Abstract
Strata of Jurassic age occur extensively across onshore Australia, but they are predominantly of non-marine origin. Marine Jurassic strata have only limited onshore exposure in northwestern and central-western Australia, with thick marine sequences lying offshore on the North West Shelf. The richest petroleum province in Australia is located at the shelf's southern end, where the Dingo Claystone represents an important source rock for oil and gas. By and large, non-marine deposits, including economic coals, are distributed in the eastern states. Jurassic stage boundaries, in the main, are poorly constrained with respect to the Australian sedimentary succession. New work on microfossils, plants, fish, and zircon dating is providing a basis for improved correlation across Australian basins, with overseas successions, and recent international IUGS geologic timescales.
Introduction
Jurassic deposits are extensive in several Australian basins (Figs. , ). In the west, they are located in the onshore Bonaparte, Canning, Carnarvon, and Perth basins and in the offshore Westralian Superbasin, which comprises the Northern Bonaparte, Browse, Roebuck, and Northern Carnarvon basins. In eastern and central-eastern Australia, they are embraced by the Laura, Carpentaria, Eromanga, Surat, Mulgildie, Clarence-Moreton, and Nambour basins. In South Australia, they are represented in the Polda and Bight basins, and the isolated deposits of the Rhaeto-Liassic Leigh Creek Coal Measures. In southeastern Australia, the oldest deposits of the Otway and Gippsland basins are probably of latest Jurassic age (Burger & Shafik Citation1996). These successions were mostly deposited in fluvial sedimentary systems.
Fig. 1 Onshore and offshore Australian basins containing Jurassic sediments (prepared by Geological Survey of Queensland, courtesy of the Information Development and Analysis Services Group, Geoscience Australia).
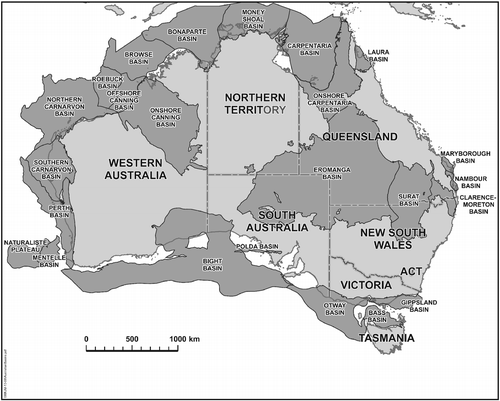
Fig. 2 Contd.
Sedimentation was not entirely continuous through the period, as widespread hiatuses occurred at its beginning (largely during the Hettangian) and during the latest Middle–early Late Jurassic. In eastern Australia, the earlier unconformity is present in the western Clarence-Moreton basin and in basins further to the west, but in the eastern Clarence-Moreton basin, a complete section across the system-period boundary appears to be present (CitationMcKellar in press). The geographic and stratigraphic extent of these hiatuses is presently not well defined. In places (e.g. in the Surat and Eromanga basins), the uppermost Jurassic is also represented by a hiatus. Moreover, in some basinal areas on or near the eastern continental margin (Nambour, northern Clarence-Moreton basins), Upper Jurassic strata are missing entirely. The hiatus extends earlier to the mid- to late Middle Jurassic in the northern Clarence-Moreton and northern Nambour basins and to the late Early Jurassic in the southern Nambour basin (e.g. CitationMcKellar in press).
Forest mires that developed into coal deposits were established in several basins during the Early to Middle Jurassic (Fig. ). The Walloon Coal Measures, which were deposited during the latter epoch in the Clarence-Moreton and Surat basins, are an important source of thermal coal, and also (in the Surat basin) coal seam gas, commercially exploited since January 2006. Contrastingly, in the northern Nambour basin, the Lower-Middle Jurassic Tiaro Coal Measures, which are extensively intruded and vary considerably in rank, have been considered uneconomic, although they have been mined previously for graphite and are being explored presently for economic coal. The uppermost Triassic–Middle Jurassic succession preserved in this northern area of the Nambour Basin was previously included in the Maryborough Basin (now restricted to Lower Cretaceous strata). For summaries of the distribution and properties of Australia's Jurassic coals, see Goscombe & Coxhead (Citation1995) and Sappal & Suwarna (Citation1997).
From 1984 to 1996, the Bureau of Mineral Resources (renamed the Australian Geological Survey Organisation, AGSO, in 1992; now named Geoscience Australia, GA) compiled and produced a series of palaeogeographic atlases of Australia for the Phanerozoic Eon, as a succession of time-slice maps for most periods. The first volumes were published in 1988, with the Jurassic assessment by Bradshaw & Yeung (Citation1990, Citation1992) coming later. More recently, the entire set was published on CD-ROM (Geoscience Australia Citation2002) and, presently, the Palaeo Atlas is freely available online at http://www.ga.gov.au/multimedia/palaeo/html/palaeo.html
The first Australian Jurassic fossils were found in the early nineteenth century by the pioneer explorers and scientists, Thomas Mitchell (in 1838) and J.W. Gregory (in 1849), but the fossils' ages were not clarified until much later (Burger & Shafik Citation1996; Grant-Mackie et al. Citation2000). Fossil plants and fish have been important from the earliest investigations, but attempts to understand the hydrogeology and hydrocarbon potential of Australia's Jurassic basins did not begin in earnest until the 1950s and 1960s, with ongoing exploration for, and development of, petroleum and coal/coal seam gas giving impetus to continuing palynostratigraphic studies (e.g. CitationMcKellar in press) and PhD project work [e.g. by Daniel Mantle at the Department of Earth Sciences, University of Queensland, on the palynology, sequence stratigraphy and palaeoenvironments of Middle to Upper Jurassic strata in the Bayu-Undan (gas condensate) Field, Timor Sea region (see below); and Natalie Sinclair, Department of Earth and Marine Sciences, Australian National University, Canberra, on the sequence stratigraphy and palynostratigraphy of the Late Jurassic in the Jansz Gas Field on the North West Shelf].
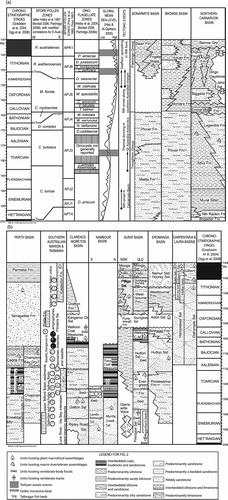
The Jurassic is now taken to cover a time range of some 54 million years, from 199.6–145.5 Ma, based on the international geologic timescale of Gradstein et al. [Citation2004; abbreviated here to GTS (Citation2004), updated by Ogg et al. Citation2008, abbreviated to GTS (Citation2008); Fig. ]. The period incorporated a broad interval of marine transgression in most parts of the world (Haq et al. Citation1987; Hallam Citation1996; Haq & Al-Qahtani Citation2005), but eastern Australia predominantly remained above sea level (Bradshaw & Yeung Citation1990, Citation1992; Burger & Shafik Citation1996). Eastern Australian basins contain many hundreds of metres of continental strata, which thus lack the appropriate marine microplankton and invertebrate indices to enable ready correlation with marine rocks in Western Australia and overseas, and with international chronostratigraphic stages. Hill & Maxwell (Citation1967, p. 55) noted thin ferruginous-oolite and associated beds rich in acritarchs in Lower Jurassic (Toarcian) strata of eastern Australia (Fig. ), but the earlier inferred marine influence for these horizons has been discounted; absence of marine fossils and sedimentologial evidence imply a lacustrine environment (Cranfield et al. Citation1994; Grant-Mackie et al. Citation2000; CitationMcKellar in press).
Grant-Mackie et al. (Citation2000) reviewed the published data on the biota and palaeobiogeography of the Australasian Jurassic and concluded that the climate was warm, deserts were few, and there were no polar icecaps; dinosaurs were the dominant land vertebrates and, in the air, by the Late Jurassic, there were pterosaurs and early birds while conifers and pteridosperms dominated the terrestrial floras. McKellar (Citation1996, Citation2004, Citationin press) has also assessed the changes in Australian palaeogeography, climate and floras from the late Palaeozoic to the early Mesozoic, and provided largely geophysical and tectonic explanations for the sequence of events leading into the beginning of the Jurassic. Based on his interpretations, the Australian region was rotated through the Permo-Triassic to earliest Jurassic into progressively lower and warmer latitudes. At the very beginning of the Jurassic (Hettangian), the palynofloral evidence from the eastern Clarence-Moreton basin suggests that there was a transition from the mesothermal climate of the Late Triassic of southeastern Queensland to a slightly warmer and more monsoonally influenced climate in the early Sinemurian (CitationMcKellar in press). These latitudinal/climatic changes enabled thermophilic and xeromorphic cheirolepidiacean conifers to thrive there during most of the Early Jurassic, under seasonally dry, warm conditions. Before the end of the epoch, the climate had become consistently humid, and remained so for the remainder of the Jurassic, with a mesothermal (warm temperate) climate becoming well established in the Late Jurassic, in accord with relocation into somewhat higher latitudes.
A resurgence of interest in the Australian Mesozoic floras has followed the remarkable discovery of the extant Wollemi Pine in the Blue Mountains Area, west of Sydney, NSW, in 1994. This discovery has particularly prompted intense study of the 200-million-year-plus history of the once cosmopolitan, but now relictual family Araucariaceae. Furthermore, discovery of new fossil assemblages has expanded the potential biogeographic, palaeoenvironmental and phylogenetic application of Australian Jurassic plants (Clifford Citation1998; Pattemore Citation2000; Pattemore & Rigby Citation2005; McLoughlin & Vajda Citation2005; Jansson et al. Citation2008b; McLoughlin & Pott Citation2009).
In the marine Jurassic sequences of Western Australia, belemnites have been recorded from the Middle Jurassic (Bajocian), but are found far more commonly in Upper Jurassic strata; ammonites from the lower Bajocian Newmarracarra Limestone, Champion Bay Group (Perth basin), allow correlation with the European standard biozones (Burger & Shafik Citation1996; Grant-Mackie et al. Citation2000). Marine vertebrates are less well known in the Jurassic of Australia (Long Citation1998; Grant-Mackie et al. Citation2000), and most vertebrate work in recent years has been conducted on non-marine, lower vertebrates (see below).
This paper provides an update on the state of work on the Jurassic in Australia, with particular focus on palaeobotanical–palynological and palaeoichthyological studies. It also highlights the current problems and deficiencies in dating and correlating Australian Jurassic strata that need to be addressed by future research.
Geological–palaeogeographical setting of Australia
The shifting pattern of environments through the Jurassic was primarily influenced by changes in global sea level/base level, changes in palaeogeographic location, and by the new tectonic regime that became established early in the period (Bradshaw & Yeung Citation1990, Citation1992; CitationMcKellar in press). These parameters were controlled not only by the break-up of Pangaea and, within its southern region, Gondwana, but also, importantly, a change in direction of rotation of Pangaea around the beginning of the Jurassic.
Tectonism and continental rotation
Australia was thermally emergent, high-riding and relatively stable in the Early Jurassic (Bradshaw & Yeung Citation1990). The break-up of eastern Gondwana began during the early Late Jurassic with the Argoland rifting event (along northwestern Australia), represented by a widespread unconformity associated with thermal uplift, normal and strike-slip faulting, rift volcanism and erosion (Bradshaw & Yeung Citation1990; Ramsay & Exon Citation1994; Exon & Colwell Citation1994; Exon & von Rad Citation1994). This tectonism, according to the cited authors, resulted in the genesis of the Argo Abyssal Plain by commencement of sea-floor spreading at 155 Ma (latest Oxfordian) when the Argo Landmass began to rift away prior to drifting northwestwards. More recently, Heine & Müller (Citation2005) have indicated that formation of the northern Gascoyne Abyssal Plain, northwest of the Exmouth Plateau and west of the Argo Abyssal Plain, was also associated with separation of this landmass, said probably to be the West Burma Block of Metcalfe (Citation1999), with the “Argo” spreading ridge continuing around the northern margin of Greater India. Their interpretation of the magnetic anomaly record, based on Gradstein et al. (Citation1994), shows that spreading started with M25 (154.1 Ma) as the first correlatable anomaly in both abyssal-plain areas, and with M26 (155.0 Ma) as the oldest identifiable in the southern Argo Abyssal Plain. Largely, however, the Gascoyne Abyssal Plain is related to the slightly younger Greater India–Australia break-up event (Heine & Müller Citation2005). Further, according to Gradstein & von Rad (Citation1991), the Argo break up unconformity preceded the onset of sea-floor spreading by about 5–10 million years. Effects of this fragmentation were widespread. They are not only evident in all basins of the Westralian Superbasin (Fig. ), but are also recorded in the Nepal Himalaya Thakkhola region by a “Callovian-Oxfordian” hiatus and the occurrence of a ferruginous oolite horizon, the distribution of which embraces southern Tibet and parts of the Indian subcontinent (Klootwijk Citation1996). During the Triassic and Jurassic, these areas (including Thakkhola) and, to the east, the Exmouth Plateau, formed part of a broad continental shelf along the south Tethyan continental margin of northeastern Gondwanan (Gradstein & von Rad Citation1991; Gradstein et al. Citation1992; Exon & von Rad Citation1994; Ogg & von Rad Citation1994). Later, further seafloor spreading in the Gascoyne and Cuvier abyssal plains commenced around 136–131.9 Ma (Early Cretaceous; Veevers Citation2000) as Greater India rifted to the west. Rifting continued in an anticlockwise sense around Australia until the Paleogene.
Although there are relatively few Jurassic intrusive and extrusive igneous rocks exposed in eastern Australia, it is clear that there was considerable volcanic activity near the eastern margin, based on the large volume of pyroclastic and epiclastic material accumulated in the region's sedimentary basins at that time. Much of this volcanism might have occurred from extensional/incipient-rifting activity on the Lord Howe Rise–Norfolk Ridge–New Zealand terranes, which in the Jurassic were united to Australia's eastern margin, only detaching to the east during the Cretaceous–Paleogene (Veevers Citation2000). Fielding (Citation1996) and Yago & Fielding (Citation1996), in considering the distribution of volcanic-ash beds in the Middle Jurassic Walloon Coal Measures (Clarence-Moreton basin), suggested that the volcanic sources might have been intra-basinal. The meridional depocentre in the eastern Clarence-Moreton basin coincides approximately with the zone encompassing the greatest concentration of volcanic-ash deposits and possibly coeval volcanic centres, lavas and intrusives of bimodal mafic and felsic composition, leading them to propose that this belt might represent the site of a failed rift. A former commonly held view maintained that the volcanic debris resulted from volcanism associated with a subducting-plate margin and arc to the east during the mid-Late Jurassic (Exon Citation1976; Veevers Citation1984; Hawlader Citation1990; He & Conaghan Citation1994).
Australia/eastern Gondwana was rotated clockwise into higher southern latitudes through most of the Jurassic (Klootwijk Citation1996). This process followed the anticlockwise rotation of the region (and Pangaea in general) during the Permo–Triassic–earliest Jurassic, which brought Australia, more or less progressively, into lower palaeolatitudes. These latest Palaeozoic–early Mesozoic changes in latitude are indicated by palaeomagnetic data and accord not only with floristic changes during the Permian to earliest Jurassic, but also the development of Rhaetian reef structures on the northwestern margin (Klootwijk Citation1996; McKellar Citation2004, Citationin press). The change from clockwise to anticlockwise rotation is also implied by the distinct Triassic–Jurassic loop in the Indian apparent polar wander path (APWP), with an apex dated approximately as latest Triassic–earliest Jurassic (Klootwijk Citation1996, fig. 19). The apices of this pole-path loop and equivalent features in other Pangaean APWPs (Piper Citation1987, p. 304, fig. 11.2; Sichler & Perrin Citation1993, pp. 13, 22–25, figs. 10, 11; Gordon et al. Citation1984; Ekstrand & Butler Citation1989; Bazard & Butler Citation1991; Molina-Garza et al. Citation2003) collectively indicate a global lithospheric event and have been held to reflect a major change in the direction of true polar wander (TPW) and, therefore, in the global pattern and distribution of crustal stresses (McKellar Citation2004, Citationin press). This interpretation, relating the changing direction of rotation of Pangaea to TPW events, is supported by the work of Steinberger & Torsvik (Citation2008) and matches an interval of global plate reorganisation (e.g. Korsch & Totterdell Citation1996). In Greater Eastern Australia, the changes in crustal stress along the ocean-continent interface led to termination of the compressional regime that dominated the Permo-Triassic, with its associated “dextral rotational force couple” then being replaced by a sinistral system of shear (Evans & Roberts Citation1980, p. 325), although the latter authors dated the change as Norian. However, the timing of the change in tectonic style and contemporaneous floristic changes (with attendant climatic signatures) in eastern Australia suggests that the anticlockwise rotation and northward displacement of the region continued into the Hettangian, before being reversed in the Sinemurian (CitationMcKellar in press).
Eastern Australian Late Permian–Triassic tectonism encompassed a series of compressional pulses that emanated from the Panthalassan hemisphere and, for eastern Australia, has been referred to, with the exception of further deformations in the Norian (Late Triassic) and latest Triassic–earliest Jurassic, as the Hunter–Bowen Orogeny (e.g. CitationMcKellar in press). On a broader scale, Du Toit (Citation1937, p. 63, 79–80, fig. 10) coined the contractional term, Samfrau Geosyncline/Orogenic Zone, to represent the “lengthy trough” that existed along the Gondwanan margin of South America–South Africa–Australia, in which strata were subjected to “several pulses” of “intermittent compression and upheaval” at this time. However, the Hunter–Bowen Orogeny, which was previously considered to have terminated about the end of the Middle Triassic (e.g. Fielding Citation1996; Stephens et al. Citation1996), continued into the beginning of the Jurassic, as similarly recognised for the Cape Orogeny (Cape Fold Belt, South Africa), which embraced a comparable series of deformational events that has been identified with the aid of 40Ar/39Ar step-heating analysis (Hälbich et al. Citation1983). Overall, the tectonic system that encompassed these compressional episodes has been viewed as being TPW-induced, consequent to tectonism generated by a changing geoid as rapid rotation-axis adjustments occurred during the Permian–earliest Jurassic (McKellar Citation2004, Citationin press).
As compressional relaxation occurred in eastern Australia with the beginning of the clockwise rotation, the subduction zone to the east apparently retreated. The new stress regime controlled sedimentation and structure during the Early Jurassic (Evans & Roberts Citation1980). Moreover, the changing patterns of global lithospheric stress in the latest Triassic–earliest Jurassic might have assisted rifting of the Pangaean supercontinent. Significantly, onset of this rifting was a Pangaea-wide event, which facilitated considerable heat loss from the Pangaean thermal anomaly. The modern relic of this process is purported to be the long-wavelength African-eastern Atlantic residual geoid high (Anderson Citation1982, Citation1989). Accumulation of heat below, and thermal elevation of, the massive, mantle-insulating supercontinent created a mass anomaly in Earth's rotation, since thermal elevation places mass at a particular latitude at a greater distance from Earth's rotation axis, thus increasing the contribution to the moment of inertia at that latitude. Such a process effects a change in the location of the planet's principal axis of inertia, with which the spin axis needs to be aligned for rotational stability (Gold Citation1955; Anderson Citation1989). These were the primary influences that seemingly controlled the anti-clockwise rotation and northwards displacement of Pangaea during the latest Palaeozoic and early Mesozoic, a long-drawn-out adjustment that was completed in the earliest Jurassic, that point in time being marked by the above-cited Indian pole-path loop apex (McKellar Citation2004, Citationin press). Soon after the beginning of the Jurassic, the Australian region was located in the lowest latitudes that it was to occupy for the entire duration of the latest Palaeozoic and Mesozoic, and more of Pangaea was located in low to middle latitudes than at any other time in the supercontinent's history. There would have been an equal distribution of Pangaea, with its underlying mass anomaly, its peripheral subduction zones (also representing geoid anomalies, but of intermediate scale), and other mass anomalies about the equator at this time – a requirement of the planet's physical system to minimise kinetic energy of rotation. This largely accounts for the traditional view, from a global perspective, that Jurassic climates were warm and highly equable (e.g. Barnard Citation1973; White Citation1986). No landmass occupied the rotation poles at the beginning of the period, as TPW located geoid highs (and mass excesses) about the equator and geoid lows about the geographic poles. That a major period of TPW did occur during the latest Palaeozoic–early Mesozoic has been strongly advocated by Anderson (Citation1982, Citation1989, Citation2007) and subsequently maintained by Marcano, van der Voo, & Mac Niocaill (Citation1999) and Steinberger & Torsvik (Citation2008).
Basin development
Mid-Mesozoic sediment accumulation in the broad epicratonic basins of eastern Australia began, on a limited scale, in the latest Triassic (Rhaetian), but tectonism, involving the final compressional pulse of the above-cited Permo–Triassic tectonic regime, gave rise to a widespread unconformity about the beginning of the Jurassic. Along the northwestern Australian margin, the end-Triassic unconformity locally embraces the Hettangian, although sedimentation may have continued in some areas without interruption across the Triassic–Jurassic boundary (Bradshaw & Yeung Citation1990, Citation1992; Exon & Colwell Citation1994). The depositional break in northwestern Australia has been associated primarily with the commencement of rifting, with rift volcanics of latest Triassic–earliest Jurassic age being erupted in areas of future break-up. In eastern Australia, the Hettangian–earliest Sinemurian unconformity is present in the western Clarence-Moreton Basin and basins further west, but, in the eastern Clarence-Moreton Basin and perhaps also in the coastal Nambour Basin, there was apparently continuous sedimentation across the period boundary (CitationMcKellar in press). Deposition recommenced elsewhere in the early Sinemurian, greatly increasing throughout the Early Jurassic. It expanded in a westerly direction, with sediments ultimately accumulating in the western Clarence-Moreton, Surat, Mulgildie, and Eromanga basins (CitationMcKellar in press; Fig. ).
Where such a hiatus is present in eastern Australia, it delimits the change from the Alisporites/Falcisporites Microflora, representative of the temperate Dicroidium Flora, to palynofloras of Jurassic aspect dominated by cheirolepidiacean pollen (Classopollis), reflecting the assumed shift to somewhat lower palaeolatitudes and warmer climates (e.g. Grant-Mackie et al. Citation2000). Where the hiatus is apparently absent (eastern Clarence-Moreton and probably Nambour basins), the Hettangian and earliest Sinemurian embrace a transitional palynoflora, although it is, by far, more closely allied to the Alisporites Microflora (based on data from N. de Jersey and CitationJ. McKellar).
A significant regional unconformity is also represented at the boundary between the Birkhead Formation and Adori Sandstone in the Eromanga Basin, and most likely at the corresponding Walloon Coal Measures–Springbok Sandstone boundary in the Surat Basin, based on palynostratigraphic assessment and lithostratigraphic extrapolation. This break has been taken to represent the Callovian–Oxfordian Argoland break-up unconformity at 155 Ma (latest Oxfordian) and extensional (incipient-rifting) influences in eastern Australia (discussed above; CitationMcKellar in press).
Six major cycles of sedimentation have been recognised in the Surat Basin Jurassic-Cretaceous succession, which are represented to a greater or lesser degree in adjoining basins, depending on the extent of the succession preserved (Exon Citation1976, Citation1980; Exon & Burger Citation1981). Each cycle commenced with sandy deposits of high-energy braided river systems and terminated with mudrock-dominated deposits of low-energy meandering rivers, lakes, and mires. Ponding in the Clarence-Moreton, Surat, and Mulgildie basins at the end of one such cycle in the Middle Jurassic led to the development of extensive forest-mire systems now represented by the Walloon Coal Measures (Fig. ). These large-scale sedimentary cycles (some spanning >10 Ma) probably developed in response to episodic flexure along the eastern continental margin, because of thermal doming and incipient rifting; and extension/incipient-rifting within the Clarence-Moreton Basin (discussed above). In contrast, the Surat, Clarence-Moreton, and northern Nambour (“Maryborough”) basins were incorrectly interpreted by Jones and Veevers (Citation1983, Citation1984) in terms of a foreland-basin system; they attributed the sedimentary cycles to periodic tectonic uplift in the orogen, and maintained that the cycles correlated with global sea-level movements to an extent that suggests global links between tectonism and eustasy. On the other hand, Exon & Burger (Citation1981) and Burger (Citation1986, Citation1989) related changing patterns of sedimentation directly to sea-level movements, using the most likely ages of associated palynofloras as a guide to correlate with the global sea-level curve of Vail et al. (Citation1977). A slight modification of these relationships, based on other curves (Vail & Todd Citation1981; Haq et al. Citation1987), was subsequently adopted (Burger Citation1994a, Citation1994b; Burger & Shafik Citation1996). However, ages assigned to Jurassic formations in the Surat Basin interpreted from correlations with sea-level curves, differ from ages obtained via correlation of their attendant palynofloras with those in New Zealand that have been accurately dated by associated marine faunas (de Jersey & Raine Citation2002; CitationMcKellar in press). This suggests that the published “global” sea-level curves may not be fully reflected in the pattern of eastern Australian sedimentation. Nonetheless, the geographically extensive deposition of oolitic ironstone in southeastern Queensland in the Surat, Mulgildie, Clarence-Moreton, and Nambour basins (e.g. Grant-Mackie et al. Citation2000) does appear to correlate with the early Toarcian peak in global sea level (CitationMcKellar in press). Such deposits were apparently formed in warm humid climates and generally in association with conditions of rising base level (Hallam Citation1975; Frakes et al. Citation1992, p. 63).
Palaeogeography and palaeoclimate
A wealth of palaeontological evidence has been compiled on the Australasian Jurassic and interpreted palaeogeographically and palaeoclimatically (Bradshaw & Yeung Citation1990, Citation1992; McKellar Citation1996, Citation2004, Citationin press; Grant-Mackie et al. Citation2000). However, Australia's palaeolatitudinal location during the Jurassic is still not well constrained. The Australian Mesozoic APWP has been little studied since publication of the largely conceptual pole path of Embleton (Citation1981, Fig. ), which was based on a few averaged poles (Klootwijk Citation1996). Klootwijk (Citation1996) indicated that the Australian pole path shows the general outline of a (Late Triassic–) Early Jurassic loop, followed by a Late Jurassic–Early Cretaceous loop. The Indian APWP is considerably better established and is characterised by an extensive Triassic–Jurassic loop with an apex around the Triassic–Jurassic boundary, followed by a Jurassic–Early Cretaceous loop with an apex around the Jurassic–Cretaceous boundary. The Indian Triassic–Jurassic loop is far more extensive than the Australian loop, suggesting that the apex of the latter is presently undefined. The Indian Jurassic–Early Cretaceous loop is less extensive than the Australian loop, leading to the conclusion that the former is possibly underdetermined. These palaeomagnetic data indicate that, during the Jurassic, eastern Gondwana rebounded towards higher southern latitudes. However, the Australian and Indian pole paths are no more than broadly determined for the Jurassic and do not provide fine detail of this southward movement (Klootwijk Citation1996). Hence, palaeogeographic reconstructions showing Australia's location during the period can be regarded as providing only a general picture. Nonetheless, Australia's rotation to higher (and cooler) latitudes through the Middle to Late Jurassic is reflected by the palynofloristic change from the Araucariacean Phase to the Podocarpacean Phase and the attendant onset of the Microcachryidites Microflora [of Helby et al. (Citation1987); see below].
Fig. 3 Representative Australian Jurassic plant macrofossils. A. Dictyophyllum sp. (dipteridacean fern), QMF12659. B. Sagenopteris nilssoniana (Brongniart) Ward (caytonialean gymnosperm), QMF12971. C. Pachydermophyllum sp. (“seed-fern”), GSQF13548. D. Masculostrobus sp. (conifer microsporangiate cone), AMF78330. E. Palissya ovalis Parris, Drinnan & Cantrill (palissyacean gymnosperm), UQF79695. F. Taeniopteris spatulata McClelland (pentoxylalean gymnosperm), UQF79721. G. Hausmannia sp. cf. H. deferrariisii Feruglio (dipteridacean fern), UQF79687. H. Podozamites jurassica White (araucariacean conifer), AMF59990. I. ?Rissikia talbragarensis White (?podocarpacean conifer), AMF72408. J. Rintoulia sp. (“seed-fern”), AMF127292. Specimens from Gatton Sandstone, Pliensbachian, Clarence-Moreton basin (A, B); Injune Creek Group, Bathonian–Callovian, northern Surat basin (C); Walloon Coal Measures, Bathonian–Callovian, Clarence-Moreton basin (D–G); and Talbragar fossil beds, latest Oxfordian–Tithonian, southern Surat basin (H–J). Scale bars = 5 mm.

This transition from the Araucariacean to the Podocarpacean Phase was viewed by Grant-Mackie et al. (Citation2000), who supported the general (global) palaeoclimatic model proposed by Frakes et al. (Citation1992), as a time of climate cooling. Based on Australian palynofloral criteria similar to those considered by Grant-Mackie et al. (Citation2000), McKellar (Citation1996, Citationin press) had likewise recognised this time as a period of cooling, but only in a regional context. Grant-Mackie et al. (Citation2000) proposed that the development of a proto-Indian Ocean, in the form of the Argo Abyssal Plain, contributed to the progressive rise in mean sea level that continued into the Early Cretaceous. This sea-level rise was said to have expanded the area of shallow epicontinental seas and hence the available extent of the maritime heat sink, transferring heat to the oceans and giving rise to generally cooler and more equable global temperatures (although the cooling was possibly ameliorated by an increase in atmospheric CO2) during the Middle and Late Jurassic.
The Araucariacean–Podocarpacean Phase transition, reflecting the purported climatic cooling in eastern Gondwana, suggests that much of the palaeolatitudinal motion revealed by Klootwijk (Citation1996) must have occurred quickly during the Late Jurassic, probably following commencement in the latest Oxfordian of sea-floor spreading associated with separation of the Argo Landmass on the northwestern margin (CitationMcKellar in press). In the Northern Hemisphere, there was a northwards shift of floral zones across Eurasia during the Late Jurassic, which was contrastingly interpreted by Vakhrameev (Citation1964), Krassilov (Citation1981), Hallam (Citation1985) and Vajda (Citation2001) as pointing to a general warming trend. Ziegler et al. (Citation1993), however, considered that these motions have no direct implications for global change as they are relative only to the land area and not to palaeolatitude. They indicated that by Middle to Late Jurassic times, a clockwise southern motion of the continents had replaced a previous general northward movement, thus supporting Klootwijk's (Citation1996) interpretation for the eastern Gondwanan sector and accounting for the differing climatic trends in opposing hemispheres during the Late Jurassic (CitationMcKellar in press).
Timescales and boundaries
AGSO published An Australian Phanerozoic Timescale (Young & Laurie Citation1996), in which Burger & Shafik (Citation1996) detailed the Australian Jurassic, including biostratigraphic zonation and international correlation. However, since publication of GTS (Citation2004), the emphasis in GA, with its Timescales and Virtual Centre of Economic Micropalaeontology and Palynology (VCEMP) Project, has been not only to maintain microfossil biozonations and develop them further, but also to define the relationship of Australian sequences with that timescale, rather than to update and maintain independent charts, nonetheless continuing to place particular emphasis on those parts of the stratigraphic column, where economic resources are important.
GTS (Citation2004) (see also Ogg et al. Citation2008) defines the Middle Jurassic as spanning a considerably shorter time interval compared to previous timescales. Although GTS (Citation2004) (see also GTS Citation2008) is the most researched and accurate chronostratigraphic scheme to date, the definitions of many stage boundaries remain problematical, and most remain informally defined, largely based on European usage. Hence, no stage boundary in Australia is well constrained, although current work is improving the biostratigraphic resolution of the Rhaetian–Hettangian (Triassic–Jurassic) and the Hettangian–Sinemurian boundaries in eastern Australia (work of N. de Jersey and JMcK in progress in eastern Clarence-Moreton Basin, see below). It is difficult to determine the age of biostratigraphic zones and hence lithostratigraphic units representing them in the Jurassic of Australia, at the level of stages, because the system is dominated by continental sequences. Marine invertebrate faunas and dinoflagellate assemblages that provide good biostratigraphic control via international correlation are recorded only in Western Australia. It has been largely by establishing relationships between the marine and continental biostratigraphies via analysis of spore–pollen assemblages that correlations and age determinations have been improved across the country. Dating of lithostratigraphic units in eastern Australia has also been facilitated by spore–pollen correlation with New Zealand, where palynological zones have been well dated using marine invertebrates (de Jersey & Raine Citation2002; CitationMcKellar in press). Generally, however, stage boundaries appear to fall within biostratigraphic zones, hence the indication above that, by and large, these chronostratigraphic units can not be well delimited.
Palaeontology
Plant macrofossils
Jurassic macrofloras (Fig. –J) are widely distributed, but poorly studied in Australia. Based on the more extensive palynological record, three major modifications of Australasian floras are recognisable during the Jurassic: the first somewhat after the beginning of the period; the second in the Toarcian; and the third in the early Late Jurassic (McKellar Citation1996, Citationin press, Grant-Mackie et al. Citation2000).
The Triassic–Jurassic transition in Australia is represented by a relatively abrupt decline of the corystosperm-dominated Dicroidium Flora, and the Jurassic is characterised by the rise of conifer and bennettite-dominated floras. However, as noted above, Hettangian strata, present in the eastern Clarence-Moreton Basin (and perhaps also in the Nambour Basin) embrace a transitional palynoflora, which displays significantly closer relationships to the Alisporites/Falcisporites Microflora (the palynofloral representation of the Dicroidium Flora) than to subsequent (Sinemurian–Pliensbachian–earliest Toarcian) palynofloras that are rich in Classopollis. The dominance of Classopollis in mid–Early Jurassic palynofloras points to significant representation of cheirolepidiacean conifers in the vegetation, but macrofossils of this group have not yet been identified with confidence from Australian deposits of this age.
The best-described Early Jurassic floras, generally associated with the later part of the epoch, derive from the Clarence-Moreton and Nambour basins (e.g. Gould Citation1968, Citation1971; Rozefelds & Sobbe Citation1983; Grant-Mackie et al. Citation2000; Pattemore & Rigby Citation2005; Jansson et al. Citation2008b). These Pliensbachian–Toarcian assemblages contain a range of lycophytes, equisetaleans, osmundaceous, dipteridaceous (Fig. ), dicksoniaceous and matoniaceous ferns, the pteridosperms Komlopteris and Rintoulia, caytonialeans [Sagenopteris (Fig. ) and Caytonia], bennettitaleans (Otozamites spp.), putative ginkgoaleans, small-leafed conifers (Plagiophyllum and Allocladus), and Palissya fructifications.
The fossil floras from Ida Bay and Lune River, in Tasmania, include a broad array of exquisitely permineralised conifers, bennettites, and especially osmundaceous and cyathealean ferns (Gould Citation1972; Tidwell Citation1987, Citation1992; Tidwell & Jones Citation1987; Tidwell et al. Citation1987; Tidwell et al. Citation1989; Tidwell et al. Citation1991; White Citation1990, Citation1991; Tidwell & Pigg Citation1993). These assemblages have traditionally been ascribed a Late Jurassic age, but radiometric dating has revealed an age of 175 ± 6.4 Ma (Toarcian–Bajocian) for basalts associated with the sediments (Hergt et al. Citation1989; Bromfield et al. Citation2007).
Australasian and Gondwanan floras generally were strongly modified over a short transitional phase through the late Early Jurassic; plant assemblage changes of this time have been used as palaeoclimatic indicators (McKellar Citation1996; Hallam Citation1996). Most notable was the decline in cheirolepidiacean conifer pollen and the rise in relative abundance of araucarian and podocarp conifers. This transition is matched in the sedimentary record of several eastern Australian basins by the occurrence of one or more beds of oolitic ironstone (e.g. Grant-Mackie et al. Citation2000) and suggests a significant perturbation of the terrestrial environment (Cranfield et al. Citation1994). Significantly, these terrestrial changes were roughly coeval with the early Toarcian peak in global sea level (see above, CitationMcKellar in press) and the early Toarcian anoxic event in the Tethys Ocean (Fig. ).
The Walloon Coal Measures and their equivalents in the Clarence-Moreton, Surat, and Mulgildie basins have been considered to be Bajocian–Bathonian in age, based on macrofloral affinity with the Clent Hills Group flora of New Zealand (Grant-Mackie et al. Citation2000), or Bathonian–Callovian in age, based on palynological evidence. The latter encompasses the oldest records of Contignisporites glebulentus (Fig. ) in the latest Temaikan marine succession at Awakino (North Island, New Zealand; de Jersey & Raine Citation2002) and of Murospora florida, a species not reported from New Zealand but occurring in the late Temaikan of New Caledonia (Grant-Mackie et al. Citation2000). In terms of international stage divisions, these occurrences (C. glebulentus, M. florida) collectively indicate a Callovian age. However, in the Walloon Coal Measures of the Surat Basin, the two species first appear, in the order cited, part of the way through the succession, although the range of C. glebulentus might extend down to just above the base of the formation (CitationMcKellar in press). In comparison, the formation in the southeastern Queensland portion of the adjacent Clarence-Moreton Basin lacks these species and may be somewhat older (Bathonian). Further, the top of the coal measures in eastern Australia is marked by the unconformity between the Middle and Late Jurassic (discussed above).
Fig. 4 Representative Australian Jurassic palynomorphs from the Surat basin, Queensland. A. Dictyotosporites complex Cookson & Dettmann, Walloon Coal Measures, Q354. B. Retitriletes circolumenus (Cookson & Dettmann) Backhouse, Westbourne Formation, Q379. C. Staplinisporites manifestus McKellar, Hutton Sandstone, Q432. D. Contignisporites glebulentus Dettmann emend. Filatoff & Price, Westbourne Formation, Q612. E. Nevesisporites vallatus de Jersey & Paten, Walloon Coal Measures, Q572. F. Foraminisporis dailyi (Cookson & Dettmann) Dettmann, Westbourne Formation, Q583. G. R. watherooensis Backhouse, Westbourne Formation, Q381. H. Leptolepidites verrucatus Couper, Westbourne Formation, Q487. I. Classopollis sp. cf. C. chateaunovi Reyre, Walloon Coal Measures, Q763. J. Ischyosporites punctatus Cookson & Dettmann, Westbourne Formation, Q446. K. Podosporites variabilis Sukh Dev, Walloon Coal Measures, Q735. L. Murospora florida (Balme) Pocock, Westbourne Formation, Q627. M. Araucariacites fissus Reiser & Williams, Walloon Coal Measures, Q759. N. Callialasporites dampieri (Balme) Sukh Dev, Westbourne Formation, Q691. Fig. 4M × 600, all other images approximately × 500. Ages of host units indicated in Fig. . Numbers prefixed by “Q” refer to the palynological specimen-locality catalogue of the Geological Survey of Queensland, Department of Employment, Economic Development and Innovation.
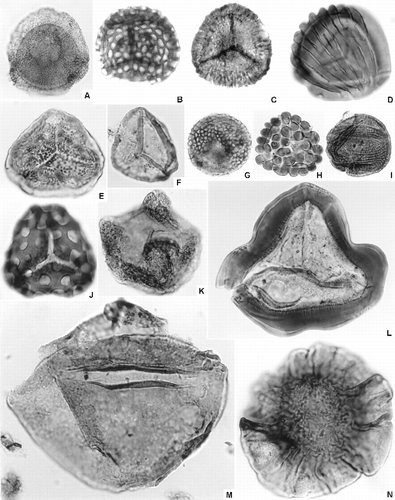
The coal measures host a typical Middle Jurassic flora incorporating liverworts, selaginellalean lycophytes, equisetaleans, osmundaceous, marattiaceous, matoniaceous, dicksoniaceous and dipteridaceous ferns (Fig. ) and araucarian, podocarpacean, and palissyacean conifers [Araucarites sp., Mataia sp., Allocladus sp., Elatocladus spp., Pagiophyllum sp., Brachyphyllum crassum, Masculostrobus sp. (Fig. ), Palissya ovalis (Fig. )], bennettitaleans (Ptilophyllum pecten, Otozamites feistmanteli, Williamsonia sp.), pentoxylaleans (Taenopteris spatulata: Fig. ), and some gymnosperms of unknown affinity (Pachypteris and Pachydermophyllum spp.: Fig. ) (Tenison-Woods Citation1882, Citation1883; Walkom Citation1917a, Citation1917b, Citation1919; Day Citation1964; Hill et al. Citation1966; Townrow Citation1967a, Citation1967b; Gould Citation1968, Citation1975, Citation1980; Grant-Mackie et al. Citation2000). Surprisingly, given their abundance in most parts of the middle to high latitudes in both hemispheres at this time, ginkgoaleans have not been recorded in the Middle Jurassic of Australia, the regional climate then being considered too warm for this group to proliferate in lowland settings, as they did in the Late Triassic and Early Cretaceous of the region (McKellar Citation1996).
The rich macroflora of the Walloon Coal Measures is encompassed by the concept of the Otozamites-Ptilophyllum Flora of Gould & Shibaoka (Citation1980), although bennettitaleans are not abundant in all assemblages (McLoughlin & Drinnan Citation1995). The presence of large dipteridaceous ferns and large-leafed araucaraiacean conifers generally favours a mesothermal climate.
Permineralised gymnosperm wood is common in Middle Jurassic volcanigenic sediments of eastern Australia, but has not been extensively studied. Following recent taxonomic revision of global Jurassic–Cretaceous wood records (Bamford & Philippe Citation2001), such fossils may find renewed application for terrestrial biostratigraphy and palaeobiogeography (Philippe et al. Citation2004). Unstudied in situ stumps occur at Miles in the Surat basin and a range of permineralised osmundaceous and dicksoniaceous fern axes have been described from nearby deposits (Tidwell & Rozefelds Citation1990, Citation1991; Tidwell & Clifford Citation1995). Pronounced growth rings in conifer woods and the abundance of matted bennettitalean-pentoxylalean leaf horizons suggest that the climate was seasonal with some plants deciduous. However, severe winters in the late Early to Late Jurassic in the Surat and adjacent basins might have been moderated by the extensive system of swamps, lakes, and rivers, a scenario mooted by Yemane (Citation1993), Ziegler et al. (Citation1993) and Kutzbach & Ziegler (Citation1993). Also, without details of the growth-ring patterns, determining likely causes of their formation is difficult, as these features can result from different environmental parameters and can be found even at low elevations at low latitudes (Fritts Citation1976).
Floras of the mid-Jurassic Araucariacean Phase are similar from all Australasian localities and correlate with assemblages in Argentina; comparable assemblages are known from peninsula India, Madagascar, and Antarctica (Grant-Mackie et al. Citation2000). Recent use of U-Pb zircon ages for calibration of Southern Hemisphere biostratigraphy has provided fresh insights into the age of plants from the non-marine Botany Bay Group, Antarctica. Hunter et al. (Citation2005) gave an absolute age of 168.9 ± 1.3–167.1 ± 1.1 Ma (Bajocian to early Bathonian, GTS Citation2004) for the reference flora, which had previously been assigned to the Early Jurassic (Gee Citation1989; Rees Citation1993; Rees & Cleal Citation2004).
Australian Late Jurassic plant macrofossils are poorly known (Young & Laurie Citation1996; Hill et al. Citation1999). They are recorded from disparate parts of the continent, but are broadly similar to those of the Early and Middle Jurassic in their representation of conifers, bennettitaleans, pentoxylaleans, pteridosperms, ferns, and equisetaleans. The Talbragar fish beds in the southern Surat basin host a rich impression flora that has traditionally been assigned a late Early Jurassic or early Middle Jurassic age, based on palynological data (Morgan Citation1984; see below). Recent radiometric dating (see below) suggests a Late Jurassic (latest Oxfordian–Kimmeridgian–Tithonian) age for these beds (Bean Citation2006b). The flora, studied most thoroughly by Walkom (Citation1921b) and White (Citation1981a), includes conifers [Podozamites jurassica (Fig. ), ?Rissikia talbragarensis (Fig. ), “Brachyphylum sp.”, “?Pagiophyllum peregrinum”, Elatocladus australis, Allocladus cribbii, Allocladus milneanus, Elatocladus australis], pentoxylaleans (Taeniopteris, Carnoconites and Sahnia spp.), ?cycadophytes (Nilssonia compta), some enigmatic seed-ferns (Rintoulia sp., Fig. ), and fern fragments. In recent years, several media and technical reports have compared Podozamites jurassica fossils, from Talbragar, to the foliage of the extant wollemi pine (Wollemia nobilis), suggesting great antiquity for the Wollemia lineage. However, P. jurassica does not provide a close match in venation pattern, leaf shape or phyllotaxy to either Wollemia or other extant araucariacean genera (McLoughlin & Vajda Citation2005). Convincing evidence for Wollemia extends back only to the Late Cretaceous.
Small fossil-plant assemblages, from the Carpentaria Basin in far northern Queensland (e.g. Gould Citation1975), the Tarlton Range, Eromanga Basin, Northern Territory (Gould Citation1978), and the northern Perth Basin (e.g. Walkom Citation1921a; Glauert Citation1923; McLoughlin & Hill Citation1996; McLoughlin & Pott Citation2009 ) have also been considered to be of Late Jurassic age. These floras contain putative lycophytes, dipteridacean, ?gleicheniacean, osmundacean and ?matoniacean ferns, bennettitaleans, pentoxylaleans, and small-leafed conifers (Pagiophyllum and Elatocladus spp.). In terms of their generic composition and the typically small-leafed conifers, they show some similarities to southeastern Australian Neocomian floras (McLoughlin et al. Citation2002), but other features (medium-leafed bennettitales) show similarities to Middle Jurassic floras of India, Patagonia, and the Antarctic Peninsula (McLoughlin & Pott Citation2009). Late Jurassic to Early Cretaceous Pachypteris leaves from the Battle Camp Formation of the Laura basin, north Queensland, are notable for the presence of lepidopteran leaf mines, one of the oldest records of phytophagy in the Australian fossil record (Rozefelds Citation1988). As several eastern Australian basins contain a near-complete record of Jurassic continental sedimentation, great scope exists for further systematic studies of the fossil floras and their application to non-marine biostratigraphy.
Palynology
A substantial database now exists on the palynofloral succession represented in Australia's Jurassic sedimentary basins. Following early systematic studies of microplankton (I.C. Cookson and co-workers) and of spores and pollen from Western Australian marginal basins (B.E. Balme), and eastern basins (N.J. de Jersey and co-workers), work was focused on developing intra- and inter-basinal palynostratigraphic schemes, primarily in response to the needs for stratigraphic correlation in hydrocarbon exploration. In the intracratonic terrestrial sequences of eastern Australia, Evans (Citation1966) largely pioneered Mesozoic miospore biostratigraphy when he defined a series of “palynological units” in the Triassic, Jurassic and Early Cretaceous successions of the Bowen-Surat and Eromanga basins in Queensland (also see Evans Citation1965). Subsequently, Burger (Citation1968, Citation1976, Citation1984, Citation1989, Citation1994a, Citation1994b) and Burger & Senior (Citation1979) modified and further qualified Evans' (Citation1965, Citation1966) scheme in order to make it more amenable to practical usage, but deficiencies remained, primarily because published details of miospore-assemblage composition and species distribution in the Middle–Late Jurassic succession were wanting. Various “in-house” and unpublished versions of Evans' scheme were also developed by palynologists working for petroleum exploration companies. However, continual modification of the zonal units and varying approaches to their application in and outside of the exploration industry generated a degree of confusion; to some extent, this was unavoidable as acquisition of palynological data and evolution of the palynostratigraphy went hand-in-hand, being not only constrained by resources available, but also driven generally by the need for immediate results. The most widely used of the private-industry zonations modified from Evans' original (Citation1966) biostratigraphy is that of Price et al. (Citation1985). Their informal zonal nomenclature, which was devised to overcome the disorder generated by the sundry versions of Evans' palynostratigraphic units, was published in brief by Filatoff & Price (Citation1988). The scheme was reviewed by Burger et al. (Citation1992) and subsequently modified and documented more fully by Price (Citation1997). That zonation bears little resemblance to Evans' (Citation1965) original scheme, and comprises range/partial range/interval zones, unlike that of Helby et al. (Citation1987; discussed below) and its subsequent modifications, in which most of the zones are Oppel zones.
Reiser & Williams (Citation1969), studying the Precipice Sandstone and Evergreen Formation of the northern Surat Basin, partly formalised Evans' (Citation1966) units by erecting their Classopollis classoides Zone (with two constituent subzones) and succeeding Tsugaepollenites segmentatus-T. dampieri “zone”; the latter was not formally instituted, as Reiser and Williams were unable to define an upper limit.
Zonation of the latest Triassic–Early Jurassic interval in the Clarence-Moreton Basin was subsequently formulated by de Jersey (Citation1975), primarily for broad comparison with other regions of Gondwana. An ensuing, but informal scheme (de Jersey Citation1976) established a somewhat finer subdivision of the latest Triassic–Early Jurassic strata in the Clarence-Moreton and Nambour basins; a modification was proposed by McKellar (Citation1981) on the basis of additional species-distribution data obtained from the latter basin.
For the latest Jurassic–Early Cretaceous succession in the Surat and Eromanga basins in Queensland, formal biostratigraphic subdivision has been undertaken by Burger (Citation1973, Citation1989). However, incomplete knowledge of latest Jurassic–earliest Cretaceous palynofloras in strata succeeding the Injune Creek Group and equivalents remains a concern and considerably impedes biostratigraphic appraisal of, and correlation in, this interval. Additionally, on a global scale, the marine faunas of the latest Jurassic–earliest Cretaceous show a high degree of provinciality inhibiting a precise correlation between marine and terrestrial ecosystems (Vajda & Wigforss-Lange Citation2006). In the southern Eromanga Basin of South Australia, Gallagher et al. (Citation2008), following a regional palynological review, recognised six subzones of the late Middle–Late Jurassic palynological unit APJ5 (of Price Citation1997). In Western Australia, Jurassic zonal schemes were established by Balme (Citation1957, Citation1964), Filatoff (Citation1975), and Backhouse (Citation1978, and see Grant-Mackie et al. Citation2000); the work of Filatoff, in particular, has facilitated comparison between the Jurassic palynofloral successions in eastern and Western Australia. However, confident correlation between east and west is complicated not only by quantitative differences in the palynofloras, but also by differences in the relative ranges of certain species and the development of slight provincialism in the Early Jurassic (Truswell et al. Citation1999; Grant-Mackie et al. Citation2000).
Much of this work provided the basis for the Mesozoic, pan-Australian, palynostratigraphic zonation proposed by Helby et al. (Citation1987). Their scheme integrated spore–pollen and microplankton biostratigraphies, and, for some intervals, is linked to the marine invertebrate record and extra-Australian dinocyst records. For the Jurassic, the Helby et al. spore–pollen zonation comprised one new zone (Callialasporites turbatus Oppel Zone) and several other zones, which represented modifications of previously established biostratigraphic units (Reiser & Williams Citation1969; Burger Citation1973; de Jersey Citation1975; Filatoff Citation1975; Backhouse Citation1978). However, the concept of the Dictyotosporites complex Oppel Zone, which was based by Helby et al. (Citation1987) on the identically named biostratigraphic unit established in the Perth Basin by Filatoff (Citation1975), is difficult to apply in eastern Australia, primarily because the nominate species (Fig. ) is rare and sporadically distributed in Middle–Late Jurassic strata. This also has impacted upon recognition (in eastern Australia) of the upper limit of their preceding C. turbatus Oppel Zone. The Contignisporites cooksoniae Oppel Zone (Helby et al. Citation1987), which succeeds the D. complex Oppel Zone, is similarly difficult to delineate in the east, because of the rare and sporadic distribution of C. cooksoniae (Balme) Dettmann (sensu Filatoff Citation1975; recorded under C. burgeri Filatoff, McKellar and Price by CitationMcKellar in press). Nonetheless, the M. florida and Retitriletes watherooensis Oppel zones (Helby et al. Citation1987) have been employed by McKellar (in press), with nomenclatural and other modifications, for subdivision of the late Middle–Late Jurassic interval in the Surat Basin. However, because of apparent differences in the relative appearances of certain miospore taxa (e.g. Fig. , L), the mutual boundary between these zones in the Surat Basin was somewhat differently interpreted from the definition given by Helby et al. (Citation1987), which was based principally on the western and northwestern Australian succession.
The zonation of Helby et al. (Citation1987) was incorporated in the AGSO Citation1996 Australian Timescale (Young & Laurie Citation1996; discussed above) and associated Wallchart (AGSO Citation1996), although alternative ages were suggested for some of their Jurassic–Cretaceous dinocyst zones by Burger & Shafik (Citation1996). Subsequently, in 2004, Helby et al., via GA's Timescales-VCEMP Project, published an updated dinocyst zonation for the Jurassic–Early Cretaceous of the North West Shelf, plotted against the AGSO Citation1996 timescale, also incorporating a comparison between the spore–pollen and dinocyst zones in the Early and early Middle Jurassic. More recently, the Timescales-VCEMP Project has facilitated a further update of this North-West-Shelf-dinocyst zonation, together with updates for the Jurassic–Early Cretaceous and Late Cretaceous–Cenozoic spore–pollen, dinocyst and palynological zonations for Australia and Late Cretaceous–Cenozoic palynological zonations for the Gippsland Basin (Monteil Citation2006, Partridge Citation2006a, Citation2006b, Citation2006c, Citation2006d), all now charted against GTS (Citation2004).
The Jurassic spore–pollen zones are grouped into two superzones, an older Callialasporites dampieri Superzone, dated by Helby et al. (Citation1987) as Hettangian–Kimmeridgian, and the younger Microcachryidites Superzone, dated as Tithonian–Albian, and characterised by distinctive ranges and abundances of key spore–pollen taxa (e.g. Fig. –N). The base of the C. dampieri Superzone records the replacement of the corystosperm-dominated Late Triassic vegetation by a flora dominated by cheirolepidiacean conifers. The latter is embodied by the Cheirolepidiacean Phase of Grant-Mackie et al. (Citation2000). The onset of this phase was dated by Helby et al. (Citation1987) as early Hettangian, whereas McKellar (Citation1996) initially favoured a late Hettangian date (in terms of significantly increased frequencies of Classopollis) and, recently, an early Sinemurian age (CitationMcKellar in press; and based on data, for eastern Australia and New Zealand, from N de Jersey and JMcK). The next major change occurred during the Toarcian with the C. dampieri Superzone being characterised by an increased representation of araucarian conifers (McKellar Citation1996, Citationin press; Grant-Mackie et al. Citation2000), representing the Araucariacean Phase of the latter authors. The third modification was dated as late Oxfordian by McKellar (Citation1996), (?)mid-Oxfordian by Grant-Mackie et al. (Citation2000), and as middle Kimmeridgian by McKellar (in press). Its onset corresponds to the C. dampieri/Microcachyridites Superzone boundary, originally dated by Helby et al. (Citation1987) as Tithonian (as inferred above). Floristic provincialism between eastern and Western Australia and a diachronous depositional hiatus across the continent may account for such a great divergence in the interpreted age of this major zonal boundary (CitationMcKellar in press). Increased frequencies of bi- and trisaccate pollen signify expansion and possibly diversification of the podocarp conifers within the vegetation at this time (Podocarpacean Phase: Grant-Mackie et al. Citation2000).
Major modifications to Australia's Jurassic floras are reflected by the quantitative and qualitative changes in spore–pollen assemblages across the three major events outlined above. Within each biozone, more subtle quantitative/qualitative variations in spore–pollen suites may reflect local facies influences or gradual palaeoecological (latitudinal, topographical, or climate-related) gradients across Australia.
In Queensland, Sajjadi and Playford (Citation2002a, Citation2002b) have published a two-part compilation of the systematic and stratigraphic palynology of the Late Jurassic–earliest Cretaceous interval in the Eromanga Basin. Furthermore, McKellar (in press) has studied the late Early to Late Jurassic palynology, biostratigraphy, and palaeogeography of the Roma Shelf area in the northwestern Surat Basin, assessing the Australia-wide phytogeographic-palaeoclimatic implications of the Callialasporites dampieri and Microcachryidites superzone palynofloras (Jurassic–Early Cretaceous) on the basis of a revised tectonic model for the continent. A new cross-Tasman palynostratigraphic compilation has been developed (by N de Jersey and JMcK) embracing work on accurate location of the Triassic–Jurassic (Rhaetian–Hettangian) and Hettangian–Sinemurian boundaries in continental eastern Australia. The results are based on correlation with the marine, ammonite-dated succession in New Zealand, then part of Greater Eastern Australia. Another major paper, on Early and Middle Jurassic spore–pollen assemblages of New Zealand (again accurately dated by associated marine invertebrate fossils) and their biostratigraphic relationships with palynofloras from continental eastern Australia, has been largely completed (see de Jersey & Raine Citation2002 for preliminary biostratigraphic results). Raine et al. (Citation2005) also provided a valuable atlas of palynomorphs for the New Zealand region (encompassing Jurassic forms), many of which also characterise Australian palynofloras.
The Jurassic palynozonation schemes developed for Australia also enable correlation to adjoining terranes of southern Gondwana. For instance, palynofloral assemblages from the MacRobertson Shelf, East Antarctica, although reworked, have demonstrated the presence of upper Lower Jurassic–Lower Cretaceous strata in that region. These palynofloras are compositionally similar to those of the Perth Basin and formerly contiguous basins in eastern India (Truswell et al. Citation1999). Other Indian subcontinent palynofloras have similarities to Australian suites (Upadhay et al. Citation2005) and Truswell et al. (Citation1999) provided an overview of palynofloral relationships between the separate southern Gondwanan regions.
Ongoing research on Australian Jurassic palynofloras is being undertaken at several institutions/consultancies and is principally aimed at providing high-resolution age and facies controls on strata intersected by hydrocarbon exploration wells. Integration of this data with seismic and downhole petrophysical data is providing improved sequence and systems-tract definition for modelling sea-level changes and basin evolution (e.g. see Longley et al. Citation2002). In particular, PhD thesis work on the Upper Jurassic in the Jansz Gas Field, on the Exmouth Plateau of the offshore Northern Carnarvon basin (North West Shelf), has been concentrating on sequence stratigraphy and palynostratigraphy (Natalie Sinclair, pers. comm.).
Other doctoral research (Mantle Citation2006), including illustration of palynomorph diversity and abundance in assemblages that include both marine (dinocysts and acritarchs) and terrestrial (spores and pollen) components, has interpreted the palynology, sequence stratigraphy and palaeoenvironments of Middle to Late Jurassic strata of the Bayu-Undan Field, Timor Sea. The Bayu-Undan Field, located in the northern Bonaparte Basin, contains thick Mesozoic–Cenozoic sedimentary successions, with the majority of target economic petroleum reservoirs confined to the Middle–Upper Jurassic. This PhD project was developed as a response to the need for further characterisation and higher resolution biostratigraphy of the Bathonian–Oxfordian reservoir intervals within the field. The comprehensive taxonomic study was based on 230 sidewall and conventional core samples from the uppermost Plover, Elang, and lower Frigate formations. The marine palynofloras display a high degree of endemism and 14 new species of dinocyst have been described (Mantle Citation2006); more cosmopolitan dinocyst species allow comparisons with ammonite-dated European and Russian assemblages, and a latest Bathonian to early Oxfordian age is ascribed to the reservoir sections. However, Mantle (Citation2006) noted that there are appreciable inter-provincial disparities between the first and last appearance datums of several dinocyst species.
Ten subzones have been recognised informally, collectively encompassing the Ternia balmei and Voodooia tabulata Interval zones (Helby et al. Citation2004) within the Bayu-Undan Field. The methods used to delimit some of these subzones (e.g. first and last common occurrences and acme events) are probably related to environmental parameters and are thus unlikely to prove reliable across a broader geographic region. However, the significant acme events (e.g. the widely recognised Durotrigia magna acme) correlate with regionally recognised flooding episodes (Mantle Citation2006).
The Elang Formation, the main lithological focus of the research, is typically divided into three third-order sequences. Detailed counts of the palynomorph assemblages allowed the recognition of various palynological markers or bioevents that correlated with most of the chronostratigraphic horizons (e.g. transgressive surfaces and maximum flooding surfaces). This enabled the lowstand, transgressive, and highstand system tracts of each sequence to be delimited. The marine-flooding episodes, in particular, are marked by distinctive compositional changes in the palynofloras (particularly acmes), an increase in stenohaline species, or an overall increase in dinoflagellate species diversity. These palynological-assemblage variations correlated extremely closely with gamma-ray log profiles traceable across the Bayu-Undan Field. Palynofacies analyses have also been used to identify depositional trends within the systems tracts and to aid in assessing the palaeoenvironments (Mantle Citation2006). However, the placement of conformable sequence boundaries, marking the gradational change from highstand to lowstand systems tracts, remains imprecise.
Various dinoflagellate species or groups were also considered useful palaeoenvironmental indicators: viz., Ternia balmei (shallow water, nearshore, euryhaline–stenohaline conditions); the Meiourogonyaulax group (nearshore, stenohaline); the Ctenidodinium group (the change from common occurrence of C. fuscibasilarum to abundant C. ancorum mirrors the progressive transgression through the Elang Formation); and the Rigaudella group (offshore, stenohaline).
Palaeoenvironmental inferences derived from analysis of the palynomorph assemblages concur with previous sedimentological interpretations of the Elang Formation (Mantle Citation2006). Thus, it is hoped that this will enable similar interpretations to be made from non-cored intervals (e.g. cuttings samples) that are based exclusively on the palynological content (Mantle Citation2006).
Vertebrate fauna
In terms of clarifying stage boundaries and events, the Jurassic vertebrate record is relatively uninformative, but it may have greater value for the definition of terrestrial biogeographic provinces and climates (Walliser Citation1996). The Australian Jurassic, with its long sequence of non-marine beds, has yielded a series of small but significant faunas (Long Citation1995; Vickers-Rich & Rich Citation1993). Molnar and Thulborn (in Grant-Mackie et al. Citation2000) reviewed knowledge of fossil vertebrates up to 1997. Since, then, new material from several vertebrates has become available from terrestrial and presumed non-marine sites. These discoveries, for instance, have led to work in progress on the classic Talbragar Fossil Fish bed in the southern Surat Basin (e.g. Pogson & Cameron Citation1999; Grant-Mackie et al. Citation2000; Bean Citation2006a, Citation2006b; and see below).
Early and Middle Jurassic fish faunas
There are sparse actinopterygian records in the Clarence-Moreton Basin. Of these, seven fragmentary semionotiform specimens (e.g. Fig. ) have been reported from the Middle Jurassic Walloon Coal Measures (Grant-Mackie et al. Citation2000). Further, the partial body outline of a semionotid fish, probably Lepidotes sp., has been reported from near Monto in the Mulgildie Basin (Thies & Turner Citation2001). It is preserved in dark, iron-rich sandstone, presumably representing the Hutton Sandstone (the unit underlying the Walloon Coal Measures), and was associated with specimens of ?Osmundacaulis, the occurrence of which supports a mid-Jurassic age.
Fig. 5 Jurassic fish: A. semionotiform fish from the Walloon Coal Measures (mid-Jurassic), Queensland [from Turner and Rozefelds (Citation1987) reproduced with permission of MQM]. B. Cavenderichthys talbragarensis, Australian Museum specimen AMF4133. C. Cavenderichthys talbragarensis, Geological Survey of New South Wales specimen MMF13561a. D. Coccolepis sp. AMF117880. E. Aphnelepis sp., unregistered specimen, Bean collection, ANU; F. Archaeomene sp., MMF23674. B–F from the Upper Jurassic Talbragar fish beds, Talbragar, NSW. Scale bars = 10 mm.
![Fig. 5 Jurassic fish: A. semionotiform fish from the Walloon Coal Measures (mid-Jurassic), Queensland [from Turner and Rozefelds (Citation1987) reproduced with permission of MQM]. B. Cavenderichthys talbragarensis, Australian Museum specimen AMF4133. C. Cavenderichthys talbragarensis, Geological Survey of New South Wales specimen MMF13561a. D. Coccolepis sp. AMF117880. E. Aphnelepis sp., unregistered specimen, Bean collection, ANU; F. Archaeomene sp., MMF23674. B–F from the Upper Jurassic Talbragar fish beds, Talbragar, NSW. Scale bars = 10 mm.](/cms/asset/9813dc18-1e1a-46b0-a97d-7f7289816492/sgff_a_392659_o_f0005g.gif)
Lepidotes Agassiz, 1832 is a genus of bony fish related to modern gars that inhabited both freshwater lakes and shallow seas. Some species exceeded 2 m in length and were covered by thick, enamelled scales. They possessed batteries of rounded, peg-like teeth, which enabled them to crush the shells of their molluscan prey. Such semionotiform fish are cosmopolitan in the Jurassic–Cretaceous (Thies Citation1989; Olsen & McClune Citation1991). Their wide geographic and aquatic distribution endows them with potential for development of a marine-to-continental-correlation scheme and a taxonomic review to this end is underway for the Jurassic (Turner Citation2008).
The Late Jurassic Talbragar fish fauna
Woodward's (Citation1895) assessment of fossil fish from Talbragar, near Gulgong, led to the first recognition of Jurassic beds in New South Wales. More recent studies by Bean (Citation2006a, Citation2006b) have provided new information on the faunal composition and palaeoenvironmental setting of these fossil fish beds.
“Leptolepis” talbragarensis Woodward Citation1895 (Fig. , C) is the most common fish species recorded in the Talbragar fauna. Long (Citation1991) considered the taxon to represent the first appearance of teleosteans in the Australian fossil record. However, doubt on generic assignment was clarified when Arratia (Citation1997) assigned the species to a new genus, Cavenderichthys. This taxonomic reappraisal and amendment was supported by Bean (Citation2006a) in her review of the actinopterygian taxa at Talbragar. Further, Woodward (Citation1895) had originally referred specimens to three species, “Leptolepis” talbragarensis, “L.” gregarious, and “L.” loweii. Arratia (Citation1997) considered all three to be synonymous and united them under C. talbragarensis, a concept that was first put forward by Wade (Citation1941). A detailed comparison of the species by one of us (LB) with taxa presently referred to Leptolepis, and also with other Late Jurassic forms, Tharsis dubius and Leptolepides sprattiformis, indicates that C. talbragarensis is most closely related to Late Jurassic members of the Leptolepididae.
Additionally, the Talbragar fauna includes to date one palaeoniscid, Coccolepis australis (Fig. ), and members of the “holostean” family Archaeomenidae, e.g. Archaeomaene tenuis (Fig. ), Aphnelepis australis (Fig. ) and Aetheolepis mirabilis (Bean Citation2006b). The genus Madariscus, erected by Wade (Citation1941) in the above family, is based on a series of imperfect specimens; those in the Australian Museum, Sydney, and the Natural History Museum, London, do not have enough detail to determine that they belong to a new genus, but, in general, they all seem to be very like a mature form of Archaeomene. The affinities of Uarbryichthys latus Wade Citation1941 are uncertain, but this taxon is no longer thought to be a macrosemiid and its affinities lie with the Macrosemiiformes (LB, work in progress). Bean (Citation2006a) found the Talbragar fish fauna to have the strongest faunal similarities with the assemblage from the Early Cretaceous (Aptian) Koonwarra biota of Victoria (e.g. Long Citation1991), since these both include a species of Coccolepis, a “leptolepid”, and archaeomenids, rather than taxa represented at older sites in the Clarence-Moreton Basin (see above).
During digs in 2006–07, several specimens of a larger amiiform caturidid fish similar to Furo Gistl, 1848, known from the Upper Jurassic of Europe and Asia (see Frickhinger Citation1996) were discovered. This new Talbragar fish grew to >30 cm, is covered with heavily enamelled scales and has a single row of teeth on both upper and lower jaws; it will be described in detail elsewhere (by LB).
The first Australian Jurassic chondrichthyan was found recently from the Talbragar fossil lagerstätten by S. Avery (NSW); this is a hybodontiform shark based on its squamation (ST pers. obs.) and will be described in due course. This exciting find underpins the potential for discovery of further freshwater or euryhaline forms (see below) and for correlation with the rich lagerstätten in Europe and elsewhere (Thies et al. Citation2007; Turner Citation2008).
The Talbragar fossil-fish assemblage indicates a Late Jurassic age (Bean Citation2006a, Citation2006b). This contrasts with Early to Middle Jurassic dates reported by most previous workers for the Purlawaugh Formation, with which the Talbragar Fossil Fish bed has been formerly associated. For example, Hind & Helby (Citation1969) recorded palynofloras referable to Evans' (Citation1966) J2 Palynozone (of Pliensbachian to early Toarcian age) from close to the base of the Purlawaugh Formation at another site near Gulgong. They recorded assemblages referable to Evans' (Citation1966) J3–J4 palynozones (of roughly mid-Toarcian to early Callovian age) from higher in the Purlawaugh Formation. No palynofloras have been recovered from the fish beds themselves but it is now apparent, based on faunal and SHRIMP (Sensitive High Resolution Ion Microprobe) dating, that these beds are somewhat younger than those parts of the Purlawaugh Formation from which the palynofloras were derived. The SHRIMP analysis of fossil fish bed samples, carried out at the ANU Research School of Earth Sciences, indicates an age for the youngest zircon population of 151.55 ± 4.27 Ma, or latest Oxfordian–Kimmeridgian–Tithonian in terms of GTS (Citation2004). The affinities of Cavenderichthys with Kimmeridgian forms in Europe accords with the isotopic date. This provides a maximum age for the strata and, given that the zircon grains show no sign of transport abrasion, suggests that the sediments were deposited at about this time. The occurrence of euhedral zircon crystals and the strongly silicified nature of the fossil fish beds support the idea, outlined further below, that ash from a felsic volcanic eruption filled the Talbragar lake and caused a mass fish kill. The fine detail preserved in both plants and fish not only promotes this view, but also implies an anaerobic burial environment.
The Talbragar Fossil Fish bed actually consists of several beds collectively 60–100 cm thick, which are divided into an upper package of very fine layers totalling 30–40 cm, and a lower rather more massive but still fine-grained bed, all of which are heavily fossiliferous (LB pers. obs.). The outcrop extends at least 200 m along strike and is thought to represent the erosional remnant of the margin of a freshwater lake-bed deposit (Percival Citation1979). Evidence for the cause of the mass death of fish now points to several anoxic events, including occasional eruptions of volcanic ash (Bean Citation2006a). The upper layers contain a high concentration of extremely well-preserved small fish, ranging in size from 4 to 12 cm, whereas the lower layer has scattered fish throughout, indicating a more normal lacustrine environment of deposition. There is little indication of desiccation, such as mud cracks or aerially exposed surfaces, so this part of the fish bed is not a mound spring deposit. Neither are the Talbragar Fossil Fish beds a typical overbank deposit, as such successions usually include coarse-grained beds with erosive bases (crevasse splay sediments), interspersed with palaeosols that developed during periodic exposure. The uppermost beds capping the main fossiliferous bed are very fine grained and represent an interval of slow deposition in still-water conditions that suddenly became anoxic.
The composition of the rock was described by Dulhunty & Eadie (Citation1969) as a hard, fine, limonitic cherty shale. However, recent thin-section and chemical analyses indicate that the clastic components of the sediments are largely reworked from underlying sandstones, which are interbedded with thin tuffs representing one or more very fine-grained ash falls (Bean Citation2006a). EDXA (Energy Dispersive X-ray Analysis) has shown no evidence of carbonate or calcium ions. The striking red-brown colour of the fossiliferous beds is post-depositional because each block has Liesegang rings associated with jointing. Manganese dioxide is commonly found infilling the fossil-fish cavities and commonly forms dendrites near block boundaries. Many of the fish and most of the plant impressions are replaced with white kaolinite and opaline quartz (Dr A. Christy pers. comm. to LB, see also White Citation1981b, Citation1986).
Recent excavations to the west of the best-known outcrop have exposed a continuation of the fish beds that has not been stained with iron oxide and is just a very fine, soft, white, and fossiliferous mudstone. This indicates that the staining of the rock and subsequent infilling of fish impressions are recent artefacts of ground-water movement (Bean, work in progress).
Fossil insects recovered from the Talbragar fossil fish beds include Cicada? lowei (Etheridge & Olliff Citation1890). Undescribed forms discovered recently were derived mainly from the southernmost extent of the deposit, although a few insects have been found in the most widely explored (northern) part of the outcrop. At least 50 insect specimens have been found, including a water boatman, rove beetle, stonefly larva, jumping plant louse, dragonfly naiads, lacewings, and assorted beetles, including at least one water beetle and one bark beetle. Small bivalve and gastropod molluscs have also been discovered, together with filamentous aquatic plants. All these taxa suggest a shallow-water environment at the lake margin (R. Beattie, pers. comm. to LB). Since none occur in the lower layers, where fish are more scattered, it seems most likely that the insects were trapped by the major ash fall that enveloped the body of water.
Tetrapods
Molnar and Thulborn (in Grant-Mackie et al. Citation2000) emphasised that Australian Jurassic dinosaur and other tetrapod body fossils are endemic. Fossil amphibians are rare. The upper jaw with alveoli of a very large, probably late Early to early Middle Jurassic “labyrinthodont” amphibian, Austropelor Longman, 1941, came from the Marburg Subgroup near Lowood, Clarence-Moreton Basin, and there is a reasonably complete skeleton of Siderops kehli from the Evergreen Formation in the Surat Basin (Grant-Mackie et al. Citation2000). Interestingly, these temnospondyl amphibians represent a phenomenon well known in southeastern Gondwana, namely that taxa persisted longer in this part of the world while becoming extinct elsewhere. This poses problems for inter-continental correlations based on higher-level taxa of terrestrial vertebrates. Another unusual occurrence is the association of freshwater plesiosaurs, which are typically marine elsewhere, but occur in Queensland in freshwater deposits, highlighted by Siderops in the Evergreen Formation (Surat Basin) and in the Razorback beds at Mt Morgan (e.g. Thulborn & Warren Citation1980; Grant-Mackie et al. Citation2000).
Only three dinosaur taxa have been found in the Jurassic (Bajocian) of Australia; one in the east, two in the west (Grant-Mackie et al. Citation2000; Rich & Vickers-Rich Citation2003). An incomplete skeleton of a sauropod, Rhoetosaurus brownei, was first found in the Hutton Sandstone in the 1920s, with subsequent collecting of parts of the same animal over 70 years. A single caudal centrum of a possible sauropod and a theropod, Ozraptor subotaii, have come from one site in the Bajocian Colalura Sandstone of Western Australia (Long Citation1998).
Traces of dinosaurs are far more common than body fossils in Australia. Footprints are more diverse and illuminating, confirming the age of the Precipice Sandstone in Queensland as Early Jurassic and suggesting a cosmopolitan distribution for ornithopod dinosaurs as early as the Early Jurassic (Beeston & Gray Citation1993). Many dinosaur footprints have been sporadically recorded in coal mines in the Clarence-Moreton Basin in southeastern Queensland, with a more recent dearth because of the shift from manual to automated longwall mining; those from the Walloon Coal Measures are around 168–161 Ma (Bathonian–Callovian: Grant-Mackie et al. Citation2000). Tracks of small to medium, agile, plant-eating dinosaurs with nails or “hooves” also occur (foot-print ichnotaxa include cf. Anomoepus, 20 cm long, Fig. ; ornithopods cf. Wintonopus, Fig. ). The latter type is widespread in the Early to Middle Jurassic worldwide (Grant-Mackie et al. Citation2000). Large three-toed and clawed predatory dinosaur prints (Fig. ; ichnotaxon Eubrontes, 54 cm long, Fig. ) from the Rosewood-Walloon coalfield resemble those of allosaurs and megalosaurs in the northern and western parts of Pangaea. A further major study of Jurassic dinosaur footprints at the Mt Morgan mines in central Queensland is soon to be published (CitationCook et al. in press).
Fig. 6 Examples of early to mid-Jurassic dinosaur footprint taxa: A. Reconstruction of possible fabrosaurid trackmaker, cf. Anomoepus from Lower Jurassic Precipice Sandstone, Carnarvon Gorge, Surat Basin. B. Large ornithopod footprint cf. Wintonopus, ca. 48 cm long, Balgowan Colliery, Dalby-Oakey coalfield, Darling Downs. C. Dinosaur footprint ca. 54 cm long, Balgowan Colliery, Dalbey-Oakey coalfield, Darling Downs. D. Large theropod footprint Eubrontes ca. 54 cm long, Oakleigh No 3 underground colliery. E. E. theropod footprint ca. 46 cm long, Lanefield Colliery, Rosewood-Walloon coalfield (Ball photo, courtesy of GSQ) B, D, from Walloon Coal Measures, Bathonian–Callovian, Clarence-Moreton basin (photos courtesy of and drawing ©T. Thulborn).

One further possibility for tetrapods in the Southern Hemisphere is the existence of a relict dicynodont population in eastern Gondwana (Thulborn & Turner Citation2003). All Jurassic basins, not just in Australia, will need to be examined for further evidence of a post-Triassic extension for these paramammals. Even mammalian remains might be expected.
Other Invertebrate faunas
Shelly marine faunas are restricted to the Western Australian marginal basins, and studied onshore assemblages are confined to the Middle and Late Jurassic (e.g. Playford et al. Citation1975; Playford et al. Citation1976; and see references in Grant-Mackie et al. Citation2000). The assemblages (particularly the ammonoids) have provided important tie points between Australian biostratigraphic zones and the international chronostratigraphic stages (Helby et al. Citation1987). Offshore Jurassic marine sequences are more continuous, but the limited material available from bore cores does not favour detailed macroinvertebrate studies. There is potential for work on microinvertebrates and shelly protists (e.g. Bown Citation1992, Grant-Mackie et al. Citation2000, Howe Citation2000), but the restricted marine conditions during deposition of some major units (e.g. the Dingo Claystone) and the general impoverishment in carbonate facies may limit scope for identifying diverse faunas of some groups. Jurassic invertebrate faunas are far better known from the neighbouring terranes of New Zealand, New Caledonia and New Guinea (Grant-Mackie et al. Citation2000; Haig et al. Citation2007; Hikuroa & Grant-Mackie Citation2008).
Four freshwater/estuarine bivalve species have been recorded from the eastern Australian Jurassic in a recent revision by Hocknull (Citation2000). These isolated records have yet to reveal significant biostratigraphic or biogeographical value (see Grant-Mackie et al. Citation2000).
Insects (Coleoptera and Blattaria) have been recovered from at least three sites in the Early Jurassic Cattamarra Coal Measures Member (Cockleshell Gully Formation), Perth Basin, Western Australia, and there appears to be scope for additional biogeographic research on this fauna (Jell Citation2004; Martin Citation2005). The Late Jurassic Talbragar fish bed (see above) also contains an apparently diverse insect fauna (R. Beattie, pers. comm), but, because the assemblage has not yet been thoroughly studied, its stratigraphic, palaeoenvironmental, and palaeobiogeographic significance cannot be evaluated here.
There is moderate potential for investigations of other non-marine fossil groups in the Australian Jurassic. For example, the oldest Australian leech cocoons have recently been described from the probable Pliensbachian Gatton Sandstone in the Clarence-Moreton Basin, which sits well above the appearance of Classopollis cf. chateaunovi that defines the Hettangian–Sinemurian boundary (Jansson et al. Citation2008). The durable proteinaceous cocoons of leeches and oligochaetes have been poorly studied, but they appear to be relatively common in the fossil record world-wide (Manum et al. Citation1991) and offer some potential as palaeoenvironmental or palaeobiogeographic indices once more records are published and their taxonomy is better resolved. Trace-fossil associations are also proving to be of substantial value as facies-demarcation tools within deltaic and shallow-marine sequences intersected in the course of hydrocarbon exploration along the Western Australian margin (Burns et al. Citation2001; Burns & Taylor Citation2005).
Discussion
Grant-Mackie et al. (Citation2000, pp. 344–345) concluded that “All marine faunas show clear Tethyan or eastern Tethyan affinities throughout the Jurassic along the northwestern and northern Australasian margins as far east as New Guinea and the Pahau Terrane”; and that “Whilst there is a clear and continuing Tethyan influence, there is in the Middle Jurassic a strong south Andean connection and a weaker Mexican link which continue into the Late Jurassic.” Further work is required to test whether this conclusion also applies to the terrestrial biota.
Australian Jurassic terrestrial fossil biotas are best represented in the eastern basins. Forests there were dominated successively by cheirolepidiacean, araucariacean, and podocarpacean conifers together with a range of seed-ferns. There are moderate differences between the Early Jurassic floras of Western and Eastern Australia, especially expressed in the distribution of the Exesipollenites association along the Tethyan margin (Grant-Mackie et al. Citation2000, Fig. 8). Later Jurassic assemblages are essentially pan-Australasian, possibly reflecting the higher latitude and more insular setting of Australia-New Zealand later in the period.
Few definitive palaeogeographic conclusions can be drawn from the sparse vertebrates, with only the possibility of a western Pacific zoogeographic province in the mid-Jurassic. In terms of the vertebrates providing useful global biostratigraphic, geochronologic, and palaeobiogeographic data, the fish of the Semionotidae were widespread in the Jurassic with several taxa already known from the Southern Hemisphere. As this group seems to have been capable of diadromy, detailed work on scales and teeth may assist in both improved zonation of continental rocks and better marine to non-marine correlation. Likewise, a good global record for non-marine sharks is emerging, with teeth of hybodonts showing potential for a refined stratigraphy (Thies et al. Citation2007).
Work is required to better correlate marine/non-marine sequences along the Western Australian margin. There are also shortcomings in the correlation of latest Middle to Late Jurassic units in eastern Australian basins. Marine microfossils are unavailable in these basins and palynozones for this interval are relatively long-ranging, hence more details of spore–pollen taxa and their ranges are required for better biostratigraphic resolution.
Conclusions and Future Directions of Research
The main problem for Southern-Hemisphere biostratigraphers emanates from the GTS (Citation2004) being Euramerican in its focus. Despite this difficulty, every effort is being undertaken to relate Australian zonal and other biostratigraphic data to it. In terms of the IGCP 506 Project, correlations and interpolations of Australian Jurassic zonations, based on this timescale, are required, providing a specific task for local biostratigraphers during the life of the project and beyond. However, many deficiencies exist in the broad spectrum of Australian biostratigraphy, and not all of them will be overcome in the short-term, because there is a serious shortage of expertise in all areas of the science. This will be exacerbated, in the next few years, by the retirement of several specialists, especially in palynology.
In the Australian region, marine sedimentation, during the Jurassic, was largely restricted to the Western Australian marginal basins. Some of the deposits therein constitute important hydrocarbon source rocks. Onshore thereto, there is very limited exposure of marine Jurassic rocks, which are known mostly from the northern Perth Basin. In the onshore subsurface, they are present in parts of the Carnarvon and Canning basins. Offshore, marine Jurassic strata are very extensive and known from hundreds of oil-exploration wells from the Carnarvon Basin northwards to the Timor Sea. In this area, particularly on the North West Shelf, palynology is presently used almost exclusively in sequences older than mid-Cretaceous and, as inferred above, a great deal of economic activity depends on it. Non-marine units (also deposited in the west) are extensively developed in eastern Australia, where marine strata are not known to occur, although, in northern Queensland (northeastern Australia), in the Laura and Carpentaria basins, the latest Jurassic-Neocomian Gilbert River Formation is lagoonal to marginal marine, and the poorly understood, Middle Jurassic-Early Cretaceous Helby beds, in the western part of the latter basin, are marine (Day et al. Citation1975; Bain & Draper Citation1997).
GA, through their Timescales and VCEMP Project, is relating biozonations to GTS (Citation2004). This national organisation has largely completed the update of the integrated spore–pollen and dinocyst zonations of Helby et al. (Citation1987, Citation2004), and their conversion to the international timescale (e.g. Partridge Citation2006b). McKellar (in press) has similarly documented the litho- and palynostratigraphic zonation of the continental Jurassic in the Surat Basin (eastern Australia), employing, for the Early–Middle Jurassic, correlation with the spore–pollen floras of New Zealand and New Caledonia, in order to establish relationships with the international timescale at stage level. The palynofloras of these regions have been dated by associated marine invertebrates (Grant-Mackie et al. Citation2000; de Jersey & Raine Citation2002). Further, N. de Jersey and JMcK have now delimited the Triassic–Jurassic (Rhaetian–Hettangian) and Hettangian–Sinemurian boundaries in the eastern Clarence-Moreton Basin (eastern Australia), in a rather exceptional continental section that apparently embraces the period/system transition without the unconformity that generally characterises the stratigraphic record at that level, based on correlation with spore–pollen floras from accurately dated marine strata in New Zealand.
As yet, no extensive isotopic-dating or magnetostratigraphic studies have been undertaken for the Australian Jurassic. Such investigations would assist calibration of Australian biozonations and complement both intra- and international biostratigraphic correlations and contribute to the establishment of relationships with the international timescale. However, with magnetostratigraphy, its expense and the availability of suitable material are limiting factors. From a biostratigraphic perspective, the most likely ways to tie Australian sequences to the Euramerican schemes will be using a combination of dinoflagellate, spore–pollen, foraminiferal, nannoplankton, and ammonite data.
The palynological zones, mainly the dinocyst zones, are now highly sophisticated and accurate within any given basin, and provide a level of resolution that will not be matched by other biostratigraphic methods in the foreseeable future. Dinoflagellates and miospores have the advantage that they are recoverable from a wide range of strata that are substantially devoid of other fossil groups. However, there is a need for improved correlation between regional Australian zonations based on them, and between the marine (dinocyst) and continental (spore–pollen) schemes, to facilitate improved dating of the latter, but this is inhibited by the commonly low abundances of miospores in marine strata.
Further to the spore–pollen studies already undertaken on the ammonite-dated (Hettangian–Sinemurian), New Zealand sequences (cited above) and additional to the work of Helby et al. (Citation1988) on dinoflagellate associations from the late Middle and Late Jurassic of that region (which were interpreted in terms of Australian zones), there is potential for obtaining similarly useful dinocyst data from the region's intervening Jurassic, for which ammonite data are also wanting. Ammonites in Western Australia are not likely to provide much more information than is already available, as all the known localities have been dated and only occasional finds are made in cores. However, foraminifera and nannofossil data are presently wanting.
For the Australian Jurassic, the following points are also highlighted:
| • | Fish may possibly assist with marine to non-marine correlation. | ||||
| • | A satisfactory definition for the J–K boundary has not yet been found, as provincialism in ammonites, caused by Late Jurassic regression in northern Europe, has fuelled debates on Tethyan vs. Boreal distributions. Also, this boundary is mostly embraced by non-marine successions in Australia, so its identification must rely on good correlation of marine and non-marine zonations. | ||||
| • | The unpublished cross-Tasman compilation of de Jersey and JMcK provides accurate palynostratigraphic location of the Triassic–Jurassic (Rhaetian–Hettangian) and Hettangian–Sinemurian boundaries in continental eastern Australia, but more extensive work is required to clarify the positions of other stage boundaries. However, it is likely that they may not necessarily coincide with local biostratigraphic, or, for that matter, lithostratigraphic boundaries. | ||||
| • | The integrated dinocyst and spore–pollen zonations (viz. Helby et al. Citation1987, Citation2004, Partridge Citation2006b) may provide resolution to sub-million year intervals, allowing recognition of high-frequency sea-level changes and enabling better characterisation of the evolution of sedimentary facies pertinent to the search for stratigraphic hydrocarbon traps. | ||||
| • | There are 5–10 km of both marine and non-marine Jurassic strata evident in seismic profiles on the North West Shelf, so the challenge for the years to come under the auspices of IGCP 506 and its successors will be to prepare an integrated high-resolution biostratigraphy calibrated to the European standard. | ||||
Acknowledgements
Professor Sha Jingeng (co-leader IGCP 506, NIGPAS) provided valuable support to enable the beginnings of this paper given by ST at the 1st IGCP 506 Symposium in Nanjing; the Australian IGCP Committee Grant-in-Aid Scheme and UNESCO-IUGS IGCP 506 are thanked for support to attend the 2005 and 2008 symposia; she and MD thank the Board of the Queensland Museum for basic support. LBB would like to thank Professor KSW Campbell (ANU), Dr GC Young (ANU) and Prof. JA Long (Museum Victoria, Melbourne) for advice on fish; Dr IS Williams (Research School of Earth Sciences, ANU, Canberra) for expertise and help with the SHRIMP analysis; Dr A. Christy (ANU) for carrying out SEM/EXDA (scanning electron microscope and energy dispersive X-ray analysis) studies; Professor RJ Arculus (ANU) for help with histological work; R. Beattie for information about fossil insects. SM was supported by Australian Research Council Discovery Grant A10020305 and a Swedish Research Council grant and a Riksmusei Vänner stipendium. We thank Drs Elizabeth Truswell (ANU, Canberra) and Eric Monteil (formerly GA, Canberra) for their helpful reviews. Further, Dr John Backhouse (University of Western Australia) provided extensive comments on the Jurassic of Western Australia, especially on the North West Shelf, and the future direction of biostratigraphic and related work in the Mesozoic of Australia; many of his comments have been incorporated into the final section. We thank Natalie Sinclair (ANU, GA) for providing a resumé of her doctoral thesis work and other data on Western Australia and Dr Daniel Mantle (GA) for providing information on the palynology of the Bonaparte basin derived largely from his unpublished PhD thesis.
Notes
This is a contribution to IGCP 506: Marine/Non-marine Jurassic correlation.
Related Research Data
References
- AGSO., 1996: Phanerozoic Timescale Wallchart: Australian and International Biozonations. Version 96-1i-7. Compiled by the Australian Geological Survey Organisation Timescale Calibration and Development Project Team, Canberra
- Anderson , D.L. 1982 . Hotspots, polar wander, Mesozoic convection and the geoid . Nature , 297 : 391 – 393 .
- Anderson , D.L. 1989 . Theory of the earth , 366 Boston : Blackwell Scientific Publications .
- Anderson , D.L. 2007 . New theory of the earth , 384 Cambridge : Cambridge University Press .
- Arratia , G. 1997 . Basal teleosts and teleostean phylogeny . Palaeo Ichthyologica , 7 : 1 – 168 .
- Backhouse , J. 1978 . Palynological zonation of the Late Jurassic and Early Cretaceous sediments of the Yarragadee formation, central Perth basin, Western Australia . : 53 Geological Survey of Western Australia, Report 7
- Bain , J.H.C. and Draper , J.J. , eds. 1997 . “ North Queensland geology ” . In Australian Geological Survey Organisation , Bulletin 240; and Queensland Department of Mines and Energy, Queensland Geology 9 600
- Balme, B.E., 1957: Spores and pollen grains from the Mesozoic of Western Australia Commonwealth Scientific and Industrial Research Organization, Australia, Coal Research Section, Technical Communication 25, 48 pp
- Balme , B.E. 1964 . “ The palynological record of Australian pre-Tertiary floras ” . In Ancient Pacific floras – the pollen story , Papers presented at the Tenth Pacific Science Congress Edited by: Cranwell , L.M. Vol. 1961 , 49 – 80 . Honolulu : University of Hawaii Press .
- Bamford , M.K. and Philippe , M. 2001 . Jurassic – Early Cretaceous Gondwanan homoxylous woods: A nomenclatural revision of the genera with taxonomic notes . Review of Palaeobotany and Palynology , 113 : 287 – 297 .
- Barnard , P.D.W. 1973 . “ Organisms and continents through time ” . In Special Papers in Palaeontology Edited by: Hughes , N.F. Vol. 12 , 175 – 187 .
- Bazard , D.R. and Butler , R.F. 1991 . Paleomagnetism of the Chinle and Kayenta formations, New Mexico and Arizona . Journal of Geophysical Research , 96 ( B6 ) : 9847 – 71 .
- Bean , L.B. 2006a . A new look at Cavernderichthys talbragarensis and other fish from the Talbragar fish bed . Alcheringa Special Issue , 1 : 433
- Bean , L.B. 2006b . The leptolepid fish Cavenderichthys talbragarensis from the Talbragar Fish Bed (Late Jurassic) near Gulgong, New South Wales . Records of the Western Australian Museum , 23 : 43 – 76 .
- Beeston , J. and Gray , A. 1993 . The Ancient rocks of Carnarvon gorge , 48 Brisbane : Department of Minerals and Energy .
- Bown , P.R. 1992 . New calcareous nannofossil taxa from the Jurassic–Cretaceous boundary interval of Sites 765 and 261, Argo Abyssal Plain . Proceedings of the Ocean Drilling Program, Scientific Results , 123 : 369 – 379 .
- Bradshaw, M. & Yeung, M., 1990: The Jurassic palaeogeography of Australia. Bureau of Mineral Resources, Australia, Record 1990/76, Palaeogeography 26, 1–60
- Bradshaw , M.T. and Yeung , M. 1992 . Palaeogeographic Atlas of Australia. Volume 8 – Jurassic , 12 Australia : Bureau of Mineral Resources .
- Bromfield , K. , Burrett , C.F. , Leslie , R.A. and Meffre , S. 2007 . Jurassic volcaniclastic–basaltic andesite-dolerite sequence in Tasmania: New age constraints for fossil plants from Lune River . Australian Journal of Earth Sciences , 54 : 965 – 974 .
- Burger, D., 1968: Stratigraphy and palynology of upper Mesozoic sections in some deep wells in the Surat basin, Queensland. Bureau of Mineral Resources, Australia, Record 1968/24. 1–35
- Burger , D. 1973 . “ Spore zonation and sedimentary history of the Neocomian, Great Artesian basin, Queensland ” . In Mesozoic and Cainozoic palynology: Essays in honour of Isabel Cookson , Geological Society of Australia, Special Publication 4 Edited by: Glover , J.E. and Playford , G. 87 – 118 .
- Burger , D. 1976 . “ Palynological observations in the Surat basin ” . In A guide to the geology of the Bowen and Surat basins in Queensland , 25th International Geological Congress, Sydney, Excursion Guide 3C Edited by: Jensen , A.R. , Exon , N.F. , Anderson , J.C. and Koppe , W.H. 51 – 54 . Appendix 5
- Burger, D., 1984: A palynological review of the Jurassic below the Injune Creek Group in the Eromanga basin, Queensland. Bureau of Mineral Resources, Australia, Record 1984/19. 1–23
- Burger , D. 1986 . Palynology, cyclic sedimentation, and palaeoenvironments in the late Mesozoic of the Eromanga basin . Geological Society of Australia, Special Publication 12 , : 53 – 70 .
- Burger, D., 1989: Stratigraphy, palynology, and palaeoenvironments of the Hooray Sandstone, eastern Eromanga basin, Queensland and New South Wales. Queensland Department of Mines, Report 3, 28 pp
- Burger , D. 1994a . “ Palynological studies of the Bundamba group and Walloon coal measures in the Clarence-Moreton basin ” . In Geology and petroleum potential of the Clarence-Moreton basin, New South Wales and Queensland , Australian Geological Survey Organisation, Bulletin 241 Edited by: Wells , A.T. and O'Brien , P.E. 164 – 180 .
- Burger , D. 1994b . “ Palynology of the uppermost Walloon Coal Measures, Kangaroo Creek Sandstone, and Grafton Formation, Clarence-Moreton basin ” . In Geology and petroleum potential of the Clarence-Moreton basin, New South Wales and Queensland , Australian Geological Survey Organisation, Bulletin 241 Edited by: Wells , A.T. and O'Brien , P.E. 181 – 188 .
- Burger , D. and Senior , B.R. 1979 . A revision of the sedimentary and palynological history of the northeastern Eromanga basin, Queensland . Journal of the Geological Society of Australia , 26 : 121 – 132 .
- Burger , D. and Shafik , S. 1996 . “ Jurassic (Charts 8 and 9) ” . In Australian Phanerozoic timescales , Edited by: Young , G.C. and Laurie , J.R. 148 – 159 . Canberra : AGSO .
- Burger, D., Foster, C.B. & McKellar, J.L., 1992: A review of Permian to Cretaceous palynostratigraphy in eastern Australia. Bureau of Mineral Resources, Australia, Record 1992/5. 1–26
- Burns, F. & Taylor, K., 2005: Trace fossils and their application to high-resolution sequence stratigraphy and associated cement distribution; Middle Jurassic and Lower Cretaceous interval, North West Shelf. Minerals and Energy Research Institute of Western Australia Report 241 (on CD-ROM)
- Burns , F. , Thompson , J. , Ainsworth , R.B. and Seggie , R. 2001 . “ Depositional modeling of a deltaic system using ichnofabric analysis; Middle Jurassic Plover Formation, Sunrise-Troubadour gas-condensate fields, Bonaparte basin, Timor Sea ” . In American Association of Petroleum Geologists , Annual Meeting Expanded Abstracts, June 2001 30
- Clifford , H.T. 1998 . First record of Caytonia in Australia . Memoirs of the Queensland Museum , 42 : 448
- Cook, A.G., Saini, N. & Hocknull, S.A., Dinosaur footprints from the Lower Jurassic of Queensland. Memoirs of the Queensland Museum (in press)
- Cranfield , L.C. , Carmichael , D.C. and Wells , A.T. 1994 . “ Ferruginous oolite and associated lithofacies from the Clarence-Moreton and related basins in southeast Queensland ” . In Geology and petroleum potential of the Clarence-Moreton basin, New South Wales and Queensland , AGSO Bulletin 241 Edited by: Wells , A.T. and O'Brien , P.E. 144 – 163 .
- Day , R.W. 1964 . Stratigraphy of the Roma-Wallumbilla area . Geological Survey of Queensland Publication 318 , : 1 – 23 .
- Day , R.W. , Whitaker , W.G. , Murray , C.G. , Wilson , I.H. and Grimes , K.G. 1975 . Queensland geology – A companion volume to the 1:2 500 000 scale geological map . Geological Survey of Queensland Publication 383 , : 194
- de Jersey , N.J. 1975 . “ Miospore zones in the lower Mesozoic of southeastern Queensland ” . In Gondwana geology , Papers presented at the Third Gondwana Symposium, Canberra, Australia, 159–172. 1973 Edited by: Campbell , K.S.W. Australian National University Press .
- de Jersey , N.J. 1976 . Palynology and time relationships in the lower Bundamba Group (Moreton basin) . Queensland Government Mining Journal , 77 : 460 – 465 .
- de Jersey , N.J. and Raine , J.I. 2002 . “ Early to Middle Jurassic miospore zonation ” . In IPC 2002 , Geological Society of Australia, Abstracts 68 41 – 42 .
- Dulhunty , J.A. and Eadie , J. 1969 . Geology of the Talbragar fossil fish bed area . Journal and Proceedings of the Royal Society of New South Wales , 102 : 1 – 4 .
- Du Toit , A.L. 1937 . Our wandering continents – an hypothesis of continental drifting , 366 Edinburgh : Oliver and Boyd .
- Ekstrand , E.J. and Butler , R.F. 1989 . Paleomagnetism of the Moenave Formation: Implications for the Mesozoic North American apparent polar wander path . Geology , 17 : 245 – 248 .
- Embleton , B.J.J. 1981 . “ A review of the paleomagnetism of Australia and Antarctica ” . In Paleoreconstruction of the continents , Geodynamics Series 2, American Geophysical Union Edited by: McElhinny , M.W. and Valencio , D.A. 77 – 92 .
- Etheridge , R.J. and Olliff , A.S. 1890 . The Mesozoic and Tertiary insects of New South Wales . Memoirs of the Geological Survey of New South Wales, Palaeontology , 7 : 1 – 12 .
- Evans, P.R., 1965: Recent advances in Mesozoic stratigraphic palynology in Australia. Bureau of Mineral Resources, Australia, Record 1965/192, 26 pp
- Evans , P.R. 1966 . Mesozoic stratigraphic palynology in Australia . Australasian Oil and Gas Journal , 12 ( 6 ) : 58 – 63 .
- Evans , P.R. and Roberts , J. 1980 . Evolution of central eastern Australia during the late Palaeozoic and early Mesozoic . Journal of the Geological Society of Australia , 26 : 325 – 340 .
- Exon , N.F. 1976 . Geology of the Surat basin in Queensland . Bureau of Mineral Resources, Australia, Bulletin 166 , : 160
- Exon , N.F. 1980 . The stratigraphy of the Surat basin, with special reference to coal deposits . Coal Geology , 1 ( 3 ) : 57 – 69 .
- Exon , N.F. and Burger , D. 1981 . Sedimentary cycles in the Surat basin and global changes of sea level . Bureau of Mineral Resources Journal of Australian Geology and Geophysics , 6 : 153 – 159 .
- Exon , N.F. and Colwell , J.B. 1994 . Geological history of the outer North West Shelf of Australia: A synthesis . AGSO Journal of Australian Geology and Geophysics , 15 : 177 – 190 .
- Exon , N.F. and von Rad , U. 1994 . “ The Mesozoic and Cainozoic sequences of the northwest Australian margin, as revealed by ODP core drilling and related studies ” . In The sedimentary basins of Western Australia , Proceedings of the Petroleum Exploration Society of Australia Symposium, Perth Edited by: Purcell , P.G. and Purcell , R.R. 181 – 199 .
- Fielding , C.R. 1996 . “ Mesozoic sedimentary basins and resources in eastern Australia – a review of current understanding ” . In Mesozoic 96, Mesozoic Geology of the Eastern Australia Plate Conference, Brisbane, 1996 , Geological Society of Australia, Extended Abstracts Vol. 43 , 180 – 185 .
- Filatoff , J. 1975 . Jurassic palynology of the Perth basin . Palaeontographica 154B , : 1 – 113 .
- Filatoff , J. and Price , P.L. 1988 . A pteridacean spore lineage in the Australian Mesozoic . Association of Australasian Palaeontologists Memoir , 5 : 89 – 124 .
- Frakes , L.A. , Francis , J.E. and Syktus , J.I. 1992 . Climate modes of the Phanerozoic: The history of the Earth's climate over the past 600 million years , 274 Cambridge : Cambridge University Press .
- Frickhinger, K.A., 1991: Fossilien Atlas Fische. Mergus, Verlag fur Natur- und Heimtierkunde Hans A. Baensch, Melle. 1088 pp. [English Edition 1996]
- Fritts , H.C. 1976 . Tree rings and climate , 567 London : Academic Press .
- Gallagher , S.M. , Wood , G.R. and Lemon , N.L. 2008 . “ Birkhead Formation chronostratigraphy on the Murteree Horst, South Australia: identifying and correlating reservoirs and seals ” . In Eastern Australasian basins Symposium III , Petroleum Exploration Society of Australia, Special Publication Edited by: Blevin , J.E. , Bradshaw , B.E. and Uruski , C. 169 – 189 .
- Gee , C.T. 1989 . Revision of the Late Jurassic/Early Cretaceous flora from Hope Bay, Antarctica . Palaeontographica , 213B : 149 – 214 .
- Geoscience Australia, 2002: Palaeogeographic Atlas of Australia (a set of ten volumes). Published on CD-ROM by Geoscience Australia, Canberra
- Glauert , L. 1923 . Notes on fossil plants from Mingenew and Irwin River . Journal of the Royal Society of Western Australia , 10 : 7 – 11 .
- Gold , T. 1955 . Instability of the Earth's axis of rotation . Nature , 175 : 526 – 529 .
- Gordon , R.G. , Cox , A. and O'Hare , S. 1984 . Paleomagnetic Euler poles and the apparent polar wander and absolute motion of North America since the Carboniferous . Tectonics , 3 : 499 – 537 .
- Goscombe , P.W. and Coxhead , B.A. 1995 . “ Clarence-Moreton, Surat, Eromanga, Nambour, and Mulgildie basins ” . In Geology of Australian Coal basins , Geological Society of Australia Coal Geology Group Special Publication 1 Edited by: Ward , C.R. , Harrington , H.J. , Mallett , C.W. and Beeston , J.W. 489 – 511 .
- Gould , R.E. 1968 . Morphology of Equisetum laterale Phillips, 1829, and E. bryanii sp. nov. from the Mesozoic of south-eastern Queensland . Australian Journal of Botany , 16 : 153 – 176 .
- Gould , R.E. 1971 . “ Report on plant fossils from Durikai, southeastern Queensland, Appendix 3 ” . In The post-Palaeozoic rocks of the Warwick 1:250 000 Sheet area, Queensland and New South Wales , Bureau of Mineral Resources, Geology and Geophysics, Australia, Report 140 Edited by: Exon , N.F. , Reiser , R.F. , Casey , D.J. and Brunker , R.L. 63 – 64 .
- Gould , R.E. 1972 . Cibotium tasmanense sp. nov. a fossil tree fern from the Tertiary of Tasmania . Australian Journal of Botany , 20 : 119 – 126 .
- Gould , R.E. 1975 . The succession of Australian pre-Tertiary megafossil floras . Botanical Review , 41 : 453 – 483 .
- Gould , R.E. 1978 . “ Report on plant fossils from the Tarlton Range, Tobermory 1:250 000 Sheet area, Northern Territory. Appendix ” . In Geology of the Eromanga Basin , Bureau of Mineral Resources, Geology and Geophysics, Australia, Bulletin 167 Edited by: Senior , B.R. , Mond , A. and Harrison , P.L. 87 – 88 .
- Gould , R.E. 1980 . The coal-forming flora of the Walloon coal measures . Australian Coal Geology , 1 : 83 – 105 .
- Gould , R. and Shibaoka , M. 1980 . Some aspects of the formation and petrographic features of coal members in Australia, with special reference to the Tasman Orogenic Zone . Coal Geology (Australia) , 2 : 1 – 29 .
- Gradstein , F.M. and von Rad , U. 1991 . Stratigraphic evolution of Mesozoic continental margin and oceanic sequences: Northwest Australia and northern Himalayas . Marine Geology , 102 : 131 – 173 .
- Gradstein , F.M. , Ogg , J.G. and Smith , A.G. 2004 . A Geologic Time Scale 2004 , 589 Cambridge : Cambridge University Press .
- Gradstein , F.M. , Agterberg , F.P. , Ogg , J.G. , Hardenbol , J. , van Veen , P. , Thierry , J. and Huang , Z. 1994 . A Mesozoic timescale . Journal of Geophysical Research , 15 : 24051 – 24074 .
- Gradstein , F.M. , von Rad , U. , Gibling , M.R. , Jansa , L.F. , Kaminski , M.A. , Kristiansen , I.-L. , Ogg , J.G. , Röhl , U. , Sarti , M. , Thurow , J.W. , Westermann , G.E.G. and Wiedmann , J. 1992 . Stratigraphy and depositional history of the Mesozoic continental margin of central Nepal . Geologisches Jahrbuch, Reihe B , 77 : 3 – 141 .
- Grant-Mackie , J.A. , Aita , Y. , Balme , B.E. , Campbell , H.J. , Crallinor , A.B. , MacFarlane , D.A.B. , Molnar , R.E. , Stevens , G.R. and Thulborn , R.A. 2000 . Jurassic palaeobiogeography of Australasia . Association of Australasian Palaeontologists Memoir , 23 : 311 – 53 .
- Haig , D.W. , McCartain , E. , Barber , L. and Backhouse , J. 2007 . Triassic–lower Jurassic foraminiferal indices for Bahamian-type carbonate-bank limestones, Cablac Mountain, East Timor . Journal of Foraminiferal Research , 37 : 248 – 64 .
- Hälbich , I.W. , Fitch , F.J. and Miller , J.A. 1983 . “ Dating the Cape Orogeny ” . In Geodynamics of the Cape Fold Belt: a contribution to the National Geodynamics Programme , Geological Society of South Africa, Special Publication 12 Edited by: Söhnge , A.P.G. and Hälbich , I.W. 149 – 164 .
- Hallam , A. 1975 . Jurassic Environments , 269 Cambridge : Cambridge University Press .
- Hallam , A. 1985 . A review of Mesozoic climates . Journal of the Geological Society of London , 142 : 433 – 445 .
- Hallam , A. 1996 . “ Major bio-events in the Triassic and Jurassic ” . In Global Events and Event Stratigraphy , Results of IGCP 216 Global Biological Events in Earth History Edited by: Walliser , O.H. 265 – 283 . Berlin : Springer .
- Haq , B.U. and Al-Qahtani , A.M. 2005 . Phanerozoic cycles of sea-level change on the Arabian Platform . GeoArabia , 10 : 127 – 160 .
- Haq , B.U. , Hardenbol , J. and Vail , P.R. 1987 . Chronology of fluctuating sea levels since the Triassic . Science , 235 : 1156 – 1167 .
- Hawlader , H.M. 1990 . Diagenesis and reservoir potential of volcanogenic sandstones – Cretaceous of the Surat basin, Australia . Sedimentary Geology , 66 : 181 – 195 .
- He , F. and Conaghan , P.J. 1994 . Diagenesis of Jurassic and Lower Cretaceous sandstones of the Eromanga basin in New South Wales . AGSO Journal of Australian Geology and Geophysics , 15 : 191 – 215 .
- Heine , C. and Müller , R.D. 2005 . Late Jurassic rifting along the Australian North West Shelf: Margin geometry and spreading ridge configuration . Australian Journal of Earth Sciences , 52 : 27 – 39 .
- Helby , R. , Morgan , R. and Partridge , A.D. 1987 . A palynological zonation of the Australian Mesozoic . Association of Australasian Palaeontologists Memoir , 4 : 1 – 94 .
- Helby , R. , Wilson , G.J. and Grant-Mackie , J.A. 1988 . “ A preliminary biostratigraphic study of Middle to Late Jurassic dinoflagellate assemblages from Kawhia, New Zealand ” . In Palynological and Palaeobotanical Studies in Honour of Basil E. Balme , Association of Australasian Palaeontologists Memoir 5 Edited by: Jell , P.A. and Playford , G. 125 – 166 .
- Helby , R. , Morgan , R. and Partridge , A.D. 2004 . Updated Jurassic-Early Cretaceous dinocyst zonation for the North West Shelf of Australia Geoscience Australia Publication, ISBN 1 920871 01 2+ http://www.ga.gov
- Hergt , J.M. , McDougall , I. , Banks , M.R. and Green , D.H. 1989 . “ Igneous Rocks: Jurassic dolerite ” . In Geology and Mineral Resources of Tasmania , Geological Society Special Publication 15 Edited by: Burrett , C.F. and Martin , E.L. 375 – 381 .
- Hikuroa , D.C.H. and Grant-Mackie , J.A. 2008 . New species of Late Jurassic Australobuchia (Bivalvia) from the Murihiku Terrane, Port Waikato – Kawhia region, New Zealand . Alcheringa , 32 : 73 – 98 .
- Hill , D. and Maxwell , W.G.H. 1967 . Elements of the Stratigraphy of Queensland , 2nd ed , 78 Brisbane : University of Queensland Press .
- Hill , D. , Playford , G. and Woods , J.T. 1966 . Jurassic Fossils of Queensland , 32 Brisbane : Queensland Palaeontographical Society .
- Hill , R.S. , Truswell , E.M. , McLoughlin , S. and Dettmann , M.E. 1999 . “ The evolution of the Australian flora: fossil evidence ” . In Flora of Australia , 2nd ed. , Edited by: Orchard , A.E. 251 – 320 . Melbourne : ABRS/CSIRO . 1 (Introduction)
- Hind , M.C. and Helby , R.J. 1969 . “ The Great Artesian basin in New South Wales ” . In Geology of New South Wales. Journal of the Geological Society of Australia Edited by: Packham , G.H. 481 – 497 . 16(1)
- Hocknull , S.A. 2000 . Mesozoic freshwater and estuarine bivalves from Australia . Memoirs of the Queensland Museum , 45 : 405 – 426 .
- Howe, R.W., 2000: Jurassic calcareous nannofossil biostratigraphy of the North West Shelf and Timor Sea areas. Minerals and Energy Research Institute of Western Australia Report 213 (on CD-ROM)
- Hunter , M.A. , Cantrill , D.J. , Flowerdew , M.J. and Millar , I.L. 2005 . Mid-Jurassic age for the Botany Bay Group: Implications for Weddell Sea basin creation and southern hemisphere biostratigraphy . Journal of the Geological Society, London , 162 : 745 – 748 .
- Jansson , I.-M. , McLoughlin , S. and Vajda , V. 2008 . Early Jurassic annelid cocoons from eastern Australia . Alcheringa , 32 : 285 – 296 .
- Jansson , I.-M. , McLoughlin , S. , Vajda , V. and Pole , M. 2008 . An Early Jurassic flora from the Clarence-Moreton basin, Australia . Review of Palaeobotany and Palynology , 150 : 5 – 21 .
- Jell , P.A. 2004 . The fossil insects of Australia . Memoirs of the Queensland Museum , 50 : 1 – 124 .
- Jones , J.G. and Veevers , J.J. 1983 . Mesozoic origins and antecedents of Australia's Eastern Highlands . Journal of the Geological Society of Australia , 30 : 305 – 322 .
- Jones , J.G. and Veevers , J.J. 1984 . “ Eastern Highlands ” . In Phanerozoic Earth History of Australia , Edited by: Veevers , J.J. 115 – 143 . New York : Oxford University Press .
- Klootwijk, C., 1996: Phanerozoic configurations of Greater Australia: evolution of the North West Shelf. Part Two: Palaeomagnetic and geologic constraints on reconstructions. Australian Geological Survey Organisation, Record 1996/52. 1–85
- Korsch , R.J. and Totterdell , J.M. 1996 . “ Mesozoic deformational events in eastern Australia and their impact on onshore sedimentary basins ” . In Mesozoic 96, Mesozoic Geology of the Eastern Australia Plate Conference, Brisbane, 1996 , Geological Society of Australia, Extended Abstracts 43 308 – 312 .
- Krassilov , V. 1981 . Changes in Mesozoic vegetation and the extinction of the dinosaurs . Palaeogeography, Palaeoclimatology, Palaeoecology , 34 : 207 – 224 .
- Kutzbach , J.E. and Ziegler , A.M. 1993 . Simulation of Late Permian climate and biomes with an atmosphere-ocean model: Comparisons with observations . Philosophical Transactions of the Royal Society of London 341B , : 327 – 340 .
- Long , J.A. 1991 . “ The long history of Australian fossil fishes ” . In Vertebrate Palaeontology of Australasia , Edited by: Vickers-Rich , P. , Monaghan , J.N. , Baird , R.F. and Rich , T.H. 337 – 428 . Melbourne : Monash University Publications Committee .
- Long , J.A. 1995 . The Rise of Fishes , 223 Sydney : University of New South Wales Press .
- Long , J.A. 1998 . Dinosaurs of Australia and New Zealand and other animals of the Mesozoic Era , 188 Sydney : University of New South Wales Press .
- Longley , I.M. , Buessenschuett , C. , Clydsdale , L. , Cubitt , C.J. , Davis , R.C. , Johnson , M.K. , Marshall , N.M. , Murray , A.P. , Somerville , R. , Spry , T.B. and Thompson , N.B. 2002 . “ The North West Shelf of Australia – a Woodside perspective ” . In The Sedimentary basins of Western Australia , Proceedings of the Petroleum Exploration Society of Australia Symposium, Perth, WA Edited by: Keep , M. and Moss , S.J. Vol. 3 , 27 – 88 .
- Mantle, D., 2006: Palynology, sequence stratigraphy, and palaeoenvironments of Middle to Late Jurassic strata, Bayu-Undan Field, Timor Sea region. Ph.D. thesis, The University of Queensland. 210 pp
- Manum , S.B. , Bose , M.N. and Sawyer , R.T. 1991 . Clitellate cocoons in freshwater deposits since the Triassic . Zoologica Scripta , 20 : 347 – 366 .
- Marcano , M.C. , van der Voo , R. and Mac Niocaill , C. 1999 . True polar wander during the Permo-Triassic . Journal of Geodynamics , 28 : 75 – 95 .
- Martin , S.K. 2005 . “ Insects of the Early Jurassic Cattamarra Coal Measures, Western Australia; pinpointing the origins of a modern fauna ” . In 19th Victorian Universities Earth and Environmental Sciences Conference; VUEESC , Geological Society of Australia Abstracts Series 80 Edited by: Corvino , A.F. and Ely , K.S. 18
- McKellar , J.L. 1981 . Palynostratigraphy of samples from the Narangba area, Nambour basin . Queensland Government Mining Journal , 82 : 268 – 273 .
- McKellar , J.L. 1996 . “ Palynofloral and megafloral indications of palaeoclimate in the Late Triassic, Jurassic, and early Cretaceous of southeastern Queensland ” . In Mesozoic Geology of the Eastern Australia Plate Conference , Geological Society of Australia Inc., Extended Abstracts no. 43 366 – 373 .
- McKellar , J.L. 2004 . Geophysical controls on late Palaeozoic - early Mesozoic geological history and floral succession: eastern Australia in perspective . Association of Australasian Palaeontologists Memoir , 29 : 47 – 83 .
- McKellar, J.L., Late Early to Late Jurassic palynology, biostratigraphy and palaeogeography of the Roma Shelf area, northwestern Surat basin, Queensland, Australia. Association of Australasian Palaeontologists Memoir (In press)
- McLoughlin , S. and Drinnan , A.N. 1995 . A Middle Jurassic flora from the Walloon coal measures, Mutdapilly, Queensland, Australia . Memoirs of the Queensland Museum , 38 : 257 – 272 .
- McLoughlin , S. and Hill , R.S. 1996 . “ The succession of Western Australian Phanerozoic terrestrial floras ” . In Gondwanan Heritage: Past, Present and Future of the Western Australian Biota , Edited by: Hopper , S.D. , Chappill , J. , Harvey , M. and George , A. 61 – 80 . Chipping Norton : Surrey Beatty & Sons .
- McLoughlin , S. and Vajda , V. 2005 . Ancient Wollemi Pines resurgent . American Scientist , 93 : 540 – 547 .
- McLoughlin , S. and Pott , C. 2009 . The stratigraphic significance of Western Australia Jurassic plant fossils in the collections of the Australian Museum, Sydney . GFF , 131 (this issue)
- McLoughlin , S. , Tosolini , A.-M. , Nagalingum , N. and Drinnan , A.N. 2002 . The Early Cretaceous (Neocomian) flora and fauna of the lower Strzelecki Group, Gippsland basin, Victoria, Australia . Association of Australasian Palaeontologists Memoir , 26 : 1 – 144 .
- Metcalfe , I. 1999 . “ Gondwanaland dispersion and Asian accretion: an overview ” . In Gondwanaland Dispersion and Asian Accretion Edited by: Metcalfe , I. , Jishuin , R. , Charvet , J. and Hada , S. 9 – 28 . Balkema, Rotterdam
- Molina-Garza , R.S. , Geissman , J.W. and Lucas , S.G. 2003 . Paleomagnetism and magnetostratigraphy of the lower Glen Canyon and upper Chinle Groups, Jurassic–Triassic of northern Arizona and northeast Utah . Journal of Geophysical Research , 108 ( B4 ) : 2181 doi:10.1029/2002JB001909
- Monteil, E., (coord.), 2006: Australian Mesozoic and Cenozoic Palynology Zonations – updated to the 2004 Geologic Time Scale. Geoscience Australia Record 2006/23 (on CD-ROM)
- Morgan , R. 1984 . “ Palynology ” . In Contributions to the geology of the Great Australian basin in New South Wales , Geological Survey of New South Wales Bulletin 31 Edited by: Hawke , J.M. and Cramsie , J.N. 143 – 159 .
- Ogg , J.G. and von Rad , U. 1994 . The Triassic of the Thakkhola (Nepal). II: Paleolatitudes and comparison with other eastern Tethyan margins of Gondwana . Geologische Rundschau , 83 : 107 – 129 .
- Ogg , J.G. , Ogg , G. and Gradstein , F.M. 2008 . The Concise Geologic Time Scale , 184 Cambridge : Cambridge University Press .
- Olsen , P.E. and McCune , A.R. 1991 . Morphology of the Semionotus elegans species group from the Early Jurassic part of the Newark Supergroup of Eastern North America with comments on the family Semionotidae (Neopterygii) . Journal of Vertebrate Paleontology , 11 : 269 – 292 .
- Partridge , A.D. 2006a . “ Jurassic-Early Cretaceous dinocyst zonations NWS Australia: 1st update of HMP 2004 ” . In Australian Mesozoic and Cenozoic Palynology Zonations – updated to the 2004 Geologic Time Scale , Geoscience Australia Record 2006/23 (on CD-ROM) Edited by: Monteil (coord.) , E.
- Partridge , A.D. 2006b . “ Jurassic-Early Cretaceous spore-pollen and dinocyst zonations for Australia ” . In Australian Mesozoic and Cenozoic Palynology Zonations – updated to the 2004 Geologic Time Scale , Geoscience Australia Record 2006/23 (on CD-ROM) Edited by: Monteil (coord.) , E.
- Partridge , A.D. 2006c . “ Late Cretaceous palynology zonations for Australia ” . In Australian Mesozoic and Cenozoic Palynology Zonations – updated to the 2004 Geologic Time Scale , Geoscience Australia Record 2006/23 (on CD-ROM) Edited by: Monteil (coord.) , E.
- Partridge , A.D. 2006d . “ Late Cretaceous-Cenozoic palynology zonations Gippsland basin ” . In Australian Mesozoic and Cenozoic Palynology Zonations – updated to the 2004 Geologic Time Scale , Geoscience Australia Record 2006/23 (on CD-ROM) Edited by: Monteil (coord.) , E.
- Pattemore , G.A. 2000 . A new Jurassic pteridosperm fructification from Queensland . Journal of African Earth Sciences , 31 : 187 – 193 .
- Pattemore , G.A. and Rigby , J.F. 2005 . Fructifications and foliage from the Mesozoic of southeast Queensland . Memoirs of the Queensland Museum , 50 : 329 – 345 .
- Percival , I.G. 1979 . “ Talbragar Fossil Site ” . In The Geological Heritage of New South Wales , Australian Heritage Commission & the Planning and Environment Commission of New South Wales 237 – 244 .
- Philippe , M. , Bamford , M. , McLoughlin , S. , Da Rosa Alves , L.S. , Falcon-Lang , H. , Gnädinger , S. , Ottone , D. , Pole , M. , Rajanikanth , A. , Shoemaker , R.E. , Torres , T. and Zamuner , A. 2004 . Biogeographic analysis of Jurassic-Early Cretaceous wood assemblages from Gondwana . Review of Palaeobotany and Palynology , 129 : 141 – 173 .
- Piper , J.D.A. 1987 . Palaeomagnetism and the continental crust , 434 Milton Keynes : Open University Press .
- Playford , P.E. , Cockbain , A.E. and Low , G.H. 1976 . Geology of the Perth basin, Western Australia . Western Australian Geological Survey Bulletin , 124 : 1 – 312 .
- Playford , P.E. , Cope , R.N. , Cockbain , A.E. , Low , G.H. and Lowry , D.C. 1975 . Phanerozoic . The Geology of Western Australia. Western Australian Geological Survey Memoir , : 223 – 433 .
- Pogson , D.J. and Cameron , R.G. 1999 . “ Surat basin ” . In Explanatory Notes, Dubbo Geological Sheet 1:250 000 S1/55-4 , Geological Survey of New South Wales, Mineral Resources of NSW 330 – 332 .
- Price , P.L. 1997 . “ Permian to Jurassic palynostratigraphic nomenclature of the Bowen and Surat basins ” . In The Surat and Bowen basins, South-east Queensland , Queensland Minerals and Energy Review Series, Queensland Department of Mines and Energy, Brisbane Edited by: Green , P.M. 137 – 178 .
- Price , P.L. , Filatoff , J. , Williams , A.J. , Pickering , S.A. and Wood , G.R. 1985 . “ Late Palaeozoic and Mesozoic palynostratigraphical units ” . CSR Oil and Gas Division Palynology Facility Report 274/225. Department of Resource Industries Open File report CR 14012 20, (unpublished report held by the Queensland Department of Employment, Economic Development and Innovation, Brisbane)
- Raine , J.I. , Mildenhall , D.C. and Kennedy , E.M. 2005 . New Zealand fossil spores and pollen: an illustrated catalogue Institute of Geological and Nuclear Sciences, Information Series 68, version 1+ http://www.gns.cri.nz/what/earthhist/fossils/spore_pollen/catalog/index.htm
- Ramsay , D.C. and Exon , N.F. 1994 . “ Structure and tectonic history of the northern Exmouth Plateau and Rowley Terrace: outer North West Shelf ” . In Geology of the outer North West Shelf, Australia , AGSO Journal of Australian Geology & Geophysics 15 Edited by: Exon , N.F. 55 – 70 .
- Rees , P.McA. 1993 . Dipterid ferns from the Mesozoic of Antarctica and New Zealand and their stratigraphical significance . Palaeontology , 36 : 637 – 656 .
- Rees , P.M. and Cleal , C.J. 2004 . Lower Jurassic floras from Hope Bay and Botany Bay, Antarctica . Special Papers in Palaeontology , 72 : 1 – 90 .
- Reiser , R.F. and Williams , A.J. 1969 . “ Palynology of the Lower Jurassic sediments of the northern Surat basin, Queensland ” . Geological Survey of Queensland Publication 339, Palaeontological Paper 15 24
- Rich , T.H. and Vickers-Rich , P. 2003 . A Century of Australian Dinosaur Collecting , 124 Queen Victoria Museum and Monash Science Centre, Launceston and Clayton .
- Rozefelds , A.C. 1988 . Lepidoptera mines in Pachypteris leaves (Corystospermaceae: Pteridospermophyta) from the Upper Jurassic/Lower Cretaceous Battle Camp Formation, north Queensland . Proceedings of the Royal Society of Queensland , 99 : 77 – 81 .
- Rozefelds , A.C. and Sobbe , I. 1983 . Fossil plants of the Durikai plant beds near Warwick, south east Queensland . The Fossil Collector Bulletin , 10 : 15 – 20 .
- Sajjadi , F. and Playford , G. 2002a . Systematic and stratigraphic palynology of Late Jurassic – earliest Cretaceous strata of the Eromanga basin, Queensland, Australia: Part One . Palaeontographica 261B , : 1 – 97 .
- Sajjadi , F. and Playford , G. 2002b . Systematic and stratigraphic palynology of Late Jurassic – earliest Cretaceous strata of the Eromanga basin, Queensland, Australia: Part Two . Palaeontographica , 261B : 99 – 165 .
- Sappal , K.K. and Suwarna , N. 1997 . “ Petrology and depositional environment of Early Jurassic coal, Western Australia ” . In Proceedings of the 30th International Geological Congress; Geology of Fossil Fuels; Coal 30 , Edited by: Yang , Q. 21 – 31 . Leiden : Brill Academic Publishers . 18B
- Sichler , B. and Perrin , M. 1993 . New early Jurassic paleopole from France and Jurassic apparent polar wander . Earth and Planetary Science Letters , 115 : 13 – 27 .
- Steinberger , B. and Torsvik , T.H. 2008 . Absolute plate motions and true polar wander in the absence of hotspot tracks . Nature , 452 : 620 – 623 .
- Stephens , C.J. , Holcombe , R.J. , Fielding , C.R. and Gust , D.A. 1996 . “ Tectonic evolution of the eastern Gondwanaland margin during the late Palaeozoic and early Mesozoic ” . In Mesozoic 96. Mesozoic Geology of the Eastern Australia Plate Conference, Brisbane, 1996 , Geological Society of Australia, Extended Abstracts 43 517 – 518 .
- Tenison-Woods , J.E. 1882 . On various deposits of fossil plants in Queensland . Proceedings of the Linnean Society of New South Wales , 7 : 95 – 98 .
- Tenison-Woods , J.E. 1883 . On the fossil flora of the coal deposits of Australia . Proceedings of the Linnean Society of New South Wales , 8 : 37 – 167 .
- Thies , D. 1989 . Lepidotes gloriae, sp. nov. (Actinopterygii: Semionotiformes) from the Late Jurassic of Cuba . Journal of Vertebrate Paleontology , 9 : 18 – 40 .
- Thies , D. , Mudroch , A. and Turner , S. 2007 . The potential of vertebrate microfossils for marine to non-marine correlation in the Jurassic . Progress in Natural Science , 17 : 655 – 663 .
- Thies , D. and Turner , S. 2001 . Jurassic actinopterygian from Queensland . Alcheringa , 25 : 381 – 386 .
- Thulborn , R.A. and Warren , A. 1980 . Early Jurassic plesiosaurs from Australia . Nature , 285 : 224 – 225 .
- Thulborn , R.A. and Turner , S. 2003 . The last dicynodont; an Australian relict . Proceedings of the Royal Society of London B , 270 : 985 – 993 . + cover
- Tidwell , W.D. 1987 . A new species of Osmundacaulis (O. jonesii sp. nov.) from Tasmania, Australia . Review of Palaeobotany and Palynology , 52 : 205 – 216 .
- Tidwell , W.D. 1992 . Millerocaulis richmondii sp. nov. an osmundaceous fern from Mesozoic strata near Little Swanport, Tasmania, Australia . Papers and Proceedings of the Royal Society of Tasmania , 126 : 1 – 7 .
- Tidwell , W.D. and Clifford , H.T. 1995 . Three new species of Millerocaulis (Osmundaceae) from Queensland, Australia . Australian Systematic Botany , 8 : 667 – 685 .
- Tidwell , W.D. and Jones , R. 1987 . Osmundacaulis nerii, a new Osmundaceous species from Tasmania, Australia . Palaeontographica , 204B : 181 – 191 .
- Tidwell , W.D. , Kim , J.-H. and Kimura , T. 1987 . Mid-Mesozoic leaves from near Ida Bay, southern Tasmania, Australia . Papers and Proceedings of the Royal Society of Tasmania , 121 : 159 – 169 .
- Tidwell , W.D. , Nishida , H. and Webster , N. 1989 . Oguracaulis banksii gen. et sp. nov. a mid-Mesozoic tree-fern stem from Tasmania, Australia . Papers and Proceedings of the Royal Society of Tasmania , 123 : 15 – 25 .
- Tidwell , W.D. , Munzing , G.E. and Banks , M.R. 1991 . Millerocaulis species (Osmundaceae) from Tasmania, Australia . Palaeontographica , 223B : 91 – 105 .
- Tidwell , W.D. and Pigg , K.B. 1993 . New species of Osmundacaulis emend. from Tasmania . Palaeontographica , 230B : 141 – 158 .
- Tidwell , W.D. and Rozefelds , A.C. 1990 . Grammatocaulis donponii gen. et sp. nov. a permineralized fern from the Jurassic of Queensland, Australia . Review of Palaeobotany and Palynology , 66 : 147 – 158 .
- Tidwell , W.D. and Rozefelds , A.C. 1991 . Yulebacaulis normanii gen. et sp. nov. a new fossil tree fern from south-eastern Queensland, Australia . Australian Systematic Botany , 4 : 421 – 132 .
- Townrow , J.A. 1967a . On Rissikia and Mataia, podocarpaceous conifers from the Lower Mesozoic of southern lands . Papers and Proceedings of the Royal Society of Tasmania , 101 : 103 – 136 .
- Townrow , J.A. 1967b . The Brachyphyllum crassum complex of fossil conifers . Papers and Proceedings of the Royal Society of Tasmania , 101 : 149 – 172 .
- Truswell , E.M. , Dettmann , M.E. and O'Brien , P.E. 1999 . Mesozoic palynofloras from the Mac. Robertson shelf, East Antarctica: geological and phytogeographic implications . Antarctic Science , 11 : 239 – 255 .
- Turner , S. 2008 . “ IGCP 506: Potential for a fish biostratigraphy for Jurassic fish for marine/non-marine correlation ” . In HPF 09411 Marine and non-marine Jurassic; Global correlation and major geological events , 33rd IGC abstracts, August 6–14, Oslo 170
- Turner , S. and Rozefelds , A. 1987 . First Jurassic actinopterygian fish from Queensland . Memoirs of the Queensland Museum , 25 : 201 – 206 .
- Upadhay , R. , Ram-Awatar Kar , R.K. and Sinha , A.K. 2005 . First record of Middle-Late Jurassic palynomorphs from the Lamayuru Complex, Indus Suture Zone, Ladakh, India . Current Science , 88 : 980 – 986 .
- Vail , P.R. and Todd , R.G. 1981 . “ Northern North Sea Jurassic unconformities, chronostratigraphy and sea-level changes from seismic stratigraphy ” . In Proceedings of the Second Conference on Petroleum Geology of the Continental Shelf of North-west Europe, London , Edited by: Illing , L.V. and Hobson , G.D. 216 – 235 . London : Heydan (on behalf of the Institute of Petroleum) .
- Vail , P.R. , Mitchum , R.M. and Thompson , S. 1977 . “ Seismic stratigraphy and global changes of sea level; Part 4: Global cycles of relative changes of sea level ” . In Seismic stratigraphy – applications to hydrocarbon exploration , The American Association of Petroleum Geologists, Memoir 26 Edited by: Payton , C.E. 83 – 97 .
- Vajda , V. 2001 . Aalenian to Cenomanian palynofloras of SW Scania, Sweden . Acta Paleontologica Polonica , 46 : 403 – 426 .
- Vajda , V. and Wigforss-Lange , J. 2006 . The Jurassic–Cretaceous transition of Southern Sweden – palynological and sedimentological interpretation . Progress in Natural Science , 16 : 1 – 38 .
- Vakhrameev , V.A. 1964 . Jurassic and Early Cretaceous Floras of Eurasia and Contemporary Paleofloristic Provinces . Nauka , : 261 (in Russian)
- Veevers , J.J. 1984 . Phanerozoic Earth History of Australia , 418 Oxford : Oxford University Press .
- Veevers , J.J. , ed. 2000 . Billion-year earth history of Australia and neighbours in Gondwanaland , 388 Sydney : GEMOC Press .
- Vickers-Rich , P. and Rich , T.H. , eds. 1993 . Wildlife of Gondwana. The 500-million-year history of vertebrate animals from the ancient southern subcontinent , 276 Chatswood, NSW : Reed .
- Wade , R.T. 1941 . The Jurassic Fishes of New South Wales . Journal & Proceedings of the Royal Society of NSW , 75 : 71 – 84 .
- Walkom , A.B. 1917a . Mesozoic floras of Queensland. Part 1.-continued. The flora of the Ipswich and Walloon Series. (c) Filicales etc . Publications of the Geological Survey of Queensland , 257 : 1 – 65 .
- Walkom , A.B. 1917b . Mesozoic floras of Queensland. Part 1.-concluded. The flora of the Ipswich and Walloon Series. (d) Ginkgoales, (e) Cycadophyta, (f) Coniferales . Publications of the Geological Survey of Queensland , 259 : 1 – 49 .
- Walkom , A.B. 1919 . On a collection of Jurassic plants from Bexhill, near Lismore, N.S.W . Proceedings of the Linnean Society of New South Wales , 44 : 180 – 190 .
- Walkom , A.B. 1921a . On the occurrence of Otozamites in Australia, with descriptions of specimens from Western Australia . Proceedings of the Linnean Society of New South Wales , 46 : 147 – 153 .
- Walkom , A.B. 1921b . Mesozoic floras of New South Wales. Part 1- Fossil plants from Cockabutta Mountain and Talbragar . Geological Survey of New South Wales, Palaeontology, Memoir , 12 : 1 – 21 .
- Walliser , O.H. , ed. 1996 . Global Events and Event Stratigraphy. Results of IGCP 216 Global Biological Events in Earth History , 333 Berlin : Springer .
- White , M.E. 1981a . Revision of the Talbragar fish bed flora (Jurassic) of New South Wales . Records of the Australian Museum , 33 : 695 – 721 .
- White , M.E. 1981b . Fish beds reveal lush fossil forest . Australian Natural History , 20 ( 7 ) : 227 – 230 .
- White , M.E. 1986 . The Greening of Gondwana , The 400 Million Year Story of Australia's plants 256 Frenchs Forest, NSW : Reed .
- White , M.E. 1990 . The Nature of Hidden Worlds , 256 Balgowlah : Reed Books .
- White , M.E. 1991 . Time in our Hands , 191 Balgowlah : Reed Books .
- Woodward , A.S. 1895 . The Fossil Fishes of the Talbragar Beds (Jurassic?) . Memoirs of the Geological Survey of New South Wales, Palaeontology , 9 : 1 – 27 .
- Yago , J.V.R. and Fielding , C.R. 1996 . “ Mesozoic 96, Mesozoic Geology of the Eastern Australia Plate Conference, Brisbane, 1996 ” . Geological Society of Australia, Extended Abstracts 43 574 – 575 .
- Yemane , K. 1993 . Contribution of Late Permian palaeogeography in maintaining a temperate climate in Gondwana . Nature , 361 : 51 – 54 .
- Young , G.C. and Laurie , J. , eds. 1996 . Australian Phanerozoic Timescales , 279 Oxford : Oxford University Press .
- Ziegler , A.M. , Parrish , J.M. , Yao , J. , Gyllenhaal , E.D. , Rowley , D.B. , Parrish , J.T. , Nie , S. , Bekker , A. and Hulver , M.L. 1993 . Early Mesozoic phytogeography and climate . Philosophical Transactions of the Royal Society of London 341B , : 297 – 305 .