
ABSTRACT
Two described specimens of the gastropod Bellerophon tenuifascia J. de C. Sowerby, 1824, from the Carboniferous (Asbian) of the Isle of Man, United Kingdom, preserve repaired injuries resulting from environmental breakage and failed, non-lethal, predatory attacks. Deep scars attributed to predatory attacks are assigned to the ichnofossil Caedichnus and were focused on the mid-dorsal area of the aperture. Abundant small-scale scallops and chipping of growth lamellae distributed around the apertural margin are mainly the result of environmental damage. A deeply incised injury in the aperture of one specimen was probably a fatal injury.
Introduction
A multitude of processes are involved in the breakdown or destruction of mollusc shells (Zuschin et al. Citation2003), but many recent palaeontological studies have focussed on predation on account of its widely accepted role as a driving force in evolution (Vermeij Citation1977, Citation1987, Citation2013; Vermeij et al. Citation1981; Alexander & Dietl Citation2003; Bicknell & Paterson Citation2018; Budd & Mann Citation2019). A general conclusion is that predation is the dominant agency for shell destruction in marine gastropods in most environments.
The gastropod shell preserves a record of life-long incremental growth, but it is also an archive of the discrete external events that have affected the individual during its life. Foremost amongst these are attacks from durophagous predators that break the shell to gain access to the soft parts within. If successful, such attacks may destroy the shell completely, but unsuccessful attacks produce non-lethal injuries which may be followed by the formation of new shell material as the gastropod strives to repair its damaged shell. Comparison of these repaired injuries with injuries in present day shells has enabled the recognition of similar damage in fossils from as far back as the Cambrian (Skovsted et al. Citation2007; Bicknell & Paterson Citation2018). Cases of failed predation with subsequent shell repair provide the best evidence of the rise of predation in the geological record, although the record may be difficult to interpret (Vermeij Citation1987; Alexander & Dietl Citation2003; Ebbestad et al. Citation2009; Budd & Mann Citation2019). Destroyed shells obviously leave little record, although some of the fragments may be diagnostic of the predator and the prey (Vermeij Citation1987; Cadée Citation1994, Citation1995; Zuschin et al. Citation2003; Harper & Kelley Citation2012; Salamon et al. Citation2014, Citation2018; Leighton et al. Citation2016) while the damage caused by lethal attacks (Clarkson et al. Citation1995) can be difficult to recognise in fossil material on account of imperfect preservation, taphonomic effects or preparation.
Bellerophon is the eponymous genus for a group of isostrophic, bilaterally symmetrical gastropods known from the Cambrian to the Triassic (Knight et al. Citation1960; Kaim & Nützel Citation2010; Bouchet et al. Citation2017). In recent years several studies have focussed on the repair of non-lethal predatory injuries in bellerophontiform gastropods (Horný Citation1997a, b, Citation1998; Ebbestad Citation1998; Ebbestad et al. Citation2009). Interestingly, Knight (Citation1941, pl. 11, ) illustrated without comment a repaired deep V-shaped injury in the apertural margin while describing type material of the type species of the genus from the Devonian of Germany. Ebbestad et al. (Citation2009) employed terminology developed by Alexander & Dietl (Citation2003) and surveyed and classified the described cases of shell repair in bellerophontiform molluscs from throughout the Palaeozoic, attributing most of them to attacks from unknown predators, although cephalopods are likely culprits (Peel Citation1984; Kröger Citation2004; Klug Citation2007). Few cases of described injuries in bellerophontoideans have been attributed to abiogenic agents, an exception being the bizarre record of a crushed bellerophontoidean by Horný (Citation1998), following the view expressed by Vermeij (Citation1987) that attempted predation is the primary cause of shell breakage.
Many extant gastropod shells, however, display injuries that are not associated with durophagous attacks. Drilling by gastropods and other invertebrates leaves prominent perforations in the shell and such holes have a long, albeit often problematic, geological record (Bengtson & Zhao Citation1992; Carriker & Yochelson Citation1968; Conway Morris & Bengtson Citation1994; Baumiller Citation1996; Reyment Citation1999; Kelley & Hansen Citation2003). Physical abrasion or breakage of the apertural margin is widespread, particularly in groups such as limpets living in high energy environments (Cadée Citation1998, Citation1999; Shanks & Wright Citation1986; Zuschin et al. Citation2003), but may also result from storm activity (Savazzi Citation1991). Some advanced predatory gastropods often damage their apertural margin while using it as a tool to prise or chip open bivalve shells (Nielsen Citation1975; Fretter & Graham Citation1994; Alexander & Dietl Citation2003; Dietl Citation2003a, Citationb, Citation2004; Dietl et al. Citation2010).
This paper describes two specimens of Bellerophon from the lower Carboniferous of the Isle of Man () which preserve evidence of biogenic and abiogenic injuries during life. The specimens are unusual amongst Palaeozoic bellerophontoideans in preserving evidence of both biogenic and abiogenic injuries. Characteristic of the abiogenic damage is that the injuries (scalloping and repeated chipping) commonly occur across most of the extent of the apertural margin with relatively little penetration into the earlier growth stages. In contrast, injuries with relatively deep penetration of the apertural margin often result from predatory attacks, although the two styles of damage are not exclusive (Ebbestad et al. Citation2009; Stafford et al. Citation2015). Additionally, one specimen may have succumbed to a major predatory attack but several earlier, deeply penetrative repaired injuries are interpreted as failed, non-lethal, attacks.
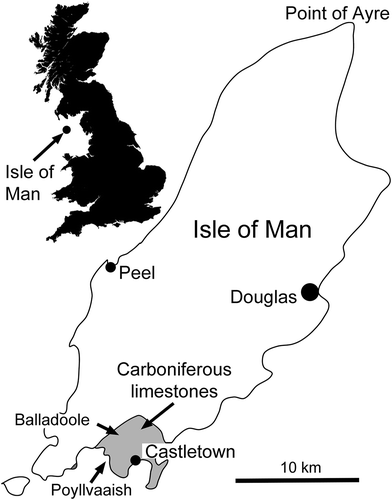
Figure 1. Locality map showing outcrops of Carboniferous limestones around Castletown, Isle of Man.
Description of material
The two specimens of Bellerophon are described from a small outcrop of lower Carboniferous (Asbian) limestones of the Balladoole Formation (including the Poolvash Limestone, equivalent to the Poyllvaaish Formation of Lewis Citation1930; Dickson et al. Citation1987; Quirk et al. Citation1990) west of Castletown, at the southern tip of the Isle of Man, UK (54⁰ 05ꞌN, 4⁰ 40ꞌW; ). Several other specimens from the same area in the collection from the Royal Museum of Scotland, Edinburgh (RMSE), show well preserved growth ornamentation without significant repaired injuries or disturbances. Specimen RMSE 4199 () is from the Dunlop Collection and identified as Bellerophon tenuifascia J. de C. Sowerby, 1824, Poolvash; as such it was mentioned by Weir (Citation1931, p. 790). Specimen RMSE 1958.1.2968 is clearly the same species and was collected at Balladoole ().
Figure 2. Bellerophon from the Balladoole Formation, Carboniferous, of the Isle of Man. A, B, specimen RMSE 4199, from Poolvash, in lateral (A) and dorsal (B) views with arrow showing principal fracture; for interpretation see Fig. 2. C–F, specimen RMSE 1958.1.2968, from Balladoole in oblique (C, D) and dorsal (E) views, locating predatory fractures a–d; detail of dorsum (F) with abiogenic disturbances arrowed, locating fracture b and selenizone (s). Scale bars: 5 mm.
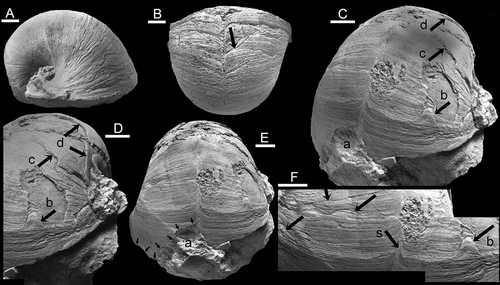
Specimen RMSE 1958.1.2968 has a maximum preserved length of about 42 mm. The specimen surface is partly disturbed by post-depositional cleavage and recrystallisation half a whorl back from the latest preserved growth stage, but it is evident that major repaired oblique fractures represent at least two substantial injuries interpreted as non-lethal predatory attacks (c and d in ), although more may be present. Both of these fractures retain step-like adapical margins, with the post-injury new shell being formed from underneath the pre-injury old shell ().
The aperture is broken in specimen RMSE 1958.1.2968 and, on the left side (right side as viewed in ), the shell has exfoliated from the internal mould to form a broken margin oblique to the growth ornamentation. A deep V-shaped fracture on the right side of the dorsal surface (left in ) can be followed from near the lateral shoulder as far as the median dorsal selenizone (a in ), marked by arrows). It probably extends across the full width of the shell, although it is not visible on the left dorsal area () due to the exfoliation. In detail, the fractured margin is seen to consist of several shallow scallops rather than a single break, indicating that the shell was broken away in several separate actions that produced an increasingly deep penetration as the selenizone was approached. The fracture traverses lunulae in the selenizone but then seems to have followed the selenizone towards the aperture since visible growth lines on the left side (right side as viewed in ) are unaffected. In total, the fracture extends back through one-eighth of the whorl. The nature of its broken margin and relationship to the sediment indicate that it was formed preburial. Evidence of repair after fracturing is not visible and the predatory attack may have been fatal.
Fracture c extends transversely across the dorsum in a series of small concave scallops. It is also an embayment but it can only be traced across the left side of the dorsum (right side in due to poor preservation in the mid-dorsal area. It increases in curvature backwards as the median selenizone is approached, in similar fashion to fracture a but it is not as strongly expressed. Subsequent shell growth is quite regular initially in the central dorsal area abapical to the scar, with parallel growth lines, but becomes increasingly disturbed as growth proceeds. The disturbances include slight realignment of the growth lines and local or more laterally extensive chipped lamellae (, arrows) and are likely abiogenic in origin. However, in one continuous line of disturbance, a small, but deep, V-shaped injury may reflect a predatory attack , arrow b). This is a typical non-lethal predation scar and the continued growth of the gastropod does not seem to have been affected; growth lines indicate that it was quickly filled. The chipped apertural margin can be traced towards the selenizone (s in ).
Fracture d, the oldest injury, extends as a broad, adaperturally concave fracture from the left umbilical area (right in ) across the mid-dorsum. This fracture was filled with new shell with regular growth lines that was broken subsequently at fracture c ().
About half a whorl of the dorsal surface is visible prior to injury d, but partially covered by inductural deposits. Repaired injuries have not been observed and the regular growth lines are only disturbed by occasional chipping.
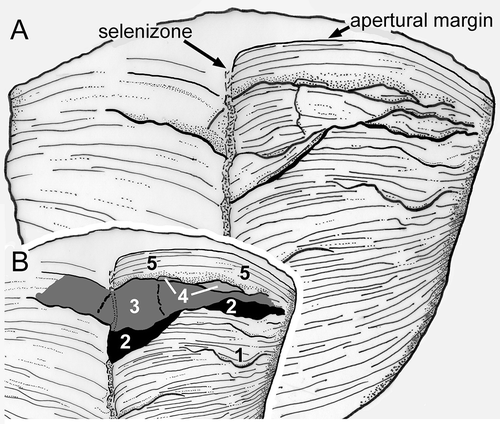
Specimen RMSE 4199 has a maximum preserved length of about 38 mm. The undisturbed apertural margin is preserved on the right side (, ). Most of the early part of the final whorl, and the aperture, consists of very fine, shallowly inclined growth laminations but these become increasingly disturbed during the last one-eighth of the whorl (). Many of the lamellae in this part of the shell are chipped and there are numerous shallow scallops that can be traced over a large part of the dorsal surface (area 1 in ). A significant fracture cuts back more than 30 percent of its transverse extent at the selenizone on the right side before being displaced forward along the selenizone and traversing the left dorsal area. The injury is partially filled by new shell (area 2 in ) but this repair is itself broken adaperturally and a new area of shell was subsequently formed across the entire dorsum (area 3 in ). Area 3 appears to be composite, with a filled repair near the selenizone, but this is not clearly delimited. Area 4 represents the trace of a subsequent growth episode emerging from under the broken edge of area 3, but most of it was broken away. Both of these terminate adaperturally in a deep transverse furrow on the right side representing a major halt in growth, although the course of this on the left side is uncertain. Beyond this, fine and regular growth lamellae define the area to the preserved apertural margin (area 5 in ).
Figure 3. Bellerophon from the Balladoole Formation, Carboniferous, of the Isle of Man. Sketch (A) and interpretation (B) of specimen RMSE 4199, from Poolvash, locating fractures and successive growth areas discussed in the text. Compare with .
Biogenic and abiogenic injuries
The shell of Bellerophon tenuifascia is thickened in the umbilical region on account of the narrowing of the umbilici by the reflexed lip (). Dorsally, individual growth lamellae are thin and feather-edged, lying at a low angle to the shell surface, and they are therefore susceptible to minor damage of the shell margin. An undisturbed apertural margin is preserved in specimen RMSE 4199 on the right side (, ). In many other species of Bellerophon, the growth lamellae are inclined at a higher angle to the dorsal surface, resulting in a rounded and robust apertural margin (Yochelson Citation1960). The presence of a tapering, feather-edged margin may indicate that gastropod soft parts did not extend beyond the apertural margin whereas the opposite may be true in species with a well-rounded margin (MacClintock Citation1967).
The frequent shallow chipping of growth lamellae at the apertural margin is dominantly abiogenic in origin, and this is likely true also for the more significant transverse injuries which follow growth ornamentation, i.e., the apertural margin. The latter include the prominent transverse depression separating areas 3 and 4 from area 5 in specimen RMSE 4199 (, see also ). The injury subsequently filled by new shell of area 2 () probably represents a predatory attack on account of its depth and focus on the selenizone. Scalloped injuries on the flanks (, ) that penetrate just a short distance may reflect predatory attacks or have an abiogenic origin.
While predators are considered to be the dominant cause of shell damage (Vermeij Citation1987; Alexander & Dietl Citation2003; Zuschin et al. Citation2003), gastropods living in littoral environments, especially the surf zone, are often subjected to mechanical damage from rolling stones (Shanks & Wright Citation1986). Repaired injuries have been widely described in patelliform shells (Bulkley Citation1968; Cadée Citation1998, Citation1999) but they also occur in other littoral forms (Rafaelli Citation1978). Cadée (Citation1999) noted that up to 75 percent of specimens of Nacella in the surf zone on King George Island (Antarctica) usually showed more than one repair per shell resulting from mechanical damage by stones and ice blocks. Storm damage may also cause chipping of the apertural margin in lower energy environments (Savazzi Citation1991; Zuschin et al. Citation2003).
At the present day, relatively shallow, transverse repaired injuries are seen in active predators such as the neogastropods Busycon, Buccinum, Fasciolaria and Sinistrofulgur which use the apertural margin to prise or chip open bivalve shells (Wells Citation1958; Nielsen Citation1975; Fretter & Graham Citation1994; Alexander & Dietl Citation2003; Dietl Citation2003a, Citationb, Citation2004; Dietl et al. Citation2010). Neither neogastropods nor this style of attack have been recognised in Palaeozoic gastropods.
Bellerophon specimen RMSE 1958.1.2968 displays three major injuries which are attributed to predatory attacks (). One of these (a in ) is a V-shaped scar extending about 40 degrees back into the whorl from the apertural margin and was probably lethal. In terms of shape, it closely resembles repaired injuries described by Ebbestad (Citation1998, fig. 5, injury A) in Bucania from the Ordovician of Sweden, and by Ebbestad et al. (Citation2009, ) in Bellerophon from the Carboniferous of Belgium. With a depth:width ratio of 0.6, the injury comfortably exceeds the parameter 0.3 used by Stafford et al. (Citation2015) to define the ichnofossil Caedichnus. It is an embayment in the terminology of Alexander & Dietl (Citation2003) and Ebbestad et al. (Citation2009). The location of the deepest part of the injury in the area of the slit and selenizone may reflect structural weakness (Lindström Citation2003; Lindström & Peel Citation2005; Ebbestad Citation1998; Ebbestad et al. Citation2009), but also reflect a focus on the chemical signal from the exhalant current (Kohn Citation1961; Sakata Citation1989; Ferner & Weissburg Citation2005).
Injuries c and d are also embayments but they can only be traced across the left side of the dorsum (right side in ) due to poor preservation in the mid-dorsal area. As with injury a, they consist of a series of concave scallops in the apertural margin but subsequent formation of new shell indicates that they represent non-lethal attacks. Initially, this new shell is quite regular with parallel growth lines; later disturbances such as slight realignment of the growth lines and chipped lamellae (), arrows) are probably mainly abiogenic. With a backwards penetration of almost a quarter of a whorl, fracture d is by far the largest injury experienced by the gastropod, substantially greater than the presumed fatal injury in the outer lip (a in ).
The increase in chipping and scalloping of the apertural margin in the later growth stages of the described specimens suggests that they may have occupied higher energy environments when they attained a larger size. However, in view of their derivation from reefoid limestone masses, probably representing living sites with considerable local variation in the degree of exposure, this change may have been coincidental rather than representing either environmental change or a distinct migration. A lack of repaired injuries resulting from non-lethal predatory attacks in the earlier growth stages is a relatively common feature in affected gastropods and can have several causes (Vermeij Citation1987; Dietl & Alexander Citation1998; Alexander & Dietl Citation2003; Griffiths & Gosselin Citation2008; Lindström & Peel Citation2005). Small specimens may have been unattractive (too small) for the predator, or they may have succumbed to attack and the lethal injuries to their shells therefore remained unrepaired. Thus, large specimens were better able to survive attacks, as confirmed by their repaired injuries, with increase in size providing a partial refuge against predation. Small specimens of the prey and larger, more mature, specimens may have occupied different environmental niches (Underwood Citation1979) which may have affected their availability or attraction to potential predators. Possibly only larger prey occupied areas or niches frequented by their hunters. The latter explanation is compatible with the increased abiogenic chipping in later growth stages, but it is clear that a larger sample is required to underpin the speculation.
Acknowledgments
Lyall I. Anderson kindly arranged the loan of material from the Royal Scottish Museum (National Museums of Scotland), Edinburgh. Jan O.R. Ebbestad (Uppsala) and Patricia H. Kelley (Wilmington, North Carolina) are thanked for reviewing the manuscript.
Disclosure statement
No potential conflict of interest was reported by the author.
Related Research Data
References
- Alexander, R.R. & Dietl, G.P., 2003: The fossil record of shell-breaking predation on marine bivalves and gastropods. In P.H. Kelley, M. Kowalewski, & T.A. Hansen (eds.): Predator-prey interactions in the Fossil Record, 141–176. New York: Kluwer Academic/Plenum Publishers.
- Baumiller, T.K., 1996: Boreholes in the Middle Devonian blastoid Heteroschisma and their implications for gastropod drilling. Palaeogeography, Palaeoclimatology, Palaeoecology 123, 343–351. doi:10.1016/0031-0182(95)00124-7
- Bengtson, S. & Zhao, Y., 1992: Predatorial borings in Late Precambrian mineralized exoskeletons. Science 257, 367–369. doi:10.1126/science.257.5068.367
- Bicknell, R.D. & Paterson, J.R., 2018: Reappraising the early evidence of durophagy and drilling predation in the fossil record: implications for escalation and the Cambrian Explosion. Biological Reviews 93, 754–784. doi:10.1111/brv.12365
- Bouchet, P., Rocroi, J.-P., Hausdorf, B., Kaim, A., Kano, Y., Nützel, A., Parkhaev, P., Schrödl, M., & Strong, E.E., 2017: Revised classification, nomenclator and typification of gastropod and monoplacophoran families. Malacologia 61, 1–526
- Budd, G.A. & Mann, R.P., 2019: Modeling durophagous predation and mortality rates from the fossil record of gastropods. Paleobiology. doi:10.1017/pab.2019.2
- Bulkley, B.T., 1968: Shell damage and repair in five members of the genus Acmaea. Veliger 11(supplement), 64–66
- Cadée, G.C., 1994: Eider, shelduck, and other predators, the main producers of shell fragments in the Wadden Sea, palaeoecological implications. Palaeontology 37, 181–202
- Cadée, G.C., 1995: Birds as producers of shell fragments in the Wadden Sea, in particular the role of the Herring gull. Geobios 28(supplement 1), 77–85. doi:10.1016/S0016-6995(95)80155-3
- Cadée, G.C., 1998: The fate of shells of the Antarctic limpet Nacella. Circumpolar Journal 12, 60–63
- Cadée, G.C., 1999: Shell damage and shell repair in the Antarctic limpet Nacella concinna from King George Island. Journal of Sea Research 41, 149–161. doi:10.1016/S1385-1101(98)00042-2
- Carriker, M.R. & Yochelson, E.L., 1968: Recent gastropod boreholes and Ordovician cylindrical borings. U.S. Geological Survey Professional Paper 593B, B1–B26
- Clarkson, E.N.K., Harper, D.A.T., & Peel, J.S., 1995: Taxonomy and palaeoecology of the mollusc Pterotheca from the Ordovician and Silurian of Scotland. Lethaia 28, 101–114. doi:10.1111/j.1502-3931.1995.tb01600.x
- Conway Morris, S. & Bengtson, S., 1994: Cambrian predators: possible evidence from boreholes. Journal of Paleontology 68, 1–23
- Dickson, J.A.D., Ford, T.D., & Swift, A., 1987: The stratigraphy of the Carboniferous rocks around Castletown, Isle of Man. Proceedings of the Yorkshire Geological Society 46, 203–228.
- Dietl, G.P., 2003a: Interaction strength between a predator and dangerous prey: Sinistrofulgur predation on Mercenaria. Journal of Experimental Marine Biology and Ecology 289, 287–301. doi:10.1016/S0022-0981(03)00047-9
- Dietl, G.P., 2003b: Coevolution of a marine gastropod predator and its dangerous bivalve prey. Biological Journal of the Linnaean Society 80, 409–436. doi:10.1046/j.1095-8312.2003.00255.x
- Dietl, G.P., 2004: Origins and circumstances of adaptive divergence in whelk feeding behavior. Palaeogeography, Palaeoclimatology, Palaeoecology 208, 279–291. doi:10.1016/j.palaeo.2004.03.011
- Dietl, G.P. & Alexander, R.R., 1998: Shell repair frequencies in whelks and moon snails from Delaware and southern New Jersey. Malacologia 39, 151–165
- Dietl, G.P., Durham, S.R., & Kelley, P.H., 2010: Shell repair as a reliable indicator of bivalve predation by shell-wedging gastropods in the fossil record. Palaeogeography, Palaeoclimatology, Palaeoecology 296, 174–184. doi:10.1016/j.palaeo.2010.07.013
- Ebbestad, J.O.R., 1998: Multiple attempted predation in the Middle Ordovician gastropod Bucania gracillima. GFF 120, 27‒33. doi:10.1080/11035899801201027
- Ebbestad, J.O.R., Lindström, A., & Peel, J.S., 2009: Predation on bellerophontiform molluscs in the Palaeozoic. Lethaia 42, 469‒485
- Ferner, M.C. & Weissburg, M.J., 2005: Slow-moving predatory gastropods track prey odors in fast and turbulent flow. Journal of Experimental Biology 208, 809–819. doi:10.1242/jeb.01438
- Fretter, V. & Graham, A., 1994: British prosobranch molluscs. Revised and updated edition, 820 pp. London: The Ray Society
- Griffiths, A.M. & Gosselin, L.A., 2008: Ontogenetic shift in susceptibility to predators in juvenile northern abalone, Haliotis kamtschatkana. Journal of Experimental Marine Biology and Ecology 360, 85–93. doi:10.1016/j.jembe.2008.04.004
- Harper, E.M. & Kelley, P.H., 2012: Predation on bivalves. In P.A. Selden (ed.): Treatise on Invertebrate Paleontology. Part N, revised Vol. 1, chapter 22, 1–21. Treatise online 44. Lawrence: Kansas University Press.
- Horný, R.J., 1997a: Shell breakage and repair in in explanate bellerophontoidean gastropods from the Middle Ordovician of Bohemia. Vestník Ceského Geologického Ústavu 72, 157–168
- Horný, R.J., 1997b: Shell breakage and repair in Sinuitopsis neglecta (Mollusca, Tergomya) from the Middle Ordovician of Bohemia. Casopis Národniho Muzea 166, 137–142
- Horný, R.J., 1998: A large injury and shell repair in Boiotremus incipiens (Mollusca, Gastropoda) from the Silurian of Bohemia. Bulletin of the Czech Geological Survey 73, 343–345
- Kaim, A. & Nützel, A., 2010: Dead bellerophontids walking — the short Mesozoic history of the Bellerophontoidea (Gastropoda). Palaeogeography, Palaeoclimatology, Palaeoecology 308, 190–199
- Kelley, P.H. & Hansen, T.A., 2003: The fossil record of drilling predation on bivalves and gastropods. In P.H. Kelley, M. Kowalewski, & T.A. Hansen (eds.): Predator–prey interactions in the Fossil Record. 113–139. New York: Kluwer Academic/Plenum Publishers.
- Klug, C., 2007: Sublethal injuries in Early Devonian cephalopod shells from Morocco. Acta Palaeontologica Polonica 52, 749–759
- Knight, J.B., 1941: Paleozoic gastropod genotypes. Geological Society of America Special Papers 32, 1–510
- Knight, J.B., Cox, L.R., Keen, A.M., Batten, R.L., Yochelson, E.L., & Robertson, R., 1960: Systematic descriptions, R.C. Moore, ed., Treatise on invertebrate paleontology, Part I, Mollusca 1, 169–310. Boulder: Geological Society of America and University of Kansas Press
- Kohn, A.J., 1961: Chemoreception in gastropod molluscs. American Zoologist 1, 291–308
- Kröger, B., 2004: Large size sublethal injuries in Middle Ordovician orthocerids. GFF 126, 311–316
- Leighton, L.R., Chojnacki, N.C., Stafford, E.S., Tyler, C.L., & Schneider, C.L., 2016: Categorization of shell fragments provides a proxy for environmental energy and predation intensity. Journal of the Geological Society 173, 711–715. doi:10.1144/jgs2015-086
- Lewis, H.P., 1930: The Avonian succession in the south of the Isle of Man. Quarterly Journal of the Geological Society of London 86, 234–290. doi:10.1144/GSL.JGS.1930.086.01-04.11
- Lindström, A., 2003: Shell breakage in two pleurotomarioid gastropods from the Upper Carboniferous of Texas, and its relation to shell morphology. GFF 125, 39–46. doi:10.1080/11035890301251039
- Lindström, A. & Peel, J.S., 2005: Repaired injuries and shell form in some Palaeozoic pleurotomarioid gastropods. Acta Palaeontologica Polonica 50, 697–704
- MacClintock, C., 1967: Shell structure of patelloid and bellerophontoid gastropods (Mollusca). Peabody Museum of Natural History Yale University Bulletin 22, 140
- Nielsen, C., 1975: Observations on Buccinum undatum L. attacking bivalves and on prey responses, with a short review on attack methods of other prosobranchs. Ophelia 13, 87–108. doi:10.1080/00785326.1974.10430593
- Peel, J.S., 1984: Attempted predation and shell repair in Euomphalopterus (Gastropoda) from the Silurian of Gotland. Bulletin of the Geological Society of Denmark 32, 163–168
- Quirk, D.G., Ford, T.D., King, J.A., Roberts, I.L., Postance, R.B., & Odell, I., 1990: Enigmatic boulders and syn-sedimentary faulting in the Carboniferous Limestone of the Isle of Man. Proceedings of the Yorkshire Geological Society 48, 99–113.
- Rafaelli, D.G., 1978: The relationship between shell injuries, shell thickness and habitat characteristics of the intertidal snail Littorina rudis Maton. Journal of Molluscan Studies 44, 166–170
- Reyment, R.A., 1999: Drilling gastropods. In Savazzi (ed.): Functional morphology of the invertebrate skeleton. 197–204. Chichester: John Wiley & Sons.
- Sakata, K., 1989: Feeding attractants and stimulants for marine gastropods. Bioorganic Marine Chemistry 3, 115–129
- Salamon, M.A., Gorzelak, P., Niedźwiedecki, R., Trzęsiok, D., & Baumiller, T.K., 2014: Trends in shell fragmentation as evidence of mid-Paleozoic changes in marine predation. Paleobiology 40, 14–23. doi:10.1666/13018
- Salamon, M.A., Leśko, K., & Gorzelak, P., 2018: Experimental tumbling of Dreissena polymorpha: implications for recognizing durophagous predation in the fossil record. Facies, 64(10), 1–22. doi:10.1007/s10347-018-0522-7
- Savazzi, E., 1991: Constructional morphology of strombid gastropods. Lethaia 24, 311–331
- Shanks, A.L. & Wright, W.G., 1986: Adding teeth to wave action: destructive effects of wave-born rocks on intertidal organisms. Oecologia 69, 420–428. doi:10.1007/BF00377065
- Skovsted, C.B., Brock, G.A., Lindström, A., Peel, J.S., Paterson, J.R., & Fuller, M.K., 2007: Early Cambrian record of failed durophagy and shell repair in an epibenthic mollusc. Biology Letters 3, 314‒317
- Sowerby, J. de, C., 1824: The Mineral Conchology of Great Britain, Vol. 5, 65–138.London: B. Meredith.
- Stafford, E.S., Dietl, G.P., Murray, P., Gingras, P., & Leighton, L.R., 2015: Caedichnus, a new ichnogenus representing predatory attack on the gastropod shell aperture. Ichnos 22, 87–102. doi:10.1080/10420940.2015.1031899
- Underwood, A.J., 1979: The ecology of intertidal gastropods. Advances in Marine Biology 16, 111–210
- Vermeij, G.J., 1977: The Mesozoic marine revolution: evidence from snails, predators and grazers. Paleobiology 3, 245–258. doi:10.1017/S0094837300005352
- Vermeij, G.J., 1987: Evolution & Escalation. An ecological history of life, 527. Princeton, New Jersey: Princeton University Press
- Vermeij, G.J., 2013: On escalation. Annual Review of Earth and Planetary Sciences 41, 1–19
- Vermeij, G.J., Schindel, D.E., & Zipser, E., 1981: Predation through geological time: evidence from gastropod shell repair. Science 214, 1024–1026. doi:10.1126/science.214.4524.1024
- Weir, J., 1931: The British and Belgian Carboniferous Bellerophontidae. Transactions of the Royal Society of Edinburgh 56, 767–861
- Wells, H.W., 1958: Predation of pelecypods and gastropods by Fasciolaria hunteria (Perry). Bulletin of Marine Science of the Gulf and Carribbean 8, 152–166
- Yochelson, E.L., 1960: Permian Gastropoda of the southwestern United States. 3. Bellerophontacea and Patellacea. Bulletin of the American Museum of Natural History 119, 205–294
- Zuschin, M., Stachowitsch, M., & Stanton, R.J., Jr, 2003: Patterns and processes of shell fragmentation in modern and ancient marine environments. Earth-Science Reviews 63, 33–82. doi:10.1016/S0012-8252(03)00014-X