
Abstract
Disarticulated sponge spicules are described in two diverse assemblages from the Holm Dal Formation of North Greenland (Cambrian Series 3, Guzhangian Stage, Lejopyge laevigata Biozone). They compare most closely with contemporaneous spicules described from the Mindyallan Stage of Queensland, Australia. Although originating on separate palaeocontinents, the Laurentian and Gondwanan spicule assemblages accumulated in equatorial latitudes, demonstrating the potential stratigraphic and palaeogeographic utility of disarticulated spicules in the Cambrian. Samples from Gustav Holm Dal are dominated by robust acanthose pentactins with widely spaced spines also present in the solariform Seqineqia bottingi n. gen. n. sp. Australispongia sinensis Dong & Knoll, 1996, Silicunculus australiensis Bengtson, 1986 and Silicunculus saaqqutit Peel, 2017 also occur but are not common. Samples from Navarana Fjord are dominated by tetractins and pentactins. Speciosuspongia cf. wangcunensis Chen & Dong, 2008 may be common. Tetractins of Tallitaniqa petalliformis n. gen. n. sp., Kuonamia fusiformis (Fedorov in Fedorov & Pereladov, 1987) and Sisamatispongia erecta n. gen. n. sp. are only recorded from Navarana Fjord.
Disarticulated spicules of sponges are common in Cambrian carbonate rocks worldwide (Bengtson Citation1986; Bengtson et al. Citation1990; Zhang & Pratt Citation1994; Dong & Knoll Citation1996; Mehl Citation1998; Sugai et al. Citation2004; Elicki Citation2011; Kouchinsky et al. Citation2011, 2015; Zheng et al. Citation2012). They provide a fossil record which complements the assemblages of articulated specimens recovered from Cambrian lagerstätten such as the Burgess Shale of Canada (Rigby Citation1986; Rigby & Collins Citation2004) and numerous earlier Cambrian assemblages from China (CitationSteiner et al. 1993; Rigby & Hou Citation1995; Yang & Zhao Citation2000; Wu Citation2004; Wu et al. Citation2005; Xiao et al. Citation2005). In North Greenland, a diverse assemblage from the Sirius Passet Lagerstätte (Cambrian Series 2, Stage 3) represents the oldest major occurrence of articulated sponges in Laurentia (Botting et al. Citation2015; Botting & Peel Citation2016; Peel & Willman Citation2018) but similarly preserved specimens have not been recovered from the carbonate-dominated succession overlying the Buen Formation (Fig. B). However, routine processing in weak acids of Cambrian carbonate samples from Greenland has produced assemblages of sponge spicules from most formations which can provide insight into the evolution of sponge faunas during the early–middle Cambrian (Cambrian Series 2–3). Some records from Greenland exist (Skovsted Citation2006; Peel et al. Citation2016; Peel Citation2017a) but this paper focusses on the spicule assemblage from the Holm Dal Formation (Cambrian Series 3, Guzhangian Stage; Figs. and ), a unit which has demonstrated utility within Laurentian Cambrian biostratigraphy (Peel Citation1988a, Citation1994).
Figure 1. A. derivation of GGU samples from the Holm Dal Formation in North Greenland. B, Cambrian stratigraphy of the southern J.P. Koch Fjord area, North Greenland.
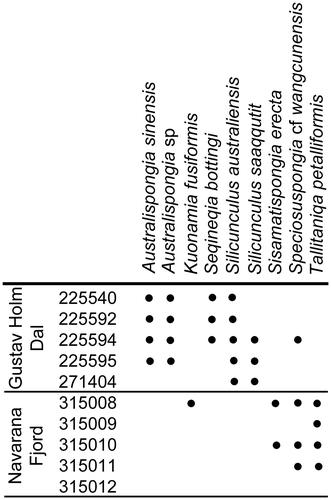
Figure 2. Distribution of described spicule taxa in the Holm Dal Formation. GGU samples are listed in numerical order.
Most described sponge spicule occurrences yield simple morphologies such as hexactins, pentactins and tetractins (=stauractins) that show little variation between samples from area to area or from horizon to horizon. Their similarity generally hinders the recognition of biological species within assemblages where both the specific assignment and the juxtaposition of the various spicule morphotypes is unknown. Fedorov & Peredalov (Citation1987) proposed several genera from the Cambrian of Siberia which were based on groups of similar spicules, e.g., diactins to hexactins, within a single morphological series in the respective type species. The figured specimens comprising these can be regarded as syntypes. However, neither the possible presence of spicules from other spicule series within the taxa nor the distribution of spicules of these types in other taxa were discussed. Suggested reconstructions of sponges by Rigby (Citation1975), Mehl (Citation1998), Sugai et al. (Citation2004) and Castellani et al. (Citation2012) included morphological transitions of spicules from more than a single series, indicating that the parent sponge wall in these taxa, as in most modern sponges (Schulze Citation1887), had several layers with spicules of distinct form.
Morphologically distinctive isolated spicules have been named under zoological nomenclature, resulting in the recognition of a number of “spicule-based” taxa (Bengtson in Bengtson et al. Citation1990; Mostler Citation1985, Citation1996; Mehl Citation1998; Chen & Dong Citation2008). The biological value of such taxa is often justifiably questioned (Castellani et al. Citation2012) but it is inappropriate to just dismiss their usefulness. It is unfortunate that size differences and taphonomic factors form a preservational barrier that is hard to cross, with the result that most descriptions of articulated sponges provide little detailed illustration of individual spicules. However, despite their small size, it is unlikely that the distinctive spicule morphologies described herein (Fig. ) have been overlooked in earlier studies of articulated specimens, indicating that study of spicules may enhance known biodiversity.
Spicules are an important source of supplementary information concerning the geological record of sponge evolution, largely due to details brought forward through the employment of the scanning electron microscopy and similar techniques that have revolutionized the study of microscopic Cambrian shelly faunas during the last half century. Exceptional details of fine surface structures of spicules can be discerned (Castellani et al. Citation2012), although surface mineralisation may produce a false surface pattern as a result of grain contact (Fig. CC). Information concerning the occurrence of biminerallic spicules in early sponges is of considerable phylogenetic significance (Botting & Butterfield Citation2005; Botting et al. Citation2012).
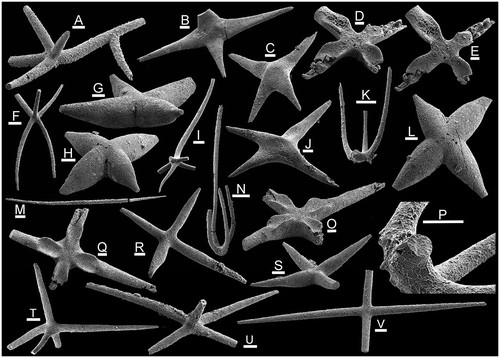
Figure 3. Spicules from the Holm Dal Formation (Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone). A, B, I, Y, EE. Silicunculus australiensis Bengtson, Citation1986. A, PMU 29996 from GGU sample 225540. B, I, PMU 30003 from GGU sample 225595. Y, PMU 29997 from GGU sample 225592. EE, PMU 29998 from GGU 225594. C–F. Silicunculus saaqqutit Peel, 2017. C, D, PMU 30002, paratype, from GGU sample 225595. E, F, PMU 29999, holotype, from GGU sample 225540. G, FF. Radiolarian primary spicules (Protoentactinia? sp.) from GGU sample 225540. G, PMU 31801. FF, PMU 31802, the tapering of the secondary rays in the lower part reflects submersion into SEM stub adhesive. H, J, K, N–Q, T–W, AA–DD, GG. Diverse sponge spicules. H, dichodiactin, PMU 31803 from GGU sample 225592. J, V, hexactin with stunted paratangential rays. J, PMU 31804 from GGU sample 225540. V, PMU 31805 from GGU sample 225540. K, acanthose pentactin, PMU 31806 from GGU sample 225594. N, triactin, PMU 31807 from GGU sample 225540. O, diactin with secondary ray, PMU 31808 from GGU sample 225592. P, DD, acanthose pentactin with claw-like paratangential rays (one broken), PMU 31809 from GGU sample 225592. Q, pentactin, PMU 31810 from GGU sample 225595. T, acanthose pentactin, PMU 31811 from GGU sample 225595. U, didactins, PMU 31812 and PMU 31813 (2 conjoined spicules) from GGU sample 225592. W, tetractin, PMU 31814 from GGU sample 315008. Z, tetractin, PMU 31818 from GGU sample 225592, with arrow indicating axial canal. AA, GG, acanthose pentactin, PMU 31815 from GGU sample 225592, with arrow indicating cast of the axial canal. BB, CC, pentactin, PMU 31816 from GGU sample 225592, with arrow indicating cast of axial canal. L, Australispongia sp., PMU 31817 from GGU sample 225594, with arrows indicating the acute flanges. M,S, Australispongia sinensis Dong & Knoll, Citation1996, PMU 31818 from GGU sample 225592. R,X, Speciosuspongia cf. wangcunensis Chen & Dong, Citation2008. R, pentactin, PMU 31819 from GGU sample 225594. X, pentactin, PMU 31820 from GGU sample 315008. Scale bars: 100 μm except CC, GG (50 μm).

The shape in cross-section of the canal produced by the axial filament has been regarded as a key discriminating character in rays of silicean sponges, being square in hexactinellids and hexagonal in demosponges (Botting & Muir Citation2013). Traces of the canal are often preserved in spicules in the form of a central canal, although this is susceptible to diagenetic modification. In describing a hexagonal axial canal in an Ordovician hexactinellid-like sponge, Botting & Muir (Citation2013) demonstrated that this form of axial canal may be widely distributed in early silicean sponges and not just a feature of demosponges. Whilst difficult to interpret due to preservational constraints (Botting & Muir Citation2013), it is evident that a largely unexplored potential exists for elucidating filament evolution and phylogeny in the earliest sponges. The occurrence of axial canals in many spicules in the present material is noted (Fig. Z, AA, CC, GG, arrows; Fig. L) but detailed study is beyond the scope of the present paper.
Figure 4. Sponge spicules from the Holm Dal Formation (Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone). A–D, F, K. Seqineqia bottingi n. gen. n. sp. A, B, F, holotype, PMU 31821 from GGU sample 225540. C, paratype, PMU 31822 from GGU sample 225592. D, paratype, PMU 31823 from GGU sample 225594; the spicule is partially submerged in adhesive. K, paratype, PMU 31824 from GGU sample 225540. E, G–J, Acanthose pentactins with widely spaced, stubby, spines. E, PMU 31825 from GGU sample 225594. G, L, PMU 31826 from GGU sample 225595, with detail of axial canal (L). H, PMU 31827 from GGU sample 225594. I, PMU 31828 from GGU sample 225594. J, PMU 31829 from GGU sample 225594. Scale bars 100 μm, except A (30 μm) and K (50 μm).

This paper employs spicule-based taxonomy to describe a number of morphologically distinct spicules (Fig. ), in full cognizance of the taxonomic limitations. Apart from partially filling gaps in the geological record and perception of the diversity of articulated sponges, this approach increases the visualization of the similarities between sponge assemblages which are now being recognized from different parts of the Cambrian world, and even their stratigraphic utility (Peel Citation2017a).
Material and localities
The Holm Dal Formation is a highly fossiliferous component of the prograding Cambrian shelf margin complex that formed on the southern margin (present day co-ordinates) of the transarctic Franklinian Basin in eastern areas of North Greenland (Higgins et al. Citation1991; Ineson & Peel Citation1997). Although restricted in distribution to the inner area of J.P. Koch Fjord (Fig. ), the formation provides a key location for the biostratigraphic correlation of late middle Cambrian (Cambrian Series 3, Guzhangian Stage) strata in Laurentia. Abundant and diverse trilobites described by Robison (Citation1988) demonstrated the co-occurrence of taxa characteristic of the lower and middle Cedaria Interval-zone of inner craton successions in North America with open shelf agnostoids characteristic of the Lejopyge laevigata Biozone. In a wider context, the North American faunas, assigned to the Dresbachian Stage in the Upper Mississippi Valley region, are now referred to the Marjuman Stage of western North American usage (Babcock et al. Citation2011, 2014), whereas the agnostoids indicate that the Holm Dal fauna belongs to the lowest biozone (Lejopyge laevigata) of the Guzhangian Stage of Cambrian Series 3 of the emerging international standard (Peng et al. Citation2009; Babcock et al. Citation2011, 2014).
Babcock (Citation1994a) extended the observations of Robison (Citation1988) concerning the interplay between inner craton and open shelf trilobite faunas in the middle Cambrian (Cambrian Series 3) of North Greenland in describing deep outer shelf, cool water, trilobite assemblages from along the northern coast. Polymeroid trilobites of Baltic aspect (Babcock Citation1994b) and agnostoids (Robison Citation1994) were described from Guzhangian Stage strata of the Kap Stanton Formation from the middle reaches J.P. Koch Fjord (Fig. A, KSF locality). Thus, cool, deep water faunas of Baltic aspect occur without major tectonic shortening only 35 km north (offshore) of sections of the Holm Dal Formation yielding admixed Laurentian inner craton and open shelf trilobites.
The Holm Dal Formation was proposed by Ineson (Citation1988) and in its type section, on the east side of Gustav Holm Dal at its junction with Fimbuldal (Fig. A), it includes 155 m of recessive, mainly thin-bedded argillaceous limestones and dolostones. The formation wedges out already just 5 km to the south, across Fimbuldal, and thins to about one third of this thickness at Navarana Fjord where it is mainly composed of dark laminated dolostones with about 5 m of fossiliferous dark limestones with shale partings about 25 m above the base (Fig. A). Northward along Gustav Holm Dal, the formation appears to thicken and become more argillaceous in poorly exposed ground. The formation represents a major regressive interval, a low stand of sea level, between the strongly prograding, cliff-forming, coarse dolostones of the Fimbuldal and Perssuaq Gletscher formations (Ineson Citation1988; Higgins et al. Citation1991; Ineson & Peel Citation1997). Phosphate covered hardground surfaces in the lower part of the formation are covered with pelmatozoan holdfasts (Peel Citation2017b), the oldest in situ record of such encrustation in Laurentia. Fossils occur mainly in the lower half of the formation which becomes increasingly dolomitic upwards.
Robison (Citation1988) described 58 species of trilobites from the Holm Dal Formation. Brachiopods were described by Zell & Rowell (Citation1988) and molluscs by Peel (Citation1988b). Peel (Citation2017a) described the sponge spicules Australispongia Dong & Knoll, Citation1996 and Silicunculus Bengtson, Citation1986, while Bergström & Ineson (Citation1988) described arthropod trace fossils.
Methods. Sponge spicules described herein from the Holm Dal Formation were hand picked from sieved residues (usually greater than the 250 μm fraction) of limestone samples dissolved in 10% acetic acid. Specimens were imaged by scanning electron microscopy and figures were assembled using Adobe Photoshop CS4. Individual residues may be dominated by organophosphatic brachiopods (Zell & Rowell Citation1988), although phosphatised Girvanella-like cyanobacteria are also common in some samples. With the exception of scattered paraconodonts, other fossils are rare.
Composition and Preservation. – All spicules appear to be preserved in silica although this varies from dominantly translucent white to matt dark brown in colouration both between, and often within, samples. Spicule surfaces vary from smooth and pristine, to coated with microcrystalline quartz. Quartz crystals may also occur as geode infillings within some articulated linguliformean brachiopods. Small patches of lustrous pyrite occur on most spicules. Axial cavities are visible in many spicules where a relatively large diameter to the cavity indicates diagenetic solution (Fig. Y). A few specimens show a general degradation or incomplete recrystallisation of the rays (Fig. D, E, Q) but most are well preserved. The distal extremities of rays are frequently broken, but to what extent this results from the preparation technique is not known. Diagenetic coating (originally pyrite?) of some spicules produces casts of the axial cavity, while the spicule skeleton itself has been dissolved diagenetically or during preparation (Fig. AA–CC, GG).
Figure 5. Sponge spicules from the Holm Dal Formation (Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone). All specimens from GGU sample 315008. A–C, J, F, M, R–V, Diverse spicules. A, pentactin with secondary axial ray, PMU 31830. B, tetractin, PMU 31831. C,J, tetractin, PMU 31832. F, pentactin, PMU 31833. M, diactin, PMU 31834. R, tetractin, PMU 31835. S, tetractin, PMU 31836. T, pentactin, PMU 31837. U, pentactin, PMU 31838. V, tetractin, PMU 31839. D, E, O, Q, Tallitaniqa petalliformis n. gen. n. sp. D, E, tetractin, paratype, PMU 31840. O, Q, tetractin, holotype, PMU 31841. G, H, L, Kuonamia fusiformis (Fedorov in Fedorov & Pereladov, Citation1987). G, tetractin, PMU 31842. H, L, tetractin, PMU 31843. I, hexactin, PMU 31844. K, N, P, Sisamatispongia erecta n. gen. n. sp. K, P, holotype, PMU 31845. N, paratype, PMU 31846. Scale bars 100 μm, except P (50 μm).
Localities.– GGU sample 225540 was collected by J.S. Peel on 7th July 1979 from 55 m above the base of the Holm Dal Formation (thickness 155 m) in its type section (Ineson Citation1988, fig. 6; Robison Citation1988, fig. ) on the east side of Gustav Holm Dal at its junction with Fimbuldal, western Peary Land (82°19.5′N, 39˚37′W; Fig. A).
GGU samples 225592, 225594, 225595 were collected by J.S. Peel on 13th July 1979 from the Holm Dal Formation in Gustav Holm Dal, western Peary Land (82°22′N, 39°41′W; Fig. A). This is locality 3 of Robison (1988, fig. 1B, 5). GGU sample 225592 is from near the middle of an approximately 100 m thick section (and probably about the same stratigraphic level as GGU sample 225540) but 225594 and 225595 are from the upper part of the section.
GGU sample 271404 was collected by P. Frykman and J.S. Peel on 16th June 1978 from 60 m above the base of the Holm Dal Formation in Gustav Holm Dal, western Peary Land (82°21′N, 39°40′W; Fig. A).
GGU samples 315008–315012 were collected by J.S. Peel on 28th June 1984 from 25 to 28 m above the base of the Holm Dal Formation on the western side of the glacier feeding into Navarana Fjord, south-east Freuchen Land (82°16.7′N, 41°22′W; Fig. A).
Institutional abbreviations. – GGU: Grønlands Geologiske Undersøgelse [Geological Survey of Greenland], now a part of the Geological Survey of Denmark and Greenland (GEUS), Copenhagen, Denmark. PMU: palaeontological type collection of the Museum of Evolution, Uppsala University, Uppsala, Sweden.
Assemblages and correlation
Differences between the spicule assemblages from the Holm Dal Formation in Gustav Holm Dal (Figs. and ) and from the locality alongside the glacier feeding into the head of Navarana Fjord (mainly illustrated in Fig. ) probably reflect the more argillaceous character of the formation at Gustav Holm Dal. Sponge spicules have not been recovered from samples from the contemporaneous deep water Kap Stanton Formation in the middle reaches of J.P. Koch Fjord (Fig. A, KSF locality).
Samples from the Holm Dal Formation in Gustav Holm Dal are dominated by robust acanthose pentactins in which the 5 rays are of similar size (Fig. E), although morphotypes with a long axial ray are also conspicuous (Figs. T; I, J). The distinctive widely spaced thorn-like spines are also present on the solariform spicules of Seqineqia bottingi n. gen. n. sp. (Fig. A, K), suggesting that they may form part of the same sponge scleritome. The solariform spicules are rare, however, no doubt reflecting their delicate structure and small size. Slender, smooth, tetractins, pentactins and rare three-rayed spicules (Fig. N) may be common. Australispongia sinensis Dong & Knoll, Citation1996 (Fig. M, S), Silicunculus australiensis Bengtson, Citation1986 (Fig. A, B, EE) and Silicunculus saaqqutit Peel, Citation2017a (Fig. C, E) are widely distributed but not common; other spicule types are rare.
Samples from alongside the glacier feeding into Navarana Fjord (Fig. A) are dominated by large, smooth tetractins in which the straight to curved rays intersect at right angles to each other or slightly obliquely (Fig. V). The axial ray in accompanying pentactins may be short (Fig. T, U) or long, straight or slightly curved, perpendicular or slightly inclined to the subequal paratangential rays (Figs. Q and T, U). However, it is strongly oblique to the paired paratangential rays in pentacts assigned to Speciosuspongia cf. wangcunensis Chen & Dong, Citation2008 (Fig. R, X); the latter are common in some samples. Pentactins of both types maybe smooth or finely acanthose. Tetractins with spindle-shaped, fusiform, rays are also common (Fig. R, S). Some rare tetractins show strongly tapering rays (Fig. B), or non-perpendicular rays with broad transverse depressions near the origin (Fig. C, J). Tetractins with curved rays which are flat on their convex side (Fig. W, Z) occur rarely both from Gustav Holm Dal and Navarana Fjord.
Tetractins of Tallitaniqa petalliformis n. gen. n. sp. (Fig. D, E, O, Q) occur in several samples from Navarana Fjord. Rare inflated tetractins assigned to Kuonamia fusiformis (Fedorov in Fedorov & Pereladov, Citation1987; Fig. G, H, L) and delicate erect tetractins of Sisamatispongia erecta n. gen. n. sp. (Fig. K, N, P) are only recorded from Navarana Fjord (Fig. ).
The sponge spicule assemblages of the Holm Dal Formation compare most closely with spicules described by Bengtson (Citation1986) from the contemporaneous Mungerebar Limestone of Queensland (Cambrian Series 3, basal Mindyallan Stage, Acmarhachis quasivespa Biozone), a correlation proposed by Robison (Citation1988) on the basis of trilobite faunas. Shared forms include Silicunculus australiensis, Australispongia sinensis, coarsely acanthose pentactins (Figs. T and I), a dichodiactin (Fig. H) and similar slender pentactins (Fig. U). Although located on different palaeocontinents, North Greenland and Australia were placed at the equator during the late Middle Cambrian, whereas South China was located in the southern tropics (Torsvik & Cocks Citation2017). Spicules from the Chefu and lower Bitiao formations (Cambrian Series 4, Furongian Series, Paibian Stage) of Hunan described by Dong & Knoll (Citation1996, figs. 23, 30) include fragments that can be assigned to Sisamatispongia erecta n. gen. n. sp. and possibly Silicunculus. A fragment of a rod-like ray with rounded longitudinal cords separated by narrow grooves (Dong & Knoll Citation1996, fig. 7.11) is indistinguishable from rare similar fragments in GGU samples 315009 and 315010 (not illustrated). A pentactin (Dong & Knoll Citation1996, fig. 7.10) appears to be Speciosuspongia cf. wangcunensis Chen & Dong, Citation2008, while a diactin spicule with a secondary ray (Fig. O) compares to diaxon triactins figured by (Dong & Knoll Citation1996, pl. 7.18 and 7.20).
Assemblages described by Mehl (Citation1998) from the Georgina Basin are mainly slightly older (Stage 5 and Drumian). They generally lack spicules similar to the Holm Dal Formation associations but Peel (Citation2017a) described spicules of this age, originally recorded from Australia, from the upper Henson Gletscher Formation of North Greenland. Considered together, the Greenland records and their corresponding Australian occurrences demonstrate both the palaeogeographic significance and potential stratigraphic utility of disarticulated sponge spicules in the Cambrian.
Radiolaria. – Rare fragments occurring together with sponge spicules in GGU sample 225540 from Gustav Holm Dal (Fig. A) are the first radiolarian spicules reported from the Cambrian of North Greenland (Fig. G, FF). They are reminiscent of Protoentactinia Kozur, Mostler & Repetski, Citation1996a illustrated by Maletz & Bruton (Citation2007) and Maletz (Citation2011) from the lower Ordovician of Spitsbergen. Won & Below (Citation1999) described diverse assemblages of radiolarians from strata assigned to Cambrian Series 3 in the Georgina Basin.
Systematic palaeontology
PHYLUM PORIFERA Grant, Citation1836
Discussion. – It has become increasingly apparent from recent studies of spicule composition, axial canals and morphology that the relationships of Cambrian sponges and their evolutionary links with crown group poriferans are unclear (Botting & Butterfield Citation2005; Botting & Muir Citation2013; Botting et al. 2013; Carrera & Botting Citation2008). Therefore, the spicules described herein are not assigned at suprageneric level, although many might be regarded as hexactinellid on account of their morphology and presumed original silica composition.
Genus Australispongia Dong & Knoll, Citation1996
Type species. – Australispongia sinensis Dong & Knoll, Citation1996
Australispongia sinensis Dong & Knoll, Citation1996
M, S
1986 Flanged pentacts, Bengtson, p. 211, fig. 7a–h.
1996 Australispongia sinensis Dong & Knoll, p. 177, figs. 5.1–5.16, 6.1, 6.2, 6.9–14.
2017a Australispongia sinensis; Peel, p. 310, fig. 3I–K.
Figured material. – PMU 29995 from GGU sample 225592, Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone, Gustav Holm Dal, western Peary Land, North Greenland (Fig. ).
Other material. – Three broken spicules from GGU sample 225540, five from GGU sample 225592 and five from GGU sample 225594; Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone, Gustav Holm Dal, western Peary Land, North Greenland (Figs. and ).
Discussion. – Peel (Citation2017a) described this pentactin with a long axial ray from spicules with broken tips from the Holm Dal Formation, and a single spicule from the Fimbuldal Formation. Maximum preserved length is 1.3 mm, similar to specimens illustrated by Bengtson (Citation1986) from Australia and Dong & Knoll (Citation1996) from China. The four short paratangential rays are circular in cross-section and lie in a single plane or slope slightly away from the dominant axial ray, the latter being straight with pronounced longitudinal flanges separated by concave interareas. Three flanges on the axial ray twist anticlockwise distally in the figured specimen from the Holm Dal Formation (Fig. M) but the five flanges in a specimen from the Fimbuldal Formation illustrated by Peel (Citation2017a) are slightly twisted clockwise distally. Dong & Knoll (Citation1996) noted three or four flanges in the type species from the Furongian Series (Paibian Stage) of Hunan, China. The flanges are weakly serrate, with widely spaced low spines. Bengtson (Citation1986) and Dong & Knoll (Citation1996) recorded specimens with slightly curved axial rays but this ray is straight in the few Greenland specimens.
In Australispongia compressa Kozur, Mostler & Repetski, Citation1996b, from the Lower Ordovician of Nevada, USA, the broad and flat axial ray is blade-like, with serrated edges. Kozur et al. (Citation1996b) also referred undescribed tetracarinate spicules from the Permian of Texas to Australispongia.
Australispongia sp.
L
Figured material. – PMU 31817 from GGU sample 225594, Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone, Gustav Holm Dal, western Peary Land, North Greenland (Fig. ).
Other material. – About 15 fragments from GGU samples 225540, 225592 and 225594, Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone, Gustav Holm Dal, western Peary Land, North Greenland (Figs. and ).
Discussion. – About 15 fragments of relatively large specimens referred to Australispongia sp. from GGU samples 225540, 225592 and 225594 are broken so that no traces of paratangential rays (if present) are preserved (Fig. L). The rays expand in width towards the rounded, presumed distal, termination; the concave surfaces separating the three acute flanges (Fig. L, arrows) may carry a single longitudinal cord. Somewhat similar blade-like spicules were described from the Mindyallan of Queensland by Bengtson (Citation1986, fig. 10A–D).
Kuonamia Doweld, Citation2016
1987 Disparella Fedorov in Fedorov & Pereladov, p. 41 (non Hessler Citation1970)
2016 Kuonamia nom. nov. Doweld, Citation2016, p. 19
Type species. – Disparella fusiformis Fedorov in Fedorov & Pereladov, Citation1987, Cambrian Series 2, Stage 4, Oryctocara Biozone, Kuonamka Formation, Yakutia, North-eastern Siberia.
Discussion. – Fedorov & Pereladov (Citation1987, pl. 13) included morphotypes with two to six rays in a spicule assemblage referred to Kuonamia fusiformis and the illustrated isolated specimens are considered to be syntypes.
Kuonamia and Cjulanciella Fedorov in Fedorov & Pereladov, Citation1987 both have inflated rays but the degree of inflation in the latter genus is greater, such that the angle between adjacent rays in tetractins is generally obtuse. Adjacent rays in Kuonamia most frequently meet at about right angles (Fig. G, H, L).
Kuonamia fusiformis (Fedorov in Fedorov & Pereladov, Citation1987 )
G, H, L
1987 Disparella fusiformis Fedorov in Fedorov & Pereladov, Citation1987, p. 42, pl. 13, figs. 3–9.
Citation2016 Kuonamia fusiformis; Doweld, p. 19
Figured material. – PMU 31842–PMU 31844 from GGU sample 315008, Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone, glacier feeding into head of Navarana Fjord, North Greenland (Fig. ).
Discussion. – Rare tetractins placed here are flattened on one surface (Fig. G) but inflated on the opposite one (Fig. H). The rays are fusiform, constricting slightly at their junction and narrowing strongly towards their distal extremity which may be pointed. The rays are slightly more fusiform than in corresponding tetractins illustrated from the Kuonamka Formation by Fedorov & Pereladov (Citation1987, pl. 13, figs. 6, 9), but are similar to those in a pentact illustrated by Fedorov in Shabanov et al. (Citation1987, pl. 34, fig. 14) from boreholes through the Cambrian in the Anabar Anticline, Siberia.
Genus Seqineqia n. gen.
LSID. – urn:lsid:zoobank.org:act:67B80F99-32A4-48D4-A007-5C078103AAFB
Type species. – Seqineqia bottingi n. gen. n. sp., Holm Dal Formation (Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone), Gustav Holm Dal, Peary Land, North Greenland (Fig. ).
Derivation of name. – From “seqineq” (Greenlandic) meaning the sun.
Diagnosis. – Solariform, with 13–15 almost coplanar, acanthose, rays radiating from central disc; diameter of central disc about equal to length of individual rays.
Discussion. – In terms of the large number of radiating rays, Seqineqia resembles Kometia Webby & Trotter, Citation1993 but the numerous secondary rays in the latter arise by subdivision of one axial ray. The underlying hexactin symmetry of Kometia is clearly evident, with well developed paratangential rays and the modified axial ray (Webby & Trotter Citation1993; Mehl Citation1998; Won et al. Citation2007; Carrera & Maletz Citation2014). In Seqineqia all rays are preserved within a single plane without differentiation of an axial ray. The unequal depth of the interspaces between rays in Seqineqia bottingi n. gen. n. sp. (Fig. A, F) may suggest that the radiating rays developed by division from an initial hexactin pattern. Seqineqia and Kometia both have acanthose rays.
Seqineqia bottingi n. gen. n. sp.
A–D, F, K
LSID. – urn:lsid:zoobank.org:act:3E1CE6DB-C9A9-4FAD-8FB6-CA50544274D7
Derivation of name. – For Joseph P. Botting in recognition of his studies of poriferan evolution.
Holotype. – PMU 31821 from GGU sample 225540 (Fig. A, B, F), Holm Dal Formation (Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone), Gustav Holm Dal, Peary Land, North Greenland (Fig. ).
Paratypes. – PMU 31824 from GGU sample 225540; PMU 31822 from GGU sample 225592; PMU 31823 from GGU sample 225594, Holm Dal Formation (Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone), Gustav Holm Dal, Peary Land, North Greenland (Figs. and ).
Other material. – Three additional specimens from GGU sample 225540.
Diagnosis. – As for genus.
Description – Spicules are solariform, with 13–15 almost coplanar, acanthose, rays radiating from the central disc, the diameter of which is about equal to the length of individual rays. Individual rays carry widely spaced, short, stubby, spines which are lower in relief and more widely spaced near their tips. The central disc is variably spinose (Fig. D, F) but individual spines are less prominent than on the proximal part of the rays.
Discussion. – Fifteen rays are preserved in the holotype (Fig. B, F), but only 13 or 14 are present in the paratypes. While the tips of the rays seem to extend a similar distance from the centre of the spicule, the interspaces between them are of unequal depth (Fig. A). In one paratype (Fig. D) the rays can be traced clearly across the central disc towards its centre, but this feature is inconspicuous in other specimens.
Polyactins from the ?upper Templetonian of the Georgina Basin (Cambrian Series 3, Stage 5) illustrated by Mehl (Citation1998, pl. 3, figs. 13, 15) have five or six co-planar rays with stubby spines similar to the individual rays of Seqineqia. The latter has a broader central disc, however, and more than twice as many rays, but there is evidence that at least some of these rays are bifurcated (Fig. A). An additional specimen figured by Mehl (Citation1998, pl. 3, fig. 14) has eight acanthose rays and a broad central disc.
Spicule association. – Acanthose pentactins are associated with the solariform spicules (Fig. E, G–J) and are common also in GGU samples 225595 (Fig. K, T, AA) and 271404. They display similarly shaped spines on the rays to those in the type series of Seqineqia bottingi, suggesting that they may form part of the same spicule skeleton (Fig. E, G–J, L). The pentactins are variable in form, with four straight to slightly curved paratangential rays lying perpendicular to each other and to the axial ray; the axial ray may be slightly curved (Fig. I) and substantially longer than the paratangential rays (Fig. J). The stubby spines are widely spaced and mainly restricted to the basal parts of the rays, usually leaving the tips smooth (Fig. E, G, H, J), as is the case also in the solariform spicules. However, spines may be more common in pentactin spicules where the axial ray is more prominent (Fig. I, J). Compact sclerites preserve narrow axial cavities but the form of these is unclear (Fig. G, L)
Genus Silicunculus Bengtson, Citation1986
Type species – Silicunculus australiensis Bengtson, Citation1986.
Silicunculus australiensis Bengtson, Citation1986
A, B, I, Y, EE
Citation1986 Silicunculus australiensis Bengtson, p. 201, fig. 3
Citation2017a Silicunculus australiensis; Peel, p. 308, fig. 2A–D, E, F,H–J
Figured material. – PMU 29996 from GGU sample 225540, PMU 29998 from GGU– sample 225594, PMU 30003 from GGU sample 225595; Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone, Gustav Holm Dal, Peary Land, North Greenland (Fig. ).
Other material. – PMU 29997 from GGU sample 225592, PMU 30001 from GGU sample 225594, PMU 30000 from GGU sample 225540 (illustrated by Peel Citation2017a). About ten mainly broken spicules from GGU sample 225592, two from GGU sample 225594, and four from GGU sample 225595. Two specimens from GGU sample 271404. Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone, Gustav Holm Dal, Peary Land, North Greenland (Figs. and ).
Discussion. – Silicunculus australiensis was defined by Bengtson (Citation1986) from the Mindyallan of Queensland and described from the contemporaneous Holm Dal Formation by Peel (Citation2017a). The pentactins from Greenland, with four short, acanthose, paratangential rays (Fig. A, B, EE) and an elongate, hooked, axial ray are closely similar to the type material figured by Bengtson (Citation1986) from Australia, although specimens illustrated by Bengtson (Citation1986) are larger. The curvature between the shaft and tip of the axial ray may be uniformly concave (Fig. EE) or angular (Fig. B); the latter form is more typical of Bengtson’s (Citation1986) material. Small barbs on the axial ray were noted by Bengtson (Citation1986, fig. 2A, G) but no such spines have been seen in the Holm Dal Formation specimens. Axial cavities visible in some spicules appear to have been diagenetically enlarged (Fig. Y).
A hooked spicule from the Bagrad Formation (Cambrian Series 2, Botoman Stage) of Bateny Ridge, south-eastern Siberia, illustrated by Sugai et al. (Citation2004, pl. 1, fig. 18) is tentatively interpreted as a broken fragment of Silicunculus australiensis, as are illustrated fragments of curved spicules from the Furongian of Hunan, China, illustrated by Dong & Knoll (Citation1996, fig. 7.2, 7.19).
Silicunculus saaqqutit Peel, Citation2017a
C–F
Citation2017a Silicunculus saaqqutit Peel, p. 309, fig., 2D, K, G, L.
Figured material. – PMU 29999, holotype (Fig. E, F), PMU 30002, paratype, from GGU sample 225595, Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone, Gustav Holm Dal, Peary Land, North Greenland (Fig. ).
Other material. – One specimen from GGU sample 225594 and four from GGU sample 225595. Two specimens from GGU sample 271404. Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone, Gustav Holm Dal, Peary Land, North Greenland (Figs. and ).
Discussion .– A full description was given by Peel (2017). The short, spinose, paratangential rays and the proximal portion of the long axial ray of Silicunculus saaqqutit cannot be distinguished from the type species S. australiensis, but the distal termination of the axial ray in S. australiensis differs in being strongly hooked (Fig. A, B, EE). The distal portion of the axial ray in Silicunculus saaqqutit has up to seven short, paired, spines on its concave surface (Fig. C–E). Silicunculus bengtsoni Webby & Trotter, Citation1993 from the late Ordovician of New South Wales has a short, central, sixth ray not seen in the type species or the Greenland material.
Genus Sisamatispongia n. gen.
LSID. – urn:lsid:zoobank.org:act:1E2F0394-6671-401C-8F75-33294537F873
Type species. – Sisamatispongia erecta n. gen. n. sp. from the Holm Dal Formation (Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone), glacier feeding into head of Navarana Fjord, North Greenland (Fig. ).
Derivation of name. – From “sisamat” (Greenlandic) meaning four, with reference to the four erect rays of the spicule.
Diagnosis. – Tetractin spicules in which the four long, slender, rays curve abruptly perpendicular to their plane of origin to become sub-parallel to each.
Discussion. – Spicules of Nabaviella Mostler & Mosleh-Yazdi, Citation1976 display robust rays which are recurved towards the prominent axial ray. An axial ray is not present in Sisamatispongia erecta which has four, long and slender recurved rays. In contrast, 7–11 diverging rays may be present in Nabaviella elegans Mostler & Mosleh-Yazdi, Citation1976, from the upper Cambrian Mila Formation of Iran, but these are short, never attain parallelism, and arise from a broad central disc.
Sisamatispongia erecta n. gen. n. sp.
K, N, P
LSID. – urn:lsid:zoobank.org:act:31FC19E5-3394-4D51-A55F-1C6F91F33751
Derivation of name. – With reference to the four erect rays of the spicule.
Holotype. – PMU 31845 from GGU sample 315008, Holm Dal Formation. Lejopyge laevigata Biozone, Cambrian Series 3, Guzhangian Stage, glacier feeding into head of Navarana Fjord, North Greenland (Figs. and ).
Paratype .– PMU 31846 from GGU sample 315008, paratype, same locality as holotype.
Other material. – One specimen from GGU sample 315010, same locality as holotype.
Diagnosis. – As for genus.
Description. – Tetractin with four rays initially diverging at right angles to each other within a single plane before curving abruptly perpendicular to this inital plane and becoming sub-parallel to each other. Rays long and slender, circular in cross-section, and with scattered short, stubby, spines at their junction (Fig. P).
Discussion. – Bengtson (Citation1986, fig. 9.I) illustrated a spicule from the Mindyallan of Queensland with strongly recurved rays seemingly disposed around an elongate, tapering axial ray. A similar specimen was illustrated by Dong & Knoll (Citation1996) from Hunan, China, but seems to lack an axial ray. The recurved rays in the Australian specimen are straight and diverge distally, unlike the slender curved rays of Sisamatispongia erecta. The latter also differs in lacking an axial ray and in having small spines proximally (Fig. P).
Genus Speciosuspongia Chen & Dong, Citation2008
Type species. – Speciosuspongia hunanensis Chen & Dong, Citation2008 from the Ptychagnostus atavus Biozone (Gapparodus bisulcatus–Westergaardodina brevidens conodont Biozone), Cambrian Series 3, Drumian Stage.
Discussion. – Mehl (Citation1998, p. 1162, pl. 3, fig. 6, 9–11) gave the appropriate name “spiderpentactins” to spicules from the Gowers Formation (Upper Floran–Lower Undillan) of the Georgina Basin of Australia which are readily assigned to Speciosuspongia Chen & Dong, Citation2008, probably to the type species Speciosuspongia hunanensis from contemporaneous strata in western Hunan, China. The axial ray is transversely swollen, shaped like an inverted heart with a pointed tip, and inclined at a low angle to the plane delimited by the tips of the four paratangential rays. The latter are not equally developed but consist of two pairs. The pair most distal from the axial ray diverge from each other at an angle of almost 90 degrees. The pair closest to the axial ray diverge at 180 degrees and are often longer and narrower than the distal pair. Thus, the spicule is not radially symmetrical about the axial ray, as is often the case in pentactins, but bilaterally symmetrical about a plane which longitudinally bisects the axial ray and the angle between the distal pair of paratangential rays.
Chen & Dong (Citation2008) described a second species as Speciosuspongia wangcunensis in which the axial ray is elongate and much more slender than in Speciosuspongia hunanensis.
Speciosuspongia cf. wangcunensis Chen & Dong, Citation2008
R, X
Figured material. – PMU 31819 from GGU sample 225594, Gustav Holm Dal, Peary Land; PMU 31820 from GGU sample 315008, glacier feeding into the head of Navarana Fjord. Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone (Fig. ).
Other material. – Common in GGU samples 315008, 315010 and 315011 from Navarana Fjord; rare in samples from Gustav Holm Dal. Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone (Figs. and ).
Discussion. – Speciosuspongia wangcunensis Chen & Dong, Citation2008 was originally described from the Ptychagnostus atavus Biozone (Gapparodus bisulcatus–Westergaardodina brevidens conodont Biozone), Cambrian Series 3, Drumian Stage of Hunan, South China.
The pentactin placed here is intermediate in shape between Speciosuspongia hunanensis and the smooth or acanthose pentactins with long axial rays that occur in many spicule assemblages (Bengtson Citation1986, fig. 8; Fig. K, Q). It differs from the latter, however, in the low angle of the axial ray to the paratangential rays, in which feature it resembles Speciosuspongia hunanensis, and the paired paratangential rays. The axial ray is often curved in pentactins, but usually it is first initially perpendicular to the plane of the paratangential rays (Fig. K, T, AA). The circular cross-section and extended tapering form separate Speciosuspongia cf. wangcunensis from Speciosuspongia hunanensis where the axial ray is short, transversely flattened and inverted heart-shaped.
Fedorov & Pereladov (Citation1987, pl. 12, figs. 7,10–12) included spicules of this type in an assemblage of tetractin and pentactin spicules from the Oryctocara Biozone (Cambrian Series 2, Stage 4) of the Kuonamka Formation, northern Siberia, which they proposed as Abnormisella insperata Fedorov n. gen. n. sp.
Numerous spicules of this type occur together with slender tetractins and pentactins in which the rays are often not perpendicular to each other in GGU samples 315008 and 315009.
Genus Tallitaniqa n. gen.
LSID. – urn:lsid:zoobank.org:act:A9183420-CC13-475D-86C7-997EE97BC66E
Type species. – Tallitaniqa petalliformis n. gen. n. sp., from the Holm Dal Formation. (Cambrian Series 3, Guzhangian Stage, Lejopyge laevigata Biozone), North Greenland (Fig. ).
Derivation of name. – From “tallit” (Greenlandic), meaning arms, referring to the four rays (arms) of the tetractin spicule.
Diagnosis. – Tetractin spicule with the four perpendicular, coplanar rays. Spicules circular in cross-section, but formed on one surface into a petal-like pattern, adjacent to their point of junction, into U-shaped depressions with raised lateral margins.
Discussion. – Simple tetractin spicules are widespread in the geological record, including several morphotypes from the Holm Dal Formation assemblages (Fig. D, R, V). Tallitaniqa n. gen. is delimited by the petalliform excavation of its rays which is seemingly unreported from elsewhere. The function of the petalliform excavations is not known, but it may result from close juxtaposition with other spicules.
Tallitaniqa petalliformis n. gen. n. sp.
D, E, O, Q
LSID. – urn:lsid:zoobank.org:act:465F8273-41A4-4EA4-ABB1-06CDC8A399D0
Derivation of name. – Referring to the petal-like form of the excavations on the rays.
Holotype. – PMU 31841 from GGU sample 315008 (Fig. O, Q), Holm Dal Formation, Cambrian Series 3, Guzhangian, Lejopyge laevigata Biozone, from the western side of the glacier feeding into the head of Navarana Fjord, south-east Freuchen Land (Figs. C and ).
Paratype. – PMU 31840, from the same sample, locality and horizon as the holotype.
Other material. – About 20 specimens from GGU samples 315008–315011; same locality and horizon as the holotype.
Diagnosis. – As for genus.
Description. – Tetractin spicule with the four perpendicular, coplanar rays which curve smoothly into each other at the axis. Individual rays are circular in cross-section and taper slightly, distally. One surface of the spicule is formed into a petal-like pattern, adjacent to their flattened point of junction of the rays, with U-shaped depressions delimited laterally and axially by raised margins (Fig. O and Q). The opposing surface is uniformly convex.
Discussion. – Tallitaniqa petalliformis n. gen. n. sp. has not been observed in samples from Gustav Holm Dal. One specimen from GGU sample 315009 has a minute axial spine on the petalloid surface.
Data archiving statement
This published work and the nomenclatural acts it contains have been registered in ZooBank: http://zoobank.org/References/BB92606A-8582-482F-BC26-4A4357DC1ECE
Disclosure statement
The author reports no potential conflict of interest.
Acknowledgements
Fieldwork during the North Greenland Project (1978-80, 1984-85) was organized by the Geological Survey of Greenland (GGU). Scanning electron microscope facilities were provided by the Microscopy and Imaging unit, Evolutionary Biology Centre, Uppsala University. Reviews by J.P. Botting and B.R. Pratt are gratefully acknowledged.
References
- Babcock, L.E., 1994a: Biogeography and biofacies patterns of Middle Cambrian polymeroid trilobites from North Greenland: palaeogeographic and palaeo-oceanographic implications. Grønlands Geologiske Undersøgelse Bulletin 169, 129–147.
- Babcock, L.E., 1994b: Systematics and phylogenetics of polymeroid trilobites from the Henson Gletscher and Kap Stanton formations (Middle Cambrian), North Greenland. Grønlands Geologiske Undersøgelse Bulletin 169, 79–127.
- Babcock, L.E., Robison, R.A. & Peng, S.C., 2011: Cambrian stage and series nomenclature of Laurentia and the developing global chronostratigraphic scale. Museum of Northern Arizona Bulletin 67, 12–26.
- Babcock, L.E., Baranoski, M.T. & Cook, A.E., 2014: Cambrian (Guzhangian Stage) trilobites from Ohio, USA, and modification of the Cedaria Zone as used in Laurentia. GFF 136, 6–15.10.1080/11035897.2013.879736
- Bengtson, S., 1986: Siliceous microfossils from the Upper Cambrian of Queensland. Alcheringa 10, 195–216.10.1080/03115518608619155
- Bengtson, S., Conway Morris, S., Cooper, B.J., Jell, P.A. & Runnegar, B.N., 1990: Early Cambrian fossils from South Australia. Memoirs of the Association of Australasian Palaeontologists 9, 1–364.
- Bergström, J. & Ineson, J.R., 1988: The arthropod trace Multipodichnus from the upper Middle Cambrian (Holm Dal Formation) of central North Greenland. Meddelelser om Grønland Geoscience 20, 113–117.
- Botting, J.P. & Butterfield, N.J., 2005: Reconstructing early sponge relationships by using the Burgess Shale fossil Eiffelia globosa, Walcott. Proceedings of the National Academy of Sciences 102, 1554–1559.10.1073/pnas.0405867102
- Botting, J.P. & Muir, L.A., 2013: Spicule structure and affinities of the Late Ordovician hexactinellid-like sponge Cyathophycus loydelli from the Llanfawr Mudstones Lagerstätte, Wales. Lethaia 46, 454–469.
- Botting, J.P. & Peel, J.S., 2016: Early Cambrian sponges of the Sirius Passet biota, North Greenland. Papers in Palaeontology 2, 463–487.10.1002/spp2.2016.2.issue-4
- Botting, J.P., Muir, L.A., Xiao, S., Li, X. & LIN, J.-P., 2012: Evidence for spicule homology in calcareous and siliceous sponges: biminerallic spicules in Lenica sp. from the Early Cambrian of South China. Lethaia 45, 463–475.10.1111/let.2012.45.issue-4
- Botting, J.P., Cárdenas, P. & Peel, J.S., 2015: A crown group demosponge from the early Cambrian Sirius Passet Biota, North Greenland. Palaeontology 58, 35–43.10.1111/pala.12133
- Carrera, M.O. & Botting, J.P., 2008: Evolutionary history of Cambrian spiculate sponges: implications for the Cambrian evolutionary fauna. Palaios 23, 124–138.10.2110/palo.2006.p06-089r
- Carrera, M.O. & Maletz, J., 2014: Ordovician sponge spicules from Spitsbergen, Nevada and Newfoundland: new evidence for hexactinellid and demosponge early diversification. Journal of Systematic Palaeontology 12, 961–981.10.1080/14772019.2013.846946
- Castellani, C., Maas, A., Haug, C., Haug, J.T. & Waloszek, D., 2012: Isolated sponge spicules from the late Cambrian Alum Shale Formation (‘Orsten’ nodules of Sweden. Bulletin of Geosciences 87, 443–460.10.3140/bull.geosci.1328
- Chen, F. & Dong, X., 2008: 2008: Middle Cambrian highly modified sponge spicules from western Hunan, South China. Acta Scientiarum Naturalium Universitatis Pekinensis 44, 883–889.
- Dong, X. & Knoll, A.H., 1996: Middle and Late Cambrian sponge spicules from Hunan, China. Journal of Paleontology 70, 173–184.
- Doweld, A.B., 2016: Kuonamia, a new generic replacement name for Disparella Fedorov and Pereladov 1987 (Porifera: Hexactinellida) non Hessler 1970 (Crustacea: Isopoda). Research & Reviews: Journal of Zoological Sciences 4, 19.
- Elicki, O., 2011: First skeletal microfauna from Cambrian Series 3 of the Jordan Rift Valley (Middle East). Memoirs of the Association of Australasian Palaeontologists 42, 153–173.
- Fedorov, A.E. & Pereladov, V.S., 1987: Spicules of siliceous sponges from the Kuonamka Formation of north-eastern Siberia. In S.P Bulynnikova & I.G. Klimova (eds.): Novye vidy drevnih rastenij i bespozvonočnyh fanerozoâ Sibiri, 36–46. SNIGGIMS, Novosibirisk, [in Russian]
- Grant, R.E., 1836: Animal kingdom. In R.B. Todd (ed.): The Cyclopedia of Anatomy and Physiology, 107–118. London, Gilbert and Piper.
- Hessler, R.R., 1970: The Desmosomatidae (Isopoda: Asellota) of the Gay Head – Bermuda Transect. Bulletin of the Scripps Institution of Oceanography 15, 1–185.
- Higgins, A.K., Ineson, J.R., Peel, J.S., Surlyk, F. & Sønderholm, M., 1991: Lower Palaeozoic Franklinian Basin of North Greenland. Bulletin Grønlands Geologiske Undersøgelse 160, 71–139.
- Ineson, J.R., 1988: Lithostratigraphy and depositional setting of the Holm Dal Formation (Middle Cambrian), central North Greenland. Meddelelser om Grønland, Geoscience 20, 9–21.
- Ineson, J.R. & Peel, J.S., 1997: Cambrian shelf stratigraphy of North Greenland. Geology of Greenland Survey Bulletin 173, 1–120.
- Kouchinsky, A., Bengtson, S., Clausen, S., Gubanov, A., Malinky, J.A. & Peel, J.S., 2011: A Middle Cambrian fauna of skeletal fossils from the Kuonamka Formation, northern Siberia. Alcheringa 35, 1–67.
- Kouchinsky, A., Bengtson, S., Clausen, S. & Vendrasco, M.J., 2015: An early Cambrian fauna of skeletal fossils from the Emyaksin Formation, northern Siberia. Acta Palaeontologica Polonica 60, 421–512.
- Kozur, H.W., Mostler, H. & Repetski, J.E., 1996a: Well-preserved Tremadocian primitive Radiolaria from the Windfall Formation of the Antelope Range, Eureka County, Nevada, U.S.A. Geologische Paläontologische Mitteilungen der Universität Innsbruck 21, 245–271.
- Kozur, H.W., Mostler, H. & Repetski, J.E., 1996b: “Modern” siliceous sponges from the lowermost Ordovician (Early Ibexian-Early Tremadocian) Windfall Formation of the Antelope Range, Eureka County, Nevada, U.S.A. Geologische Paläontologische Mitteilungen der Universität Innsbruck 21, 201–221.
- Maletz, J., 2011: Radiolarian skeletal structures and biostratigraphy in the early Palaeozoic (Cambrian–Ordovician). Palaeoworld 20, 116–133.10.1016/j.palwor.2010.12.007
- Maletz, J. & Bruton, D.L., 2007: Lower Ordovician (Chewtonian to Castlemainian) radiolarians of Spitsbergen. Journal of Systematic Palaeontology 5, 245–288.10.1017/S1477201907002039
- Mehl, D., 1998: Porifera and Chancelloriidae from the Middle Cambrian of the Georgina Basin, Australia. Palaeontology 41, 1153–1182.
- Mostler, H., 1985: Neue heteractinide Spongien (Calcispongia) aus dem Unter- und Mittelkambrium Südwestsardinien. Berichte des naturwissenschaftlichen-medizinischen Verein Innsbruck 72, 7–32.
- Mostler, H., 1996: Polyactinellide Schwämme, eine auf das Paläzoikum beschränkte Calcispongien-Gruppe. Geologische Paläontologische Mitteilungen der Universität Innsbruck 21, 223–243.
- Mostler, H. & Mosleh-Yazdi, A., 1976: Neue Poriferen aus oberkambrischen Gesteinen der Milaformationen im Elburzgebirge (Iran). Geologische Paläontologische Mitteilungen der Universität Innsbruck 5, 1–36.
- Peel, J.S. (ed.), 1988a: Stratigraphy and palaeontology of the Holm Dal Formation (late Middle Cambrian), central North Greenland. Meddelelser om Grønland Geoscience 20, 1–168.
- Peel, J.S., 1988b: Molluscs of the Holm Dal Formation (late Middle Cambrian), central North Greenland. Meddelelser om Grønland Geoscience 20, 145–168.
- Peel, J.S. (ed.), 1994: Palaeontology, stratigraphy and environmental setting of Middle Cambrian outer shelf deposits, North Greenland. Grønlands Geologiske Undersøgelse Bulletin 169, 1–155.
- Peel, J.S., 2017a: First records from Laurentia of some middle Cambrian (Series 3) sponge spicules. Alcheringa 41, 306–314.10.1080/03115518.2017.1282983
- Peel, J.S., 2017b: The oldest pelmatozoan encrusted hardground and holdfasts from Laurentia (Cambrian Series 2–3). GFF 139, 195–204.10.1080/11035897.2017.1347196
- Peel, J.S. & Willman, S., 2018: The Buen Formation (Cambrian Series 2) Biota of North Greenland. Papers in Palaeontology. doi:10.1002/spp2.11120 (published on line)
- Peel, J.S., Streng, M., Geyer, G., Kouchinsky, A. & Skovsted, C.B., 2016: Ovatoryctocara granulata assemblage (Cambrian Series 2–Series 3 boundary) of Løndal, North Greenland. Australasian Palaeontological Memoirs 49, 241–282.
- Peng, S., Babcock, L.E., Zuo, J. LIn, H., Zhu, X., Yang, X., Robison, R.A., Qui, Y., Bagnoli, G. & Chen, Y., 2009: The Global Boundary Stratotype Section and Point (GSSP) of the Guzhangian Stage (Cambrian) in the Wuling Mountains, Northwestern Hunan, China. Episodes 32, 41–55.
- Rigby, J.K., 1975: Some unusual hexactinellid sponge spicules from the Cambrian Wilberns Formation of Texas. Journal of Paleontology 49, 412–415.
- Rigby, J.K., 1986: Sponges of the Burgess Shale (Middle Cambrian), British Columbia. Palaeontographica Canadiana 2, 1–105.
- Rigby, J.K. & Collins, D., 2004: Sponges of the Middle Cambrian Burgess Shale and Stephen Formations, British Columbia. Royal Ontario Museum Contributions in Science 1, 1–155.
- Rigby, J.K. & Hou, X.-G., 1995: Lower Cambrian demosponges and hexactinellid sponges from Yunnan, China. Journal of Paleontology 69, 1009–1019.10.1017/S0022336000037999
- Robison, R.A., 1988: Trilobites of the Holm Dal Formation (late Middle Cambrian), central North Greenland. Grønlands Geologiske Undersøgelse Bulletin 169, 23–103.
- Robison, R.A., 1994: Agnostoid trilobites from the Henson Gletscher and Kap Stanton formations (Middle Cambrian), North Greenland. Grønlands Geologiske Undersøgelse Bulletin 169, 25–77.
- Schulze, F.E., 1887: Report on the Hexactinellida collected by H.M.S. ‘Challenger’ during the years 1873-1876. Report on the Scientific Results of the Voyage of H.M.S. ‘Challenger’, 1873-1876. Zoology 21, 1–514.
- Shabanov, Yu.Ya., Astashkin, V.A., Pegel, T.V., Egorova, L.I., Zhuravleva, I.T., Pelman, Yu.L., Sundukov, V.M., Stepanova, M.V., Sukhov, S.S., Fedorov, A.B., Shiskin, B.B., Vaganova, N.V., Ermak, V.I., Ryabukha, K.V., Yadrenkina, A.G., Abaimova, G.P., Lopushinskaya, T.V., Sychev, O.V. & Moskalenko, T.A., 1987: Nizhnii paleozoi yugo-zapadnogo sklona Anabarskoi anteklizy (pro materialum bureniya). Nauka, Novosibirsk, 206 pp. [in Russian]
- Skovsted, C.B., 2006: Small shelly fauna from the upper Lower Cambrian Bastion and Ella Island Formations, North-East Greenland. Journal of Paleontology 80, 1087–1112.10.1666/0022-3360(2006)80[1087:SSFFTU]2.0.CO;2
- Steiner, M., Mehl, D., Reitner, J. & Erdtmann, B.-D., 1993: Oldest entirely preserved sponges and other fossils from the lowermost Cambrian and a new facies reconstruction of the Yangtze Platform (China). Berliner Geowissenschaften Abhandlungen (E) 9, 293–329.
- Sugai, Y., Iwata, K., Sennikov, N.V., Obut, O.T. & Khlebnikova, T.V., 2004: Sponge spicules from the Lower Cambrian of Bateny Ridge and Gorny Altai (Bagrad and Shashkunar formations). Novosti paleontologii i stratigrafii. Pridozhenyie k Zhurnaly Geologiya i Geofyzika 6, 59–73.
- Torsvik, T.H. & Cocks, L.R.M., 2017: Earth History and Palaeogeography. Cambridge UP, 332 pp.10.1017/9781316225523
- Webby, B.D. & Trotter, J., 1993: Ordovician sponge spicules from New South Wales, Australia. Journal of Paleontology 67, 20–28.
- Won, M. & Below, R., 1999: Cambrian Radiolaria from the Georgina Basin, Queensland, Australia. Micropaleontology 45, 325–363.10.2307/1486119
- Won, M., Iams, W.J., Reed, K. & Below, R., 2007: Review of the Family Anakrusidae and its relationship to Kometia (Porifera: Hexactinellida). Journal of the Paleontological Society of Korea 23, 73–91.
- Wu, W., 2004: Fossil sponges from the early Cambrian Chengjiang Fauna, Yunnan, China. Nanjing Institute of Geology and Palaeontology, Chinese Academy of Sciences, Nanjing, China, 1–190 [In Chinese with English abstract].
- Wu, W., Yang, A.-H., Janussen, D., Steiner, M. & Zhu, M.-Y., 2005: Hexactinellid sponges from the Early Cambrian black shale of South Anhui, China. Journal of Paleontology 79, 1043–1051.
- Xiao, S., Hu, J., Yuan, X., Parsley, R.L. & Cao, R., 2005: Articulated sponges from the lower Cambrian Hetang Formation in southern Anhui, South China: their age and implications for the early evolution of sponges. Palaeogeography, Palaeoclimatology, Palaeoecology 220, 89–117.10.1016/j.palaeo.2002.02.001
- Yang, X.-L. & Zhao, Y.-L., 2000: Sponges of the Lower Cambrian Niutitang Formation Biota in Zunyi, Guizhou, China. Journal of Guizhou University of Technology (Natural Science Edition) 29, 30–36.
- Zell, M.G. & Rowell, A.J., 1988: Brachiopods of the Holm Dal Formation (late Middle Cambrian), central North Greenland. Meddelelser om Grønland Geoscience 20, 119–144.
- Zhang, X. & Pratt, B.R., 1994: New and extraordinary sponge spicule assemblage from China. Geology 22, 43–46.10.1130/0091-7613(1994)022<0043:NAEECS>2.3.CO;2
- Zheng, Y., Li, Y. & Guo, J., 2012: Sponge spicule fossils of Qiongzhusian Age in Zhenba County of southern Shaanxi Province. Journal of Earth Sciences and Environment 34, 24–30.