extraction, and PCR
We collected ear tissue samples from diverse cattle breeds located in twenty different regions across China, sourced specifically from designated conservation and state-owned farms. The genomic DNA from 407 samples was extracted by the standard phenol-chloroform method.Citation30 Following this, through utilizing the BGVD (Bovine Genome Variation Database and Selective Signatures) online database, we accessed the genomic signature section and inputted the TRPM2 gene along with its chromosomal position to retrieve the missense mutation site c.805A > G: p. Met269Val (rs527146862.Citation31 Referring to the published nucleotide sequence information of the bovine TRPM2 gene in NCBI (GenBank accession NC_037328.1) and incorporating the SNP missense mutation site, we designed a pair of oligonucleotide primers, synthesized by Sangon Biotech Co. Ltd. which is as follows: Forward 5′- CCACCTCTCTCCATCCCCA −3′ and Reverse: 5′-GGTCCCATCGTCCACAAGAA-3′.
To determine the most suitable annealing temperature, we conducted a gradient PCR. The optimal annealing temperature was identified as 60 °C, with the optimal cycle number set at 32. Following PCR amplification, conventional agarose gel electrophoresis was conducted and images were captured under the imaging system, revealing a distinct and consistent target band with no nonspecific amplification. The remaining amplicon fragments underwent sequencing by Sangon Biotech Co. Ltd. (Shanghai, China.). The sequencing results were subsequently analyzed SEQMAN TMIIv 6.1 (DNASTAR, Inc., Madison, USA). Following a thorough quality check via agarose gel electrophoresis, the selected tissue samples exhibited consistently uniform and bright bands, devoid of any noticeable specific amplification. The decision to choose these samples was made after careful comparison, resulting in the identification of a 122 bp sequence length. Analyzing the genotyping results derived from sequencing was facilitated using Chromas software (technelysium.com).
Statistical analysis
Genotypic and allelic frequencies were calculated based on the observed genotypes in the analysed breeds. Hardy–Weinberg equilibrium (HWE)Citation32 was tested based on the likelihood ratio for different locus–population combinations, and the number of observed and effective alleles was determined using POPGENE software. Mean annual temperature (T), and humidity (H) were taken from the National Oceanic and Atmospheric Administration (1995–2021) (Supplement Table 1) (Supplement Figure 2),Citation33 and the classical index was used to evaluate the degree of thermal stress, which is calculated as:
where T is the temperature, and RH is the relative humidity.
SPSS (Version 26.0, Inc, Armonk, New York, NY, USA) software was used to analyse the relationship between the genotypes and climatic data and the linear model:
Yi represents the value of the environmental parameter; μ is the overall mean; Gi is the fixed genotype effect; Bi is the fixed-effect variety; and ei is the random residual effect. Taking p < 0.01 as the criterion for having a significant difference.
The TRPM2 protein sequence (XP_015317659.1) was used as a control. The homology testing of missense mutation site c.805A > G: p. Met269Val (rs527146862) for assessment of alterations in protein structure and to predict nucleotide changes, we employed the SWISS-MODEL software.Citation34
Results
PCR amplification and sequencing
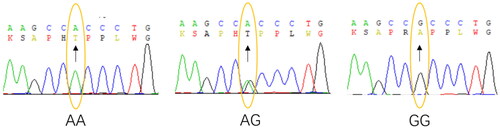
All the obtained bands of amplicon of 122 bp sequence lengths were uniform and bright, affirming successful priming and amplification. The outcome of the PCR product can be visualized in agarose gel (Supplementary Figure 1). Furthermore, the Oscillographic preview also demonstrated clear sequence peaks of nucleotides across 122 bp fragments and confirmed successful sequencing. The Chromas-generated results revealed the presence of three distinct genotypes—AA (wild type), AG (partial mutation), and GG (complete mutation)—across the three groups of twenty unrelated cattle samples. Additionally, a geographical distribution pattern of these genotypes was also observed ().
Figure 1. Sequencing map for NC_037328.1: c.805A > G:p. Met269Val locus of the TRPM2 gene.
Allele frequency analysis and association analysis
The data of association analysis and subsequent estimations with three environmental parameters—T, RH, and THI—as well as the genotyped allele frequencies are summarized in .
Table 1. Results of correlation analysis of annual mean temperature, humidity, and humidity-temperature index.
The findings indicate a significant correlation (P < 0.01) between the distribution of three genotypes (AA, AG, and GG) and environmental parameters. This underscores the pivotal role of a missense mutation in the TRPM2 gene in conjunction with climatic factors. Moreover, the mutation in Chinese cattle appears of robust positive selection influenced by climate variables such as temperature and humidity.
The summary statistics of association analysis of 407 samples revealed an allele frequency of 0.5479 for allele A and 0.4521 for allele G, as detailed in and the sequencing results used for statistical analyses are listed in Supplement Table 2. Further scrutiny of genotypes and complete mutant genotypes within the northern cattle population affirmed the presence of mutants in regions from north to south. The calculation of genetic diversity is list in Supplement Table 3 and the allele frequencies of the different breeds are presented as pie charts on a standard Chinese map ().
Figure 2. Distribution of a and G alleles and AA, AG, and GG genotypes in 20 Chinese cattle breeds.
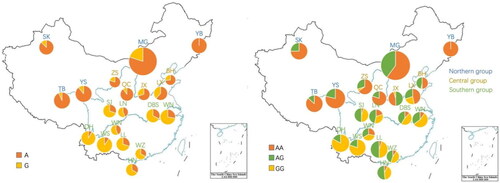
Table 2. Species, population, and genotyping tables for a sample of 407 individuals.
Mutation analysis
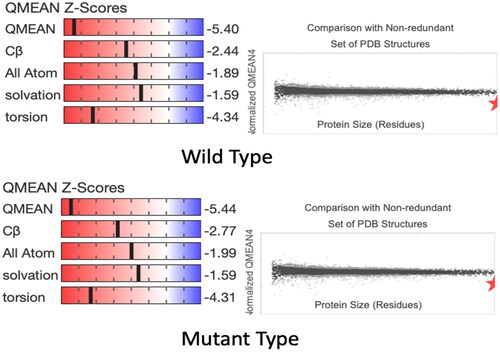
The substitution at the mutation site c.805A > G: p.Met269Val involves replacing A with G allele, resulting in the alteration of amino acid number 268 from threonine to alanine (). The analysis of the predicted three-dimensional structure (3D) using SWISS-MODEL software further elucidates the impact of this missense mutation at the protein structural level. Four structural descriptors—C-Beta, All Atom, solvation, and torsion—, revealed a significant difference in C-Beta () and this distinction in domain differences explains the consequence of the missense mutation. While predictions of the protein structure of the TRPM2 gene may not exhibit notable change due to individual amino acid shifts, however, alterations in the model undoubtedly highlight the presence of the missense mutation.
Figure 3. Protein structural changes.
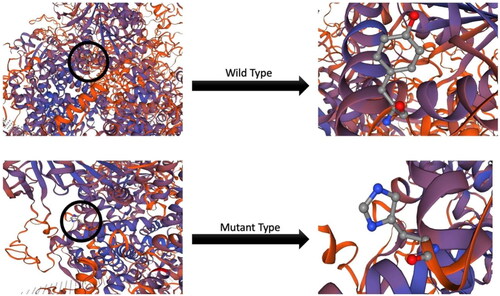
Figure 4. QMEAN Z Score results.
Discussion
Biological evolution occurs over a long and complex course. Within this process, cattle undergo adaptation to climate change, wherein mutations at various gene loci favour environmental adaptation based on selective retention. Guided by contemporary evolutionary theory, the direction of species evolution is shaped by natural selection, and every individual is subjected to the forces of natural selection, eliminating those ill-suited to the environment and perpetuating those better adapted.Citation35
Cattle have developed various mechanisms to adapt to different temperatures, which are crucial for their survival and well-being.Citation36 After being introduced into China 5,000 years ago, cattle continued to undergo natural selection across the various regions of China. China spans several temperature zones, including temperate continental, temperate monsoon, subtropical monsoon, tropical monsoon, and alpine plateau climate.Citation37 It also boasts complex landforms, such as the highest Qinghai-Tibet Plateau, which is found in the southwestern region. The Tibetan cattle population adapted to anoxic environments at high altitudes and this breed stands apart from other southern cattle breeds. Its distinctive characteristics align more closely with those of northern cattle populations, a trait attributed to extensive and enduring selective processes. Moreover, Chinese cattle breeds have been subjected to intense selective pressures under such diverse environmental conditions, resulting in notable population differentiation. Northern cattle display fewer heat-tolerant traits than southern cattle, hence these breeds have fewer heat-tolerant mutations due to selection pressures.
TRPM2 gene is a key player in thermoregulation and warmth sensation in animals and is responsible for detecting non-noxious warmth. It is expressed in autonomic neurons, suggesting its involvement in thermoregulation and immune responses.Citation38 This functional phenomenon was validated in mice experiments where 10% of somatosensory neurons in the dorsal root ganglion in mice are activated by the heat response to TRPM2.Citation39 A past study has also demonstrated that TRPM2 is intrinsically heat-sensitive and can be activated within the physiological range of body temperature, making it an essential component of the animal to sense and respond to temperature change.Citation40 The expression of the TRPM2 gene in these animals promotes the ability to detect and regulate body temperature, thereby enabling them to seek out optimal thermal environments. In this way the screened missense mutation c.805A > G: p. Met269Val (rs527146862) of TRPM2 might help the cattle during evolutionary adaptation to hot and humid environments of the tropics.
Results from this analysis showed that the frequency of allele A decreased in northern, central, and southern cattle. Notably, there was a significant downward trend in frequency from north to south, suggesting that missense mutation is more prevalent among cattle breeds in environments with high temperatures and humidity. Furthermore, we found that the missense mutation is significantly associated with annual average T, H, and THI where the cattle are distributed. The A allele has a decreasing trend from south to north. The AA genotype was found to be highly frequent in northern cattle, particularly Yanbian and Tibetan cattle. On the other hand, certain mutant types were relatively abundant in central plains cattle, especially in Dehong, Hainan, and Wenshan cattle. Southern cattle of China are known for heat tolerance, and identification of TRPM2 mutation which might assimilate temperature information, orchestrating autonomic and behavioural adaptations crucial for maintaining thermal homeostasis. In addition, protein structure prediction analysis showed a missense mutation that might cause the amino acid changes linked to the large structure of the TRPM2 protein. Furthermore, it may affect the physicochemical properties of the protein, and however, follow-up studies to unravel the substitution effect of alleles can be another research direction in this study.
Collectively, this research contributes to the functional attributes of TRP family members in mammals. In addition, it enhanced our comprehension of the TRPM2 gene’s pivotal role in temperature regulation. Notably, this study pioneers the exploration of the correlation between genetic polymorphism of TRPM2 and climate adaptation and offers valuable insights and benchmarks for the selection of breeding cattle residing in diverse temperature environments.
Conclusion
In summary, Our study reveals a significant TRPM2 gene missense mutation (c.805A > G:p.Met269Val) in Chinese cattle, intimately tied to geographic and climatic factors. The patterning of allele frequency shifts in northern, central, and southern cattle groups elucidate the potential linkage of the TRPM2 gene with diverging climatic conditions, conferring heightened resilience to heat stress in heterozygous and homozygous carriers. This discovery holds practical implications for cattle breeding, enabling informed decisions to boost livestock productivity and resilience amid ongoing climate fluctuations.
Supplemental Material
Download MS Excel (11.9 KB)Supplemental Material
Download MS Excel (13.6 KB)Supplemental Material
Download MS Excel (11.5 KB)Supplemental Material
Download PNG Image (9.9 MB){kind=link}
Supplemental Material
Download PNG Image (3 MB){kind=link}
Acknowledgments
Thanks to the High-Performance Computing (HPC) of Northwest A&F University (NWAFU) for providing computing resources.
Disclosure statement
No potential conflict of interest was reported by the author(s).
Additional information
Funding
References
- Lei CZ, Chen H, Zhang HC, et al. Origin and phylogeographical structure of Chinese cattle. Anim Genet. 2006;37(6):579–582.
- Zhang K, Lenstra JA, Zhang S, Liu W, Liu J. Evolution and domestication of the Bovini species. Anim Genet. 2020; 51(5):637–657.
- Utsunomiya YT, Milanesi M, Fortes MRS, et al. Genomic clues of the evolutionary history of Bos indicus cattle. Anim Genet. 2019;50(6):557–568.
- Cai D, Sun Y, Tang Z, et al. The origins of Chinese domestic cattle as revealed by ancient DNA analysis. J Archaeolog Sci. 2014;41:423–434.
- Chen S, Lin BZ, Baig M, et al. Zebu cattle are an exclusive legacy of the South Asia neolithic. Mol Biol Evol. 2010;27(1):1–6.
- Gao Y, Gautier M, Ding X, et al. Species composition and environmental adaptation of indigenous Chinese cattle. Sci Rep. 2017;7(1):16196.
- Li R, Xie WM, Chang ZH, et al. Y chromosome diversity and paternal origin of Chinese cattle. Mol Biol Rep. 2013;40(12):6633–6636.
- Jia S, Zhou Y, Lei C, et al. A new insight into cattle’s maternal origin in six Asian countries. J Genet Genomics. 2010;37(3): 173–180.
- Gong M, Yang P, Fang W, Li R, Jiang Y. Building a cattle pan-genome using more de novo assemblies. J Genet Genomics. 2022;49(9):906–908.
- Buggiotti L, Yurchenko AA, Yudin NS, et al. Demographic history, adaptation, and NRAP convergent evolution at amino acid residue 100 in the world northernmost cattle from Siberia. Mol Biol Evol. 2021;38(8):3093–3110.
- Xia X, Zhang F, Li S, et al. Structural variation and introgression from wild populations in East Asian cattle genomes confer adaptation to local environment. Genome Biol. 2023;24(1):211.
- Sinding MS, Ciucani MM, Ramos-Madrigal J, et al. Kouprey (Bos sauveli) genomes unveil polytomic origin of wild Asian Bos. iScience. 2021;24(11):103226.
- Chen N, Cai Y, Chen Q, et al. Whole-genome resequencing reveals world-wide ancestry and adaptive introgression events of domesticated cattle in East Asia. Nat Commun. 2018;9(1):2337.
- Tan CH, McNaughton PA. TRPM2 and warmth sensation. Pflugers Arch. 2018; 470(5):787–798.
- Vulczak A, Catalão CHR, Freitas LAP, Rocha MJA. HSP-Target of Therapeutic Agents in Sepsis Treatment. Int J Mol Sci. 2019; 20(17):4255.
- Lindström JB, Pierce NT, Latz MI. Role of TRP Channels in Dinoflagellate Mechanotransduction. Biol Bull. 2017; 233(2):151–167.
- Yang KT, Chang WL, Yang PC, et al. Activation of the transient receptor potential M2 channel and poly(ADP-ribose) polymerase is involved in oxidative stress-induced cardiomyocyte death. Cell Death Differ. 2006;13(10):1815–1826.
- Wang S, Liang T, Luo Q, et al. H9N2 swine influenza virus infection-induced damage is mediated by TRPM2 channels in mouse pulmonary microvascular endothelial cells. Microb Pathog. 2020;148:104408.
- Lu J, Shen Y, Qian HY, et al. Effects of mild hypothermia on the ROS and expression of caspase-3 mRNA and LC3 of hippocampus nerve cells in rats after cardiopulmonary resuscitation. World J Emerg Med. 2014;5(4):298–305.
- Malko P, Jiang LH. TRPM2 channel-mediated cell death: An important mechanism linking oxidative stress-inducing pathological factors to associated pathological conditions. Redox Biol. 2020; 37:101755.
- Tan CH, McNaughton PA. The TRPM2 ion channel is required for sensitivity to warmth. Nature. 2016; 536(7617):460–463.
- Nagamine K, Kudoh J, Minoshima S, et al. Molecular cloning of a novel putative Ca2+ channel protein (TRPC7) highly expressed in brain. Genomics. 1998;54(1):124–131.
- Moore C, Gupta R, Jordt SE, Chen Y, Liedtke WB. Regulation of Pain and Itch by TRP Channels. Neurosci Bull. 2018;34(1):120–142.
- Moran MM. TRP channels as potential drug targets. Annu Rev Pharmacol Toxicol. 2018;58(1):309–330.
- Shao Y, Chen C, Zhu T, et al. TRPM2 contributes to neuroinflammation and cognitive deficits in a cuprizone-induced multiple sclerosis model via NLRP3 inflammasome. Neurobiol Dis. 2021; 160:105534.
- Wertheim BM, Lin YD, Zhang YY, et al. Isolating pulmonary microvascular endothelial cells ex vivo: Implications for pulmonary arterial hypertension, and a caution on the use of commercial biomaterials. PLoS One. 2019;14(2):e0211909.
- Miller BA. TRPM2 in Cancer. Cell Calcium. 2019;80:8–17.
- Togashi K, Hara Y, Tominaga T, et al. TRPM2 activation by cyclic ADP-ribose at body temperature is involved in insulin secretion. Embo J. 2006;25(9):1804–1815.
- Lange I, Penner R, Fleig A, Beck A. Synergistic regulation of endogenous TRPM2 channels by adenine dinucleotides in primary human neutrophils. Cell Calcium. 2008; 44(6):604–615.
- Wu C, McCune TS. Implementation of research on potential drug target cloning and characterization in a biochemistry laboratory. Biochem Mol Biol Educ. 2020;48(2):108–117.
- Chen N, Fu W, Zhao J, et al. BGVD: an integrated database for bovine sequencing variations and selective signatures. Genomics Proteomics Bioinformatics. 2020;18(2):186–193.
- Mayo O. A century of Hardy-Weinberg equilibrium. Twin Res Hum Genet. 2008;11(3):249–256.
- Saud S, Wang D, Fahad S, et al. Comprehensive impacts of climate change on rice production and adaptive strategies in China. Front Microbiol. 2022;13:926059.
- Waterhouse A, Bertoni M, Bienert S, et al. SWISS-MODEL: homology modelling of protein structures and complexes. Nucleic Acids Res. 2018;46(W1):W296–W303.
- Kuratani S, Uesaka M, Irie N. How can recapitulation be reconciled with modern concepts of evolution? J Exp Zool B Mol Dev Evol. 2022; 338(1-2):28–35.
- Rashamol VP, Sejian V, Bagath M, et al. Physiological adaptability of livestock to heat stress: an updated review. JABB. 2018;6(3):62–71.
- Chen N, Ren L, Du L, et al. Ancient genomes reveal tropical bovid species in the Tibetan Plateau contributed to the prevalence of hunting game until the late Neolithic. Proc Natl Acad Sci U S A. 2020;117(45):28150–28159.
- Kaneko S, Kawakami S, Hara Y, et al. A critical role of TRPM2 in neuronal cell death by hydrogen peroxide. J Pharmacol Sci. 2006;101(1):66–76.
- Vilar B, Tan CH, McNaughton PA. Heat detection by the TRPM2 ion channel. Nature. 2020;584(7820):E5–E12.
- Bartók Á, Csanády L. Dual amplification strategy turns TRPM2 channels into supersensitive central heat detectors. Proc Natl Acad Sci USA. 2022;119(48):e2212378119.