ABSTRACT
Idaho National Laboratory's Test Area North is the site of a trichloroethene (TCE) plume resulting from waste injections. Previous investigations revealed that TCE was being attenuated relative to two codisposed internal tracers, tritium and tetrachloroethene, with a half-life of 9 to 21 years. Biological attenuation mechanisms were investigated using a novel suite of assays, including enzyme activity probes designed for the soluble methane monooxygenase (sMMO) enzyme. Samples were analyzed for chlorinated solvents, tritium, redox parameters, primary substrates, degradation products, bacterial community methanotrophic potential, and bacterial DNA. The enzyme probe assays, methanotrophic enrichments and isolations, and DNA analysis documented the presence and activity of indigenous methanotrophs expressing the sMMO enzyme. Three-dimensional groundwater data showed plume-wide aerobic conditions, with low levels of methane and detections of carbon monoxide, a by-product of TCE cometabolism. The TCE half-life attributed to aerobic cometabolism is 13 years relative to tritium, based on the tracer-corrected method. Similarly, a half-life of 8 years was estimated for cis-dichloroethene (DCE). Although these rates are slower than most anaerobic degradation processes, they can be significant for large plumes. This investigation is believed to be the first documentation of intrinsic aerobic TCE and DCE cometabolism in an aquifer by indigenous methanotrophs.
INTRODUCTION
Monitored natural attenuation (MNA) has recently gained favor as a viable remedy for chlorinated-solvent contaminated groundwater under certain conditions (CitationEPA, 1999). Several degradation pathways have been elucidated, though the environmental significance of some remains uncertain. Under anaerobic conditions, chlorinated ethenes can be degraded by reductive dechlorination (CitationFreedman and Gossett, 1989; CitationVogel et al., 1987) and by abiotic reduction (CitationButler and Hayes, 1999; CitationLee and Batchelor, 2002). Trichloroethene (TCE), cis-DCE (dichloroethene), trans-DCE, 1,1-DCE, and vinyl chloride (VC) have also been repeatedly shown to be susceptible to cometabolic oxidation under aerobic conditions (CitationAlvarez-Cohen and Speitel, 2001). In addition, DCE and VC have been shown to undergo direct oxidation under both aerobic (CitationVogel et al., 1987, CitationBradley and Chapelle, 2000) and anoxic conditions (CitationBradley and Chapelle, 1998), whereas tetrachloroethene (PCE) has been shown to be resistant to both direct and cometabolic oxidation under aerobic conditions (CitationMcCarty, 1997).
Although cometabolic oxidation of chlorinated ethenes has been widely studied, its importance under natural environmental conditions has not. Several primary substrates have been shown to induce cometabolic oxidation, including methane, propane, butane, phenol, toluene, and ammonia (CitationAlvarez-Cohen and Speitel, 2001). Among the most-studied of the TCE-cometabolizing microbes are methanotrophic bacteria, which are aerobic gram-negative organisms that use methane as their sole source of carbon and energy (CitationAnthony, 1982). Methane monooxygenase (MMO), an enzyme produced by methanotrophic bacteria, catalyzes the oxidation of methane to methanol. Some methanotrophs have the capacity to synthesize both soluble (sMMO) and particulate membrane-bound (pMMO) forms of the enzyme; however, only the particulate form of the enzyme is present in all known methanotrophs (CitationSemrau et al., 1995). Both types of MMO are capable of cometabolic TCE oxidation (CitationDiSpirito et al., 1992), but sMMO oxidizes TCE many times faster than pMMO (CitationTsien et al., 1989; CitationOldenhuis et al., 1991) and has a much broader substrate range (CitationLontoh and Semrau, 1998).
Although aerobic cometabolism has been applied as an engineered in situ remedy at several sites (CitationHopkins and McCarty, 1995; CitationSemprini et al., 1990), conventional thinking is that cometabolism is likely of limited importance under intrinsic conditions due to competitive inhibition and the specific environmental conditions needed for significant degradation (CitationMcCarty, 1997; CitationSewell and Gibson, 1997). Numerous studies have detected and isolated methanotrophic bacteria, and although some have measured activity in a laboratory setting (CitationBowman et al., 1993; CitationAuman et al., 2000; CitationMiguez et al., 1997), few have documented intrinsic cometabolism in the field as an important attenuation mechanism, or enzyme activity in unaltered samples. Even if cometabolism were important at a site, detection of this mechanism is problematic due to the completely mineralized contaminants and the short-lived intermediates. Thus, the primary evidence for degradation is the disappearance of the contaminants themselves. However, measurement of decreasing contaminant concentrations in groundwater alone is of limited usefulness in assessing the importance of intrinsic cometabolism as an attenuation mechanism independent of dispersion and dilution. This difficulty can be overcome when it is possible to employ the tracer-corrected method for degradation rate estimation to quantify attenuation of a degrading contaminant relative to some internal, conservative tracer (CitationSorenson et al., 2000). Although this approach can effectively provide degradation rates that are independent of dispersion and dilution, it provides little insight as to the actual mechanism for degradation.
Further, methods commonly used for monitoring bacteria that are capable of cometabolism, such as colony and direct counting, and various molecular approaches, do not provide evidence of enzymatic activity, but rather only indicate the presence of the organism or of particular genes or gene expression levels. Enzyme activity probes that undergo cometabolic transformation by specific oxygenases to yield quantifiable fluorescent products only when the enzyme of interest is actively functioning have recently been developed (CitationKeener and Watwood, 1997; CitationKeener et al., 1998, Citation2001; Kaufmann et al., 2003). For example, sMMO activity can be evaluated with a rapid fluorescent assay based on coumarin (nonfluorescent) transformation to 7-hydroxycoumarin (fluorescent) (CitationKeener and Watwood, 1997). The transformation of these and other enzyme activity probes yields a product that enables either (a) direct visualization of cometabolic activity within bacterial cells or (b) measurement of activity in solution. This class of probes provides direct evidence of cometabolic enzyme activity in situ through the use of unaltered samples representing natural and noninduced conditions. As such, these probes provide direct evidence of enzymatic activity that, in turn, provides strong evidence for cometabolic activity toward chlorinated solvents at contaminated sites. Without other means for direct measurement, these data are essential for documenting that cometabolism of the contaminant is occurring in a given environment (CitationMiller et al., 2002). In addition to the activity-dependent enzyme probes, a series of molecular gene probes has been developed or adapted to investigate the genetic potential of methane-oxidizing microbial populations. Whereas activity-dependent enzyme probes measure actual activity and provide data that are otherwise not obtainable, molecular techniques can be used to provide context for the activity-dependent enzyme probes.
The conceptual model for the Test Area North (TAN) facility of the Idaho National Laboratory (INL) is thoroughly discussed by CitationSorenson (2000), and a summary relevant to the aerobic portion of the plume is presented here. The historical injection of liquid wastes and concentrated sludges into the Snake River Plain Aquifer (SRPA) using Well TSF-05 has resulted in a nearly 3-km-long TCE, PCE, and tritium plume in groundwater. The depth to water at TAN is approximately 60 m, and the contaminated thickness extends to nearly 185 m in downgradient portions of the plume. The aquifer and most of the unsaturated zone are comprised primarily of layered basalt flows, intercalated with sedimentary interbeds deposited during periods of volcanic quiescence. Groundwater flow in the aquifer is controlled by the highly transmissive zones that occur at the contacts between individual basalt flows, and to a lesser extent, by fractured zones within flow interiors. The scale of the basalt geology dictates that preferential flow can be very important at spatial scales less than approximately 100 m, after which a transition to continuum behavior occurs and the aquifer can be thought of essentially as a macroporous medium.
The distribution of TCE at TAN exemplifies the fringe and core hypothesis for the anatomy of chlorinated solvent plumes by CitationCherry (1997). A relatively large, low-concentration fringe surrounds and emanates from a much smaller, high concentration core (). Within the core is a very small residual source area that contaminates clean groundwater flowing through from upgradient. The transition from the scale of the residual source where preferential flow is significant to the scale of the fringe, where significant vertical communication has been present along the flow path, results in a relatively well-mixed, predictable groundwater plume.
FIGURE 1 Test Area North TCE plume showing wells sampled.
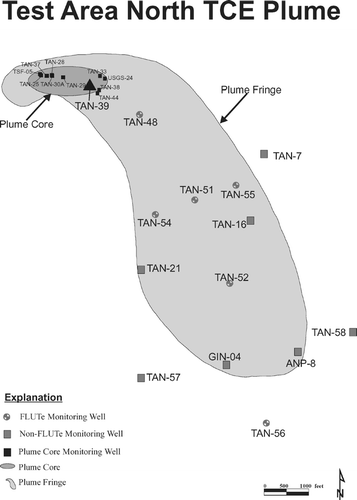
Previous investigations conducted in the aerobic portion of the TCE plume at TAN led to the refinement and application of the tracer-corrected method (CitationSorenson et al., 2000), which indicated that intrinsic TCE degradation was occurring at a rate that was significant relative to the required remedial time frame, but identification of the mechanism as aerobic cometabolism was initially tentative. Results showed that TCE decreased with distance from the source area in relation to two internal, codisposed tracers, PCE and tritium, with a relative half-life for TCE of 9 to 21 years (CitationSorenson et al., 2000). The fact that the tracers were subject to the same advective and dispersive transport mechanisms as TCE demonstrated that dispersion was not responsible for the observed concentration decreases. Given that this entire portion of the plume was aerobic, and that previous laboratory studies had shown that TCE sorption to the basalt matrix was insignificant (data not shown), these results indicated that the attenuation mechanism was destructive, and suggested that aerobic cometabolism was responsible.
Despite the initial findings at the TAN site, several data gaps, including a more definitive determination of the attenuation mechanism, remained. For example, the sampling activities that generated these initial data were performed using submersible pumps following standard high-flow-rate purge and sampling techniques, which are known to integrate samples across well screens and potentially dilute contaminant concentrations (CitationChurch and Granato, 1996; CitationWilliams and Conger, 1990). In addition, the wells included in these sampling activities had limited coverage of the entire contaminated thickness, as most were screened in the upper 30 m of the aquifer. The purpose of the current study was to address these data gaps to provide confirmation of degradation rates and a more definitive identification of the degradation mechanism.
The results herein present the first-ever combined application of activity-dependent enzyme probes and molecular methods to unaltered field samples for assessment of intrinsic aerobic cometabolism of chlorinated solvents as an important attenuation mechanism. These assays, along with numerous control studies, were conducted to implicate the sMMO enzyme as being responsible for the observed attenuation of TCE and cis-DCE at the site. This study also presents the groundwater chemistry associated with the intrinsic degradation collected from a network of multi-level sampling systems, and quantifies the degradation rates of TCE and cis-DCE under these conditions. This is the first field study to our knowledge to measure intrinsic activity of TCE cometabolizing enzymes, to characterize the associated groundwater chemistry, and to estimate in situ degradation rates.
MATERIALS AND METHODS
Groundwater Sample Collection
Multilevel sampling was performed to confirm that the observed attenuation of TCE at the site was not an artifact of a poor understanding of vertical contaminant distribution. Field testing of several multilevel sampling systems and approaches showed that a commercially available multilevel sampler, the Flexible Liner Underground Technology (FLUTe) sampling system, was best suited for application at TAN (CitationWymore et al., 2007). FLUTe systems were installed into six wells (), resulting in a total of 51 discrete multilevel sampling locations (CitationWymore et al., 2007).
Sampling was accomplished over two field seasons through the collection of samples from both FLUTe and non-FLUTe wells in the aerobic fringe of the plume. The wells equipped with FLUTe systems were sampled following the established protocols (CitationWymore et al., 2007; CitationCherry et al., 2007), and are represented by the round symbols on . The non-FLUTe wells were sampled following low-flow principles (i.e., minimal drawdown) (CitationPuls and Barcelona, 1996) using submersible pumps, and are represented by the square symbols on . Analyses included in this investigation were standard groundwater chemistry parameters, activity-dependent enzyme probe assays, and molecular techniques. Groundwater chemistry parameters included Volatile Organic Compound contaminants, redox-sensitive parameters, biological activity indicators, biological nutrients, and water quality parameters, which were analyzed using a combination of fixed and field laboratory techniques (). In addition, dissolved gas samples were collected following the bubble strip method (CitationChapelle et al., 1997) and analyzed for methane, hydrogen, and carbon monoxide by reduced gas analysis.
TABLE 1 Analytes and Analytical Methods
During the first year of the investigation, all depths in FLUTe wells TAN-51, -52, -54, and -55 were sampled for all of the groundwater chemistry parameters listed in . In addition, activity-dependent enzyme probe and molecular samples were collected in triplicate from TAN-7, and from three discrete depths in TAN-51, -52, and -55. During the second year of the investigation, all wells shown in except TAN-7 were sampled for chlorinated ethenes, dissolved gases, and tritium. Also in the second year, activity-dependent enzyme probe and molecular samples were collected from three depths in each of TAN-51, -52, and -57, as well as from a single depth in each of TAN-56, -58, and -16.
The depths from which activity-dependent enzyme probe samples were collected in FLUTe wells TAN-51 and TAN-52 were different during each year of this investigation. This strategy led to a total of 24 activity-dependent enzyme probe and molecular sampling locations, as well as extensive sampling for groundwater chemistry parameters from wells throughout the fringe of the plume and across the entire contaminated thickness.
Enzyme Probe and Molecular Sample Collection
Samples of groundwater (20 to 60 L) from each depth were filtered inline during field sample collection through 142-mm, 0.2-μ m hydrophilic Supor membranes (VWR) placed on sterile, autoclaved stainless-steel vacuum manifolds (Fisher Scientific). Filters were stored directly on ice (4°C) or on dry ice (for DNA extraction), whereas nonfiltered samples corresponding to each depth were collected in acid-washed carboys (capacity 10 to 19 L) and stored separately on ice for transport to the laboratory. All samples were analyzed for enzyme activity within 4 to 8 h of collection.
Coumarin Oxidation Assay
Each filter (142 mm) was cut aseptically into four quarters, which were placed into separate glass Petri plates. Two milliliters of 5 mM coumarin solution in phosphate buffer was pipetted onto each filter, and incubated for 10 min at room temperature. Following the incubation, each plate was positioned at a 35° angle, and 2 ml of modified nitrate mineral salts medium (NMS; CitationWhittenbury et al., 1970) were used to wash the product from each filter. The medium was repipetted over the filter six times to ensure quantitative recovery of the product. Solution fluorescence was determined (excitation wavelength 338 nm, emission wavelength 450 nm) using a Hitachi F-2000 fluorescence spectrophotometer (Hitachi, Tokyo, Japan), with a quartz cuvette of 1 cm pathlength (Amersham). Fluorescent scans were performed in triplicate for each of the samples (three of the four quarters of each filter).
In addition, groundwater samples were exposed to coumarin as described previously (CitationKeener and Watwood, 1997) with minor modifications. Two milliliters of groundwater were pipetted into a 10-ml glass serum vial and sparged for 2 to 4 min with compressed air. Coumarin was added to a final concentration of 500 μ M, and vials were incubated for approximately 10 min. The fluorescence of each solution was analyzed as described above.
To verify the dependence of product formation on sMMO and not other oxygenase activity, phenylacetylene was added to a separate set of whole water samples as an MMO inhibitor. Phenylacetylene (PA) is a differential inhibitor of the soluble and particulate forms of the enzyme at relatively low concentrations (CitationStirling and Dalton, 1979; CitationPrior and Dalton, 1985; CitationLontoh et al., 1999; CitationLontoh et al., 2000a). At a low concentration (100 μ M), PA causes almost no inhibition of pMMO, but causes greater than 90% inhibition of the sMMO (CitationLontoh et al., 2000a). Water samples (2.5 ml) were placed in 10-ml serum vials, and 1 ml of PA (100 μ M final concentration) was added to each. Vials were sealed and incubated for 20 min at room temperature. The caps were removed to vent the headspace, coumarin was added (500 μ M), and the solutions were assayed for fluorescence after a 10-min incubation period.
Groundwater samples were also evaluated for enzyme activity in the presence of methane, which should serve as a competitive inhibitor of the coumarin oxidation by sMMO. The fluorescence of oxidized coumarin in a sample saturated with methane should be significantly less than a sample that contains low, noncompetitive concentrations of methane. Whole-water cell suspensions were filtered onto Supor filters and evaluated with the coumarin assay as previously described. Following the initial fluorescence measurement (eluate) without methane, the filter was sealed in the serum vial, sparged with a 1:1 ultra-high-purity methane/air mixture (v/v) (Specialty Chemical Products, South Houston, Texas) for 30 s and incubated at room temperature for 15 min. After the incubation period, the caps were removed, the filters washed with NMS medium, and the eluate assayed for fluorescence. The samples were then sparged with compressed air for 3 to 5 min to remove residual methane. Following incubation in the presence of additional coumarin (5 mM) and without methane, the cell samples were analyzed once more for fluorescence to determine if enzyme activity against coumarin had recovered.
Unfiltered whole water samples were also evaluated with the coumarin assay in the presence of 20 mM sodium formate. Cells that are cometabolizing TCE often require a reductant, as the cometabolic reaction is often reductant limited. Formate, a readily reducible substrate and electron donor, has been shown to enhance the rates of both methane and TCE oxidation and, as such, should increase sMMO activity (CitationLontoh and Semrau, 1998; CitationLontoh et al., 1999; CitationAlvaraz-Cohen and McCarty, 1991). This increase in sMMO activity should be reflected in an elevated fluorescence response with the coumarin assay, in the presence of formate. Groundwater samples (2.5 ml) were added to 10-ml serum vials containing 5 mM coumarin and 20 mM formate and assayed for fluorescence after incubation for 10 min at room temperature.
Nonmethanotrophic controls consisted of microorganisms that do not express the sMMO gene being exposed to coumarin and assayed for fluorescence. Pseudomonas putida F1 and Burkholderia cepacia G4, which express toluene oxygenases, were grown under oxygenase-induced conditions, and served as control organisms for the enzyme probe. These organisms were exposed to coumarin as described above.
Naphthalene Oxidation Assay
A modified version of the naphthalene assay (CitationChu and Alvarez-Cohen, 1998; CitationBrusseau et al., 1990) was used to verify the enzyme probe results. This assay is based on the assumption that only sMMO can oxidize naphthalene to 1- or 2-naphthol in methane-oxidizing bacteria. The primary modification from the previously reported methods was that for this application, the assay was applied to the natural population of cells from groundwater samples rather than to laboratory cultures. A second modification was that the appearance of the product was not measured over time, but rather the change in color was qualitatively observed in solution.
Briefly, the assay was conducted by adding 1 ml of groundwater, along with 20 mM formate and 1 naphthalene crystal to a 10-ml serum vial sealed with a Teflon-lined rubber septum (Wheaton). The mix was incubated at 35°C on a shaker at 160 rpm for 1 h before the addition of 100 μ l of freshly prepared 0.2% (w/v) tetrazotized o-dianisidine. Soluble MMO activity was indicated by the appearance of a purple color in the mixture. Reactions containing only cells and formate were used as negative controls.
Extraction of Total DNA from Groundwater
DNA was isolated from groundwater using the DNEasy tissue isolation kit as described by the manufacturer (Qiagen, Valencia, CA, USA). These methods consistently yielded high-quality bacterial DNA, based on agarose gel band analysis.
Polymerase chain reaction (PCR) amplifications were performed as described previously (CitationMcDonald et al., 1995) using the mmoX f882 and r1403 primers, which amplify an approximate 564-bp fragment from M. capsulatus Bath (CitationMcDonald et al., 1995). DNA from Methylosinus trichosporium OB3b was used in PCR as a positive control to verify that mmoX primers targeted organisms in the samples harboring the gene for the soluble form of the enzyme; DNA extracted from Pseudomonas putida F1 and Burkholderia cepacia G4 was used as a nonmethanotrophic control for the mmoX-targeted amplifications.
DNA Analysis
To confirm that the PCR-amplified DNA was consistent with known mmoX sequences, DNA sequencing of the mmoX gene fragment, taken directly from the agarose gels, was carried out on both strands using the ABI Prism BigDye terminators sequencing kit (PE Applied Biosystems, Foster City, CA). The sequencing reactions and analyses were performed by the Idaho State University Molecular Core Research Facility (MCRF) using an Applied Biosystems 3100 Sequencer. The mmoX sequences were aligned with translated mmoX sequences obtained from the GenBank database, BLAST nucleotide-to-nucleotide match (http://www.ncbi.nlm.nih.gov/BLAST/).
Enrichment Cultures
TAN groundwater samples were serially diluted 1:100 in 50 ml NMS medium and added to 100-ml glass serum vials. The vials were sealed, and the headspace was modified to create a 1:1 methane-air atmosphere. The vials were incubated at 30°C for up to 21 days. After the incubation period, aliquots of enrichment medium were then spread-plated onto semisolid NMS medium (1.5% agarose w/v) and incubated under 1:1 methane:air atmosphere for 1 week at 30°C. Three successive transfers onto fresh minimal media plates were performed. The final transfer of colonies was made into sterile NMS liquid medium in 10-ml serum vials that were subsequently incubated for 5 to 7 days with a 1:1 methane:air headspace. The turbidity of the primary and secondary liquid enrichments was scored as positive or negative and cell colony morphologies were recorded. Isolates were stored in 25% glycerol stocks at −20°C for future analysis. Sample enrichments and isolates were tested for sMMO activity using the coumarin and naphthalene oxidation assays, and PCR amplification of mmoX genes was performed as described above. To identify the isolates, DNA was extracted from each isolate, PCR amplified using universal 16S rDNA primers, purified, and sequenced. Subsequently, sequences were aligned with other 16S rDNA sequences obtained by BLAST searches, as described above.
Total Cell Counts
Total cells were determined after staining with 4,6-diamidino-2-phenylindole (DAPI) for 10 min at room temperature in the dark (final concentration 3 μ g/ml). Briefly, water samples (10 ml) were filtered onto 0.2-μ m black polycarbonate filters under low vacuum and stained with DAPI (250 μ l onto filter surface). Samples were then washed twice with 1 ml of nanopure water and placed on glass slides. Filters were immediately covered with immersion oil and a cover slip. The stock solution of DAPI (50 μ g/mL) was prepared in sterile, nanopure water and stored in the dark at 4°C until use. Total cells were visualized with a Wide UV filter set (Exciter filter BP 340-380, dichroic mirror DM 400, and barrier filter BA 445).
RESULTS
Groundwater Chemistry Results
All locations sampled during the 2-year investigation were aerobic, with DO ranging from 2 to 10 mg/L. Sulfate was present near background levels of 30 to 40 mg/L in all wells sampled. Nitrate was present in all wells, with concentrations ranging from 0.4 to 1.4 mg/L. Ferrous iron was detected, but was present at concentrations less than 0.10 mg/L at all but one sampling location. Although methane was detected at low levels (in the nM range), it is not attributed to local methanogenic conditions based both on the low levels and on the other redox parameters. These multiple lines of evidence show that redox conditions in the fringe of the plume are aerobic, which is consistent with the low levels of dissolved organic carbon observed in the system (< 10 mg/L). The dissolved organic carbon levels were nominally consistent with the concentrations attributable to natural humic and fulvic acids (CitationFreeze and Cherry, 1979). Copper, which has been shown in laboratory studies to inhibit sMMO activity when present above approximately 16 μ g/L (CitationTsien et al, 1989), was sampled extensively during the first year of this investigation. It was measured above its detection limit of 3.2 μ g/L in only 1 of the 51 locations, at a concentration of 14 μ g/L.
TCE concentrations ranged from non-detect to a maximum of 240 μ g/L in well TAN-51. Wells TAN-56 and -58 did not have definitive detections of any contaminants, while well TAN-57 had TCE and PCE concentrations above detection but below drinking water standards. cis-DCE was detected at low levels, with the maximum concentration of 9.1 μ g/L being far below the drinking water standard (maximum contaminant level) of 70 μ g/L. Tritium was measured above detection in wells TAN-51, -52, -54, and -55. Representative contaminant results are shown in and are also presented in .
FIGURE 2 TCE and tritium profiles from TAN-51 and TAN-55.
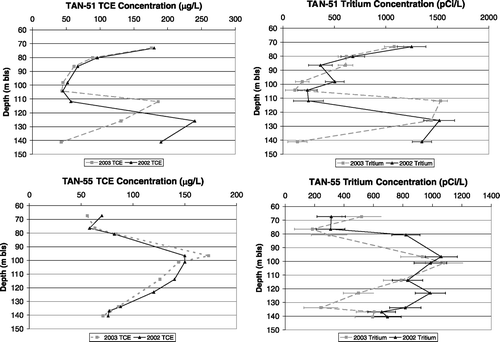
TABLE 2 Results of Groundwater Analysis for the Enzyme Probe Coumarin, Naphthalene, and Controls
Collection of samples from the wells equipped with FLUTe systems provided the ability to assess three-dimensional plume anatomy in the aerobic fringe. In general, significant vertical variations were observed in contaminant profiles within individual wells. In addition, trends observed were not consistent among wells. Representative illustrations are provided in , which shows TCE and tritium profiles from two separate monitoring wells, TAN-51 and TAN-55. Well TAN-51 showed depressed concentrations of both TCE and tritium in the middle 40 m of the aquifer, with zones of higher concentration detected in the upper and lower portions. In contrast, TAN-55 showed higher concentrations in the middle portions, with lower concentration horizons found in the upper and lower portions of the aquifer. Despite these disparate trends between wells, the similar vertical concentration distribution of both TCE and tritium within a given well suggests that the vertical concentration trends are due to local-scale heterogeneities in permeability and other physical aquifer parameters, as opposed to large-scale preferential flow or differences in degradation kinetics and chemical processes. This is further supported by the fact that the variation in ratios of TCE:PCE within an individual well is small (data not shown).
CitationChapelle et al. (1997) suggests that submersible pumps can generate hydrogen during operation, which can skew the measured concentrations of hydrogen as well as other dissolved gases. The fact that the dissolved gas results from wells sampled using the submersible pumps were 1 to 3 orders of magnitude greater than the equilibrium concentrations suggests that this was the case at TAN. Therefore, the investigation results considered herein include only dissolved gas data from wells equipped with FLUTe systems.
In spite of the plume's aerobic conditions, low levels of methane and carbon monoxide appeared widespread both inside and outside the TAN TCE plume. Although atmospheric contamination of the dissolved gas samples was unlikely because of (1) the closed-loop system used to collect the dissolved gas samples and (2) the short period of time between sample collection and analysis, measured concentrations were compared to concentrations that would be expected if the sample were in equilibrium with the atmosphere. The extent to which each gas partitions into the aqueous phase was determined using Henry's Law (CitationManahan, 1994; CitationSander, 1999), with corrections for elevation of the site (4700 feet above mean sea level) and temperature (12°C) of the groundwater. For methane, the atmospheric equilibrium concentration was determined to be 2.2 nM, whereas the atmospheric equilibrium concentration for carbon monoxide was 0.11 nM.
Methane was detected at all sampling locations, with concentrations ranging from 0.2 to 3.9 nM during the 2-year investigation. Approximately 22% of the methane concentrations were greater than or equal to the atmospheric equilibrium concentration. Carbon monoxide was also detected at all sampling locations during the 2-year investigation, with concentrations ranging from 0.02 to 7.5 nm. However, in contrast to the methane data, approximately 80% of the samples were greater than or equal to the equilibrium value. Based on comparisons of dissolved gas measurements taken several weeks apart (data not shown), even methane concentrations less than or equal to the calculated equilibrium values are unlikely to be due solely to contamination. These comparisons showed that equilibration between groundwater collected and stored in glass bulbs and atmospheric gases can take several weeks, whereas analyses were all completed within 24 to 48 h.
Enzyme Probe and Molecular Results
In general, bacterial abundance as measured by DAPI in the samples analyzed over the entire sampling period (2002 to 2003) and at all depths, was similar with an average of 4.8 × 105 ± 1.4 × 105 cells/ml. The results of the coumarin assay performed on TAN groundwater are shown in . All of the TAN groundwater samples tested using the coumarin oxidation assay yielded fluorescence. Fluorescence in all samples was between 14 and 21 intensity units per minute (fluorescence intensity) in year 1 (2002) and was significantly more variable (p < .05), 4 to 23 intensity units per minute, in year 2 (2003). Nonmethanotrophic controls not expressing sMMO (Pseudomonas putida F1 and Burkholderia cepacia G4) did not yield fluorescence when exposed to coumarin (). It was confirmed that the product derived from the transformation of coumarin exhibited a fluorescence spectrum similar to that of coumarin when reacted with M. trichosporium OB3b and that of 7-hydroxycoumarin (CitationMiller et al., 2002), the fluorescence product that is generated in a successful coumarin oxidation assay.
All of the groundwater samples tested in both years, when incubated with naphthalene for at least 1 h, then exposed to tetrazotized o-dianisidine, turned a deep purple color, further indicating the presence of active sMMO in the samples (). No color was formed in any of the controls not expressing sMMO.
The combined presence of coumarin and formate caused an increase (average 53% ± 2.1%) relative to the activity observed when cells were exposed to coumarin alone (). In all of the groundwater samples tested, development of fluorescence was strongly inhibited when samples were preincubated with 5 mM phenylacetylene ().
All of the groundwater samples tested in the presence of methane and coumarin, showed 96% to 98% reduction in the fluorescence signal, the basis of the coumarin oxidation assay (). However, once the methane was removed, sMMO activity directed toward coumarin oxidation was recovered to an average of 80% ± 3.4% of the original value ().
TABLE 3 Fluorescent Signals in Response to the Methane Reversible Inhibitor Control Study
The PCR primers for the mmoX gene amplified sequences from all of the groundwater samples (direct extraction), yielding the predicted PCR product of 564 bp, but no products were detected when sMMO-negative strains were examined (data not shown). The second year of testing (2003) revealed similar results, with the predicted PCR product being amplified from DNA extracted from cells present in all of the groundwater samples that were tested. The primers also amplified 564-bp products from all of the organisms enriched with methane in laboratory experiments (data not shown). These results confirm that the sMMO gene was present in all groundwater and enrichment samples analyzed.
In all reactions containing groundwater samples, an mmoX-amplification product of correct size was generated, which is consistent with previous findings (CitationNewby et al., 2004; CitationErwin et al., 2005). PCR products were sequenced from the enrichments and matched (BLAST nucleotide-to-nucleotide match), based on 16S primers. The level of similarity between the amplified fragments and fragments obtained from the NCBI database, ranged from 97% to 99% with known sMMO-producing organisms. The alignments revealed clone identity matches with Methylocystis, Methylosinus, and uncultured methanotrophs. Again, these results supplement the enzyme probe data in that they demonstrate that methanotrophs were detected throughout the TAN plume and the SRPA in general.
DISCUSSION
The tracer-corrected method for TCE degradation rate estimation was applied to these new monitoring data, including the vertically discrete sampling data from the FLUTe systems. The results of this analysis are presented in , which shows the ratios of TCE:tritium with distance from the upgradient edge of the aerobic fringe (represented by well TAN-39 on ). Tritium concentrations were corrected for its 12.3-year half-life based on the estimated groundwater velocity used in the analysis, as in CitationSorenson et al. (2000). This plot confirms the previous hypothesis in that it shows a negative slope, indicating TCE attenuation relative to tritium is occurring. In this case, the updated half-life estimate for TCE degradation was 13 years, with a range 12 to 15 years based on a 95% confidence interval on the slope of the regression. The addition of the vertically discrete data to this analysis not only supports the previous conclusions of CitationSorenson et al. (2000), but also actually narrows the 95% confidence interval of the rate estimate (previously estimated at 11 to 17 years), demonstrating that the apparent degradation was not merely an artifact of inadequate characterization of the contaminant distribution.
FIGURE 3 Application of the tracer-corrected method (CitationSorenson et al., 2000) for TCE (a) and c-DCE (b).
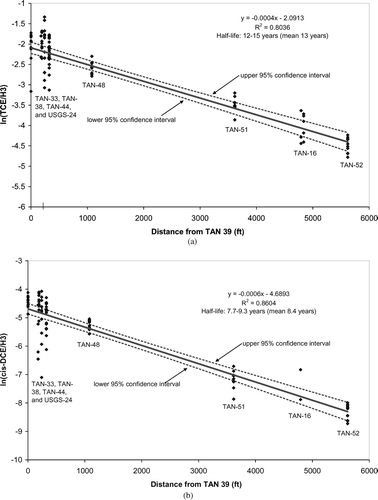
The initial application of the tracer-corrected method included estimates for TCE attenuation only (CitationSorenson et al., 2000). However, given that cis-DCE is also susceptible to aerobic cometabolism by the same microbial populations that cometabolize TCE (CitationAlvarez-Cohen and Speitel, 2001), the method was also applied to cis-DCE data from the aerobic fringe of the plume. The results, shown in , show that cis-DCE is also attenuating relative to tritium, with a half-life of 8.4 years, and a range of 7.7 to 9.3 years based on a 95% confidence interval. This half-life is significantly shorter than the TCE half-life, which is consistent with the fact that cis-DCE is more rapidly cometabolized than TCE by the sMMO enzyme system (CitationAlvarez-Cohen and Speitel, 2001). In addition, cis-DCE can undergo direct oxidation (CitationVogel et al., 1987; CitationBradley and Chapelle, 1998; CitationBradley and Chapelle, 2000), which might contribute to the observed degradation.
Groundwater chemistry data were consistent with the conditions required for aerobic cometabolism. The overall assessment of redox conditions indicated that the entire distal zone was aerobic. Also, the widespread detections of low levels of methane indicate the presence of a primary substrate and inducer of sMMO. The source of the subsurface methane is unknown, but is speculated to be from a deep, hydrogen-driven biosphere (e.g., CitationPedersen, 1997). For example, CitationStevens et al. (1993) found significant activity from iron-reducers, sulfate-reducers, and methanogens in samples collected from two anaerobic basaltic aquifers in southeast Washington. CitationStevens and McKinley (1995) studied these and other ecosystems in the Columbia River Basalt Group and found that autotrophic methanogenesis was the dominant metabolism.
Although the presence of a deep hydrogen-driven biosphere has not been confirmed at TAN, the conditions in the SRPA beneath TAN are similar to the conditions in the studies referenced above. The concentrations of methane measured during this and previous investigations at TAN were consistent with results obtained by CitationNewby et al. (2004), who performed investigations throughout the SRPA (CitationNewby et al., 2004). They also reported that stable carbon isotope analyses of methane and dissolved inorganic carbon in the SRPA from four wells outside of the TAN plume indicate that the methane is likely of microbial origin and is actively undergoing oxidation. Although the stable carbon isotope assays were not routinely performed during this investigation, screening results from samples collected from three intervals in well TAN-51 were consistent with these isotopic signatures (data not shown).
Carbon monoxide was included in the monitoring program because it can be a potentially important byproduct of cometabolic oxidation of TCE in the pH range observed at TAN (CitationVogel and McCarty, 1987; CitationFogel et al., 1986; CitationLontoh et al., 2000b). Although the source of the carbon monoxide is uncertain, no other known sources (e.g., portable generators or running vehicles) that could have contributed to the observed concentrations were present. Carbon monoxide was also detected during previous sampling activities (data not shown). Its presence is consistent with cometabolic oxidation of TCE by indigenous methanotrophs.
The results discussed above provide several lines of indirect evidence for aerobic TCE attenuation at TAN. They also document the groundwater chemistry conditions under which the degradation is occurring, and quantify the in situ degradation rate. The most compelling evidence for intrinsic cometabolism, however, is the enzyme probe and molecular results. Based on previous studies (CitationMiller et al., 2002), the positive probe responses observed when TAN groundwater was exposed to coumarin is evidence of active sMMO enzymes within the aerobic portion of the plume. Control studies including a known sMMO inhibitor, phenylacetylene (CitationLontoh et al., 2000a), and a competitive reversible inhibitor, methane, inhibited coumarin transformation in all samples tested in both years. These findings imply that the transformation of coumarin to a fluorescent product is dependent on the sMMO activity present in the samples. The presence of this activity in groundwater from the TAN site strongly supports the hypothesis of CitationSorenson et al. (2000) that TCE is undergoing natural transformation in situ via aerobic cometabolic oxidation. Additional studies with probes targeting aromatic oxygenases confirm the presence of cometabolic activity in this aquifer (CitationLee et al., 2007); these studies also provided direct evidence of TCE degradation concomitant with probe activity by indigenous microbial populations from TAN.
In the present study, we enriched for and isolated groundwater bacteria to provide support to our enzyme activity probe analysis. Isolates were grown with methane as their sole carbon source and exhibited sMMO activity based on both coumarin and naphthalene assays. In addition, DNA was amplified using primers specific for mmoX genes, and the resulting amplicons exhibited high sequence homology (97% to 99%) with genes from sMMO-containing organisms. Similar results were obtained using these primers on groundwater samples both inside and outside the plume. The observation of sMMO activity in, and successful enrichment of methanotrophs from, areas outside of the TCE plume indicates that methanotrophic bacteria are members of the indigenous microbial community in the SRPA, and that their activity is not dependent upon the contamination in the plume. This is further supported by the fact that similar enzyme activity was observed at discrete sampling locations that had different contaminant concentrations. Also, this study supports recent findings of methane-oxidizers in numerous wells across the SRPA (CitationNewby et al., 2004; CitationErwin et al., 2005).
The data presented represent two discrete sampling times (2002 and 2003) and clearly show that the appropriate organisms for methanotrophic TCE cometabolism (a) were present, (b) had the enzyme potential to cometabolize TCE, and (c) had actively functioning enzymes known to degrade TCE. This investigation documents what is believed to be the first reported case of intrinsic aerobic TCE and cis-DCE cometabolism by indigenous methanotrophs. It is also significant in the regulatory context as it supported a first-of-its-kind Record of Decision Amendment changing much of the site remediation strategy from pump and treat to monitored natural attenuation of TCE under aerobic conditions.
From a broader perspective, these results show the efficacy of the approach used herein, namely combining groundwater chemistry with activity-dependent enzyme probes and molecular methods for documenting the occurrence of intrinsic cometabolism at a chlorinated solvent contaminated site. Most importantly, these results potentially contradict conventional thinking that intrinsic degradation of TCE in groundwater cannot be important under aerobic conditions. It remains to be seen whether the aerobic cometabolism of TCE (or cis-DCE or VC) documented at the TAN site is common at other sites.
ACKNOWLEDGEMENTS
The authors would like to thank the many people that were integral members of the project team that performed this investigation. They would also like to extend our gratitude to Frank Roberto for his role in the development of the coumarin probe and Brady Lee for use of analytical equipment and facilities at the INL Research Center. Riena Carroll, Dana Dettmers, Patrick Lebow, Kevin Harris, Al Cram, and Kory Edelmayer provided sampling and field test kit support. Cathy Rae and Mark Delwiche performed fixed laboratory analysis for chloroethene and dissolved gas samples. Project management and technical oversight were provided by Lance Peterson, Joe Rothermel, and Lee Nelson. This work was performed under United States Department of Energy Idaho Operations Office Contract Number DE-AC07-99ID13727.
Notes
a Intensity units per minute. Values represent the average of three replicates.
b Phenylacetylene.
a Intensity units per minute.
REFERENCES
- Alvarez-Cohen , L. and McCarty , P. L. 1991 . Effects of toxicity, aeration, and reductant supply on trichloroethylene transformation by a mixed methanotropic culture . Appl. Environ. Microbiol. , 57 : 228 – 235 .
- Alvarez-Cohen , L. and Speitel , G. E. Jr. 2001 . Kinetics of aerobic cometabolism of chlorinated solvents . Biodegradation , 2 : 105 – 126 .
- Anthony , C. 1982 . The biochemistry of methylotrophs , London : Academic Press .
- Auman , A. J. , Stolyar , S. , Costello , A. and Lidstrom , M. E. 2000 . Molecular characterization of methanotrophic isolates from freshwater lake sediments . Appl. Environ. Microbiol. , 66 : 5259 – 5266 .
- Aziz , C. E. , Georgiou , G. and Speitel , G. E. Jr. 1999 . Cometabolism of chlorinated solvents and binary chlorinated solvent mixtures using M. trichosporium OB3b PP358 . Biotechnol. Bioengin. , 65 : 100 – 107 .
- Bowman , J. , Jimenez , L. , Rosario , I. , Hazen , T. and Sayler , G. 1993 . Characterization of the methanotrophic bacterial community present in a trichloroethylene-contaminated subsurface groundwater site . Appl. Environ. Microbiol. , 59 : 2380 – 2387 .
- Bradley , P. M. and Chapelle , F. H. 2000 . Aerobic microbial mineralization of dichloroethene as sole carbon substrate . Environ. Sci. Technol , 34 : 221 – 223 .
- Bradley , P. M. and Chapelle , F. H. 1998 . Humic acids as electron acceptors for anaerobic microbial oxidation of vinyl chloride and dichloroethene . Appl. Environ. Microbiol. , 64 : 3102 – 3105 .
- Brusseau , G. A. , Tsien , H.-C. , Hanson , R. S. and Wackett , L. P. 1990 . Optimization of trichloroethylene oxidation by methanotrophs and the use of a colorimetric assay to detect soluble methane monooxygenase activity . Biodegradation , 1 : 19 – 29 .
- Butler , E. and Hayes , K. 1999 . Kinetics of the transformation of trichloroethylene and tetrachloroethylene by iron sulfide . Environ. Sci. Technol. , 33 : 2021 – 2027 .
- Chapelle , F. H. , Vroblesky , D. A. , Woodward , J. C. and Lovley , D. R. 1997 . Practical considerations for measuring hydrogen concentrations in groundwater . Environ. Sci. Technol. , 31 : 2873 – 2877 .
- Cherry , J. A. . Conceptual models for chlorinated solvent plumes and their relevance to intrinsic remediation . Proceedings of the Symposium on Natural Attenuation of Chlorinated Organics in Groundwater . pp. 31 – 32 . Washington, DC : Office of Research and Development, U.S. Environmental Protection Agency . EPA/540/R-97/504
- Cherry , J. A. , Parker , B. L. and Keller , C. 2007 . A new depth-discrete multilevel monitoring approach for fractured rock . Groundwater Monitoring and Remediation , 2 : 57 – 70 .
- Chu , K.-H. and Alvarez-Cohen , L. 1998 . Effect of nitrogen source on growth and trichloroethylene degradation by methane-oxidizing bacteria . Appl. Environ. Microbiol. , 64 : 3451 – 3457 .
- Church , P. E. and Granato , G. E. 1996 . Bias in ground-water data caused by well-bore flow in long-screen wells . Groundwater , 34 : 262 – 273 .
- DiSpirito , A. , Gulledge , J. , Murrell , J. C. , Shiemke , A. K. , Lidstrom , M. E. and Krema , C. L. 1992 . Trichloroethylene oxidation by the membrane-associated methane monooxygenase in type I, type II, and type x methanotrophs . Biodegradation , 2 : 151 – 164 .
- EPA . 1999 . Use of Monitored Natural Attenuation at Superfund, RCRA Corrective Action, and Underground Storage Tanks , Washington, DC : Office of Solid Waste and Emergency Response, U.S. Environmental Protection Agency . Directive 9200.4–17P
- Erwin , D. P. , Erickson , I. K. , Delwiche , M. E. , Colwell , F. S. , Strap , J. L. and Crawford , R. L. 2005 . Diversity of oxygenase genes from methane- and ammonia-oxidizing bacteria in the eastern snake river plain aquifer . Appl. Environ. Microbiol. , 71 : 2016 – 2025 .
- Fogel , M. M. , Taddeo , A. R. and Fogel , S. 1986 . Biodegradation of chlorinated ethenes by a methane-utilizing mixed culture . Appl. Environ. Microbiol. , 51 : 720 – 724 .
- Freeze , R. A. and Cherry , J. A. 1979 . Groundwater , 86 Englewood Cliffs, NJ : Prentice Hall .
- Freedman , D. L. and Gossett , J. M. 1989 . Biological reductive dechlorination of tetrachloroethylene and trichloroethylene to ethylene under methanogenic conditions . Appl. Environ. Microbiol. , 55 : 2144 – 2151 .
- Hopkins , G. D. and McCarty , P. L. 1995 . Field evaluation of in situ aerobic cometabolism of trichloroethylene and three dichloroethylene isomers using phenol and toluene as the primary substrates . Environ. Sci. Technol. , 29 : 1628 – 1637 .
- Kauffman , M. E. , Keener , W. K. , Clingenpeel , S. R. , Watwood , M. E. , Lehman , M. and Reed , D. 2003 . Activity-dependent fluorescent labeling of bacteria that degrade toluene via 3-methylcatechol . J. Microb. Methods , 55 : 801 – 805 .
- Keener , W. K. and Watwood , M. E. 1997 . “ Probes for enzyme-dependent fluorescent labeling of bacteria degrading trichloroethylene ” . In In Situ and On-Site Bioremediation , Edited by: Alleman , B. C. and Leeson , A. Vol. 5 , 327 – 332 . Columbus, OH : Battelle Press . Proceedings of the Fourth International In Situ and On-site Bioremediation Symposium
- Keener , W. K. , Watwood , M. E. and Apel , W. A. 1998 . Activity-dependent fluorescent labeling of bacteria that degrade toluene via toluene 2,3-dioxygenase . Appl. Microbiol. Biotechnol. , 49 : 455 – 462 .
- Keener , W. K. , Watwood , M. E. , Schaller , K. D. , Walton , M. R. , Partin , J. K. , Smith , W. A. and Clingenpeel , S. R. 2001 . Use of selective inhibitors and chromogenic substrates to differentiate bacteria based on toluene oxygenase activity . J. Microb. Methods , 46 : 171 – 185 .
- Lee , M. H. , Clingenpeel , S. C. , Leiser , O. P. , Wymore , R. A. , Sorenson , K. S. Jr. and Watwood , M. E. 2007 . Activity-dependent labeling of oxygenase enzymes in a trichloroethylene-contaminated groundwater site . Environ. Poll. , in press
- Lee , W. and Batchelor , B. 2002 . Abiotic reductive dechlorination of chlorinated ethylenes by iron-bearing soil minerals. 1. Pyrite and magnetite . Environ. Sci. Technol. , 36 : 5147 – 5154 .
- Lontoh , S. , DiSpirito , A. A. , Krema , C. L. , Whittaker , M. R. , Hooper , A. B. and Semrau , J. D. 2000a . Differential inhibition in vivo of ammonia monooxygenase, soluble methane monooxygenase and membrane-associated monooxygenase by phenylacetylene . Environ. Microbiol. , 2 : 485 – 494 .
- Lontoh , S. , DiSpirito , A. A. and Semrau , J. D. 1999 . Dichloromethane and trichloroethylene inhibition of methane oxidation by the membrane-associated methane monooxygenase of methylosinus trichosporium OB3b . Arch. Microbiol. , 171 : 301 – 308 .
- Lontoh , S. and Semrau , J. D. 1998 . Methane and trichloroethylene degradation by methylosinus trichosporium OB3b expressing particulate methane monoxygenase . Appl. Environ. Microbiol. , 64 : 1106 – 1114 .
- Lontoh , S. , Zahn , J. A. , DiSipirito , A. A. and Semrau , J. D. 2000b . Identification of intermediates of in vivo trichloroethylene oxidation by the membrane-associated methane monooxygenase . FEMS Microbiol. Lett. , 186 : 109 – 113 .
- Manahan , S. E. 1994 . Environmental chemistry, , 6th ed. , Boca Raton, FL : Lewis Publishers .
- McCarty , P. L. . Biotic and abiotic transformations of chlorinated solvents in groundwater . Symposium on Natural Attenuation of Chlorinated Organics in Groundwater . pp. 7 – 11 . Washington, DC : Office of Research and Development, U.S. Environmental Protection Agency . EPA/540/R-97/504
- McDonald , I. R. , Kenna , E. M. and Murrell , J. C. 1995 . Detection of methanotrophic bacteria in environmental samples with the PCR . Appl. Environ. Microbiol. , 61 : 116 – 121 .
- Miguez , C. B. , Bourque , D. , Sealy , J. A. , Greer , C. W. and Groleau , D. 1997 . Detection and isolation of methanotrophic bacteria possessing soluble methane monooxygenase (sMMO) genes using the polymerase chain reaction (PCR) . Microb. Ecol. , 33 : 21 – 31 .
- Miller , A. R. , Roberto , F. , Keener , W. and Watwood , M. E. 2002 . Development of a fluorescence-based assay for soluble methane monooxygenase . Appl. Microbiol. Biotechnol. , 58 : 183 – 188 .
- Newby , D. T. , Reed , D. W. , Petzke , L. M. , Igoe , A. L. , Delwiche , M. E. , Roberto , F. F. , McKinley , J. P. , Whiticar , M. J. and Colwell , F. S. 2004 . Diversity of methanotroph communities in a basalt aquifer . FEMS Microbiol. Ecol. , 48 : 333 – 344 .
- Oldenhuis , R. , Oedzes , J. Y. , Waarde , J. J. V. D. and Janssen , D. B. 1991 . Kinetics of chlorinated hydrocarbon degradation by methylosinus trichosporium OB3b and toxicity of trichloroethylene . Appl. Environ. Microbiol. , 57 : 7 – 14 .
- Oldenhuis , R. R. L. , Vink , J. M. , Vink , M. , Janssen , D. B. and Witholt , B. 1989 . Degradation of chlorinated aliphatic hydrocarbons by Methylosinus trichosporium OB3b expressing soluble methane monooxygenase . Appl. Environ. Microbiol. , 55 : 2819 – 2826 .
- Pedersen , K. 1997 . Microbial life in deep granitic rock . FEMS Microbiol. Rev. , 20 : 399 – 414 .
- Prior , S. D. and Dalton , H. 1985 . Acetylene as a suicide substrate and active site probe for methane monooxygenase from methylococcus capsulatus (Bath) . FEMS Microbiol. Lett. , 29 : 105 – 109 .
- Puls , R. W. and Barcelona , M. J. 1996 . Low-Flow (Minimal Drawdown) Groundwater Sampling Procedures , Washington, DC : Office of Research and Development and Office of Solid Waste and Emergency Response, U.S. Environmental Protection Agency . EPA/540/S-95/504
- Sander , R. 1999 . Compilation of Henry's Law Constants for Inorganic and Organic Species of Potential Importance in Environmental Chemistry , Mainz, , Germany : Air Chemistry Department, Max-Planck Institute of Chemistry . http://www.mpch-mainz.mpg.de/∼sander/res/henry.html
- Semprini , L. , Roberts , P. V. , Hopkins , G. D. and McCarty , P. L. 1990 . A field evaluation of in-situ biodegradation of chlorinated ethenes: Part 2, Results of biostimulation and biotransformation experiments . Ground Water , 28 : 715 – 727 .
- Semrau , J. D. , Chistoserdov , A. , Lebron , J. , Costello , A. , Davagnino , J. , Kenna , E. , Holmes , A. J. , Finch , R. , Murrell , J. C. and Lidstrom , M. E. 1995 . Particulate methane monooxygenase genes in methanotrophs . J. Bacteriol. , 177 : 3071 – 3079 .
- Sewell , G. W. and Gibson , S. A. . Microbial Ecology of Adaptation and Response . Symposium on Natural Attenuation of Chlorinated Organics in Groundwater . pp. 16 – 18 . Washington, DC : Office of Research and Development, U.S. Environmental Protection Agency . EPA/540/R-97/504
- Sorenson , K. S. 2000 . Intrinsic and enhanced in situ biodegradation of trichloroethene in a deep, fractured basalt aquifer , PhD Dissertation University of Idaho .
- Sorenson , K. S. , Peterson , L. N. , Ely , R. L. and Hinchee , R. E. 2000 . An evaluation of aerobic trichloroethene attenuation using first-order rate estimation . Bioremediat. J. , 4 : 337 – 358 .
- Stevens , T. O. and McKinley , J. P. 1995 . Lithoautotrophic microbial ecosystems in deep basalt aquifers . Science , 270 : 450 – 454 .
- Stevens , T. O. , McKinley , J. P. and Fredrickson , J. K. 1993 . Bacteria associated with deep, alkaline, anaerobic groundwaters in Southeast Washington . Microb. Ecol. , 25 : 35 – 50 .
- Stirling , D. L. and Dalton , H. 1979 . Properties of the methane mono-oxygenase from extracts of methylosinus trichosporium OB3b and evidence for its similarity to the enzyme from methylococcus capsulatus . Eur. J. Biochem. , 96 : 205 – 212 .
- Tsien , H.-C. , Brusseau , G. A. , Hanson , R. S. and Wackett , L. P. 1989 . Biodegradation of trichloroethylene by Methylosinus trichosporium OB3b . Appl. Environ. Microbiol. , 55 : 3155 – 3161 .
- Vogel , T. M. , Criddle , C. S. and McCarty , P. L. 1987 . Transformations of halogenated aliphatic compounds . Environ. Sci. Technol. , 21 : 722 – 736 .
- Whittenbury , R. , Phillips , K. C. and Wilkinson , J. K. 1970 . Enrichment isolation, and some properties of methane-utilizing bacteria . J. Gen. Microbiol. , 59 : 2771 – 2776 .
- Williams , J. H. and Conger , R. W. 1990 . Preliminary delineation of contaminated water-bearing fractures intersected by open-hole bedrock wells . Ground Water Monit. Remediat. , 10 : 118 – 126 .
- Wymore , R. A. , Sorenson , K. S. Jr. and Steven Porter , P. 2007 . Evaluation of multi-level sampling systems for assessment of contaminant distribution in a deep, fractured basalt aquifer . Ground Water Monit. Remediat. , Submitted